

Abstract
Decision-makers in wildlife policy require reliable population size estimates to justify interventions, to build acceptance and support in their decisions and, ultimately, to build trust in managing authorities. Traditional capture-recapture approaches present two main shortcomings, namely, the uncertainty in defining the effective sampling area, and the spatially-induced heterogeneity in encounter probabilities. These limitations are overcome using spatially explicit capture-recapture approaches (SCR). Using wolves as case study, and non-invasive DNA monitoring (faeces), we implemented a SCR with a Poisson observation model in a single survey to estimate wolf density and population size, and identify the locations of individual activity centres, in NW Iberia over 4,378 km2. During the breeding period, posterior mean wolf density was 2.55 wolves/100 km2 (95%BCI = 1.87–3.51), and the posterior mean population size was 111.6 ± 18.8 wolves (95%BCI = 81.8–153.6). From simulation studies, addressing different scenarios of non-independence and spatial aggregation of individuals, we only found a slight underestimation in population size estimates, supporting the reliability of SCR for social species. The strategy used here (DNA monitoring combined with SCR) may be a cost-effective way to generate reliable population estimates for large carnivores at regional scales, especially for endangered species or populations under game management.
Introduction
Estimating the abundance of species is one of the most contentious issues in conservation and applied ecology1,2. Decision-makers in wildlife policy require reliable population size and density estimates to adopt and justify interventions. Reliability is essential to build acceptance and support in management decisions and, ultimately, trust in managing authorities. Otherwise, speculation and distrust can emerge after decisions are made, and may undermine entire management or conservation strategies1,3. Incorrect population estimates may lead to misinterpretations of the status of populations, the impact of interventions (e.g., hunting quotas or culling programs), or the degree to which conservation goals have been achieved.
The management of large carnivores is controversial due to the multiple political, socio-economic and conservation interests involved. Information on population size or the impact of interventions is in constant demand, not only by managers, researchers and conservationists, but also by other interest groups. This is exemplified by recurrent debates around large carnivore numbers, particularly centred on endangered and charismatic species, such as in the case of tigers (Panthera tigris), lions (Panthera leo) or wolves (Canis lupus)4–8. Clear population targets are often established by managing authorities, and have become political issues, with reliable assessments of changes in large carnivore ranges and population size required to justify actions9.
Wolves are a good example of a species whose estimates of population size and range are systematically demanded by multiple actors (e.g., governments, livestock producers, conservationists), and whose population targets are often established, compared to other wildlife. Wolf estimates are required to meet legal obligations in both Europe10 and the US11. The Spanish authorities approved a short-term recovery goal of 15 packs for the endangered Sierra Morena wolf population12. The Swedish parliament decided to maintain its wolf population within a minimum of 20 reproductions and a maximum of 210 individuals13. The Norwegian parliament has established a national target of four to six wolf litters a year14. The recovery goal for wolves in the Northern Rocky Mountains after reintroduction into Yellowstone National Park and central Idaho, US, was set to >300 wolves and >30 breeding pairs evenly distributed among the recovery areas for three consecutive years15.
The number of wolf packs or reproductions are often the basis of wolf monitoring16–18, and may make wolf population size estimates more homogeneous in a transboundary and regional context. Notwithstanding, many wolf management strategies still combine the number of packs/reproductions with the number of individuals. In Europe, for example, estimates reporting the number of individuals still prevail19, requiring a conversion exercise from the number of packs/reproductions (usually using estimates on pack size). However, this conversion remains a challenge20 and, in most cases, the data required to calculate conversion factors properly is not available. While the number of packs or reproductions are a reasonable target for wolf monitoring at regional scales17, apart from mismatches between management goals (i.e., often based on the number of individuals, for instance, to establish hunting quotas) and monitoring targets (i.e., packs or reproductions), there are cases where estimating the number of wolves may be important. Examples include small and endangered wolf populations, such as the Sierra Morena12 or Mexican21 wolf populations, and populations under game management6.
Different field methods and analytical approaches have been used to address the challenge of surveying large carnivore populations at regional scales17,22–26, including non-invasive DNA monitoring21,23,27. This method, combined with traditional capture-recapture procedures, is often presented as a promising strategy to achieve robust, feasible and economically affordable population size estimates. Nevertheless, several constraints have been identified when implementing this monitoring strategy. A recurrent limitation is the imprecision when it comes to defining the effective sampling area28. When using traditional capture-recapture procedures, additional spatial information is required in order to define the effective sampling area, such as information from collared individuals29,30. However, gathering this information may not be budgetary feasible in many cases, particular at regional scales. Moreover, the minimum number and type of collared individuals needed to capture the movement parameters in a population must be representative. On the other hand, encountering probabilities are heterogeneous among those individuals exposed to sampling31.
However, the development of spatially explicit capture-recapture approaches (SCR)–linking population size with space by estimating a latent variable representing the location and number of individual’s activity centres–allows the estimation of density, defined as the local intensity of a spatial point process32, taking into account heterogeneity in encounter probabilities. In SCR approaches, the effective sampling area is estimated based on the underlying spatial point process, using information from individual’s encounters across detectors31. Although it is possible to integrate other sources of spatial information, such as telemetry data33. For large carnivores, within the SCR framework, several field sampling methods have been used to generate spatial encounter data, mainly camera-trapping data34–36 and non-invasive DNA monitoring37–39. Occasions (i.e., repeated opportunities for observation) can be accomplished either in space or time (e.g., one site sampled multiple times or multiple sites sampled once). Under this framework, SCR approaches based on a single survey and using a Poisson observation model allows density estimates40,41.
SCR assumes that distributions of animal activity centres are uniformly and independently distributed over the state space (S) –the area that includes all individuals potentially exposed to sampling32. However, both assumptions are violated by multiple species. For example, in social and territorial carnivores, like wolves or lions, a large segment of the population lives in packs or prides, respectively. In solitary carnivores, like many felid species or bears42, adult females and juveniles (i.e., family groups) stay together for several months within the annual cycle before dispersion. In wolves, the functional unit is the pack, but wolf populations show a varying proportion of non-resident individuals43. Although the size of wolf home ranges is influenced by individual attributes (e.g., age, sex, reproductive status)43,44, pack cohesion varies throughout the annual cycle45,46 and pack members may use space differently within pack territories47, spatial overlap of home ranges for pack members being high44,47.
Here, we combined non-invasive DNA monitoring (faeces) with a SCR Poisson observation model in a single survey to demonstrate the performance of this monitoring strategy in estimating wolf density and population size, and identify the activity centres of wolves at regional scales. Using wolves as illustrative example, we used simulations to show how SCR models perform with species violating the assumptions that distributions of animal activity centres and animal movements are independent32. In addition, to provide additional support to this monitoring strategy, we compared estimates of the spatial scale parameter sigma (σ) -related with the movement of individuals- obtained from the SCR model, with empirical data from a set of collared wolves in the study area.
Materials and Methods
Sample collection and genetic analyses
We tested the use of non-invasive DNA monitoring (faeces) with a SCR Poisson observation model in a single survey in the wolf population of Costa da Morte and surroundings (hereafter CM; Galicia, NW Spain) (Figs 1 and S4), a segment of the NW Iberian wolf population18 covering ca. 4,378 km2, and where 11 breeding packs were detected in 201348. During the 2013 wolf breeding period (May-October, including the pup-rearing period), 317 wolf-like faeces were collected across CM. In collaboration with rangers from the Regional Government of Galicia, faeces were searched along transects distant from human settlements26. Random sampling is not effective to locate wolf faeces; therefore, scat sampling was focused on landscape elements often used by wolves as marking places (paths and trails, particularly focusing on junctions and mountain passes)26. A minimum of 10 km of transect length was invested per each 10 × 10 km UTM cell, surveying a total of ca. 750 km over the study area (Figs S1 and S2). Surveying a minimum of 10 km per cell resulted in a probability of detecting wolves of >0.617, which is likely influenced by the persistence of wolf faeces. All samples were georeferenced using a GPS and preserved in 96% ethanol at room temperature.
Figure 1.
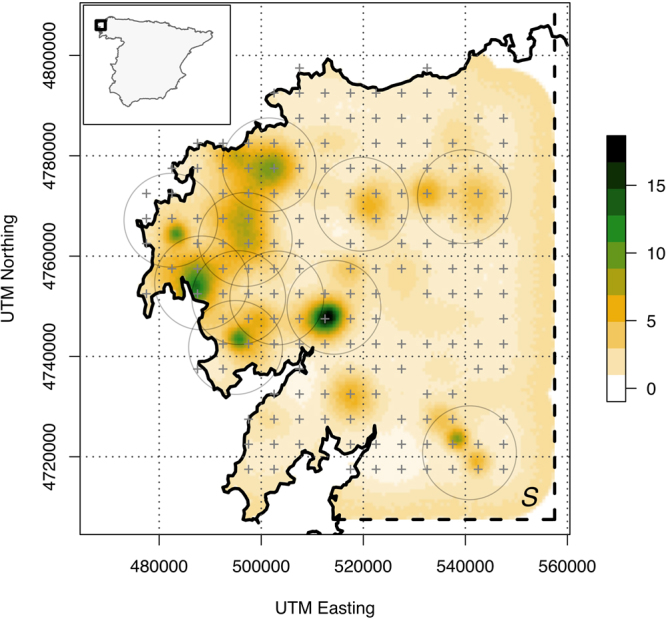
Posterior mean density of activity centres for wolves in the study area. The spatial locations of detectors are denoted by “+” (grey). S: state space. We included the approximate location of reproductive pack territories (grey circles) by creating a conservative buffer area centred on the rendezvous sites of known breeding packs in 201348. The selection of the size of the circles was based on previous information on the mean and SD of wolf home range sizes of subadult/adult wolves belonging to packs in NW Iberia (122.1 ± 93.6 km2)52 (the area was equal to the mean plus SD = 215 km2). The figure was produced by José Jiménez using R55.
Details on genotyping are provided in Pacheco et al.49 and Appendix S1. MtDNA species identification was achieved for 288 samples (91%). Based on DNA quality, 172 samples were further considered for individual identification using four replicas of 18 Ancestral Informative Markers50. After a Bayesian analysis to exclude dogs (see details in Pacheco et al.49), ninety-five wolf samples achieved a consensus genotype (55%) with <20% missing data, following rules defined in Godinho et al.50. Multiple factors can influence the success rate of genotyping and individual identification, such as the age of the wolf scat (related to the presence of odour)51. Nevertheless, the success rate observed in this study was similar to other values reported in studies using non-invasive DNA51. Estimated allelic dropout across loci was ADO = 0.033, and estimated false alleles were FA = 0.00549. After regrouping of identical genotypes49, we identified 65 individual wolves. This dataset had a probability of identity (PID) = 4.19 × 10−8 and a PIDsiblings = 4.01 × 10−4 (Appendix S1).
Wolf estimates
We applied a SCR modelling approach to estimate wolf density, population size and the distribution of individual’s activity centres in CM. Conceptually, activity centres are points si representing “where the animal i of the N population size lives”. Formally, the sequence of points s1, …, sN is a realization of a latent spatial point process, and allows the estimation of density using SCR models. SCR assumes that every individual i in the population has its own activity centre si, and that all activity centres are distributed across the study area. Moreover, encounter probability (pij) is a decreasing function of distance between the activity centre of the individual i (si) and the location of the detector j (xj). Therefore, SCR addresses the movement of individuals by assuming that each individual has an activity centre, and that the probability of capture individuals is a function of the distance from the activity centres to detector locations. In our case, because the collection of wolf faeces was not attached to physical traps, our detectors were the centroids of the cells within the sample grid, and the effort invested in searching samples inside cells was used as a “detector level” covariate.
We built a grid cell layer with the size of cells being small compared to typical wolf home ranges. Considering previous information on the spatial ecology of wolves in NW Iberia, we created a 5 × 5 km (25 km2) cell grid layer over the study area. Cell size was well below the average (±SD) home range size for subadult/adult wolves belonging to packs in NW Iberia (122.1 ± 93.6 km2)52. This cell size was selected to avoid an excessive loss of resolution in the scale parameter sigma (σ) –a parameter that determines the decline of detection frequency of individuals in detectors with increasing distance from their activity centres–, resulting in 174 5 × 5 km cells in CM (Fig. S2). Each cell centroid was considered as a detector location (Fig. S1). Thus, we modelled centroids (detector locations) as count detectors because the same wolf can be detected at multiple cells during the sampling, and more than one individual can be detected in the same detector32. All genotyped faeces were assigned to its detector. The encounter histories are a bi-dimensional matrix y (i × j), because we used a single occasion.
Therefore, the number of times that an individual i is located in a detector j is Poisson distributed with mean λij:
| 1 |
The link function between the location of detectors and the activity centres follows a half-normal distribution32:
| 2 |
where dij is the distance between the activity centre for each individual si and the detector location xj, and λ0 is the baseline encounter probability. The baseline encounter probability is modelled with a log function being dependent on sampling effort within each cell, and σ is the Gaussian scale parameter for the distance function between activity centres and detector locations (Fig. S2):
| 3 |
As several cells share land and sea surfaces in CM (Fig. 1), we used the density of transects (km km−2) in each terrestrial section of the cells as a covariate accounting for sampling effort . Density of transects was scaled and centred, and α0 and α1 were the parameters to be estimated. The total number of activity centres (N) were calculated using the data augmentation approach32. All-zero encounter histories (500 potential individuals) were added to encounter histories of detected individuals. We generated the state space (S) adding a 10-km buffer to the detector grid. This distance must be >2.5 × σ32. The resulting S was ca. 7,500 km2. We used an application of the “zero’s trick” to buffer an irregular detector array (i.e., near the coastline) without discretizing the state space53. We scaled and centred S dividing the spatial coordinates (m) by 10,00032.
We fitted a null Poisson distributed observation model (M0) in a Bayesian framework using Nimble v.0.6-254 and R55 (Appendix S2). We ran 3 chains of the MCMC sampler with 50,000 iterations each, yielding 150,000 total samples from the joint posterior distributions. To check for chain convergence, we assessed MCMC convergence and mixing by visually inspecting trace plots for each parameter of interest, and we calculated the Gelman-Rubin statistic R-hat56 using the R package coda57, where values below 1.1 indicated convergence. We evaluated the goodness of fit of the model by using the Bayesian p-value approach and two fit statistics (described in Royle et al.)32: i) individual encounter frequencies, which evaluates heterogeneity in encounter frequencies due to space (i.e., in SCR models, the explicit factor space explains part of the heterogeneity); and ii) detector frequencies, which is based on aggregating over individuals and replicates to form centroid-encounter frequencies (Appendix S2).
SCR approaches assume that all individuals are uniformly and independently distributed over the state space S. We explored the influence of such violations on population density estimates by carrying out three simulation studies (described in Appendix S3). We tested for the influence of the randomness in the location of wolf packs across the state space, because the identification of the spatial location of activity centres is more accurate inside the detector grid.
Spatial information on wolves
We evaluated the performance of this modelling approach by comparing σ from M0 with two sigma parameter estimates based on wolf spatial behaviour (. We used spatial information from 18 wolves collared in the study area between 2006 and 2014. Wolves were captured with Belisle© leg-hold snares (Edouard Belisle, Canada), chemically immobilised by intramuscular injection of medetomidine (Domitor®, Merial, France; 0.10 mg/kg) and equipped with GPS-GSM collars (Followit, Sweden). Our dataset was comprised of 9 males and 9 females, of ≥2 years, and considering individuals integrated and non-integrated within packs. We used a total dataset of 5,432 locations (2 locations per day and wolf; mean number of locations per Wolf = 302, range 34–789). The average monitoring period for collared wolves was 161 days (range 14–397). In order to provide additional support to this modelling approach, we also compared sigma estimates during the same period of the year (, to account for variations in wolf spatial behaviour along the annual cycle43. To do this, we subset our dataset on wolf spatial information considering only those locations between March and October, corresponding with the period of DNA monitoring plus two previous months given by the potential persistence of wolf faeces in the field. This dataset was composed by 3,650 locations from 16 wolves (range 26–549 locations per individual). We estimated individual home ranges (km2) using the 95% fixed-kernel method with the R package adehabitatHR58.
We calculated the scale parameters (considering all wolf locations) and (only considering those wolf locations overlapping with the DNA monitoring period) considering the average spatial requirements of wolves in CM, as follows:
| 4 |
where was the value of a Chi-square with 2 degrees of freedom (α = 0.05, ) and A was the average home range area (m2)32.
We also evaluated the spatial mismatch between the spatially explicit posterior mean density of activity centres for wolves across S and the location of known breeding packs in CM. We used the information on the number and spatial location of wolf breeding packs from the official wolf monitoring survey carried out in CM in 201348 (wolf monitoring was based on the location of breeding packs following Llaneza, García & López-Bao 2014)26.
Finally, we estimated the proportion of wolves that were part of known breeding packs. To do this, we assumed that in late summer, breeding wolf packs oscillated between 7 and 9 individuals, including the year’s pups. Although pack size is variable in wolves43, our assumption was based on 88 field observations of breeding packs at rendezvous sites (i.e., sites used by pack members and pups approximately in the first 5 months of pups’ age)59 in N Iberia, where the average number of observed pups was 4.82 (95%CI 4.57–5.08), the average number of subadults was 1.67 (95%CI 1.25–2.08) and the average number of adults was 1.98 (95%CI 1.74–2.22)60. Considering the number of known breeding packs in CM in 2013 (11 packs)48 and the range of individuals per pack in late summer (7–9 wolves), we transformed the number of packs into number of wolves belonging to packs.
Ethics statement
Wolves were captured under permits 19/2006, 71/2009, 86/2011, 095/2013 and 8765/2015 from the Regional Government of Galicia. All field procedures were carried out in accordance with animal welfare regulations.
Results
From DNA analyses, forty-nine wolves (75%) were detected once, nine wolves were detected twice and, respectively, three, one and three individuals were detected 3, 4 and 5 times (the average number of captures by wolf was 1.46). The number of individuals captured per detector is shown in Table S1. Based on raw data (65 individual genotypes and 95 captures), asymptotic rarefaction curves were not reached (Fig. S3). All parameters from the null model (M0) showed convergence and R-hat < 1.1. The posterior density estimate for wolves in CM was 2.55 wolves/100 km2 (95% Bayesian Credible Interval (BCI) = 1.87–3.51; Table 1; Fig. S4). The posterior mean abundance of wolves, (±SD) was 111.6 ± 18.8 individuals (95%BCI = 81.8–153.6). The posterior estimate for sigma was 0.33 (m, scaled by 104) (95%BCI = 0.27–0.40; Table 1). Bayesian p-values showed a good fit for the case of individual encounter frequencies (p-value = 0.457), but not for detector-encounter frequencies (p-value = 0.000) (Fig. S5).
Table 1.
Posterior summaries of parameters estimated from the SCR Poisson encounter model (null model, M0) to estimate wolf density.
| Mean | SD | BCI | |||
|---|---|---|---|---|---|
| 2.5% | 50% | 97.5% | |||
| Density () | 2.55 | 0.42 | 1.87 | 2.50 | 3.51 |
| alpha1 () | −1.21 | 0.25 | −1.38 | −1.21 | −0.74 |
| alpha2 () | 0.24 | 0.13 | −0.01 | 0.24 | 0.50 |
| psi (ψ) | 0.38 | 0.07 | 0.27 | 0.38 | 0.53 |
| sigma () | 0.33 | 0.03 | 0.27 | 0.32 | 0.40 |
Estimates were based on 3 chains of 50,000 iterations, and thin rate = 1, yielding 150,000 total samples from the joint posterior. BCI = Bayesian Credible Intervals. alpha1 () and alpha2 () are used to model the baseline encounter rate, being dependent on sampling effort (see main text for details): , where Lj is the density of transect in each cell; psi (ψ) is a parameter of the augmented data, and sigma (σ), or movement parameter, is the half-normal scale parameter that describes the rate at which detection probability declines as a function of distance.
From simulation studies, using clusters of individuals and repulsion between clusters (packs in our case) we tested the potential role of spatial aggregation and non-independence of individuals, and we only found a slight underestimation in population size estimates (Appendix S3). The adequacy of the Root-Mean-Squared-Error for the posterior mean for the parameter , and the coverage of 95% Highest Posterior Density intervals performed better when all clusters of individuals were located within the area covered by the detector grid (Appendix S3). Estimations of slightly decreased in accuracy and precision when clusters of individuals where totally or partially outside the detector grid (Appendix S3).
The average total home range estimate for our set of collared wolves was 266.4 km2; whereas this figure was 233.1 km2 when we considered only locations within the interval March-October. The estimates for and were very similar, 0.37 and 0.35, respectively (m, scaled by 104). These values were within the 95% BCI of the posterior estimate for sigma from M0 (0.27–0.40; Table 1), indicating a good biological performance of the model. The posterior mean density of activity centres across the state space spatially matched all the approximate locations of detected breeding packs in CM in 2013 (Fig. 1). However, we also detected some activity centres that were not overlapping with known breeding packs, indicating potential pack locations. Finally, the estimated number of wolves not linked to packs was 23.6 ± 8.7 individuals (i.e., 16–25% of the total number of wolves estimated using SCR; 88.0 ± 16.7 wolves were estimated to belong to packs).
Discussion
Controversy often surrounds population size and density estimates of large carnivores. Different pitfalls are usually highlighted, such as the lack of standardisation and homogenisation in sampling protocols, lack of replicability or high uncertainties in estimates. For example, the majority of population size estimates for large carnivores in Europe lack rough calculations of uncertainty18,19. There is a demand for development of reliable monitoring tools, including those that allow for the quantification of uncertainty. In this regard, several powerful approaches, paying attention to accuracy, precision and optimization, have been proposed17,34,61,62. Improving the reliability of population estimates is expected to increase the credibility of surveys and improve trust in the agencies and stakeholders in charge of large carnivore monitoring.
We combined non-invasive DNA monitoring (faeces) and a SCR approach, with a Poisson observation model using a single survey, to estimate density, population size and the location of activity centres in wolves at a regional scale. The number of sampling surveys can constrain the application of SCR approaches at regional scales. We used the null model M0 to estimate the number of wolves with a measure of uncertainty (111.6 ± 18.8) across an area of >4,000 km2. A critical point in SCR models is the need for sufficient spatial recaptures for reliable estimates of σ32. Considering the average number of captures by wolf (1.46), the number of individuals captured across detectors (65), and the space among detectors (5 km), our spatial parameters can be considered reliable31,32. Count-based observation models, such as the Poisson model, using multiple detections of the same individual at the same detector, and being based on a single survey, allows the estimation of model parameters successfully. For situations in which behavioural or time-related covariates –across the sampling occasions- are not available, the property of additivity of the Poisson distribution allows the aggregation of the observation data, facilitating the use of the Poisson model based on a single survey to estimate model parameters (the observation model is the same for all values of K)32. Nevertheless, when implementing encounter frequency models, like a Poisson model, encounter frequencies over short periods are mainly due to individual behaviour. Moreover, it can be questionable to consider that all encounters are independent, which deserves further investigation.
The location of wolf activity centres fully matched the approximate locations of detected breeding packs in CM. But we also detected non-overlapping aggregations of activity centres (Fig. 1), probably corresponding to other pack locations where no reproduction was detected in 2013, or where the criteria used to assign wolf reproduction were not satisfied26,48. In fact, the spatial location of some aggregations of activity centres corresponded with the location of previously known breeding packs in the study area50. The combination of both wolf surveys (packs and individuals), together with information on pack size60, allowed us to approximate the number of wolves not linked to breeding packs in late summer, between 16 and 25% of wolves in our case. This estimate is similar to those obtained during winter wolf monitoring in other study areas across the wolf range. The percent of non-resident wolves in winter monitoring in different North American wolf populations ranged from 7 to 20%43. Interestingly, the above comparisons suggest that when using the number of packs as a target for wolf monitoring, underestimation in population estimates may occur.
For species with large spatial requirements, such as large carnivores, and short study periods, the area of activity centres used during the sampling period may represent only a section of the total animals’ home range. In our case, the sampling period spanned several months during the breeding and pup-rearing periods, and the posterior estimate for sigma () was close to the sigma calculation using the total home range from our dataset of collared wolves (), as well as the restricted dataset of locations for the period March-October () (note that both estimates were within the 95%BCI for sigma from M0; Table 1). Thus, using the SCR Poisson approach, we were able to capture some aspects of the spatial behaviour of wolves. Differences in scent marking patterns among individuals26,27 may be behind the small differences observed in sigma, which could lead to some small bias in population estimates. Young and lone wolves more often deposit their faeces off-trail compared to adult and territorial individuals27,63. On the other hand, the persistence of wolf faeces may facilitate the detection of scats and the occurrence of spatial recaptures.
The assumptions that all individuals are uniformly and independently distributed over the state space S32 are violated by multiple species, which raises concerns when using SCR methodologies64, and may also have prevented their use in some cases. In this study, our simulations show that SCR can be reliable in species violating these assumptions. The aggrupation of wolves in packs, and their territorial behaviour, had a minimum impact on population size estimates. We found a slight underestimation in population size estimates (Appendix S3)32,65. Importantly, our results highlight the importance of setting the detector grid appropriately when monitoring species in which individuals are aggregated. Outside the detector grid, the individuals from a cluster (i.e., pack), and randomly located, were detected fewer times and with fewer spatial and non-spatial captures (Appendix S3), resulting in less accuracy and precision for and σ. Therefore, we recommend that the detector grid be extended to cover all previous suspected/known clusters of individuals in a given study area, and the use of simulations to detect possible bias in estimates (Appendix S3).
The combination of non-invasive DNA monitoring (faeces, hair, etc.) and SCR modelling approaches can be used to monitor species at regional scales accurately, such as large carnivores and large herbivores. Reliable population size estimates are particularly important for these species considering that ca. 60% of the world´s largest carnivores and herbivores are classified as threatened with extinction on the International Union for the Conservation of Nature (IUCN) Red List66 and different populations are under game management. The use of a single survey is expected to be less costly compared to multiple visits to detectors64. Moreover, the use of this monitoring strategy is expected to have positive implications for the adaptive management of these species. Adaptive management is a systematic approach to improve resource management by learning from management outcomes67. Because it is a learning-based process68, monitoring is an essential step in the adaptive management process. In our case, the ability to obtain reliable population estimates – quantifying uncertainty - that are reproducible over time is fundamental to assess the impact of interventions properly, which can be achieved with this monitoring strategy.
Apart from funding to carry out genetic analyses, the main challenge in implementing this monitoring strategy is an appropriate sample collection to ensure a sufficient number of recaptures in a single survey. In this regard, an alternative to optimize the use of limited resources, without compromising the collection of samples, may be the integration of multiple trained stakeholders and volunteers in sample collection (see an example for brown bears, Ursus arctos, in Europe)69, which should be properly designed to record all required information, including sampling effort.
Electronic supplementary material
Acknowledgements
We are in debt to the staff of the Regional Government of Galicia for the administrative and logistical support, and all the rangers for their incredible work collecting wolf samples under the framework of the Spanish wolf monitoring program, promoted by the Spanish Ministry of Agriculture, Food and Environment and the Autonomous Regions. J.V.L.B. was supported by a Ramon & Cajal research contract (RYC-2015-18932) from the Spanish Ministry of Economy, Industry and Competitiveness. R.G. was supported by research contract (IF/00564/2012) from the Portuguese Foundation for Science and Technology (FCT). We thank D. Castro and S. Lopes for lab assistance and P. Perú, A. Rivadulla and V. Sazatornil for field assistance. We thank two anonymous referees for their constructive comments on a previous version of this article. This work was partially supported by the project PTDC/BIA-EVF/2460/2014 (FCT). This is scientific paper 20 of the Iberian Wolf Research Team (IWRT).
Author Contributions
J.V.L.B. and J.J. conceived the idea; L.L., F.J.L., E.G., V.P. and J.V.L.B. collected the data; L.L., V.P., E.G., and J.V.L.B. were involved in wolf collaring; R.G. and C.P. run the laboratory analyses; J.J. and J.V.L.B. analysed the data; J.V.L.B. led the writing of the manuscript, with particular support from J.J. All authors commented the final draft and gave final approval for publication.
Competing Interests
The authors declare that they have no competing interests.
Footnotes
Electronic supplementary material
Supplementary information accompanies this paper at 10.1038/s41598-018-20675-9.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Hayward MW, et al. Ecologists need robust survey designs, sampling and analytical methods. J. Appl. Ecol. 2015;52:286–290. doi: 10.1111/1365-2664.12408. [DOI] [Google Scholar]
- 2.Stephens PA, Pettorelli N, Barlow J, Whittingham MJ, Cadotte MW. Management by proxy? The use of indices in applied ecology. J. Appl. Ecol. 2015;52:1–6. doi: 10.1111/1365-2664.12383. [DOI] [Google Scholar]
- 3.Popescu VD, Artelle KA, Pop MI, Manolache S, Rozylowicz L. Assessing biological realism of wildlife population estimates in data‐poor systems. J. Appl. Ecol. 2016;53:1248–1259. doi: 10.1111/1365-2664.12660. [DOI] [Google Scholar]
- 4.Bauer H, et al. Lion (Panthera leo) populations are declining rapidly across Africa, except in intensively managed areas. PNAS. 2015;112:14894–14899. doi: 10.1073/pnas.1500664112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Gopalaswamy AM, Delampady M, Karanth KU, Kumar N, Macdonald DW. An examination of index‐calibration experiments: counting tigers at macroecological scales. Methods Ecol. Evol. 2015;6:1055–1066. doi: 10.1111/2041-210X.12351. [DOI] [Google Scholar]
- 6.Creel S, et al. Questionable policy for large carnivore hunting. Science. 2015;350:1473–1475. doi: 10.1126/science.aac4768. [DOI] [PubMed] [Google Scholar]
- 7.Riggio J, et al. Lion populations may be declining in Africa but not as Bauer et al. suggest. PNAS. 2016;113:E107. doi: 10.1073/pnas.1521506113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Harihar, A., Chanchani, P., Pariwakam, M., Noon, B. R. & Goodrich, J. Defensible inference: Questioning global trends in tiger populations. Cons. Lett. 10, 502–505.
- 9.Duchamp C, et al. Wolf monitoring in France: a dual frame process to survey time-and space-related changes in the population. Hystrix. 2011;23:14–28. [Google Scholar]
- 10.Epstein Y, López-Bao JV, Chapron G. A legal-ecological understanding of favorable conservation status for species inEurope. Cons. Lett. 2016;9:81–88. doi: 10.1111/conl.12200. [DOI] [Google Scholar]
- 11.Beyer, D. E., Peterson, R. O., Vucetich, J. A. & Hammill, J. H. Wolf population changes in Michigan. In Wydeven, A. P., VanDeelen, T. R., Heske, E. J. (Eds) Recovery of gray wolves in the Great Lakes region of the United States (pp. 65–85). Springer Publishing, New York, New York, USA (2009).
- 12.López-Bao JV, et al. Toothless wildlife protection laws. Biodivers. Conserv. 2015;24:2105–2108. doi: 10.1007/s10531-015-0914-8. [DOI] [Google Scholar]
- 13.Swedish Parliament. A new predator management. Environment and Agriculture Committee report 2009/10:MJU8 (2009).
- 14.Norwegian Environment Agency, http://www.environment.no/goals/1.-biodiversity/target-1.2/status-of-specific-threatened-species/four-wolf-litters/ (2016).
- 15.U.S. Fish and Wildlife Service [USFWS]. The reintroduction of gray wolves to Yellowstone National Park and Central Idaho. Appendix 9. Final Environmental Impact Statement. Denver, Colorado, USA (1994).
- 16.Ausband DE, et al. Surveying predicted rendezvous sites to monitor gray wolf populations. J. Wildl. Manage. 2010;74:1043–1049. doi: 10.2193/2009-303. [DOI] [Google Scholar]
- 17.Jiménez J, et al. Multimethod, multistate Bayesian hierarchical modeling approach for use in regional monitoring of wolves. Conserv. Biol. 2016;30:883–893. doi: 10.1111/cobi.12685. [DOI] [PubMed] [Google Scholar]
- 18.Chapron G, et al. Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science. 2014;346:1517–1519. doi: 10.1126/science.1257553. [DOI] [PubMed] [Google Scholar]
- 19.Kaczensky, P. et al. Status, management and distribution of large carnivores: bear, lynx, wolf and wolverine: in Europe. Report to the EU Commission, p 272 (2013).
- 20.Chapron G, et al. Estimating wolf (Canis lupus) population size from number of packs and an individual based model. Ecol. Mod. 2016;339:33–44. doi: 10.1016/j.ecolmodel.2016.08.012. [DOI] [Google Scholar]
- 21.Piaggio AJ, et al. A noninvasive method to detect Mexican wolves and estimate abundance. Wildl. Soc. Bull. 2016;40:321–330. doi: 10.1002/wsb.659. [DOI] [Google Scholar]
- 22.Boitani, L. & Powell, R. A. Carnivore ecology and conservation: a handbook of techniques. Oxford University Press (2012).
- 23.Long, R. A., MacKay, P., Zielinski, W. J. & Ray, J. C. Noninvasive survey methods for carnivores. Island Press, Washington D. C. (2008).
- 24.O’Connell, A. F., Nichols, J. D. & Karanth, K. U. Camera traps in animal ecology. Methods and analyses. Springer, Tokyo (2011).
- 25.Palacios V, López-Bao JV, Llaneza L, Fernández C, Font E. Decoding group vocalizations: The acoustic energy distribution of chorus howls is useful to determine wolf reproduction. PLOS ONE. 2016;11:e0153858. doi: 10.1371/journal.pone.0153858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Llaneza L, García EJ, López-Bao JV. Intensity of territorial marking predicts wolf reproduction: Implications for wolf monitoring. PLOS ONE. 2014;9:e93015. doi: 10.1371/journal.pone.0093015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Marucco F, et al. Wolf survival and population trend using non-invasive capture–recapture techniques in the Western Alps. J. Appl. Ecol. 2009;46:1003–1010. doi: 10.1111/j.1365-2664.2009.01696.x. [DOI] [Google Scholar]
- 28.Efford MG, Dawson DK, Robbins CS. DENSITY: Software for analysing capture-recapture data from passive detector arrays. Anim. Biodiv. Conserv. 2004;27:217–228. [Google Scholar]
- 29.Furnas BJ, Landers RH, Callas RL, Matthews SM. Estimating population size of fishers (Pekania pennanti) using camera stations and auxiliary data on home range size. Ecosphere. 2017;8:e01747. doi: 10.1002/ecs2.1747. [DOI] [Google Scholar]
- 30.Popescu VD, Iosif R, Pop MI, Chiriac S. Bouroș, G. & Furnas, B.J. Integrating sign surveys and telemetry data for estimating brown bear (Ursus arctos) density in the Romanian Carpathians. Ecol. Evol. 2017;7:7134–7144. doi: 10.1002/ece3.3177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Royle, J. A., Fuller, A. K. & Sutherland, C. Unifying population and landscape ecology with spatial capture-recapture. Ecography, in press, 10.1111/ecog.03170.
- 32.Royle, J. A., Chandler, R. B., Sollman, R. & Gardner, B. Spatial capture-recapture. Elsevier. Academic Press (2014).
- 33.Tenan S, Pedrini P, Bragalanti N, Groff C, Sutherland C. Data integration for inference about spatial processes: A model-based approach to test and account for data inconsistency. PLOS ONE. 2017;12:e0185588. doi: 10.1371/journal.pone.0185588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Elliot NB, Gopalaswamy AM. Towards accurate and precise estimates of lion density. Conserv. Biol. 2016;31:934–943. doi: 10.1111/cobi.12878. [DOI] [PubMed] [Google Scholar]
- 35.Goldberg JF, et al. Examining temporal sample scale and model choice with spatial capture-recapture models in the common leopard Panthera pardus. PLOS ONE. 2015;10:e0140757. doi: 10.1371/journal.pone.0140757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Molina S, Fuller AK, Morin DJ, Royle JA. Use of spatial capture – recapture to estimate density of Andean bears in northern Ecuador. Ursus. 2017;28:117–126. doi: 10.2192/URSU-D-16-00030.1. [DOI] [Google Scholar]
- 37.Bischof R, Brøseth H, Gimenez O. Wildlife in a politically divided world: Insularism inflates estimates of brown bear abundance. Cons. Lett. 2016;9:122–130. doi: 10.1111/conl.12183. [DOI] [Google Scholar]
- 38.Sollmann R, Gardner B, Belant JL, Wilton CM, Beringer J. Habitat associations in a recolonizing, low-density black bear population. Ecosphere. 2016;7:1–11. doi: 10.1002/ecs2.1406. [DOI] [Google Scholar]
- 39.Morehouse AT, Boyce MS. Grizzly bears without borders: Spatially explicit capture-recapture in southwestern Alberta. J. Wildl. Manage. 2016;80:1152–1166. doi: 10.1002/jwmg.21104. [DOI] [Google Scholar]
- 40.Efford MG. Estimation of population density by spatially explicit capture–recapture analysis of data from area searches. Ecology. 2011;92:2202–2207. doi: 10.1890/11-0332.1. [DOI] [PubMed] [Google Scholar]
- 41.Efford MG, Dawson DK, Borchers DL. Population density estimated from locations of individuals on a passive detector array. Ecology. 2009;90:2676–2682. doi: 10.1890/08-1735.1. [DOI] [PubMed] [Google Scholar]
- 42.Sandell, M. The mating tactics and spacing patterns of solitary carnivores. In Gittleman, J. L. (Ed). Carnivore Behavior, Ecology, and Evolution (pp. 164–182). Ithaca. Cornell University Press (1989).
- 43.Mech, L. D. & Boitani, L. Wolves: Behavior, ecology, and conservation. University of Chicago Press (2003).
- 44.Jedrzejewski W, Schmidt K, Theuerkauf J, Jedrzejewska B, Kowalczyk R. Territory size of wolves Canis lupus: linking local (Bialowieza Primeval Forest, Poland) and Holarctic-scale patterns. Ecography. 2007;30:66–76. [Google Scholar]
- 45.Fritts, S. H. & Mech, L. D. Dynamics, movements, and feeding ecology of a newly protected wolf population in northwestern Minnesota. Wildl. Monogr. 80 (1981).
- 46.Peterson, R. O., Woolington, J. D. & Bailey, T. N. Wolves of the Kenai Peninsula, Alaska. Wildl. Monogr. 88 (1984).
- 47.Benson JF, Patterson BR. Spatial overlap, proximity, and habitat use of individual wolves within the same packs. Wildl. Soc. Bull. 2015;39:31–40. doi: 10.1002/wsb.506. [DOI] [Google Scholar]
- 48.Llaneza, L., García, E. J., Palacios, V. & López-Bao, J. V. Wolf monitoring in Galicia, 2013–2014. Report to the Spanish Ministry of Agriculture, Food and Environment, Spain (2015).
- 49.Pacheco C, et al. Spatial assessment of wolf-dog hybridization in a single breeding period. Sci. Rep. 2017;7:42475. doi: 10.1038/srep42475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Godinho R, et al. Real‐time assessment of hybridization between wolves and dogs: combining non-invasive samples with ancestry informative markers. Mol. Ecol. Res. 2015;15:317–328. doi: 10.1111/1755-0998.12313. [DOI] [PubMed] [Google Scholar]
- 51.Nakamura M, et al. Evaluating the predictive power of field variables for species and individual molecular identification on wolf non-invasive samples. Eur. J. Wildl. Res. 2017;63:53. doi: 10.1007/s10344-017-1112-7. [DOI] [Google Scholar]
- 52.Llaneza, L. Wolves in human-dominated landscapes of Northwestern Iberian Peninsula. PhD thesis. University ofSantiago de Compostela, Spain (2016).
- 53.Chandler, R. Unmarked Workshop, http://sites.google.com/site/spatialcapturerecapture/workshop–athens-ga-3-2015/day-4 (2015).
- 54.NIMBLE Development Team. NIMBLE: An R Package for Programming with BUGS models (2015).
- 55.R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/ (2017).
- 56.Gelman, A., Carlin, J. B., Stern, H. S. & Rubin, D. B. Bayesian data analysis (Vol. 2). Boca Raton, FL, USA: Chapman & Hall/CRC (2013).
- 57.Plummer M, Best N, Cowles K, Vines K. CODA: Convergence diagnosis and output analysis for MCMC. R news. 2006;6:7–11. [Google Scholar]
- 58.Calenge, C. Home range estimation. adehabitatHR package for R. Version 0.3 (2013).
- 59.Sazatornil V, et al. The role of human-related risk in breeding site selection by wolves. Biol. Cons. 2016;201:103–110. doi: 10.1016/j.biocon.2016.06.022. [DOI] [Google Scholar]
- 60.Llaneza, L., García, E. J., Palacios, V., Sazatornil, V. & López-Bao, J. V. Pack size in the Iberian wolf (Canis lupus signatus). Oral communication. Abstract-book of the III Iberian Wolf Congress: p. 19 (2012).
- 61.Schmidt JH, Rattenbury KL, Robison HL, Gorn TS, Shults BS. Using non-invasive mark-resight and sign occupancy surveys to monitor low-density brown bear populations across large landscapes. Biol. Cons. 2017;207:47–54. doi: 10.1016/j.biocon.2017.01.005. [DOI] [Google Scholar]
- 62.Sollmann R, et al. Combining camera-trapping and noninvasive genetic data in a spatial capture–recapture framework improves density estimates for the jaguar. Biol. Cons. 2013;167:242–247. doi: 10.1016/j.biocon.2013.08.003. [DOI] [Google Scholar]
- 63.Rothman RJ, Mech LD. Scent-marking in lone wolves and newly formed pairs. Anim. Behav. 1979;27:750–760. doi: 10.1016/0003-3472(79)90010-1. [DOI] [Google Scholar]
- 64.Stevenson BC, et al. A general framework for animal density estimation from acoustic detections across a fixed microphone array. Methods Ecol. Evol. 2015;6:38–48. doi: 10.1111/2041-210X.12291. [DOI] [Google Scholar]
- 65.Reich B, Gardner B. A spatial capture-recapture model for territorial species. Environmetrics. 2014;25:630–637. doi: 10.1002/env.2317. [DOI] [Google Scholar]
- 66.Ripple WJ, et al. Saving the world’s terrestrial megafauna. BioScience. 2016;66:807–812. doi: 10.1093/biosci/biw092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Johnson, N. C., Malk, A. J., Szaro, R. C. & Sexton, W. T. Ecological stewardship: A common reference for ecosystem management. Oxford Elsevier Science Ltd (1999).
- 68.Bormann, B. T., Haynes, R. W. & Martin, J. R. Adaptive management of forest ecosystems: did some rubber hit the road? BioScience57, 186–191.
- 69.LIFE DinAlp Bear, http://dinalpbear.eu/en/ (2017).
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.