

Abstract
Members of the anaerobic gut fungi (AGF) reside in rumen, hindgut, and feces of ruminant and non-ruminant herbivorous mammals and reptilian herbivores. No protocols for gene insertion, deletion, silencing, or mutation are currently available for the AGF, rendering gene-targeted molecular biological manipulations unfeasible. Here, we developed and optimized an RNA interference (RNAi)-based protocol for targeted gene silencing in the anaerobic gut fungus Pecoramyces ruminantium strain C1A. Analysis of the C1A genome identified genes encoding enzymes required for RNA silencing in fungi (Dicer, Argonaute, Neurospora crassa QDE-3 homolog DNA helicase, Argonaute-interacting protein, and Neurospora crassa QIP homolog exonuclease); and the competency of C1A germinating spores for RNA uptake was confirmed using fluorescently labeled small interfering RNAs (siRNA). Addition of chemically-synthesized siRNAs targeting D-lactate dehydrogenase (ldhD) gene to C1A germinating spores resulted in marked target gene silencing; as evident by significantly lower ldhD transcriptional levels, a marked reduction in the D-LDH specific enzymatic activity in intracellular protein extracts, and a reduction in D-lactate levels accumulating in the culture supernatant. Comparative transcriptomic analysis of untreated versus siRNA-treated cultures identified a few off-target siRNA-mediated gene silencing effects. As well, significant differential up-regulation of the gene encoding NAD-dependent 2-hydroxyacid dehydrogenase (Pfam00389) in siRNA-treated C1A cultures was observed, which could possibly compensate for loss of D-LDH as an electron sink mechanism in C1A. The results demonstrate the feasibility of RNAi in anaerobic fungi, and opens the door for gene silencing-based studies in this fungal clade.
Keywords: RNA interference, Pecoramyces ruminantium, D-lactate dehydrogenase, Anaerobic gut fungi
Introduction
The role played by non-coding RNA (ncRNA) molecules in epigenetic modulation of gene expression at the transcriptional and post-transcriptional levels is now well recognized (Catalanotto, Cogoni & Zardo, 2016). Small interfering RNAs (siRNA) are short (20–24 nt) double stranded RNA molecules that mediate post-transcriptional regulation of gene expression and gene silencing by binding to mRNA in a sequence-specific manner (Quoc & Nakayashiki, 2015). The process of RNA interference (RNAi) has been independently documented in fungi (Chang, Zhang & Liu, 2012; Cogoni & Macino, 1997b; Romano & Macino, 1992), animals and human cell lines (Atayde, Tschudi & Ullu, 2011; Chiu & Rana, 2002), as well as plants (Fang & Qi, 2016). The fungal RNAi machinery has been investigated in several model fungi, e.g., Neurospora crassa (Romano & Macino, 1992), Mucor circinelloides (Nicolás, Torres-Martínez & Ruiz-Vázquez, 2003), and Magnaporthe oryzae (Kadotani et al., 2003; Kadotani et al., 2004), and encompasses: (1) Dicer (Dic) enzyme(s): RNaseIII dsRNA-specific ribonucleases that cleave double stranded RNA (dsRNA) to short (20–25 bp) double stranded siRNA entities; (2) Argonaute (Ago) protein(s), the core component of the RNA-induced silencing complex (RISC) which binds to the dicer-generated siRNAs and other proteins and cleaves the target mRNA; (3) RNA-dependent RNA polymerase (RdRP) enzyme (present in the majority, but not all fungi) that aids in amplifying the silencing signal through the production of secondary double stranded siRNA molecules from single stranded mRNAs generated by the RISC complex; (4) DNA helicase, Neurospora crassa QDE-3 homolog (Pickford et al., 2002), that aids in the production of the aberrant RNA to be targeted by RdRP; and (5) Argonaute-interacting protein, Neurospora crassa QIP homolog (Maiti, Lee & Liu, 2007), an exonuclease that cleaves and removes the passenger strand from the siRNA duplex.
The phenomenon of RNA interference could induce gene silencing due to the action of endogenously produced microRNA (miRNA), or could be triggered due to the introduction of foreign siRNA (e.g., due to viral infection or genetic manipulation). Under normal physiological conditions, RNAi is thought to play a role in endogenous regulation of gene expression (Bartel, 2004), development of resistance to viruses (Hammond et al., 2008a; Segers et al., 2007; Sun, Choi & Nuss, 2009; Zhang et al., 2008), and silencing the expression of transposons (Murata et al., 2007; Nolan et al., 2005). On the other hand, the introduction of foreign siRNA could be utilized for targeted, sequence-specific, gene knockdown in fungi (Quoc & Nakayashiki, 2015; Chang, Zhang & Liu, 2012; Romano & Macino, 1992). Indeed, demonstration of the feasibility of RNAi approaches for targeted gene silencing has been shown in Ascomycota (Romano & Macino, 1992; Abdel-Hadi et al., 2011; Barnes, Alcocer & Archer, 2008; Eslami et al., 2014; Jöchl et al., 2009; Kalleda, Naorem & Manchikatla, 2013; Li et al., 2012; Moazeni et al., 2012; Moazeni et al., 2014; Mousavi et al., 2015; Penn et al., 2015; Prakash, Manjrekar & Chattoo, 2016), Basidiomycota (Caribé dos Santos et al., 2009; Matityahu et al., 2008; Nakade et al., 2011; Namekawa et al., 2005; Skowyra & Doering, 2012), and Mucoromycota (Gheinani et al., 2011; Nicolas et al., 2008); and RNAi-based protocols were used to infer the putative roles of several genes or simply as a proof of principle.
The anaerobic gut fungi (AGF) represent a basal fungal phylum (Neocallimastigomycota) that resides in the herbivorous gut and plays an important role in enhancing plant biomass metabolism by the host animals (Gruninger et al., 2014). The AGF have multiple potential biotechnological applications such as a source of lignocellulolytic enzymes (Cheng et al., 2014; Kwon et al., 2016; Lee et al., 2015; Morrison, Elshahed & Youssef, 2016; Wang, Chen & Hseu, 2014; Wei et al., 2016a; Wei et al., 2016b), direct utilization of AGF strains for sugar extraction from plant biomass in enzyme-free biofuel production schemes (Ranganathan et al., 2017), additives to biogas production reactors (Nkemka et al., 2015; Procházka et al., 2012), and feed additives for livestock (Dey et al., 2004; Lee, Ha & Cheng, 2000; Paul et al., 2011; Paul et al., 2004; Saxena et al., 2010; Sehgal et al., 2008; Tripathi et al., 2007). However, the strict anaerobic nature of AGF renders genetic manipulation procedures involving plating and colony selection extremely cumbersome. Consequently, there are currently no protocols for transformation, gene insertion, gene deletion, or sequence-specific homologous recombination-based genetic manipulation in AGF, hindering in-depth investigation of their biotechnological potential.
We here report on the development of an RNAi-based protocol for targeted gene knockdown in the anaerobic gut fungal isolate Pecoramyces ruminantium strain C1A. The protocol does not involve transformation, and does not require homologous recombination, or colony selection. We demonstrate the uptake of chemically synthesized short double stranded siRNA by germinating spores of P. ruminantium strain C1A, and subsequently demonstrate the feasibility of using this approach for silencing D-lactate dehydrogenase (ldhD) gene. We finally examine the off-target effects of ldhD gene knockdown, as well as the impact of inhibiting D-lactate production on the glycolytic and fermentation pathways in C1A.
Materials and Methods
Microorganism and culture maintenance
Pecoramyces ruminantium strain C1A was isolated previously in our laboratory (Hanafy et al., 2017) and maintained by biweekly transfers into an antibiotic-supplemented rumen-fluid-cellobiose medium (RFC) as described previously (Calkins et al., 2016).
Identification and phylogeny of RNAi complex in anaerobic fungi
The occurrence of genes encoding Dic, Ago, RdRP, QIP, and QDE3 proteins was examined in the genome of P. ruminantium C1A (Youssef et al., 2013) (Genbank accession number ASRE00000000.1), as well as in three additional publicly available Neocallimastigomycota genomes (Solomon et al., 2016) (Genbank accession numbers: MCOG00000000.1, MCFG00000000.1, MCFH00000000.1). The phylogeny of the translated amino acid sequences of identified homologues was compared to fungal and eukaryotic homologues in MEGA7. Representative sequences were aligned using ClustalW and the aligned sequences were manually refined and used to construct Neighbor Joining trees in Mega7 (Kumar, Stecher & Tamura, 2016) with bootstrap values calculated based on 100 replicates.
RNAi experimental design
Choice of delivery procedure
Delivery of the inhibitory RNA molecules to fungal cultures is commonly achieved using appropriate vectors that either express short hairpin RNA (Hammond et al., 2008b; Hammond & Keller, 2005; Nakayashiki et al., 2005), or individual sense and antisense RNA strands that will subsequently be annealed into dsRNA (Cogoni & Macino, 1997b; Patel et al., 2008). The process involves transformation (PEG-CaCl2-mediated into protoplasts, Li acetate-mediated, Agrobacterium-mediated, or via electroporation) and necessitates transformants’ selection on marker (usually hygromycin) plates. Alternatively, direct delivery of exogenous, chemically synthesized short double stranded RNA (siRNA) has also been utilized for targeted gene silencing in fungi (Eslami et al., 2014; Jöchl et al., 2009; Kalleda, Naorem & Manchikatla, 2013; Mousavi et al., 2015; Khatri & Rajam, 2007). This approach exploits the machinery for nucleic acids uptake, and the natural competence of the germinating spore stage observed in the filamentous fungus Aspergillus (Jöchl et al., 2009). Due to the strict anaerobic nature of AGF which would hinder the process of transformation and selection on plates, we opted for direct addition of chemically synthesized siRNA to C1A germinating spores, in spite of its reported lower efficacy (Kalleda, Naorem & Manchikatla, 2013).
dsRNA synthesis
We targeted D-lactate dehydrogenase (ldhD) gene encoding D-LDH enzyme (EC 1.1.1.28). D-LDH is an NAD-dependent oxidoreductase that reduces pyruvate to D-lactate, a major fermentation end product in C1A (Ranganathan et al., 2017). Only a single copy of ldhD (996 bp in length) was identified in C1A genome (IMG accession number 2511055262 within the C1A genome: https://img.jgi.doe.gov/cgi-bin/m/main.cgi?section=TaxonDetailpage=taxonDetailtaxon_oid=2510917007). A 21-mer siRNA targeting positions 279-298 in the ldhD gene transcript (henceforth ldhD-siRNA) was designed using Dharmacon® siDesign center (http://dharmacon.gelifesciences.com/design-center/) with the sense strand being 5′-CGUUAGAGUUCCAGCCUAUUU-3′, and the antisense strand being 5′-AUAGGCUGGAACUCUAACGUU-3′. Included within the designed siRNAs were 3′ overhanging UU dinucleotides to increase the efficiency of target RNA degradation as suggested before (Elbashir et al., 2001). The siRNA was ordered from Dharmacon® (LaFayette, CO, USA) as 21-mer duplex (double stranded) with a central 19-bp duplex region and symmetric UU dinucleotide 3′ overhangs on each end. The 5′ end of the antisense strand was modified with a phosphate group required for siRNA activity (Chiu & Rana, 2003), while the 5′ end of the sense strand was modified with a Cy-3 fluorescent dye to facilitate visualization of the siRNA uptake by C1A germinating spores. In addition, a 21-mer duplex that should not anneal to any of C1A’s mRNA transcripts (henceforth unrelated-siRNA) was also designed and used as a negative control with the sense strand being 5′-UCGUUGGCGUGAGCUUCCAUU-3′, and the antisense strand being 5′-UGGAAGCUCACGCCAACGAUU-3′. The unrelated-siRNA was modified in the same way as the ldhD siRNA.
RNAi protocol
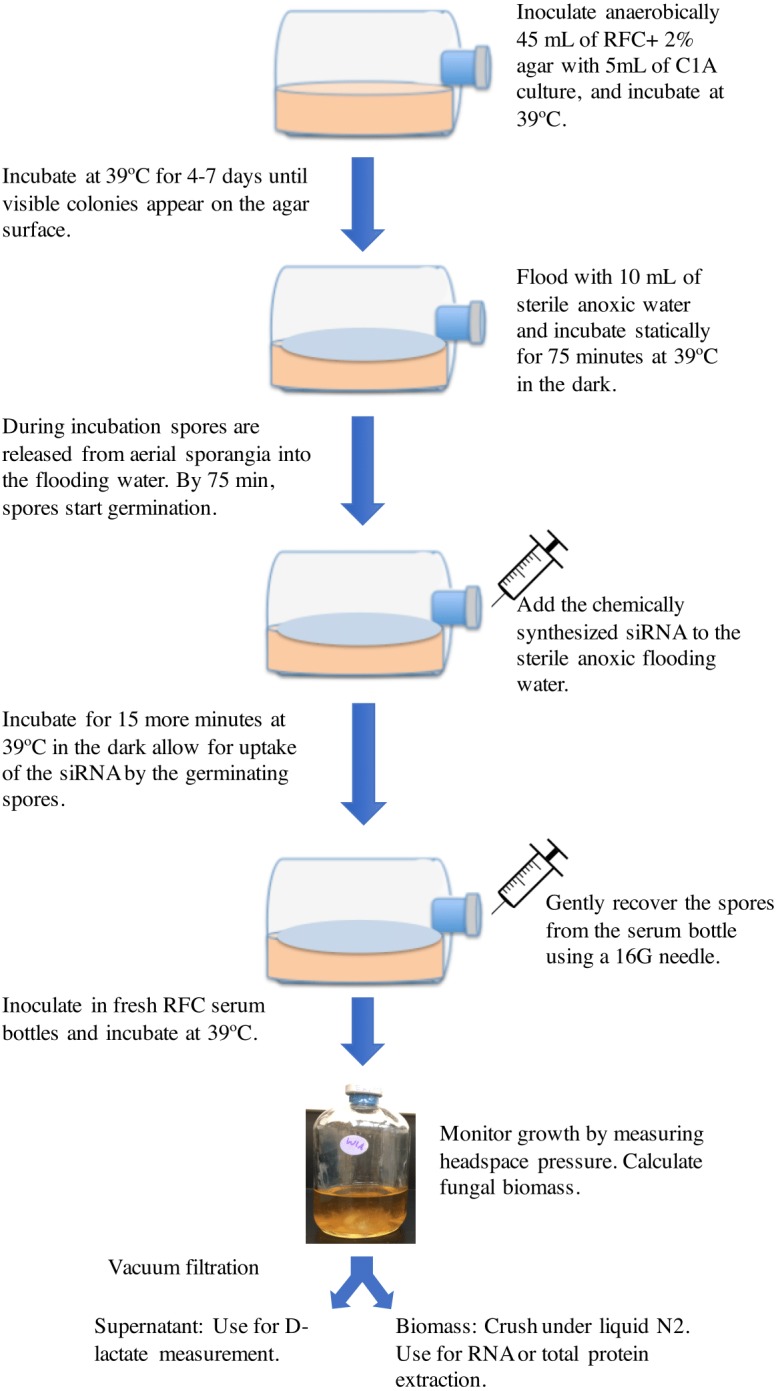
The basic protocol employed is shown in Fig. 1. Strain C1A was grown on RFC-agar medium in serum bottles at 39 °C in the dark as described previously (Calkins et al., 2016) until visible surface colonies are observed (usually 4–7 days). Surface growth was then flooded by adding 10 ml sterile anoxic water followed by incubation at 39 °C (Calkins et al., 2016). During this incubation period, spores are released from surface sporangia into the anoxic water. Previous work has shown that the duration of incubation with the flooding solution has a major impact on the spore developmental stage, where exclusively active flagellated spores were observed in incubations shorter than 30 min, while 90–100-minute incubation exclusively produced germinating spores. The onset of spore germination was observed at 75–80 min during incubation with the flooding solution (Calkins et al., 2016). Germinating spores were previously shown to be most amenable for accumulating the highest amount of exogenously added nucleic acids (Jöchl et al., 2009). We, therefore, reasoned that addition of chemically synthesized siRNA to the sterile anoxic flooding water at the onset of spore germination (at around 75 min from the onset of flooding) followed by re-incubation at 39 °C for 15 additional minutes (for a total of 90-minute incubation period) would allow for uptake of the siRNA by the germinating spores. Chemically synthesized siRNA was added from a stock solution constituted in a sterile anoxic RNase-free siRNA buffer (60 mM KCl, 6 mM HEPES-pH 7.5, and 0.2 mM MgCl2) to the desired final concentration. Initial experiments were conducted using Cy3-labeled ldhD-siRNA molecules to test the uptake of siRNA by the germinating spores. Subsequent experiments were conducted using unlabeled siRNA. Following siRNA addition and incubation, spores were gently recovered from the serum bottle using a 16G needle and used to inoculate fresh RFC media bottles (CalFkins et al., 2016), and the impact of silencing ldhD gene on gene expression, enzyme activities, and D-lactate concentrations was assessed in these cultures. Controls included treatments with unrelated-siRNA, as well as cultures with no siRNA addition.
Figure 1. A cartoon depicting the RNAi gene knockdown protocol used in this study.
Impact of ldhD gene knockdown on ldhD transcriptional levels, D- LDH enzyme activity, and D-lactate production in strain C1A
The supernatant of both siRNA-treated and control C1A cultures was periodically sampled (0.5 ml) and used for D-lactate quantification. The amount of fungal biomass at the time of quantification was derived from the headspace pressure as previously described (Ranganathan et al., 2017). The fungal biomass was vacuum filtered on 0.45 µm filters, and immediately crushed in a bath of liquid nitrogen using a mortar and pestle as described previously (Calkins & Youssef, 2016). The crushed cells were then poured into 2 separate 15-mL plastic falcon tubes, and stored at −80 °C for subsequent RNA, and protein extraction, respectively.
D-Lactate quantification
D-lactate was determined in the culture supernatant using the D-Lactate Assay Kit (BioAssay Systems, Hayward, CA, USA) following the manufacturer’s instructions.
RNA extraction, qRT-PCR, and RNA-seq
RNA was extracted following the protocol in Epicentre® MasterPure™ Yeast RNA Purification Kit, with few modifications as detailed previously (Calkins & Youssef, 2016). RNA concentrations were measured using the Qubit® RNA HS Assay Kit (Life Technologies®, Carlsbad, CA, USA). Total RNA was utilized for both transcriptional studies using qRT-PCR, as well as for transcriptomic analysis using RNA-seq.
For transcriptional studies, replicate samples were chosen to cover a range of fungal biomass ranging from 6–22 mgs corresponding to various growth stages. Reverse transcription (cDNA synthesis) was performed using the Superscript IV First-Strand Synthesis System kit for RT-PCR (Life Technologies®, Carlsbad, CA, USA), following the manufacturer’s protocols. Quantitative reverse transcription PCR (qRT-PCR) was conducted on a MyIQ thermocycler (Bio-Rad Laboratories, Hercules, CA, USA). ldhD, as well as the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH), were amplified using primers designed by the OligoPerfect™ Designer tool (Life Technologies, Carlsbad, CA, USA) (ldhD-forward primer: AGACCATGGGTGTCATTGGT, ldhD-reverse primer TTCATCGGTTAATGGGCAGT; GAPDH-forward primer: ATTCCACTCACGGACGTTTC, GAPDH-reverse primer: CTTCTTGGCACCACCCTTTA). The reactions contained 1 µl of C1A cDNA, and 0.5 µM each of the forward and reverse primers. Reactions were heated at 50 °C for 2 min, followed by heating at 95 °C for 8.5 min. This was followed by 50 cycles, with one cycle consisting of 15 s at 95 °C, 60 s at 50 °C, and 30 s at 72 °C. Using the ΔCt method, the number of copies of ldhD is reported relative to the number of copies of GAPDH used as the normalizing control.
Transcriptomic analysis was used both to evaluate off-target effects of the chemically synthesized ldhD siRNA (transcripts that will be down-regulated in siRNA-treated versus untreated cultures), and to examine the effect of ldhD knockdown on other NADH-oxidizing mechanisms to compensate for loss of D-LDH as an electron sink in C1A (transcripts that will be up-regulated in siRNA-treated versus untreated cultures). For transcriptomic analysis, RNA from untreated (Quoc & Nakayashiki, 2015 biological replicates) as well as siRNA-treated (two biological replicates) cultures was sequenced using Illumina-HiSeq. RNA sequencing as well as sequence processing were as described previously (Couger et al., 2015). Briefly, de novo assembly of the generated RNA-Seq reads was accomplished using Trinity (Haas et al., 2013), and quantitative levels of assembled transcripts were obtained using Bowtie2 (Langmead & Salzberg, 2012). Quantitative values in Fragments Per Kilobase of transcripts per Million mapped reads (FPKM) were calculated in RSEM. edgeR (Robinson, McCarthy & Smyth, 2010) was used to determine the transcripts that were significantly up- or down-regulated based on the Benjamini–Hochberg adjusted p-value (False discovery rate, FDR). We used a threshold of 10% FDR as the cutoff for determining significantly differentially expressed transcripts.
Total protein extraction and D-Lactate dehydrogenase enzyme assay
For total protein extraction, replicate samples were chosen to cover a range of fungal biomass ranging from 6–22 mgs corresponding to various growth stages. C1A cells crushed in liquid nitrogen were suspended in 0.5 mL of Tris-Gly buffer (3 g Tris base, 14.4 g Glycine, H2O up to 1L, pH 8.3), and mixed briefly. Cell debris were pelleted by centrifugation (12,500× g for 2 min at 4 °C) and the sample supernatant containing the total protein extract was carefully transferred into a sterile microfuge tube. Protein concentrations were quantified in cellular extracts using Qubit™ Protein assay kit (Life Technologies, Carlsbad, CA, USA). D-LDH enzyme activity was quantified in the cell extracts using the Amplite™ Colorimetric D-Lactate Dehydrogenase Assay Kit (ATT Bioquest®, Sunnyvale, CA, USA), following the manufacturer’s protocols.
Statistical analysis
To evaluate the effect of the siRNA treatment on the transcriptional level of ldhD relative to the housekeeping gene gapdh, the D-LDH specific activity, as well as the total amount of D-lactate in the culture supernatant, Student t-tests were conducted to test for the significance of difference between untreated cultures and ldhD-siRNA treated cultures, and p-values were compared. To evaluate the effect of the concentration of ldhD-siRNA exogenously added to C1A cultures on the level of inhibition of ldhD (at the RNA (the transcriptional level of ldhD relative to the housekeeping gene gapdh), protein (D-LDH specific activity), and metabolite (the total amount of D-lactate in the culture supernatant) levels), Student t-tests were conducted to test for the significance of difference between samples treated with different concentrations of ldhD-siRNA, and p-values were compared.
Nucleotide accession
This Transcriptome Shotgun Assembly project has been deposited at DDBJ/EMBL/GenBank under the accession GFSU00000000. The version described in this paper is the first version, GFSU01000000.
Results
RNAi machinery in the Neocallimastigomycota
The four examined Neocallimastigomycota genomes harbored most of the genes constituting the backbone of the RNAi machinery: ribonuclease III dicer, argonaute, QDE3-homolog DNA helicase, and QIP-homolog exonuclease. Phylogenetically, these genes were closely related to representatives from basal fungal lineages (Figs. 2–4). Gene copies in various genomes ranged between 1 to 4 (Figs. 2–4). However, it is notable that all four examined genomes lacked a clear homolog of RNA-dependent RNA polymerase (RdRP) gene. RdRP has been identified in the genomes of diverse organisms including Caenorhabditis elegans (Smardon et al., 2000), plants, and the majority of examined fungi (Cogoni & Macino, 1997a) but is absent in the genomes of vertebrates and flies; in spite of their possession of a robust RNAi machinery that mediates sequence-specific gene silencing in response to exogenously added dsRNAs.
Figure 2. Neighbor joining phylogenetic tree.

Neighbor joining phylogenetic tree depicting the phylogenetic relationship between Pecoramyces ruminantium strain C1A predicted Dicer sequences and those from other fungal and eubaryotic species. Trees were constructed in Mega7 with bootstrap support based on 100 replicates. Bootstrap values are shown for branches with >50 bootstrap support.
Figure 4. Neighbor joining phylogenetic tree.

Neighbor joining phylogenetic tree depicting the phylogenetic relationship between Pecoramyces ruminantium strain C1A predicted QDE-3 helicase (A) and QIP exonuclease (B) sequences and those from other fungal and eukaryotic species. Trees were constructed in Mega7 with bootstrap support based on 100 replicates. Bootstrap values are shown for branches with >50 bootstrap support.
Figure 3. Neighbor joining phylogenetic tree.

Neighbor joining phylogenetic tree depicting the phylogenetic relationship between Pecoramyces ruminantium strain C1A predicted Argonaute sequences and those from other fungal and eukaryotic species. Trees were constructed in Mega7 with bootstrap support based on 100 replicates. Bootstrap values are shown for branches with >50 bootstrap support.
Uptake of synthetic siRNA by C1A germinating spores and effect on growth
The addition of fluorescently labeled siRNA targeting ldhD transcript to C1A spores at the onset of germination followed by a 15-minute incubation at 39 °C resulted in the uptake of the siRNA by the germinating spores as evident by their fluorescence (Fig. S1A). Several fields of vision were examined and the number of spores with Cy3-fluoresence, indicative of siRNA uptake, as a percentage of the total number of spores (stained with the nuclear stain DAPI) was evaluated. Under the examined conditions, the majority of the germinating spores picked up the siRNA since 80–90% of spores stained with the nuclear stain DAPI also exhibited Cy3-fluoresence (results from at least four separate experiments). ldhD-siRNA-treated spores were collected and used to inoculate fresh RFC liquid media, and the growth rate of these cultures were compared to siRNA-untreated controls. As shown in Fig. S1B, ldhD-siRNA treatment had no significant effect on either the rate of fungal growth or the final fungal biomass yield.
Knockdown of ldhD-gene by exogenously added ldhD-siRNA
Inhibition at the mRNA level
Table 1 shows the effect of adding exogenous ldhD-siRNA on ldhD transcriptional level relative to the housekeeping gene glyceraldehyde-3-phosphate dehydrogenase. Results from qRT-PCR revealed that there was an observable decrease in ldhD transcription levels in samples treated with ldhD-specific siRNA compared to siRNA-untreated samples or unrelated siRNA-treated samples. This effect was significant (Student t-test p-values ≤ 0.0012). The inhibitory effect increased with the concentration of ldhD-specific siRNA added, and this observed increase in inhibitory effect was significant at siRNA concentrations higher than 50 nM (Student t-test p-values <0.05). The highest level of inhibition was obtained when 100 nM of the ldhD-siRNA was exogenously added to C1A germinating spores, where a four-fold decrease in transcription was observed. C1A cultures treated with the unrelated siRNA showed no significant difference in the transcriptional level of ldhD when compared to siRNA-untreated cultures.
Table 1. Effect of the uptake of exogenous ldhD-siRNA by C1A germinating spores on the transcriptional level of ldhD relative to the housekeeping gene gapdh.
| Treatment | Final siRNA conc. (nM) | Copies of ldhD relative to gapdh1,2 | Fold change in transcription level (ΔΔCt) compared to untreated samples | Number of biological replicates | Fungal biomass yield (mg) at the time of sacrificing1 |
|---|---|---|---|---|---|
| ldhD-siRNA | 20 | 4.2E−03 ± 3E−03∗∗∗,a | 0.02 | 4 | 12.3 ± 5 |
| 50 | 4.4E−03 ± 2E−03∗∗∗,b | 0.02 | 5 | 9.3 ± 5.2 | |
| 75 | 3.6E−04 ± 1.8E−04∗∗∗,ab | 0.0017 | 4 | 15.4 ± 3.7 | |
| 100 | 6.1E−05 ± 2.4E−05∗∗∗,ab | 0.0003 | 4 | 15.9 ± 6 | |
| Untreated | NA | 0.21 ± 0.04 | 1 | 5 | 9.6 ± 2 |
| Unrelated-siRNA | 50 | 0.26 ± 0.07NS | 1.29 | 2 | 13.5 ± 3.8 |
Notes.
Values are average ± standard deviation. Student t-test was used to test the significance of the difference of the siRNA-treated samples averages from that of the untreated sample. All ldhD-siRNA treated samples showed a significant decrease in the transcriptional level of ldhD relative to the housekeeping gene gapdh compared to the untreated control. P-value of the significant difference; ***, p-value <0.00002; **, p-value = 0.0012; NS, not significant.
a,b When comparing the transcriptional level of ldhD relative to the housekeeping gene gapdh in samples treated with different concentration of ldhD-siRNA, a significant difference was observed with concentrations higher than 50 nM (p-value <0.05). Samples with the same letter were significantly different.
Inhibition at the protein level
Similar to the effect of treatment on the mRNA level, ldhD-siRNA-treated samples exhibited a marked decrease in the specific D-LDH activity (Table 2). This effect was significant (Student t-test p-value <2 ×10−8). The decrease in D-LDH specific activity was dependent on the concentration of siRNA added and ranged from 71–84% reduction compared to siRNA-untreated samples. The highest level of inhibition was obtained when 100 nM of the ldhD-siRNA was exogenously added to C1A germinating spores, where an 84% decrease in D-LDH specific activity was observed. When comparing samples treated with different concentration of ldhD-siRNA to one another, D-LDH specific activities in samples treated with 20 nM ldhD-siRNA were not significantly different from those in samples treated with 50 nM ldhD-siRNA (p > 0.05). Similarly, D-LDH specific activities in samples treated with 75 nM ldhD-siRNA were not significantly different from those in samples treated with 100 nM ldhD-siRNA (p > 0.05). However, D-LDH specific activities in samples treated with 20 nM ldhD-siRNA were significantly different from those in samples treated with 75 nM or 100 nM ldhD-siRNA, and similarly, samples treated with 50 nM ldhD-siRNA were significantly different from those in samples treated with 75 nM or 100 nM ldhD-siRNA (p < 0.05). C1A cultures treated with the unrelated siRNA showed no significant difference in D-LDH specific activities when compared to siRNA-untreated cultures (Table 2).
Table 2. Effect of the uptake of ldhD-siRNA by C1A germinating spores on the D-LDH specific activity.
| Treatment | siRNA concentration (nM) | D-LDH specific activity (U/mg protein)1 | Fold change in D-LDH specific activity compared to untreated samples | Total number of biological replicates | Fungal biomass yield (mg) at the time of sacrificing1 |
|---|---|---|---|---|---|
| ldhD-siRNA | 20 | 332.2 ± 90∗∗∗ | 0.29 | 6 | 16.5 ± 5.8 |
| 50 | 331.9 ± 144.5∗∗∗ | 0.29 | 17 | 10 ± 4.3 | |
| 75 | 194.2 ± 79∗∗∗ | 0.17 | 6 | 12.8 ± 5.3 | |
| 100 | 180.6 ± 131∗∗∗ | 0.16 | 6 | 12.7 ± 7.4 | |
| Untreated | NA | 1157.6 ± 308.6 | 1 | 13 | 10.9 ± 2.9 |
| Unrelated-siRNA | 50 | 926.4 ± 69NS | 0.8 | 2 | 13.5 ± 3.8 |
Notes.
Values are average ± standard deviation. Student t-test was used to test the significance of the difference of the siRNA-treated samples averages from that of the untreated sample. All ldhD-siRNA treated samples showed a significant decrease in the D-LDH specific activity compared to the untreated control. P-value of the significant difference; ***, p-value <2 ×10−8; NS, not significant. When comparing samples treated with different concentration of ldhD-siRNA to one another, D-LDH specific activities in samples treated with 20 nM ldhD-siRNA were not significantly different from those in samples treated with 50 nM ldhD-siRNA (p > 0.05). Similarly, D-LDH specific activities in samples treated with 75 nM ldhD-siRNA were not significantly different from those in samples treated with 100 nM ldhD-siRNA (p > 0.05). However, D-LDH specific activities in samples treated with 20 nM ldhD-siRNA were significantly different from those in samples treated with 75 nM or 100 nM ldhD-siRNA, and similarly, samples treated with 50 nM ldhD-siRNA were significantly different from those in samples treated with 75 nM or 100 nM ldhD-siRNA (p < 0.05).
Effect of ldhD gene knockdown on the extracellular levels of D-lactate in culture supernatants
D-lactate production in C1A culture supernatant is non-linear, with higher amounts of D-lactate produced at later stages of growth (Fig. 5A). D-lactate production in ldhD-siRNA-treated cultures was invariably significantly lower when compared to controls (Student t-test p-value <0.05) (Fig. 5B), with the difference especially pronounced at later stages of growth. The level of reduction was dependent on the siRNA concentration added and ranged from 42–86% in the early log phase, 49–67% in the mid log phase, and 57–86% in the late log-early stationary growth phase (Fig. 5B, and Table S1).
Figure 5. D-lactate in culture supernatant.

(A) Pattern of D-lactate production in C1A culture supernatant as a factor of fungal biomass. The majority of the D-lactate production occurs at the late log-early stationary phase. Data is shown for both siRNA-untreated cultures (green), as well as ldhD-specific siRNA-treated cultures with final concentration 20 nM (dark blue), 50 nM (orange), 75 nM (grey), and 100 nM (yellow). (B) A bar-chart depicting average ± standard deviation (from at least two replicates) of D-lactate levels in C1A culture supernatant during early log (6–13 mg biomass), mid-log (14–17 mg biomass), and late log/early stationary (18–23 mg) phases. Data is shown for both siRNA-untreated cultures (green), as well as ldhD-specific siRNA-treated cultures with final concentration 20 nM (dark blue), 50 nM (orange), 75 nM (grey), and 100 nM (yellow).
Transcriptomic analysis
Differential gene expression patterns between ldhD-siRNA-treated and siRNA-untreated samples were analyzed to identify possible off-target effects of siRNA treatment, i.e., transcripts that were significantly down-regulated in the siRNA-treated cultures. Only 29 transcripts were significantly (FDR <0.1) down-regulated (Fig. 6). Predicted functions of these transcripts are shown in Table S2 and included hypothetical proteins (n = 11), several glycosyl hydrolases (n = 5), and other non-fermentation related functions. Comparison of the siRNA sequence to these 29 transcripts revealed matches to the first seven bases of the ldhD-siRNA sequence to only three of the down-regulated transcripts indicating that the off-target effect was mainly not sequence-specific.
Figure 6. Volcano plot of the distribution of gene expression for C1A cultures when treated with ldhD-specific siRNA (50 nM) versus untreated cultures.

The fold change (log2 (average FPKM in siRNA-treated cultures/average FPKM in control cultures)) is shown on the X-axis, while the significance of the change (−log10 (false discovery rate)) is shown on the Y-axis. Red data points are those transcripts that were significantly down-regulated (n = 29), while green data points are those transcripts that were significantly up-regulated (n = 53). The corresponding IMG gene accession numbers and the predicted functions for these genes are shown in Table S1. The orange data point corresponds to the D-lactate dehydrogenase transcript (targeted in the RNAi experiment) with 2.5-fold decrease in FPKM compared to the untreated control, while the purple data point corresponds to the NAD-dependent 2-hydroxyacid dehydrogenase (Pfam 00389) transcript (possibly acting to compensate for the loss of NADH oxidation that occurred as a result of ldhD knockdown) with 1,542-fold increase in FPKM compared to the untreated control.
In an attempt to decipher the impact of inhibiting the D-lactate dehydrogenase enzyme (one of the major electron sinks in C1A) on the glycolytic and fermentation pathways in C1A, we investigated the significantly up-regulated transcripts in the siRNA-treated cultures. A total of 53 transcripts were significantly upregulated in the siRNA-treated cultures (FDR < 0.1) (Fig. 6). Predicted functions of these transcripts are shown in Table S2. One transcript encoding NAD-dependent 2-hydroxyacid dehydrogenase (Pfam 00389) was significantly upregulated (1,542-fold) in the siRNA-treated cultures (P-value = 0.02). Enzymes belonging to this family act specifically on the D-isomer of their substrates (Dengler et al., 1997). In case of D-LDH inhibition in the siRNA-treated cultures, the Pfam 00389 enzyme might act to compensate for the loss of NADH oxidation by acting on an alternate substrate (e.g., hydroxypyruvate, 2-oxoisocaproate, or other 2-oxo carboxylic acids) and reducing it as a sink of electrons to regenerate NAD. However, it is difficult to know the actual substrate based on sequence data alone. Transcripts of other glycolytic and fermentative enzymes of C1A were not differentially expressed in siRNA-treated cultures (Table S2).
Discussion
Here, we explored the feasibility of RNA interference for targeted gene silencing in the anaerobic gut fungi (phylum Neocallimastigomycota) via the exogenous addition of synthetic double stranded siRNAs targeting the ldhD gene to Pecoramyces ruminantium strain C1A germinating spores. We show that ds-siRNA was uptaken by germinating spores, and, as a consequence, the transcription of the target gene (ldhD) was down-regulated (Table 1), leading to lower D-LDH enzymatic activity (Table 2) and lower D-lactate concentration in the culture supernatant (Fig. 5).
In general, the fungal RNAi machinery encompasses Dicer (Dic) enzyme(s), Argonaute (Ago) protein(s), RNA-dependent RNA polymerase (RdRP) enzyme, QDE3-like DNA helicase, and Argonaute-interacting exonuclease (QIP-like). Genomes of Neocallimastigomycota representatives belonging to four genera (Pecoramyces, Neocallimastix, Piromyces, and Anaeromyces) encode at least one copy of Dic, Ago, QDE3-like helicase, and QIP exonuclease. However, all genomes lacked a clear homolog of RdRP. The absence of an RdRP homolog is not uncommon. While present in almost all studied fungi, RdRP seems to be missing from the genomes of other basal fungal phyla (Chytridiomycota and Blastocladiomycota) representatives (Choi et al., 2014; Farrer et al., 2017). The absence of clear RdRP homologues in the Neocallimastigomycota and related basal fungal phyla despite their presence in other fungi could suggest that either an RdRP is not involved in dsRNA-mediated mRNA silencing as shown before in mammals (Stein et al., 2003). Alternatively, RNA-dependent RNA polymerase activity could be mediated through a non-canonical RdRP in basal fungi, e.g., the RNA polymerase II core elongator complex subunit Elp1 shown to have RdRP activity in Drosophila, as well as Caenorhabditis elegans, Schizosaccharomyces pombe, and human (Birchler, 2009; Lipardi & Paterson, 2009).
We chose as a gene knockdown target the D-Lactate dehydrogenase gene (ldhD) that mediates NADH-dependent pyruvate reduction to D-lactate, for several reasons. First, the gene is present as a single copy in the genome. Second, quantification of the impact of ldhD gene knockdown is readily achievable in liquid media at the RNA (using RT-PCR and transcriptomics), and protein (using specific enzyme activity assays) levels, as well as phenotypically (by measuring D-lactate accumulation in the culture media); providing multiple lines of evidence for the efficacy of the process. Finally, D-lactate dehydrogenase is part of the complex mixed acid fermentation pathway in P. ruminantium (Ranganathan et al., 2017; Youssef et al., 2013) and other anaerobic gut fungi, and we sought to determine how blocking one route of electron disposal could lead to changes in C1A fermentation end products.
ldhD-siRNA-treated cultures showed a significant reduction in ldhD gene transcription and D-LDH enzyme activity. Both of these effects were dependent on the concentration of siRNA added (Tables 1 and 2) similar to previous reports in filamentous fungi (Eslami et al., 2014; Jöchl et al., 2009; Kalleda, Naorem & Manchikatla, 2013; Mousavi et al., 2015). We show that the addition of 100 nM of ldhD-siRNA resulted in a four-fold reduction in ldhD transcription, 84% reduction in D-LDH specific activity, and 86% reduction in D-lactate concentration in culture supernatant. The fact that targeted gene silencing using exogenously added gene-specific siRNA results in reducing rather than completely abolishing gene function is an important advantage of RNAi approaches allowing functional studies of housekeeping or survival-essential genes.
While initial studies of gene silencing using exogenously added siRNAs suggested that the process was highly sequence-specific (Elbashir et al., 2001; Tuschl et al., 1999), subsequent studies showed silencing of off-target genes based on less than perfect complementarity between the siRNA and the off-target gene (Jackson et al., 2003). Here, we used RNA-seq to quantify the off-target effects of ldhD-siRNA. In contrast to previous studies that used similar approaches to quantify RNAi off-targets (Li-Byarlay et al., 2013), we show here that the off-target effects of ldhD silencing were minimal (only 29 transcripts out of 55,167 total transcripts were differentially down-regulated as a result of siRNA treatment) and appeared to be not sequence-specific.
Currently, and due to their strict anaerobic nature, there are no established procedures for genetic manipulations (e.g., gene silencing, insertion, deletion, and mutation) of AGF leading to a paucity of molecular biological studies of the phylum. This is in stark contrast to the rich body of knowledge available on genetic manipulations of various aerobic fungal lineages (Eslami et al., 2014; Kalleda, Naorem & Manchikatla, 2013; Mousavi et al., 2015; Khatri & Rajam, 2007; Michielse et al., 2008; Minz & Sharon, 2010). Our work here represents a proof of principal of the feasibility of the RNAi approach in AGF, and opens the door for genetic manipulation and gene function studies in this important group of fungi.
Conclusions
Anaerobic gut fungi (AGF) have a restricted habitat in the herbivorous gut. Due to their anaerobic nature, gene manipulation studies are limited hindering gene-targeted molecular biological manipulations. We used an AGF representative, Pecoramyces ruminantium strain C1A, to study the feasibility of using RNA interference (RNAi) for targeted gene silencing. Using D-lactate dehydrogenase (ldhD) gene as a target, we show that RNAi is feasible in AGF as evidenced by significantly lower gene transcriptional levels, a marked reduction in encoded enzymatic activity in intracellular protein extracts, and a reduction in D-lactate levels accumulating in the culture supernatant. To our knowledge, this is the first attempt of gene manipulation studies in the AGF lineage and should open the door for gene silencing-based studies in this fungal clade.
Supplemental Information
Figure S1. Uptake of fluorescently (Cy3) tagged siRNA by C1A spores. (A) The ldhD-specific siRNA was added to the flooding solution 75 minutes after the onset of flooding followed by incubation for 15 more minutes at 39°C. Samples (a few microliters) were taken at regular intervals for visualization. The same field is shown for DAPI-, and Cy3-labeled germinating spores (Note that the spores were concurrently stained with DAPI and fluorescing green indicating the uptake of the Cy3-labeled siRNA) (bar =2000B5). (B) Effect of the siRNA treatment on fungal growth rate. siRNA-treated spores were collected and used to inoculate fresh RFC medium. Control cultures were started at the same time using siRNA-untreated spores. Headspace pressure was measured daily and used to calculate fungal biomass as described previously (1). Error bars are standard deviations from at least three replicate cultures for each condition.
Table S1. Silencing efficiency (based on the total number of mmoles of D-lactate in the culture supernatant) of ldhD-siRNA during early log (6–13 mg biomass), mid-log (14–17 mg biomass), and late log/early stationary (18–23 mg) phases a.
Table S2. Transcripts with a significant (False Discovery Rate (FDR) <0.1) fold change in the ldhD-siRNA-treated cultures.
Funding Statement
This work was supported by the National Science Foundation Grant award number 1557102. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Additional Information and Declarations
Competing Interests
The authors declare there are no competing interests.
Author Contributions
Shelby S. Calkins performed the experiments, analyzed the data, wrote the paper, prepared figures and/or tables, reviewed drafts of the paper.
Nicole C. Elledge performed the experiments, reviewed drafts of the paper.
Katherine E. Mueller performed the experiments, analyzed the data, reviewed drafts of the paper.
Stephen M. Marek reviewed drafts of the paper.
MB Couger performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, reviewed drafts of the paper.
Mostafa S. Elshahed analyzed the data, contributed reagents/materials/analysis tools, wrote the paper, prepared figures and/or tables, reviewed drafts of the paper.
Noha H. Youssef conceived and designed the experiments, analyzed the data, contributed reagents/materials/analysis tools, wrote the paper, prepared figures and/or tables, reviewed drafts of the paper.
Data Availability
The following information was supplied regarding data availability:
This Transcriptome Shotgun Assembly project has been deposited at DDBJ/EMBL/GenBank under the accession GFSU00000000. The version described in this paper is the first version, GFSU01000000.
References
- Abdel-Hadi et al. (2011).Abdel-Hadi AM, Caley DP, Carter DR, Magan N. Control of aflatoxin production of Aspergillus flavus and Aspergillus parasiticus using RNA silencing technology by targeting aflD (nor-1) gene. Toxins. 2011;3:647–659. doi: 10.3390/toxins3060647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Atayde, Tschudi & Ullu (2011).Atayde VD, Tschudi C, Ullu E. The emerging world of small silencing RNAs in protozoan parasites. Trends in Parasitology. 2011;27:321–327. doi: 10.1016/j.pt.2011.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barnes, Alcocer & Archer (2008).Barnes SE, Alcocer MJC, Archer DB. siRNA as a molecular tool for use in Aspergillus niger. Biotechnology Letters. 2008;30:885–890. doi: 10.1007/s10529-007-9614-0. [DOI] [PubMed] [Google Scholar]
- Bartel (2004).Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. doi: 10.1016/S0092-8674(04)00045-5. [DOI] [PubMed] [Google Scholar]
- Birchler (2009).Birchler JA. Ubiquitous RNA-dependent RNA polymerase and gene silencing. Genome Biology. 2009;10:243–243. doi: 10.1186/gb-2009-10-11-243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calkins et al. (2016).Calkins S, Elledge NC, Hanafy RA, Elshahed MS, Youssef N. A fast and reliable procedure for spore collection from anaerobic fungi: application for RNA uptake and long-term storage of isolates. Journal of Microbiological Methods. 2016;127:206–213. doi: 10.1016/j.mimet.2016.05.019. [DOI] [PubMed] [Google Scholar]
- Calkins & Youssef (2016).Calkins S, Youssef NH. Insights into the utility of the focal adhesion scaffolding proteins in the anaerobic fungus Orpinomyces sp. C1A. PLOS ONE. 2016;11:e0163553. doi: 10.1371/journal.pone.0163553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caribé dos Santos et al. (2009).Caribé dos Santos AC, Sena JAL, Santos SC, Dias CV, Pirovani CP, Pungartnik C, Valle RR, Cascardo JCM, Vincentz M. dsRNA-induced gene silencing in Moniliophthora perniciosa, the causal agent of witches’ broom disease of cacao. Fungal Genetics and Biology. 2009;46:825–836. doi: 10.1016/j.fgb.2009.06.012. [DOI] [PubMed] [Google Scholar]
- Catalanotto, Cogoni & Zardo (2016).Catalanotto C, Cogoni C, Zardo G. MicroRNA in control of gene expression: an overview of nuclear functions. International Journal of Molecular Sciences. 2016;17:E1712. doi: 10.3390/ijms17101712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang, Zhang & Liu (2012).Chang SS, Zhang Z, Liu Y. RNA interference pathways in fungi: mechanisms and functions. Annual Review of Microbiology. 2012;66:305–323. doi: 10.1146/annurev-micro-092611-150138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng et al. (2014).Cheng YS, Chen CC, Huang CH, Ko TP, Luo W, Huang JW, Liu JR, Guo RT. Structural analysis of a glycoside hydrolase family 11 xylanase from Neocallimastix patriciarum: insights into the molecular basis of a thermophilic enzyme. Journal of Biological Chemistry. 2014;289:11020–11028. doi: 10.1074/jbc.M114.550905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiu & Rana (2002).Chiu Y-L, Rana TM. RNAi in human cells. Molecular Cell. 2002;10:549–561. doi: 10.1016/S1097-2765(02)00652-4. [DOI] [PubMed] [Google Scholar]
- Chiu & Rana (2003).Chiu YL, Rana TM. siRNA function in RNAi: a chemical modification analysis. RNA. 2003;9:1034–1048. doi: 10.1261/rna.5103703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi et al. (2014).Choi J, Kim K-T, Jeon J, Wu J, Song H, Asiegbu FO, Lee Y-H. funRNA: a fungi-centered genomics platform for genes encoding key components of RNAi. BMC Genomics. 2014;15:S14. doi: 10.1186/1471-2164-15-S9-S14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cogoni & Macino (1997a).Cogoni C, Macino G. Conservation of transgene-induced post-transcriptional gene silencing in plants and fungi. Trends in Plant Science. 1997a;2:438–443. doi: 10.1016/S1360-1385(97)90028-5. [DOI] [Google Scholar]
- Cogoni & Macino (1997b).Cogoni C, Macino G. Isolation of quelling-defective (qde) mutants impaired in posttranscriptional transgene-induced gene silencing in Neurospora crassa. Proceedings of the National Academy of Sciences of the United States of America. 1997b;94:10233–10238. doi: 10.1073/pnas.94.19.10233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Couger et al. (2015).Couger MB, Youssef NH, Struchtemeyer CG, Liggenstoffer AS, Elshahed MS. Transcriptomic analysis of lignocellulosic biomass degradation by the anaerobic fungal isolate Orpinomyces sp. strain C1A. Biotechnology for Biofuels. 2015;8 doi: 10.1186/s13068-015-0390-0. Article 208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dengler et al. (1997).Dengler U, Niefind K, Kieß M, Schomburg D. Crystal structure of a ternary complex of d-2-hydroxyisocaproate dehydrogenase from Lactobacillus casei, NAD+ and 2-oxoisocaproate at 1.9 Å resolution. Journal of Molecular Biology. 1997;267:640–660. doi: 10.1006/jmbi.1996.0864. [DOI] [PubMed] [Google Scholar]
- Dey et al. (2004).Dey A, Sehgal JP, Puniya AK, Singh K. Influence of an anaerobic fungal culture (Orpinomyces sp.) administration on growth rate, ruminal fermentation and nutrient digestion in calves. Asian-Australasian Journal of Animal Sciences. 2004;17:820–824. doi: 10.5713/ajas.2004.820. [DOI] [Google Scholar]
- Elbashir et al. (2001).Elbashir SM, Martinez J, Patkaniowska A, Lendeckel W, Tuschl T. Functional anatomy of siRNAs for mediating efficient RNAi in Drosophila melanogaster embryo lysate. EMBO Journal. 2001;20:6877–6888. doi: 10.1093/emboj/20.23.6877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eslami et al. (2014).Eslami H, Khorramizadeh MR, Pourmand MR, Moazeni M, Rezaie S. Down-regulation of sidB gene by use of RNA interference in Aspergillus nidulans. Iranian Biomedical Journal. 2014;18:55–59. doi: 10.6091/ibj.1217.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fang & Qi (2016).Fang X, Qi Y. RNAi in plants: an argonaute-centered view. The Plant Cell. 2016;28:272–285. doi: 10.1105/tpc.15.00920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farrer et al. (2017).Farrer RA, Martel A, Verbrugghe E, Abouelleil A, Ducatelle R, Longcore JE, James TY, Pasmans F, Fisher MC, Cuomo CA. Genomic innovations linked to infection strategies across emerging pathogenic chytrid fungi. Nature Communications. 2017;8 doi: 10.1038/ncomms14742. Article 14742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gheinani et al. (2011).Gheinani AH, Jahromi NH, Feuk-Lagerstedt E, Taherzadeh MJ. RNA silencing of lactate dehydrogenase gene in Rhizopus oryzae. Journal of RNAi and Gene Silencing. 2011;7:443–448. [PMC free article] [PubMed] [Google Scholar]
- Gruninger et al. (2014).Gruninger RJ, Puniya AK, Callaghan TM, Edwards JE, Youssef N, Dagar SS, Fliegerova K, Griffith GW, Forster R, Tsang A, McAllister T, Elshahed MS. Anaerobic fungi (phylum Neocallimastigomycota): advances in understanding their taxonomy, life cycle, ecology, role and biotechnological potential. FEMS Microbiology Ecology. 2014;90:1–17. doi: 10.1111/1574-6941.12383. [DOI] [PubMed] [Google Scholar]
- Haas et al. (2013).Haas BJ, Papanicolaou A, Yassour M, Grabherr M, Blood PD, Bowden J, Couger MB, Eccles D, Li B, Lieber M, MacManes MD, Ott M, Orvis J, Pochet N, Strozzi F, Weeks N, Westerman R, William T, Dewey CN, Henschel R, LeDuc RD, Friedman N, Regev A. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nature Protocols. 2013;8:1494–1512. doi: 10.1038/nprot.2013.084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammond et al. (2008a).Hammond TM, Andrewski MD, Roossinck MJ, Keller NP. Aspergillus mycoviruses are targets and suppressors of RNA silencing. Eukaryotic Cell. 2008a;7:350–357. doi: 10.1128/EC.00356-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammond et al. (2008b).Hammond TM, Bok JW, Andrewski MD, Reyes-Dominguez Y, Scazzocchio C, Keller NP. RNA silencing gene truncation in the filamentous fungus Aspergillus nidulans. Eukaryotic Cell. 2008b;7:339–349. doi: 10.1128/EC.00355-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammond & Keller (2005).Hammond TM, Keller NP. RNA silencing in Aspergillus nidulans is independent of RNA-dependent RNA polymerases. Genetics. 2005;169:607–617. doi: 10.1534/genetics.104.035964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanafy et al. (2017).Hanafy RA, Elshahed MS, Liggenstoffer AS, Griffith GW, Youssef NH. Pecoramyces ruminantium, gen. nov, sp. nov, an anaerobic gut fungus from the feces of cattle and sheep. Mycologia. 2017;109:231–243. doi: 10.1080/00275514.2017.1317190. [DOI] [PubMed] [Google Scholar]
- Jackson et al. (2003).Jackson AL, Bartz SR, Schelter J, Kobayashi SV, Burchard J, Mao M, Li B, Cavet G, Linsley PS. Expression profiling reveals off-target gene regulation by RNAi. Nature Biotechnology. 2003;21:635–637. doi: 10.1038/nbt831. [DOI] [PubMed] [Google Scholar]
- Jöchl et al. (2009).Jöchl C, Loh E, Ploner A, Haas H, Hüttenhofer A. Development-dependent scavenging of nucleic acids in the filamentous fungus Aspergillus fumigatus. RNA Biology. 2009;6:179–186. doi: 10.4161/rna.6.2.7717. [DOI] [PubMed] [Google Scholar]
- Kadotani et al. (2003).Kadotani N, Nakayashiki H, Tosa Y, Mayama S. RNA silencing in the phytopathogenic fungus Magnaporthe oryzae. Molecular Plant-Microbe Interactions. 2003;16:769–776. doi: 10.1094/MPMI.2003.16.9.769. [DOI] [PubMed] [Google Scholar]
- Kadotani et al. (2004).Kadotani N, Nakayashiki H, Tosa Y, Mayama S. One of the two Dicer-like proteins in the filamentous fungi Magnaporthe oryzae genome is responsible for hairpin RNA-triggered RNA silencing and related small interfering RNA accumulation. Journal of Biological Chemistry. 2004;279:44467–44474. doi: 10.1074/jbc.M408259200. [DOI] [PubMed] [Google Scholar]
- Kalleda, Naorem & Manchikatla (2013).Kalleda N, Naorem A, Manchikatla RV. Targeting fungal genes by diced siRNAs: a rapid tool to decipher gene function in Aspergillus nidulans. PLOS ONE. 2013;8:e75443. doi: 10.1371/journal.pone.0075443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khatri & Rajam (2007).Khatri M, Rajam MV. Targeting polyamines of Aspergillus nidulans by siRNA specific to fungal ornithine decarboxylase gene. Medical Mycology. 2007;45:211–220. doi: 10.1080/13693780601158779. [DOI] [PubMed] [Google Scholar]
- Kumar, Stecher & Tamura (2016).Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis Version 7.0 for bigger datasets. Molecular Biology and Evolution. 2016;33:1870–1874. doi: 10.1093/molbev/msw054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwon et al. (2016).Kwon M, Song J, Park HS, Park H, Chang J. Characterization of heterologously expressed acetyl xylan esterase1 isolated from the anaerobic rumen fungus Neocallimastix frontalis PMA02. Asian-Australasian Journal of Animal Sciences. 2016;29:1576–1584. doi: 10.5713/ajas.16.0336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Langmead & Salzberg (2012).Langmead B, Salzberg SL. Fast gapped-read alignment with Bowtie 2. Nature Methods. 2012;9:357–359. doi: 10.1038/nmeth.1923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee et al. (2015).Lee SM, Guan LL, Eun JS, Kim CH, Lee SJ, Kim ET, Lee SS. The effect of anaerobic fungal inoculation on the fermentation characteristics of rice straw silages. Journal of Applied Microbiology. 2015;118:565–573. doi: 10.1111/jam.12724. [DOI] [PubMed] [Google Scholar]
- Lee, Ha & Cheng (2000).Lee SS, Ha JK, Cheng KJ. Influence of an anaerobic fungal culture administration on in vivo ruminal fermentation and nutrient digestion. Animal Feed Science and Technology. 2000;88:201–217. doi: 10.1016/S0377-8401(00)00216-9. [DOI] [Google Scholar]
- Li et al. (2012).Li ZZ, Tao LL, Zhang J, Zhang HJ, Qu JM. Role of NOD2 in regulating the immune response to Aspergillus fumigatus. Inflammation Research. 2012;61:643–648. doi: 10.1007/s00011-012-0456-4. [DOI] [PubMed] [Google Scholar]
- Li-Byarlay et al. (2013).Li-Byarlay H, Li Y, Stroud H, Feng S, Newman TC, Kaneda M, Hou KK, Worley KC, Elsik CG, Wickline SA, Jacobsen SE, Ma J, Robinson GE. RNA interference knockdown of DNA methyl-transferase 3 affects gene alternative splicing in the honey bee. Proceedings of the National Academy of Sciences of the United States of America. 2013;110:12750–12755. doi: 10.1073/pnas.1310735110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lipardi & Paterson (2009).Lipardi C, Paterson BM. Identification of an RNA-dependent RNA polymerase in Drosophila involved in RNAi and transposon suppression. Proceedings of the National Academy of Sciences of the United States of America. 2009;106:15645–15650. doi: 10.1073/pnas.0904984106. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Maiti, Lee & Liu (2007).Maiti M, Lee HC, Liu Y. QIP, a putative exonuclease, interacts with the Neurospora Argonaute protein and facilitates conversion of duplex siRNA into single strands. Genes and Development. 2007;21:590–600. doi: 10.1101/gad.1497607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matityahu et al. (2008).Matityahu A, Hadar Y, Dosoretz CG, Belinky PA. Gene silencing by RNA interference in the white rot fungus Phanerochaete chrysosporium. Applied and Environmental Microbiology. 2008;74:5359–5365. doi: 10.1128/AEM.02433-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michielse et al. (2008).Michielse CB, Hooykaas PJJ, Van den Honde CAMJJ, Ram AFJ. Agrobacterium-mediated transformation of the filamentous fungus Aspergillus awamori. Nature Protocols. 2008;3:1671–1678. doi: 10.1038/nprot.2008.154. [DOI] [PubMed] [Google Scholar]
- Minz & Sharon (2010).Minz A, Sharon A. Electroporation and Agrobacterium-mediated spore transformation. In: Sharon A, editor. Molecular and cell biology methods for fungi. Humana Press; Totowa: 2010. pp. 21–32. [DOI] [PubMed] [Google Scholar]
- Moazeni et al. (2012).Moazeni M, Khoramizadeh MR, Kordbacheh P, Sepehrizadeh Z, Zeraati H, Noorbakhsh F, Teimoori-Toolabi L, Rezaie S. RNA-mediated gene silencing in Candida albicans: inhibition of hyphae formation by use of RNAi technology. Mycopathologia. 2012;174:177–185. doi: 10.1007/s11046-012-9539-6. [DOI] [PubMed] [Google Scholar]
- Moazeni et al. (2014).Moazeni M, Khoramizadeh MR, Teimoori-Toolabi L, Noorbakhsh F, Rezaie S. The effect of EFG1 gene silencing on down-regulation of SAP5 gene, by use of RNAi technology. Acta Medica Iranica. 2014;52:9–14. [PubMed] [Google Scholar]
- Morrison, Elshahed & Youssef (2016).Morrison JM, Elshahed MS, Youssef NH. Defined enzyme cocktail from the anaerobic fungus Orpinomyces sp. strain C1A effectively releases sugars from pretreated corn stover and switchgrass. Scientific Reports. 2016;6:29217. doi: 10.1038/srep29217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mousavi et al. (2015).Mousavi B, Hedayati MT, Teimoori-Toolabi L, Guillot J, Alizadeh A, Badali H. cyp51A gene silencing using RNA interference in azole-resistant Aspergillus fumigatus. Mycoses. 2015;58:699–706. doi: 10.1111/myc.12417. [DOI] [PubMed] [Google Scholar]
- Murata et al. (2007).Murata T, Kadotani N, Yamaguchi M, Tosa Y, Mayama S, Nakayashiki H. siRNA-dependent and -independent post-transcriptional cosuppression of the LTR-retrotransposon MAGGY in the phytopathogenic fungus Magnaporthe oryzae. Nucleic Acids Research. 2007;35:5987–5994. doi: 10.1093/nar/gkm646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakade et al. (2011).Nakade K, Watanabe H, Sakamoto Y, Sato T. Gene silencing of the Lentinula edodes lcc1 gene by expression of a homologous inverted repeat sequence. Microbiological Research. 2011;166:484–493. doi: 10.1016/j.micres.2010.09.004. [DOI] [PubMed] [Google Scholar]
- Nakayashiki et al. (2005).Nakayashiki H, Hanada S, Quoc NB, Kadotani N, Tosa Y, Mayama S. RNA silencing as a tool for exploring gene function in ascomycete fungi. Fungal Genetics and Biology. 2005;42:275–283. doi: 10.1016/j.fgb.2005.01.002. [DOI] [PubMed] [Google Scholar]
- Namekawa et al. (2005).Namekawa SH, Iwabata K, Sugawara H, Hamada FN, Koshiyama A, Chiku H, Kamada T, Sakaguchi K. Knockdown of LIM15/DMC1 in the mushroom Coprinus cinereus by double-stranded RNA-mediated gene silencing. Microbiology. 2005;151:3669–3678. doi: 10.1099/mic.0.28209-0. [DOI] [PubMed] [Google Scholar]
- Nicolas et al. (2008).Nicolas FE, Calo S, Murcia-Flores L, Garre V, Ruiz-Vazquez RM, Torres-Martinez S. A RING-finger photocarotenogenic repressor involved in asexual sporulation in Mucor circinelloides. FEMS Microbiology Letters. 2008;280:81–88. doi: 10.1111/j.1574-6968.2007.01044.x. [DOI] [PubMed] [Google Scholar]
- Nicolás, Torres-Martínez & Ruiz-Vázquez (2003).Nicolás FE, Torres-Martínez S, Ruiz-Vázquez RM. Two classes of small antisense RNAs in fungal RNA silencing triggered by non-integrative transgenes. EMBO Journal. 2003;22:3983–3991. doi: 10.1093/emboj/cdg384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nkemka et al. (2015).Nkemka VN, Gilroyed B, Yanke J, Gruninger R, Vedres D, McAllister T, Hao X. Bioaugmentation with an anaerobic fungus in a two-stage process for biohydrogen and biogas production using corn silage and cattail. Bioresource Technology. 2015;185:79–88. doi: 10.1016/j.biortech.2015.02.100. [DOI] [PubMed] [Google Scholar]
- Nolan et al. (2005).Nolan T, Braccini L, Azzalin G, De Toni A, Macino G, Cogoni C. The post-transcriptional gene silencing machinery functions independently of DNA methylation to repress a LINE1-like retrotransposon in Neurospora crassa. Nucleic Acids Research. 2005;33:1564–1573. doi: 10.1093/nar/gki300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patel et al. (2008).Patel RM, Van Kan JA, Bailey AM, Foster GD. RNA-mediated gene silencing of superoxide dismutase (bcsod1) in Botrytis cinerea. Phytopathology. 2008;98:1334–1339. doi: 10.1094/PHYTO-98-12-1334. [DOI] [PubMed] [Google Scholar]
- Paul et al. (2011).Paul SS, Deb SM, Punia BS, Das KS, Singh G, Ashar MN, Kumar R. Effect of feeding isolates of anaerobic fungus Neocallimastix sp. CF 17 on growth rate and fibre digestion in buffalo calves. Archives of Animal Nutrition. 2011;65:215–228. doi: 10.1080/1745039x.2011.559722. [DOI] [PubMed] [Google Scholar]
- Paul et al. (2004).Paul SS, Kamra DN, Sastry VRB, Sahu NP, Agarwal N. Effect of administration of an anaerobic gut fungus isolated from wild blue bull (Boselaphus tragocamelus) to buffaloes (Bubalus bubalis) on in vivo ruminal fermentation and digestion of nutrients. Animal Feed Science and Technology. 2004;115:143–157. doi: 10.1016/j.anifeedsci.2004.01.010. [DOI] [Google Scholar]
- Penn et al. (2015).Penn TJ, Wood ME, Soanes DM, Csukai M, Corran AJ, Talbot NJ. Protein kinase C is essential for viability of the rice blast fungus Magnaporthe oryzae. Molecular Microbiology. 2015;98:403–419. doi: 10.1111/mmi.13132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pickford et al. (2002).Pickford AS, Catalanotto C, Cogoni C, Macino G. Quelling in Neurospora crassa. Advances in Genetics. 2002;46:277–303. doi: 10.1016/S0065-2660(02)46010-5. [DOI] [PubMed] [Google Scholar]
- Prakash, Manjrekar & Chattoo (2016).Prakash C, Manjrekar J, Chattoo BB. Skp1, a component of E3 ubiquitin ligase, is necessary for growth, sporulation, development and pathogenicity in rice blast fungus (Magnaporthe oryzae) Molecular Plant Pathology. 2016;17:903–919. doi: 10.1111/mpp.12336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Procházka et al. (2012).Procházka J, Mrázek J, Štrosová L, Fliegerová K, Zábranská J, Dohányos M. Enhanced biogas yield from energy crops with rumen anaerobic fungi. Engineering in Life Sciences. 2012;12:343–351. doi: 10.1002/elsc.201100076. [DOI] [Google Scholar]
- Quoc & Nakayashiki (2015).Quoc NB, Nakayashiki H. RNA silencing in filamentous fungi: from basics to applications. In: Van den Berg MA, Maruthachalam K, editors. Genetic transformation systems in fungi. Vol 2. Springer International Publishing; Gewerbestrasse: 2015. pp. 107–124. [Google Scholar]
- Ranganathan et al. (2017).Ranganathan A, Smith OP, Youssef NH, Struchtemeyer CG, Atiyeh HK, Elshahed MS. Utilizing anaerobic fungi for two-stage sugar extraction and biofuel production from lignocellulosic biomass. Frontiers in Microbiology. 2017;8 doi: 10.3389/fmicb.2017.00635. Article 635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson, McCarthy & Smyth (2010).Robinson MD, McCarthy DJ, Smyth GK. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics. 2010;26:139–140. doi: 10.1093/bioinformatics/btp616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Romano & Macino (1992).Romano N, Macino G. Quelling: transient inactivation of gene expression in Neurospora crassa by transformation with homologous sequences. Molecular Microbiology. 1992;6:3343–3353. doi: 10.1111/j.1365-2958.1992.tb02202.x. [DOI] [PubMed] [Google Scholar]
- Saxena et al. (2010).Saxena S, Sehgal J, Puniya A, Singh K. Effect of administration of rumen fungi on production performance of lactating buffaloes. Beneficial Microbes. 2010;1:183–188. doi: 10.3920/BM2009.0018. [DOI] [PubMed] [Google Scholar]
- Segers et al. (2007).Segers GC, Zhang X, Deng F, Sun Q, Nuss DL. Evidence that RNA silencing functions as an antiviral defense mechanism in fungi. Proceedings of the National Academy of Sciences of the United States of America. 2007;104:12902–12906. doi: 10.1073/pnas.0702500104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sehgal et al. (2008).Sehgal JP, Jit D, Puniya AK, Singh K. Influence of anaerobic fungal administration on growth, rumen fermentation and nutrient digestion in female buffalo calves. Journal of Animal and Feed Sciences. 2008;17:510–518. doi: 10.22358/jafs/66678/2008. [DOI] [Google Scholar]
- Skowyra & Doering (2012).Skowyra ML, Doering TL. RNA interference in Cryptococcus neoformans. Methods in Molecular Biology. 2012;845:165–186. doi: 10.1007/978-1-61779-539-8_11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smardon et al. (2000).Smardon A, Spoerke JM, Stacey SC, Klein ME, Mackin N, Maine EM. EGO-1 is related to RNA-directed RNA polymerase and functions in germ-line development and RNA interference in C. legans. Current Biology. 2000;10:169–178. doi: 10.1016/S0960-9822(00)00323-7. [DOI] [PubMed] [Google Scholar]
- Solomon et al. (2016).Solomon KV, Haitjema CH, Henske JK, Gilmore SP, Borges-Rivera D, Lipzen A, Brewer HM, Purvine SO, Wright AT, Theodorou MK, Grigoriev IV, Regev A, Thompson DA, O’Malley MA. Early-branching gut fungi possess a large, comprehensive array of biomass-degrading enzymes. Science. 2016;351:1192–1195. doi: 10.1126/science.aad1431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stein et al. (2003).Stein P, Svoboda P, Anger M, Schultz RM. RNAi: mammalian oocytes do it without RNA-dependent RNA polymerase. RNA. 2003;9:187–192. doi: 10.1261/rna.2860603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun, Choi & Nuss (2009).Sun Q, Choi GH, Nuss DL. A single Argonaute gene is required for induction of RNA silencing antiviral defense and promotes viral RNA recombination. Proceedings of the National Academy of Sciences of the United States of America. 2009;106:17927–17932. doi: 10.1073/pnas.0907552106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tripathi et al. (2007).Tripathi VK, Sehgal JP, Puniya AK, Singh K. Effect of administration of anaerobic fungi isolated from cattle and wild blue bull (Boselaphus tragocamelus) on growth rate and fibre utilization in buffalo calves. Archives of Animal Nutrition. 2007;61:416–423. doi: 10.1080/17450390701556759. [DOI] [PubMed] [Google Scholar]
- Tuschl et al. (1999).Tuschl T, Zamore PD, Lehmann R, Bartel DP, Sharp PA. Targeted mRNA degradation by double-stranded RNA in vitro. Genes and Development. 1999;13:3191–3197. doi: 10.1101/gad.13.24.3191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang, Chen & Hseu (2014).Wang HC, Chen YC, Hseu RS. Purification and characterization of a cellulolytic multienzyme complex produced by Neocallimastix patriciarum J11. Biochemical and Biophysical Research Communications. 2014;451:190–195. doi: 10.1016/j.bbrc.2014.07.088. [DOI] [PubMed] [Google Scholar]
- Wei et al. (2016a).Wei YQ, Long RJ, Yang H, Yang HJ, Shen XH, Shi RF, Wang ZY, Du JG, Qi XJ, Ye QH. Fiber degradation potential of natural co-cultures of Neocallimastix frontalis and Methanobrevibacter ruminantium isolated from yaks (Bos grunniens) grazing on the Qinghai Tibetan Plateau. Anaerobe. 2016a;39:158–164. doi: 10.1016/j.anaerobe.2016.03.005. [DOI] [PubMed] [Google Scholar]
- Wei et al. (2016b).Wei YQ, Yang HJ, Luan Y, Long RJ, Wu YJ, Wang ZY. Isolation, identification and fibrolytic characteristics of rumen fungi grown with indigenous methanogen from yaks (Bos grunniens) grazing on the Qinghai-Tibetan Plateau. Journal of Applied Microbiology. 2016b;120:571–587. doi: 10.1111/jam.13035. [DOI] [PubMed] [Google Scholar]
- Youssef et al. (2013).Youssef NH, Couger MB, Struchtemeyer CG, Liggenstoffer AS, Prade RA, Najar FZ, Atiyeh HK, Wilkins MR, Elshahed MS. The genome of the anaerobic fungus Orpinomyces sp. strain C1A reveals the unique evolutionary history of a remarkable plant biomass degrader. Applied and Environmental Microbiology. 2013;79:4620–4634. doi: 10.1128/AEM.00821-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang et al. (2008).Zhang X, Segers GC, Sun Q, Deng F, Nuss DL. Characterization of Hypovirus-derived small RNAs generated in the chestnut blight fungus by an inducible DCL-2-dependent pathway. Journal of Virology. 2008;82:2613–2619. doi: 10.1128/JVI.02324-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1. Uptake of fluorescently (Cy3) tagged siRNA by C1A spores. (A) The ldhD-specific siRNA was added to the flooding solution 75 minutes after the onset of flooding followed by incubation for 15 more minutes at 39°C. Samples (a few microliters) were taken at regular intervals for visualization. The same field is shown for DAPI-, and Cy3-labeled germinating spores (Note that the spores were concurrently stained with DAPI and fluorescing green indicating the uptake of the Cy3-labeled siRNA) (bar =2000B5). (B) Effect of the siRNA treatment on fungal growth rate. siRNA-treated spores were collected and used to inoculate fresh RFC medium. Control cultures were started at the same time using siRNA-untreated spores. Headspace pressure was measured daily and used to calculate fungal biomass as described previously (1). Error bars are standard deviations from at least three replicate cultures for each condition.
Table S1. Silencing efficiency (based on the total number of mmoles of D-lactate in the culture supernatant) of ldhD-siRNA during early log (6–13 mg biomass), mid-log (14–17 mg biomass), and late log/early stationary (18–23 mg) phases a.
Table S2. Transcripts with a significant (False Discovery Rate (FDR) <0.1) fold change in the ldhD-siRNA-treated cultures.
Data Availability Statement
The following information was supplied regarding data availability:
This Transcriptome Shotgun Assembly project has been deposited at DDBJ/EMBL/GenBank under the accession GFSU00000000. The version described in this paper is the first version, GFSU01000000.