

Abstract
MicroRNAs (miRNAs/miRs) are involved in the regulation of various types of cancer, either as oncogenes or tumor suppressors. miR302a has been reported that it could suppress tumor cell proliferation by inhibiting Akt in prostate cancer. The present study examined the effect of miR302a on proliferation and invasion in esophageal cancer cell lines. The expression levels of miR302a in esophageal cancer cell lines was determined by reverse transcription-polymerase chain reaction. Subsequently, miR302a mimics were transfected into esophageal cancer cells, and cell viability and invasion were assessed by MTT and Transwell assays. In addition, the effects of miR302a on the mitogen-activated protein kinase (MAPK) and phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) signaling pathways were investigated by western blot analysis. The results revealed that miR302a expression was significantly decreased in the esophageal cancer cell lines compared with a healthy esophagus epithelium cell line. Upregulation of miR302a inhibited the proliferation and invasion of esophageal cancer cells, and decreased the phosphorylation of extracellular signal-regulated kinase 1/2 and Akt. Taken together, the results of the present study indicated that miR302a overexpression inhibited the proliferation and invasion of esophageal cancer cells through suppression of the MAPK and PI3K/Akt signaling pathways, indicating the potential value of miR302a as a treatment target for human esophageal cancer.
Keywords: microRNA 302a, esophagus cancer, viability, invasion, mitogen-activated protein kinase, phosphoinositide 3-kinase/protein kinase B, phosphorylated extracellular signal-regulated kinase 1/2, phosphorylated protein kinase B
Introduction
Esophageal cancer is a common primary malignant tumor of the digestive tract, and its morbidity and mortality rank eighth and sixth, respectively, among malignant tumors globally (1). Esophageal cancer affects >450,000 people worldwide and the incidence rate is increasing (2). The overall 5-year survival rate of patients with esophageal carcinoma ranges from 13–19%, based on 2006–2012 data (3–5). Although developments in chemotherapy have improved the prognosis of esophageal cancer, drug resistance remains the primary obstacle to successful treatment (6). Therefore, in order to improve treatment efficacy and prognosis in patients, further investigation of the genesis and underlying developmental mechanisms of esophageal carcinoma is crucial.
MicroRNAs (miRNAs/miRs) are short (~22 nucleotides), non-protein-coding RNAs. A growing body of evidence indicates that miRNAs serve crucial functions in diverse cellular processes through gene regulation (7). Furthermore, miRNA expression profiling has revealed that certain miRNAs are associated with tumor development, progression and response to therapy (7). For example, miR302a, a member of the microRNA-302 family, was initially identified in human embryonic stem cells and human embryonic carcinoma cells (8). Increasing reports have indicated that miR302a participates in various cell differentiation or tumor metastasis-associated signaling pathways via targeting different molecules. Guo et al (9) demonstrated that miR302a is involved in the inhibition of ovarian cancer cell proliferation, and enhances cell apoptosis through targeting syndecan 1. In addition, miR302a suppresses tumor cell proliferation by inhibiting protein kinase B (AKT) in prostate cancer (10). However, miR302a expression and its involvement in esophageal cancer remain undetermined, and the underlying mechanisms of miR302a in esophageal cancer cells remain unknown.
The present study revealed that miR302a expression was decreased in esophageal cancer cell lines compared with a healthy esophageal epithelial cell line. Furthermore, miR302a overexpression inhibits esophageal cancer cell proliferation and invasion. Finally, miR302a was revealed to act as a tumor suppressor via regulation of the mitogen-activated protein kinase (MAPK) and phosphoinositide 3-kinase (PI3K)/Akt signaling pathway in the cell lines TE-10 and ECA109.
Materials and methods
Cell culture
The human esophageal cancer cell lines, TE-1, TE-10, TE-11, ECA109, and the healthy human esophageal epithelial cell line Het-1A were purchased from Shanghai Jining Industry Co., Ltd. (Shanghai, China) TE-1 (cat. no. JN-B1846; http://www.shjning.com/plus/view.php?aid=42526), TE-10 (cat.no. JN-B1582; http://www.shjning.com/plus/view.php?aid=42241), TE-11 (cat. no. JN-A2623; http://www.shjning.com/plus/view.php?aid=39797), ECA109 (cat. no. JN-B1684; http://www.shjning.com/plus/view.php?aid=42362) and Het-1A (cat. no. JN-4897; http://www.shjning.com/plus/view.php?aid=39249) cells were grown in Dulbecco's modified Eagle's medium (DMEM) medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), supplemented with 10% fetal bovine serum (FBS; Hangzhou Sijiqing Biology Engineering Materials Co., Ltd., Hangzhou, China). Cells were cultured at 37°C in a 5% CO2 atmosphere. Cells were passaged with 0.25% trypsin (Sigma-Aldrich; Merck KGaA, Damstadt, Germany) at a 1:3 ratio every 3 days.
Cell transfection
Cells (TE-10, ECA109) were maintained until mid-log phase, and then digested with trypsin and seeded into 6-well plates (Sigma-Aldrich; Merck KGaA) at a density of 1×105 cells per well. Following culture for 48 h, when cells reached 80% confluence, cells were processed for transfection. miR302a mimics (5′-ACUUAAACGUGGAUGUACUUGCU-3′), miR302a inhibitor (5′-UGAAUUUGCACCUACAUGAACGA-3′) and their negative control oligonucleotides (5′-UCGUUCAUGUAGGUGCAAAUUCA-3′; all synthesized by Guangzhou Ribobio Co., Ltd, Guangzhou, China) were transfected into TE-10 and ECA109 cells using Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to the manufacturer's protocol. The reagents, including 1 µg miR302a mimics or inhibitors and 3 µl Lipofectamine 2000, were diluted into 50 µl Opti-Minimal Essential medium (Gibco; Thermo Fisher Scientific, Inc.), and incubated for 20 min at room temperature. Then, the miRNA-lipid complex was added to the cells (10 µl/well) and gently blended. After 48 h of incubation at 37°C, the transfected cells were collected for analysis of miRNA and protein expression.
MTT assays
Cells at the mid-log phase were seeded in 96-well plates (Sigma-Aldrich; Merck KGaA) at a density of 1×104 cells/well in 100 µl DMEM medium, with 5 replicates for each group. Cells were incubated with 5 mg/ml MTT solution (10 µl) at 37°C in a 5% CO2 atmosphere for 4 h. Following incubation, the medium was removed, 150 µl DMSO (Sigma-Aldrich; Merck KGaA). The absorbance at 490 nm was detected using a spectrophotometer (Thermo Fisher Scientific, Inc.), and cell viability was measured once per day for 7 days.
Transwell assays
Cell invasion assays were performed in a 24-well Transwell chambers with 8-mm pore size inserts according to the manufacturer's protocol (Corning Incorporated, Corning, NY, USA). Matrigel matrix gel (100 µl; BD Biosciences, Franklin Lakes, NJ, USA) was diluted with DMEM without FBS overnight at a ratio of 1:6 and was added to the upper chamber of the 24-well Transwell plate. For the assay, 1×105 cells were seeded into the upper chamber with 100 µl DMEM without FBS. In the lower chamber, 600 µl DMEM supplemented with 10% FBS was added. Following incubation for 24 h at 37°C in a 5% CO2 atmosphere, the non-invading cells were removed from the plate with a cotton swab. Cells on the lower surface of the membrane were fixed with 4% paraformaldehyde for 10 min at room temperature and stained with 0.1% crystal violet for 3 min at room temperature. The cells were then counted in at least 5 random fields using an inverted microscope (Olympus Corporation, Tokyo, Japan).
Reverse transcription-polymerase chain reaction (RT-PCR)
Total RNA was extracted using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.). To quantify miR302a expression, total RNA was first polyadenylated and reverse-transcribed using the NCode miRNA first-Strand cDNA synthesis kit (Takara Biotechnology Co., Ltd., Dalian, China), according to the manufacturer's protocol, followed by PCR using 1 µg cDNA via TaqMan® Universal Master Mix (Life technology, Carlsbad, CA, USA), according to the manufacturer's protocol. The thermocycler conditions were as follows: 95°C for 10 sec, followed by 40 amplification cycles (95°C for 5 sec, 60°C for 30 sec) and 60°C for 12 min. Primers (Sangon Biotech Co., Ltd., Shanghai, China) used for miR302a were as follows: Forward, 5′-CGTGGATGTACTTGCTTTGAA-3′ and reverse, 5′-TCACCAAAACATGGAAGCAC-3′. β-actin was used as an internal control, and every sample was replicated at least five times, β-actin forward: 5′-ACCGAGCGCGGCTACAG-3′, β-actin reverse: 5′-CTTAATGTCACGCACGATTTCC-3′. PCR products were run on a 3% agarose gel in 1X TAE (0.04 M Tris, 1 mM EDTA, 0.02 M acetic acid, pH 8.2–8.4) buffer, and then the gels were stained with ethidium bromide to a final concentration of 0.5 µg/ml (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at 55°C for 1 min, observed and pictured using Image J version 2 software (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The band intensities were quantified using Image J software. For relative quantification, miR302a expression was normalized to β-actin expression in the corresponding sample.
Protein extraction and western blot analysis
After 48 h of incubation at 37°C, the transfected cells were collected for cell lysis by treating with radio immunoprecipitation assay (Solarbio, Beijing, China) lysis buffer and protease inhibitor (Beyotime, Shanghai, China) on ice for 30 min. Then, it was centrifuged at 12,000 × g for 5 min at 4°C, the supernatant was extracted and the protein concentration was measured using a BCA kit (Beyotime), according to the manufacturer's instructions. Extracted protein (30 µg) was added to 50 µl Laemmli sample buffer (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), and boiled for 5 min. The protein was then loaded and separated on 10% Ready Gels, and then transferred to a polyvinylidene fluoride membrane (EMD Millipore, Billerica, MA, USA). Membranes were blocked with 5% non-fat dry milk for 1 h at room temperature. Primary monoclonal mouse anti-human β-actin antibodies (1:5,000; cat no. ab7813), anti-phosphorylated (p-) extracellular signal-regulated kinase (ERK)1/2 (1:500; cat no. ab214362), anti-ERK1/2 (1:500; cat no. ab17942), anti-p-protein kinase B (Akt; 1:500; cat no. ab38499) and total Akt (1:500; cat no. ab6789; all from Abcam, Cambridge, MA, USA) were added and incubated overnight at 4°C. Following washing three times in 0.1% Tween 20-TBS (TBST), the membranes were incubated with secondary polyclonal goat anti-mouse horseradish peroxidase (HRP)-conjugated antibodies (1:5,000; cat no. ab179463; Abcam, Cambridge, MA, USA) overnight at 4°C, and then washed three times in TBST. The immunoreactive bands were visualized using enhanced chemiluminescence reagents, according to the manufacturer's protocol (Western Lightning Plus-ECL; PerkinElmer, Inc., Waltham, MA, USA), and then assessed using the Quantity One software version 4.62 (Bio-Rad Laboratories, Inc.).
Statistical analysis
All statistical analyses were performed using SPSS statistical software, version 19.0 (IBM Corp., Armonk, NY, USA). Values are presented as the mean ± standard deviation. Statistical differences among the groups were tested by one-way analysis of variance, followed by Fisher's least significant difference tests. P<0.05 was considered to indicate a statistically significant difference.
Results
Esophageal cancer cell lines have reduced miR302a expression compared with healthy esophageal cells
To investigate the function of miR302a in esophageal cancer cell lines, the expression of miR302a was measured by RT-PCR in four esophageal cancer cell lines (TE-1, TE-10, TE-11, ECA109) and a healthy human esophageal epithelial cell (Het-1A). As presented in Fig. 1, miR302a expression in all four esophageal cancer cell lines was significantly decreased compared with the healthy esophageal epithelial cell line. Then, two esophageal cancer cell lines with similar miR302a levels, TE-10, and ECA109, were selected for follow-up experiments.
Figure 1.
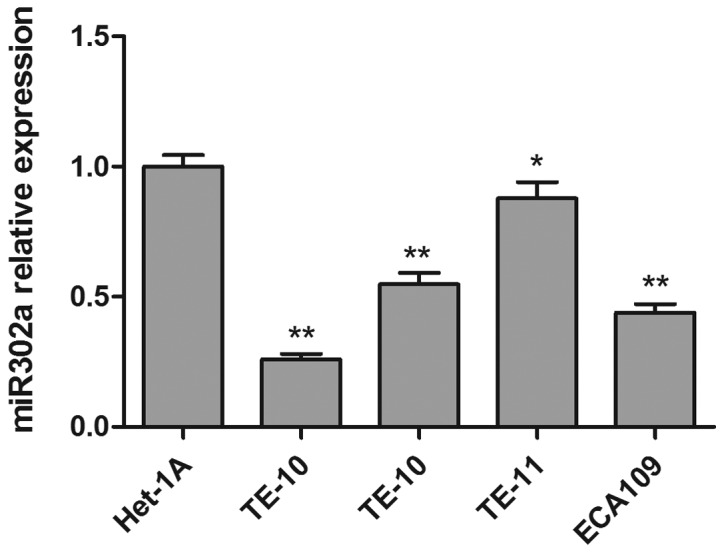
miR302a expression in Het-1A cells and 4 esophageal cancer cell lines (TE-1, TE-10, TE-11 and ECA109). Data are presented as the mean ± standard deviation. *P<0.05 and **P<0.01 vs. Het-1A. miR302a, microRNA302a.
miR302a significantly inhibits esophageal cancer cell proliferation
To further evaluate the effect of miR302a, miR302a mimics or inhibitors were stably overexpressed in TE-10 and ECA109 cell lines, and the expression of miR302a was confirmed by RT-PCR. Compared with the control group, miR302a expression was significantly increased in the miR302a mimic group (P<0.01), and miR302a expression was significantly decreased in the miR302a inhibitor group in TE-10 and ECA109 cells (P<0.01; Fig. 2).
Figure 2.
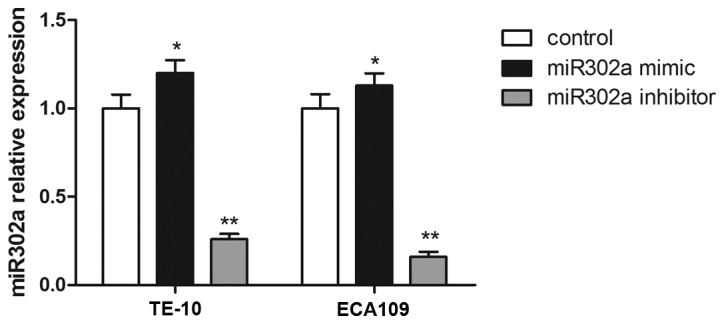
miR302a expression in transfected TE-10 and ECA109 esophageal cancer cells. Data are presented as the mean ± standard deviation. *P<0.05 and **P<0.01 vs. control. miR302a, microRNA302a.
Then, MTT assays were performed to examine whether miR302a overexpression inhibited esophageal cancer cell proliferation in vitro. Compared with the control group, cell proliferation in the miR302a mimic group was significantly decreased (P<0.01), and cell proliferation in the miR302a inhibitor group was significantly increased (P<0.01), from the third day to the seventh day in TE-10 and ECA109 cells (Fig. 3). The results demonstrated that miR302a significantly inhibited the viability of esophageal cancer cells.
Figure 3.

Cell viability was determined using an MTT assay in TE-10 and ECA109 cells. Data are presented as the mean ± standard deviation. *P<0.05 and **P<0.01 vs. control. miR302a, microRNA302a; OD, optical density.
miR302a significantly inhibits esophageal cancer cell invasion
The aforementioned results revealed that miR302a significantly inhibited the proliferation of esophageal cancer cells. Next, invasion was assessed in esophageal cancer cells transfected with miR302a mimics or inhibitors using Transwell assays. As presented in Fig. 4, the number of invasive cells in the miR302a mimics group was significantly decreased compared with the control group in TE-10 and ECA109 cells (P<0.01), and the number of invasive cells in the miR302a inhibitor group was significantly increased in TE-10 and ECA109 cells compared with the control group (P<0.01). These results revealed that overexpression of miR302a inhibited the invasion of esophageal cancer cells.
Figure 4.

Overexpression of miR302a inhibited the invasion of esophageal cancer cells. (A) Cell invasion was measured using Transwell assays. Representative images of crystal violet-stained invasive cells are pictured (magnification, ×400). (B) Quantification of the invasion assay. Data are presented as the mean ± standard deviation. *P<0.05 and **P<0.01 vs. control; ##P<0.01 vs. miR302a mimic group. miR302a, microRNA302a.
Overexpression of miR302a suppresses the phosphorylation of proteins in the MAPK and PI3K/Akt signaling pathways
As miR302a has been demonstrated to be associated with cell viability and invasion in esophageal cancer cells, the underlying mechanisms of these functions were subsequently explored. The expression of p-Akt and p-ERK1/2, which are crucial downstream effectors of the MAPK and PI3K/Akt signaling pathways, were assessed in the different treatment groups. As presented in Figs. 5 and 6, overexpression of miR302a had no significant effect on total Akt and ERK1/2 expression levels, while the phosphorylation level of Akt and ERK1/2 was significantly decreased in the miR302a mimic group (P<0.05) and increased in the miR302a inhibitor group (P<0.01), compared with the control group in the two cell lines.
Figure 5.

Expression of p-ERK1/2 and total ERK1/2 in TE-10 and ECA109 cells transfected with miR302a mimics or miR302a inhibitors, assessed using western blotting. Expression of p-ERK1/2 and total ERK1/2 in (A) TE-10 cells and (B) ECA109 cells transfected with miR302a mimics or miR302a mimic inhibitor. Data are presented as the mean ± standard deviation. *P<0.05 and **P<0.01 vs. control; ##P<0.01 vs. miR302a mimic group. p-, phosphorylated; ERK, extracellular signal-regulated kinase; miR302a, microRNA302a.
Figure 6.
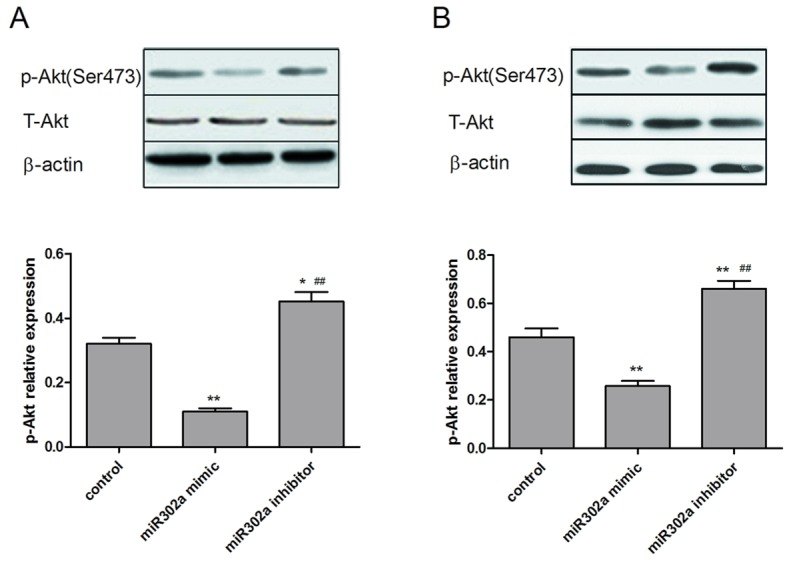
Expression of p-Akt and total Akt in TE-10 and ECA109 cells transfected with miR302a mimics or miR302a inhibitors, assessed using western blotting. Expression of p-Akt and total Akt in (A) TE-10 cells and (B) ECA109 cells transfected with miR302a mimics or miR302a mimic inhibitors. *P<0.05 and **P<0.01 vs. control; ##P<0.01 vs. miR302a mimic group. p-, phosphorylated; Akt, protein kinase B; miR302a, microRNA302a.
Discussion
The results of the present study revealed that miR302a expression was significantly downregulated in several esophageal cancer cell lines compared with a healthy esophageal epithelial cell line. Furthermore, overexpression of miR302a significantly inhibited the proliferation and invasion of esophageal cancer cells. Finally, increased miR302a expression significantly inhibited p-Akt and p-ERK1/2 protein expression levels, indicating that miR302a may act as an upstream regulator of MAPK and PI3K/Akt signaling in esophagus cancer.
miR302 family members have been reported to regulate cell proliferation, invasion and apoptosis in certain types of cancer cell. Maadi et al (11) demonstrated that miR-302 inhibited cancer cell proliferation, angiogenesis and invasion by reversing the epithelial-mesenchymal transition in A-375 melanoma cells and HT-29 colorectal cancer cells. miR302a also increases 5-fluorouracil-induced cell death and inhibits cell viability in human colon cancer cells by inhibiting Akt signaling (12). miRNA expression profiles have been reported in esophageal cancer, but the function of miR302a in esophageal cancer remains unknown (13). The present study revealed that miR302a was significantly downregulated in esophageal cancer cell lines (TE-1, TE-10, TE-11, ECA109) compared with the healthy esophageal cancer cell line Het-1A and, to the best of our knowledge, its downregulation was revealed to be associated with cell proliferation and invasion in TE-10 and ECA109 cell lines for the first time. Furthermore, overexpression of miR302a significantly inhibited the viability and invasion of esophageal cancer cells, and silencing of miR302a elicited the opposite effects. The results of the present study may aid the development and improvement of diagnosis and treatment of esophageal cancer in the future.
MAPK signal transduction pathways are among the most widespread cellular regulation mechanisms (14). The Ras/ERK pathway is the prototypic MAPK pathway, being a key signaling pathway that is involved in the regulation of healthy cell proliferation, survival, growth and differentiation. Ras protein is the most frequently mutated protein in human tumors, and in its active state affects cell growth, differentiation, cytoskeleton, protein transport and secretion (15). Dysregulation of the ERK pathway contributes to ~1/3 of all types of human cancer (16). Certain reports have demonstrated that p-ERK inhibits cell proliferation and induces the apoptosis of gastric cancer cells treated with β-elemene (17), and sustained, β-sitosterol-induced ERK phosphorylation impedes renal tumor transformation promotion and maintenance (18). Increased phosphorylated ERK could promote tumor cell proliferation and invasion, and affects cell differentiation, inhibition of apoptosis. In addition, the PI3K/Akt signaling pathway is also involved in the regulation of cell growth, metabolism, proliferation, glucose homeostasis and vesicle trafficking, and dysregulation of PI3K/Akt pathway components accounts for ~ 30% of all cases of human cancer (19). In particular, the Akt family of serine/threonine kinases has emerged as a critical target of PI3K in human tumors. P-Akt increases cell proliferation and inhibits cell apoptosis of multiple types of cancer. For example, p-Akt promotes cell proliferation and survival in vitro and serves an important function in prostate cancer progression, as well as the prediction of recurrence (20), and decreased expression of p-Akt inhibits polydatin-induced cell proliferation and induces apoptosis in laryngeal cancer and HeLa cells (21). Furthermore, overexpression of p-Akt promotes the proliferation and tumorigenesis of the esophageal cancer cell line ECA-109 (22). Therefore, miR302a may have targeted p-ERK1/2 and p-Akt through the MAPK and PI3K signaling pathways, respectively, and this may be its primary contribution to the inhibition of cell proliferation in TE-10 and ECA109 cells. Consistent with previous results, in the present study, the phosphorylation levels of Akt and ERK1/2 were significantly decreased in the miR302a mimics group (P<0.05), and increased in the miR302a inhibitor group (P<0.01) (23–26). Furthermore, total Akt and ERK1/2 expression levels were also determined by western blot analysis in TE-10 and ECA109 cells, and overexpression of miR302a did not markedly differ between groups. miR302a overexpression has been demonstrated to significantly inhibit cell proliferation by inhibiting Akt in colon cancer and prostate cancer (10,27). Furthermore, miR-302 has also been reported to inhibit cancer cell proliferation via the Akt signaling pathway, and the microRNA-302-367 cluster suppressed the proliferation of cervical carcinoma cells through the novel target Akt1 (28). Sun et al (29) also revealed that miR302a inhibited Akt expression by directly binding to its 3′ untranslated region, resulting in subsequent alterations of the Akt-glycogen synthase kinase 3β-cyclin D1 pathway. However, to date, few studies have examined miR302a-induced inhibition of cell proliferation via the MAPK signaling pathway in cancer cells, and to the best of our knowledge no prior studies have been conducted concerning whether miR302a affected the proliferation of esophageal cancer cells by regulating the MAPK or PI3K/Akt signaling pathways. The present study revealed that miR302a expression was decreased in esophageal cancer cell lines compared with a healthy esophageal epithelial cell line, and investigated the relevance of miR302a expression to the MAPK and PI3K/Akt signaling pathways.
In conclusion, our group demonstrated that miR302a overexpression inhibited the viability and invasion of esophageal cancer cells via the MAPK and PI3K/Akt signaling pathways. These results may provide a potential therapeutic target to suppress the development, invasion and metastasis of esophageal cancer cells. However, the effects of miR302a determined by the present study were all based on in vitro experiments, and further in vivo studies should be performed in the future.
References
- 1.Stewart BW, Wild CP, editors. World cancer report 2014. Lyon, France: Int Agency Res Cancer; 2014. [Google Scholar]
- 2.Pennathur A, Gibson MK, Jobe BA, Luketich JD. Oesophageal carcinoma. Lancet. 2013;381:400–412. doi: 10.1016/S0140-6736(12)60643-6. [DOI] [PubMed] [Google Scholar]
- 3.Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65:87–108. doi: 10.3322/caac.21262. [DOI] [PubMed] [Google Scholar]
- 4.Siegel RL, Miller KD, Jemal A. Cancer statistics, 2015. CA Cancer J Clin. 2015;65:5–29. doi: 10.3322/caac.21254. [DOI] [PubMed] [Google Scholar]
- 5.Valmasoni M, Pierobon ES, Ruol A, De Pasqual CA, Zanchettin G, Moletta L, Salvador R, Costantini M, Merigliano S. Endoscopic tumor length should be reincluded in the esophageal cancer staging system: Analyses of 662 consecutive patients. PLoS One. 2016;11:e0153068. doi: 10.1371/journal.pone.0153068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ghidini M, Petrelli F, Hahne JC, De Giorgi A, Toppo L, Pizzo C, Ratti M, Barni S, Passalacqua R, Tomasello G. Clinical outcome and molecular characterization of brain metastases from esophageal and gastric cancer: A systematic review. Med Oncol. 2017;34:62. doi: 10.1007/s12032-017-0919-0. [DOI] [PubMed] [Google Scholar]
- 7.Iorio MV, Croce CM. MicroRNA dysregulation in cancer: Diagnostics, monitoring and therapeutics. A comprehensive review. EMBO Mol Med. 2012;4:143–159. doi: 10.1002/emmm.201100209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Barroso-Deljesus A, Lucena-Aguilar G, Sanchez L, Ligero G, Gutierrez-Aranda I, Menendez P. The Nodal inhibitor Lefty is negatively modulated by the microRNA miR-302 in human embryonic stem cells. FASEB J. 2011;25:1497–1508. doi: 10.1096/fj.10-172221. [DOI] [PubMed] [Google Scholar]
- 9.Guo T, Wei Y, Lv S, Zhang C, Tian Y. MiR-302a inhibits the tumorigenicity of ovarian cancer cells by suppression of SDC1. Int J Clin Exp Pathol. 2015;8:4869–4880. [PMC free article] [PubMed] [Google Scholar]
- 10.Zhang GM, Bao CY, Wan FN, Cao DL, Qin XJ, Zhang HL, Zhu Y, Dai B, Shi GH, Ye DW. MicroRNA-302a suppresses tumor cell proliferation by inhibiting Akt in prostate cancer. PLoS One. 2015;10:e0124410. doi: 10.1371/journal.pone.0124410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Maadi H, Moshtaghian A, Taha MF, Mowla SJ, Kazeroonian A, Haass NK, Javeri A. Multimodal tumor suppression by miR-302 cluster in melanoma and colon cancer. Int J Biochem Cell Biol. 2016;81:121. doi: 10.1016/j.biocel.2016.11.004. [DOI] [PubMed] [Google Scholar]
- 12.Liu N, Li J, Zhao Z, Han J, Jiang T, Chen Y, Hou N, Huang C. MicroRNA-302a enhances 5-fluorouracil-induced cell death in human colon cancer cells. Oncol Rep. 2017;37:631–639. doi: 10.3892/or.2016.5237. [DOI] [PubMed] [Google Scholar]
- 13.Feber A, Xi L, Luketich JD, Pennathur A, Landreneau RJ, Wu M, Swanson SJ, Godfrey TE, Litle VR. MicroRNA expression profiles of esophageal cancer. J Thorac Cardiovasc Surg. 2008;135:255–260. doi: 10.1016/j.jtcvs.2007.08.055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Turjanski AG, Vaqué JP, Gutkind JS. MAP kinases and the control of nuclear events. Oncogene. 2007;26:3240–3253. doi: 10.1038/sj.onc.1210415. [DOI] [PubMed] [Google Scholar]
- 15.Roberts PJ, Der CJ. Targeting the Raf-MEK-ERK mitogen-activated protein kinase cascade for the treatment of cancer. Oncogene. 2007;26:3291–3310. doi: 10.1038/sj.onc.1210422. [DOI] [PubMed] [Google Scholar]
- 16.Schubbert S, Shannon K, Bollag G. Hyperactive Ras in developmental disorders and cancer. Nat Rev Cancer. 2007;7:295–308. doi: 10.1038/nrc2175. [DOI] [PubMed] [Google Scholar]
- 17.Li P, Zhou X, Sun W, Sheng W, Tu Y, Yu Y, Dong J, Ye B, Zheng Z, Lu M. Elemene induces apoptosis of human gastric cancer cell line BGC-823 via extracellular signal-regulated kinase (ERK)1/2 signaling pathway. Med Sci Monit. 2017;23:809–817. doi: 10.12659/MSM.903197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sharmila R, Sindhu G. Evaluate the antigenotoxicity and anticancer role of β-sitosterol by determining oxidative DNA damage and the expression of phosphorylated mitogen-activated protein kinases', C-fos, C-jun, and endothelial growth factor receptor. Pharmacogn Mag. 2017;13:95–101. doi: 10.4103/0973-1296.197634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Shaw RJ, Cantley LC. Ras, PI(3)K and mTOR signalling controls tumour cell growth. Nature. 2006;441:424–430. doi: 10.1038/nature04869. [DOI] [PubMed] [Google Scholar]
- 20.Hammerich KH, Frolov A, Li R, Ittmann M, Ayala GE. Cellular interactions of the phosphorylated form of AKT in prostate cancer. Hum Pathol. 2017;63:98–109. doi: 10.1016/j.humpath.2017.02.014. [DOI] [PubMed] [Google Scholar]
- 21.Li H, Shi B, Li Y, Yin F. Polydatin inhibits cell proliferation and induces apoptosis in laryngeal cancer and HeLa cells via suppression of the PDGF/AKT signaling pathway. J Biochem Mol Toxicol. 2017;31 doi: 10.1002/jbt.21900. [DOI] [PubMed] [Google Scholar]
- 22.Hong Y, Wen C, Du X, Hong Y, Chen W, Du X, Ning H, Chen H, Shi R, Lin S, et al. Upregulation of sex-determining region Y-box 9 (SOX9) promotes cell proliferation and tumorigenicity in esophageal squamous cell carcinoma. Oncotarget. 2015;6:31241–31254. doi: 10.18632/oncotarget.5160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Mundi PS, Sachdev J, McCourt C, Kalinsky K. AKT in cancer: New molecular insights and advances in drug development. Br J Clin Pharmacol. 2016;82:943–956. doi: 10.1111/bcp.13021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Briest F, Grabowski P. PI3K-AKT-mTOR-signaling and beyond: The complex network in gastroenteropancreatic neuroendocrine neoplasms. Theranostics. 2014;4:336–365. doi: 10.7150/thno.7851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Carpenter RL, Sirkisoon S, Zhu D, Rimkus T, Harrison A, Anderson A, Paw I, Qasem S, Xing F, Liu Y, et al. Combined inhibition of AKT and HSF1 suppresses breast cancer stem cells and tumor growth. Oncotarget. 2017;8:73947–73963. doi: 10.18632/oncotarget.18166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hamilton G, Abraham AG, Morton J, Sampson O, Pefani DE, Khoronenkova S, Grawenda A, Papaspyropoulos A, Jamieson N, McKay C, et al. AKT regulates NPM dependent ARF localization and p53mut stability in tumors. Oncotarget. 2014;5:6142–6167. doi: 10.18632/oncotarget.2178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Liu N, Li J, Zhao Z, Han J, Jiang T, Chen Y, Hou N, Huang C. MicroRNA-302a enhances 5-fluorouracil-induced cell death in human colon cancer cells. Oncol Rep. 2017;37:631–639. doi: 10.3892/or.2016.5237. [DOI] [PubMed] [Google Scholar]
- 28.Cai N, Wang YD, Zheng PS. The microRNA-302-367 cluster suppresses the proliferation of cervical carcinoma cells through the novel target AKT1. RNA. 2013;19:85–95. doi: 10.1261/rna.035295.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sun S, Zhang G, Wu Z, Shi W, Yang B, Li Y. MicroRNA-302a functions as a putative tumor suppressor in colon cancer by targeting Akt. PLoS One. 2014;9:e115980. doi: 10.1371/journal.pone.0115980. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]