

ABSTRACT
Chromosome instability (CIN) contributes to the development of many cancer. In this paper, we summarize our recent finding that a novel pathway by which FBW7 loss promotes Centromere Protein A (CENP-A) phosphorylation on Serine 18 through Cyclin E1/CDK2, therefore promoting CIN and tumorigenesis. Our finding demonstrates the importance of CENP-A post-translational modification on modulating centromere and mitotic functions in cancer.
KEYWORDS: chromosome instability, centromere, CENP-A, Cyclin E1, FBW7, tumorigenesis
Chromosome instability (CIN) contributes to tumor heterogeneity, drug resistance and cancer progression, and high levels of CIN are associated with poor patient survival for many cancer types.1,2 Major mechanisms proposed for CIN include oncogene-induced replication stress, telomere dysfunction, and aberrant mitosis.1 FBW7 is a tumor suppressor protein frequently mutated in multiple cancer types, and belongs to the F-box protein family as part of the SCF ubiquitin E3 ligase complex.3 Cyclin E1 is a well-characterized FBW7 substrate that regulates G1/S cell cycle entry. Aberrant accumulation of Cyclin E1 due to overexpression or FBW7 mutation leads to polyploidy4, and is prevalent in many types of cancers. Existing evidence strongly suggests that Cyclin E1 misregulation results in genomic instability primarily through excessive replication origin firing and oncogene-induced replication stress.5 However, we have recently identified a novel pathway where FBW7 modulates phosphorylation of an essential centromere protein CENtromere Protein A (CENP-A) through Cyclin E1/CDK2, presenting a new paradigm for how this tumor suppressor regulates CIN and tumorigenesis through centromere and mitotic functions.6
The centromere is the specialized chromatin locus that recruits the kinetochore, and is crucial for proper mitosis and genome maintenance. The centromere is enriched for CENP-A, an important histone H3 variant that is considered a key epigenetic mark for centromere identity and propagation.7,8 Too little or too much of CENP-A can disrupt genome integrity. CENP-A depletion displaces the downstream components of the Constitutive Centromere-Associated Network (CCAN) and the KMN network (KNL1 complex, MIS12 complex, NDC80 complex) from centromeres and kinetochores, resulting in chromosome missegregation.9-12 Moreover, ectopic localization of CENP-A to non-centromeric loci due to overexpression or targeted recruitment leads to fragmented chromosomes in Drosophila melanogaster and human cell lines.13-16 At least in Drosophila this is through formation of neo-centromeres and ectopic kinetochores.13-16 Therefore, faithful chromosome segregation requires tight regulation of CENP-A protein levels, to ensure proper CENP-A nucleosome assembly only at centromeres.17 In tumors, overexpression of centromere and kinetochore genes is prevalent.18,19 Importantly, centromere gene upregulation strongly correlates with CIN and poor prognosis in numerous human cancer types18-23, and predicts enhanced cancer cell and patient sensitivity to genotoxic adjuvant therapies.18 However, it is unclear whether centromere misregulation contributes to malignancy, or is merely a consequence of other changes during tumor progression.
Post-translational modifications of centromere proteins also influence centromere functions. These include phosphorylation of Ser16 and Ser18 residues within the CENP-A N terminal tail24, as well as Ser68 in the histone fold domain.25,26 Ser to Ala mutations at these sites lead to defective CENP-A deposition or mitotic defects, although there is some debate about how essential these modifications are in normal cells.26,27 Nevertheless, CENP-A and other centromere and kinetochore protein genes are rarely mutated in large TCGA patient datasets18, thus the clinical relevance of these modifications and the potential roles of CENP-A regulation in cancer progression remain poorly understood.
We found that increased Cyclin E1 levels promote CIN and tumor growth through centromere misregulation.6 Specifically, loss of the tumor suppressor FBW7 results in increased Cyclin E1/CDK2 activity, leading to hyper-phosphorylation of CENP-A at the N- terminal Ser18 site, and reduced CENP-A, CENP-B and HEC1 levels at centromeres. Mechanistically, our study demonstrated that Cyclin E1/CDK2 is necessary and sufficient for CENP-A Ser18 phosphorylation both in vitro and in cultured cells. Further, excessive CENP-A Ser18 phosphorylation enhances CIN, including chromosome missegregation and micronucleus formation, and promotes anchorage-independent growth and tumor progression (Fig. 1). Strong evidence for the relevance of their findings to human clinical cancers was demonstrated using a disease relevant FBW7 mutation, human clinical cancer tissues and a xenograft mouse model. Moreover, our results suggest that Ser18 hyper-phosphorylation due to increased Cyclin E1 activity and/or FBW7 loss reduces efficient CENP-A deposition at centromeres. Mechanistically, these results suggest that the HJURP chaperone and assembly factor does not interact with CENP-A S18D (Ser to Asp) mutant as efficiently as with WT proteins. Together with the data from clinical human breast cancer samples, we identify an important new function for aberrant Cyclin E1/CDK2 activation in cancer, distinguishable from its well-established role in the G1/S transition.6 Previous research on Cyclin E1 in oncogenesis primarily focused on its role in replication initiation, which supports oncogene-induced replication stress.5 However, the results from us suggest that an additional mechanism acting through centromere misregulation also occurs in a significant proportion of human cancers where FBW7 is lost or Cyclin E1 is overexpressed.6
Figure 1.
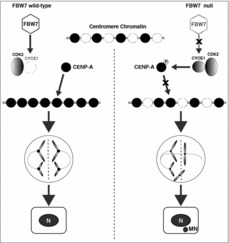
A schematic model of FBW7 defects leading to Cyclin E1 overexpression, CENP-A Ser18 hyper-phosphorylation and chromosomal instability. In FBW7 wild-type cells, CENP-A is successfully deposited at centromeres in late mitosis and early G1 cell cycle phases in the absence of ectopic Cyclin E1/CDK2 activity. In contrast, FBW7 null cells accumulate Cyclin E1, and Cyclin E1 itself is frequently amplified in many cancers. Excessive Cyclin E1/CDK2 activity promotes aberrant CENP-A phosphorylation at the Ser18 residue, reduced CENP-A deposition at the centromere, lagging chromosomes and bridges in mitosis, and micronuclei (MN) formation associated with tumor progression.
The exact mechanism responsible for reduced interaction between HJURP and CENP-A upon CENP-A phosphorylation on Ser18 is currently unclear. Previous structural and molecular studies indicated direct interactions between the HJURP N-terminal region and the CENP-A histone fold domain.28,29 In our study, the failure of a phospho-mimetic CENP-A to be efficiently recruited to LacO arrays by LacI-HJURP, and decreased in vitro binding of HJURP in CENP-A S18D mutants, imply a reduced ability to form a competitive pre-nucleosomal complex or less competitive chromatin incorporation.6 Perhaps CENP-A Ser18 phosphorylation perturbs the stable interaction between HJURP N terminal Scm3 domain and CENP-A histone fold domain. Additionally, it is possible that centromeric CENP-A phosphorylated at Ser18 reduces efficient HJURP recruitment for new CENP-A loading at endogenous centromeres.
Regardless, these results suggest that the CENP-A N terminal tail, and specifically levels of Ser18 phosphorylation, modulate proper centromeric CENP-A nucleosome assembly and centromere function. Previous domain replacement experiments using histone H3 N terminal tail suggested that CENP-A N terminal tail is not absolutely essential for cell viability.30 However, the possibility that phosphorylation modulates centromere function in a cancer context could not be ruled out in those experiments. The results shown in our research are consistent with many findings in the field showing that the CENP-A N terminal tail regulates centromere functions in multiple species31,32, including proper CENP-B function in human cells30,33, epigenetic stability of centromeres in fission yeast34, CENP-A protein stability in budding yeast and Drosophila31,32, and meiosis and organismal fertility in Arabidopsis.35 Interestingly, ectopic overexpression of a CENP-A S18A mutation led to modest but statistically insignificant chromosome missegregation in HeLa cells where most CENP-A is phosphorylated24; while in our publication, CENP-A S18D or forced hyper-phosphorylation in DLD1 cells induces micronuclei.6 The mechanistic details of this pathway await further investigation to better understand its role in carcinogenesis, and for future applications to therapeutic intervention.
Funding Statement
The authors also acknowledge grant support from the National Institutes of Health (NIH) R01 GM119011 (to G.K), NIH (K99/R00 CA160351), Department of Defense Peer Reviewed Cancer Research Program Career Development Award (W81XWH-15-1-0599), The V Foundation Scholar Award, Kimmel Scholar Award, Mary Kay Foundation Award and Susan G. Komen Career Catalyst Award (to Q.Z.).
Acknowledgements
This research was partially supported by University Cancer Research Fund (UCRF) to QZ. The authors also acknowledge grant support from the National Institutes of Health (NIH) R01 GM119011 (to G.K), NIH (K99/R00 CA160351), Department of Defense Peer Reviewed Cancer Research Program Career Development Award (W81XWH-15-1-0599), The V Foundation Scholar Award, Kimmel Scholar Award, Mary Kay Foundation Award and Susan G. Komen Career Catalyst Award (to Q.Z.).
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed
References
- [1].McGranahan N, Burrell RA, Endesfelder D, Novelli MR, Swanton C. Cancer chromosomal instability: therapeutic and diagnostic challenges. EMBO Rep. 2012;13:528-38. doi: 10.1038/embor.2012.61. PMID:22595889 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Holland AJ, Cleveland DW. Losing balance: the origin and impact of aneuploidy in cancer. EMBO Rep. 2012;13:501-14. doi: 10.1038/embor.2012.55. PMID:22565320 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Wang Z, Inuzuka H, Zhong J, Wan L, Fukushima H, Sarkar FH, Wei W. Tumor suppressor functions of FBW7 in cancer development and progression. FEBS Lett. 2012;586:1409-18. doi: 10.1016/j.febslet.2012.03.017. PMID:22673505 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Spruck CH, Won KA, Reed SI. Deregulated cyclin E induces chromosome instability. Nature. 1999;401:297-300. doi: 10.1038/45836. PMID:10499591 [DOI] [PubMed] [Google Scholar]
- [5].Jones RM, Mortusewicz O, Afzal I, Lorvellec M, Garcia P, Helleday T, Petermann E. Increased replication initiation and conflicts with transcription underlie Cyclin E-induced replication stress. Oncogene. 2013;32:3744-53. doi: 10.1038/onc.2012.387. PMID:22945645 [DOI] [PubMed] [Google Scholar]
- [6].Takada M, Zhang W, Suzuki A, Kuroda TS, Yu Z, Inuzuka H, Gao D, Wan L, Zhuang M, Hu L, et al.. FBW7 Loss Promotes Chromosomal Instability and Tumorigenesis via Cyclin E1/CDK2-Mediated Phosphorylation of CENP-A. Cancer Res. 2017;77:4881-93. doi: 10.1158/0008-5472.CAN-17-1240. PMID:28760857 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Westhorpe FG, Straight AF. Functions of the centromere and kinetochore in chromosome segregation. Curr Opin Cell Biol. 2013;25:334-40. doi: 10.1016/j.ceb.2013.02.001. PMID:23490282 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Allshire RC, Karpen GH. Epigenetic regulation of centromeric chromatin: old dogs, new tricks? Nat Rev Genet. 2008;9:923-37. doi: 10.1038/nrg2466. PMID:19002142 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Howman EV, Fowler KJ, Newson AJ, Redward S, MacDonald AC, Kalitsis P, Choo KH. Early disruption of centromeric chromatin organization in centromere protein A (Cenpa) null mice. Proc Natl Acad Sci U S A. 2000;97:1148-53. doi: 10.1073/pnas.97.3.1148. PMID:10655499 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Blower MD, Karpen GH. The role of Drosophila CID in kinetochore formation, cell-cycle progression and heterochromatin interactions. Nat Cell Biol. 2001;3:730-9. doi: 10.1038/35087045. PMID:11483958 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Oegema K, Desai A, Rybina S, Kirkham M, Hyman AA. Functional analysis of kinetochore assembly in Caenorhabditis elegans. J Cell Biol. 2001;153:1209-26. doi: 10.1083/jcb.153.6.1209. PMID:11402065 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Goshima G, Kiyomitsu T, Yoda K, Yanagida M. Human centromere chromatin protein hMis12, essential for equal segregation, is independent of CENP-A loading pathway. J Cell Biol. 2003;160:25-39. doi: 10.1083/jcb.200210005. PMID:12515822 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Heun P, Erhardt S, Blower MD, Weiss S, Skora AD, Karpen GH. Mislocalization of the Drosophila centromere-specific histone CID promotes formation of functional ectopic kinetochores. Dev cell. 2006;10:303-15. doi: 10.1016/j.devcel.2006.01.014. PMID:16516834 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Barnhart MC, Kuich PH, Stellfox ME, Ward JA, Bassett EA, Black BE, Foltz DR. HJURP is a CENP-A chromatin assembly factor sufficient to form a functional de novo kinetochore. The Journal of cell biology. 2011;194:229-43. doi: 10.1083/jcb.201012017. PMID:21768289 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Mendiburo MJ, Padeken J, Fulop S, Schepers A, Heun P. Drosophila CENH3 is sufficient for centromere formation. Science. 2011;334:686-90. doi: 10.1126/science.1206880. PMID:22053052 [DOI] [PubMed] [Google Scholar]
- [16].Mishra PK, Au WC, Choy JS, Kuich PH, Baker RE, Foltz DR, Basrai MA. Misregulation of Scm3p/HJURP causes chromosome instability in Saccharomyces cerevisiae and human cells. PLoS Genet. 2011;7:e1002303. doi: 10.1371/journal.pgen.1002303. PMID:21980305 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Black BE, Cleveland DW. Epigenetic centromere propagation and the nature of CENP-a nucleosomes. Cell. 2011;144:471-9. doi: 10.1016/j.cell.2011.02.002. PMID:21335232 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Zhang W, Mao JH, Zhu W, Jain AK, Liu K, Brown JB, Karpen GH. Centromere and kinetochore gene misexpression predicts cancer patient survival and response to radiotherapy and chemotherapy. Nat Commun. 2016;7:12619. doi: 10.1038/ncomms12619. PMID:27577169 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Tomonaga T, Matsushita K, Yamaguchi S, Oohashi T, Shimada H, Ochiai T, Yoda K, Nomura F. Overexpression and mistargeting of centromere protein-A in human primary colorectal cancer. Cancer Res. 2003;63:3511-6. PMID:12839935 [PubMed] [Google Scholar]
- [20].Athwal RK, Walkiewicz MP, Baek S, Fu S, Bui M, Camps J, Ried T, Sung MH, Dalal Y. CENP-A nucleosomes localize to transcription factor hotspots and subtelomeric sites in human cancer cells. Epigenetics Chromatin. 2015;8:2. doi: 10.1186/1756-8935-8-2. PMID:25788983 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Lacoste N, Woolfe A, Tachiwana H, Garea AV, Barth T, Cantaloube S, Kurumizaka H, Imhof A, Almouzni G. Mislocalization of the centromeric histone variant CenH3/CENP-A in human cells depends on the chaperone DAXX. Mol Cell. 2014;53:631-44. doi: 10.1016/j.molcel.2014.01.018. PMID:24530302 [DOI] [PubMed] [Google Scholar]
- [22].Montes de Oca R, Gurard-Levin ZA, Berger F, Rehman H, Martel E, Corpet A, de Koning L, Vassias I, Wilson LO, Meseure D, et al.. The histone chaperone HJURP is a new independent prognostic marker for luminal A breast carcinoma. Mol Oncol. 2015;9:657-74. doi: 10.1016/j.molonc.2014.11.002. PMID:25497280 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Hu Z, Huang G, Sadanandam A, Gu S, Lenburg ME, Pai M, Bayani N, Blakely EA, Gray JW, Mao JH. The expression level of HJURP has an independent prognostic impact and predicts the sensitivity to radiotherapy in breast cancer. Breast Cancer Res: BCR. 2010;12:R18. doi: 10.1186/bcr2487. PMID:20211017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Bailey AO, Panchenko T, Sathyan KM, Petkowski JJ, Pai PJ, Bai DL, Russell DH, Macara IG, Shabanowitz J, Hunt DF, et al.. Posttranslational modification of CENP-A influences the conformation of centromeric chromatin. Proc Natl Acad Sci U S A. 2013;110:11827-32. doi: 10.1073/pnas.1300325110. PMID:23818633 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Sekulic N, Bassett EA, Rogers DJ, Black BE. The structure of (CENP-A-H4)(2) reveals physical features that mark centromeres. Nature. 2010;467:347-51. doi: 10.1038/nature09323. PMID:20739937 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Yu Z, Zhou X, Wang W, Deng W, Fang J, Hu H, Wang Z, Li S, Cui L, Shen J, et al.. Dynamic phosphorylation of CENP-A at Ser68 orchestrates its cell-cycle-dependent deposition at centromeres. Dev Cell. 2015;32:68-81. doi: 10.1016/j.devcel.2014.11.030. PMID:25556658 [DOI] [PubMed] [Google Scholar]
- [27].Fachinetti D, Logsdon GA, Abdullah A, Selzer EB, Cleveland DW, Black BE. CENP-A Modifications on Ser68 and Lys124 Are Dispensable for Establishment, Maintenance, and Long-Term Function of Human Centromeres. Dev Cell. 2017;40:104-13. doi: 10.1016/j.devcel.2016.12.014. PMID:28073008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Shuaib M, Ouararhni K, Dimitrov S, Hamiche A. HJURP binds CENP-A via a highly conserved N-terminal domain and mediates its deposition at centromeres. Proc Natl Acad Sci U S A. 2010;107:1349-54. doi: 10.1073/pnas.0913709107. PMID:20080577 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Hu H, Liu Y, Wang M, Fang J, Huang H, Yang N, Li Y, Wang J, Yao X, Shi Y, et al.. Structure of a CENP-A-histone H4 heterodimer in complex with chaperone HJURP. Genes Dev. 2011;25:901-6. doi: 10.1101/gad.2045111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Fachinetti D, Folco HD, Nechemia-Arbely Y, Valente LP, Nguyen K, Wong AJ, Zhu Q, Holland AJ, Desai A, Jansen LE, et al.. A two-step mechanism for epigenetic specification of centromere identity and function. Nat Cell Biol. 2013;15:1056-66. doi: 10.1038/ncb2805. PMID:23873148 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Au WC, Dawson AR, Rawson DW, Taylor SB, Baker RE, Basrai MA. A novel role of the N terminus of budding yeast histone H3 variant Cse4 in ubiquitin-mediated proteolysis. Genetics. 2013;194:513-8. doi: 10.1534/genetics.113.149898. PMID:23525333 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Malik HS, Vermaak D, Henikoff S. Recurrent evolution of DNA-binding motifs in the Drosophila centromeric histone. Proc Natl Acad Sci U S A. 2002;99:1449-54. doi: 10.1073/pnas.032664299. PMID:11805302 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Fachinetti D, Han JS, McMahon MA, Ly P, Abdullah A, Wong AJ, Cleveland DW. DNA Sequence-Specific Binding of CENP-B Enhances the Fidelity of Human Centromere Function. Developmental cell. 2015;33:314-27. doi: 10.1016/j.devcel.2015.03.020. PMID:25942623 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Folco HD, Campbell CS, May KM, Espinoza CA, Oegema K, Hardwick KG, Grewal SI, Desai A. The CENP-A N-tail confers epigenetic stability to centromeres via the CENP-T branch of the CCAN in fission yeast. Curr Biol. 2015;25:348-56. doi: 10.1016/j.cub.2014.11.060. PMID:25619765 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Ravi M, Shibata F, Ramahi JS, Nagaki K, Chen C, Murata M, Chan SW. Meiosis-specific loading of the centromere-specific histone CENH3 in Arabidopsis thaliana. PLoS Genet. 2011;7:e1002121. doi: 10.1371/journal.pgen.1002121. PMID:21695238 [DOI] [PMC free article] [PubMed] [Google Scholar]