

Abstract
Latent membrane protein 1 (LMP1) is a primary oncogene encoded by the Epstein‐Barr virus, and various portions of LMP1 are detected in nasopharyngeal carcinoma (NPC) tumor cells. LMP1 has been extensively studied since the discovery of its transforming property in 1985. LMP1 promotes cancer cell growth during NPC development and facilitates the interaction of cancer cells with surrounding stromal cells for invasion, angiogenesis, and immune modulation. LMP1 is detected in 100% of pre‐invasive NPC tumors and in approximately 50% of advanced NPC tumors. Moreover, a small population of LMP1‐expressing cells in advanced NPC tumor tissue is proposed to orchestrate NPC tumor tissue maintenance and development through cancer stem cells and progenitor cells. Recent studies suggest that LMP1 activity shifts according to tumor development stage, but it still has a pivotal role during all stages of NPC development.
Keywords: cancer stem cell, Epstein‐Barr virus, immune evasion, latent membrane protein 1, nasopharyngeal carcinoma
1. INTRODUCTION
Epstein‐Barr virus (EBV) was discovered in Burkitt's lymphoma tumor cells in 1964 and is a causative agent of various human cancers.1, 2 Most of these cancers arise in the lymphatic organ, especially B lymphocyte‐origin tumors.3, 4 Nasopharyngeal carcinoma (NPC), an EBV‐associated malignant disease, is unique in that it is an epithelial origin tumor and expresses EBV primary oncogene latent membrane protein 1 (LMP1) at the premalignant stage.2, 5 Normal pharyngeal epithelial cells are a proliferating site for EBV, and EBV infection results in lysis of the infected cell. Thus, switching from lytic EBV infection to a latent one in the epithelium is an initial step in NPC carcinogenesis.6 Inactivation of the p16 tumor suppressor and overexpression of cyclin D1, commonly detected in the premalignant and dysplastic nasopharyngeal epithelium, may enable the transition to an EBV latent infection in the nasopharyngeal epithelium.7, 8 Of the 3 types of latent infection, NPC is categorized as a type II infection, which is associated with the expression of 3 virus‐encoded proteins (EBV nuclear antigen 1, LMP1, and LMP2) and some virus‐encoded RNAs, EBV‐encoded small RNAs (EBERs) and BamHI A rightward transcripts (BARTs). Among these EBV gene products, LMP1 has a major role in NPC carcinogenesis (Table 1).3, 9 In addition, the importance of the tumor microenvironment is becoming clear with recent progress in the cancer research field.
Table 1.
Epstein‐Barr virus (EBV) gene expression and viral latency
| EBV latency | EBV gene transcription | Infected cell types and tumors |
|---|---|---|
| Type 0 | EBERs | Memory B cell |
| Type I | EBERs, EBNA1, BARTs | Burkitt's lymphoma |
| Type II | EBERs, EBNA1, BARTs, LMP1, LMP2 | Nasopharyngeal carcinoma, gastric cancer,a Hodgkin's lymphoma, NK/T lymphoma |
| Type III | EBERs, EBNA1, EBNA‐LP, EBNA2, EBNA3A‐C, BARTs, LMP1, LMP2 | Lymphoblastoid cell (infectious mononucleosis), post‐transplant lymphoproliferative disease, patients with immunosuppression |
BART, BamHI A rightward transcript; EBER, EBV‐encoded small RNA; EBNA, EBV nuclear antigen; EBNA‐LP, EBV nuclear antigen leader protein.
In this review, we tentatively categorized in latency type II.
Expression of latent membrane protein 1 (LMP1) in gastric cancer and EBV latency of gastric cancer is controversial.
Here, we review the pathogenic roles of LMP1 in the tumor microenvironment during the development of NPC.
2. LATENT MEMBRANE PROTEIN 1 EXPRESSION IN NPC
Latent membrane protein 1 was recognized as a primary transforming gene product of EBV after its transforming property in rodent fibroblasts was reported.10 More than 30 years of LMP1 research revealed its multipotency to modulate various cell signal transduction pathways, leading to the proliferation and simultaneous suppression of apoptotic programs in LMP1‐expressing cells.11, 12, 13 It is involved in not only cell transformation, but also invasion, metastasis upregulation, cytokine production, immune modulation, and tumor angiogenesis.
Although LMP1 protein was detected in only 20%‐40% of NPC samples in a series of previous reports, more sensitive approaches using immunohistochemical staining have revealed LMP1 expression in almost 100% of primary NPC specimens.12, 14 However, LMP1 expression is highly heterogeneous among NPC tumor cells. The population of LMP1‐positive cells is low, and LMP1 is usually detected in sparse cells and small clusters. Latent membrane protein 1 is detected in almost 100% of premalignant lesions and is especially concentrated in the basal layer, which suggests that LMP1 mainly contributes to the early stage of tumor development, and only a small population of cancer cells retains a high level of LMP1 expression in advanced NPC (Table 2).5, 14 Latent membrane protein 1 is a double‐edged sword because abundant LMP1 expression inhibits cell growth and induces apoptosis in epithelial cells.15 High LMP1 expression may be involved in the maintenance of stem cell properties and LMP1‐expressing epithelial cells may represent a cancer stem cell or progenitor‐like cell phenotype.15, 16, 17 These reports suggest that the pathogenic role of LMP1 may change during NPC progression (Figure 1).
Table 2.
Comparison of latent membrane protein 1 expression in early and advanced nasopharangeal carcinoma
| Premalignant | Advanced | |
|---|---|---|
| Detection rate (IHC) | 100% | 20%‐50% |
| Localization | Basal layer | Sporadically in tumor |
| Expression level | Rather homogeneous | Heterogeneous |
IHC, immunohistochemistry.
Figure 1.
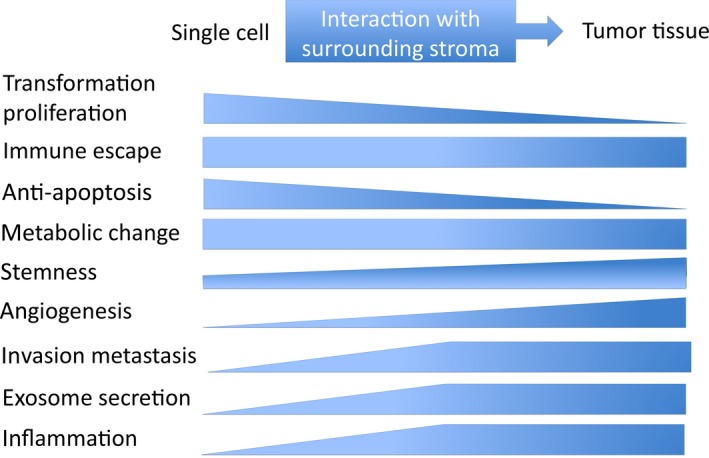
Role of latent membrane protein 1 (LMP1) shifts with tumor progression. The role of LMP1 changes according to the progression of the tumor. The transforming and anti‐apoptotic property of LMP1 is essential at early carcinogenic stages. As LMP1‐positive nasopharyngeal cancer cells progress to clinical tumor tissues surrounded by stromal cells, the role of LMP1 changes to upregulate invasive, angiogenic pathways and to maintain or increase the tumor tissue bulk
3. INTERPLAY WITH THE MICROENVIRONMENT
The development of cancer tissue depends on the interaction between cancer cells and various stromal components. Here, we focus on the recent progress in understanding the pathogenic role of LMP1 within the NPC microenvironment.
The role of LMP1 in invasion and metastasis has been thoroughly examined since the induction of MMP9 by LMP1 was reported.18 Invasion and metastasis is composed of a multistep interaction between tumor and stromal cells. In addition to the initial transforming property, LMP1 modulates expression of various receptors, enzymes, and cytokines in LMP1‐expressing cells, which affects the surrounding microenvironment and promotes metastasis of LMP1‐expressing cells.19, 20 The epithelial‐mesenchymal transition (EMT) produces cells with properties of stem cells, which suggests that EMT activity may be involved in generating stemness in cancer.21, 22 In particular, LMP1‐transformed epithelial cells cause fibroblasts to lose polarity, and upregulation of EMT by LMP1 occurs through upregulation of Twist, Snail, and SatB1 in LMP1‐expressing cells.23, 24, 25
Most oncogenic viruses are capable of stabilizing or enhancing the transcriptional activity of the angiogenic factor hypoxia‐inducible factor‐1α (HIF‐1α), which integrates several biological pathways for viral carcinogenesis.26, 27 Latent membrane protein 1 also induces HIF‐1α.28 Angiogenesis and lymphangiogenesis surrounding LMP1‐expressing cells were also promoted through upregulation of angiogenic factors such as vascular endothelial growth factor, basic fibroblast growth factor, and interleukin‐8 (IL‐8).29, 30, 31, 32, 33
4. LATENT MEMBRANE PROTEIN 1 AND THE WARBURG EFFECT
Aerobic glycolysis, glucose consumption, and lactate production even in the presence of oxygen is called the Warburg effect and is a main hallmark of cancer.34 The Warburg effect can also be induced by EBV.35 Latent membrane protein 1 upregulates the HIF‐1α pathway regardless of oxygen pressure in the tumor microenvironment. Moreover, glucose is phosphorylated into glucose‐6‐phosphate in the glycolytic pathway.36 Hexokinase 2 (HK2) catalyzes the first step in this glycolytic pathway, and thus, overexpression of HK2 is a common feature of cancer.
Latent membrane protein 1 reprograms glycolysis by upregulating HK2, which binds to mitochondria in NPC, and c‐Myc is required for LMP1‐induced upregulation of HK2.35 In addition, LMP1 upregulates the receptor for advanced glycation end products (RAGE) in NPC through the activation of nuclear factor‐κB, and microvessel count in NPC tumors is associated with the expression of LMP1 and RAGE.37
Hypoxic conditions help maintain stemness in advanced bulky tumors. Latent membrane protein 1 activates the hypoxic signaling pathway for cells with cancer stem‐ or progenitor‐like properties before these cells become advanced tumor cells under a hypoxic microenvironment, which helps to maintain cancer stem cells in an early stage of NPC development.12, 17
5. LATENT MEMBRANE PROTEIN 1 IS SECRETED THROUGH EXOSOMES
Exosomes are a recently discovered mechanism through which cancer cells and virally infected cells can manipulate the microenvironment. Exosomes are 40‐100 nm endosomal‐derived vesicles containing transfer proteins, mRNAs, and microRNAs secreted from many cell types to neighboring or distant cells to modulate immune function, angiogenesis, cell proliferation, tumor invasion, and cell‐to‐cell communication.38, 39
Latent membrane protein 1 localizes at the Golgi and multivesicular compartments, and is packaged into exosomes for release from the cell.40 Exosomes containing LMP1 have been detected in the serum of NPC patients, which induces T‐cell anergy and escape from immune surveillance.41, 42 A nanogram amount of purified recombinant LMP1 strongly suppresses activation of T cells and natural killer (NK) cells.42, 43, 44 In addition to LMP1 exosomal secretion, LMP1 also increases the concentration and release of basic fibroblast growth factor into exosomes.32 Latent membrane protein 1 induces the expression of the epidermal growth factor receptor in an EBV‐negative epithelial cell line, and exosomes produced by these cells also contain high levels of the epidermal growth factor receptor in exosomes.41, 45 Latent membrane protein 1 increases the levels of HIF‐1α in exosomes, which are then delivered to surrounding tumor cells for EMT induction and pro‐metastatic effects.46 These findings suggest that the effects of EBV and LMP1 on cellular expression also modulate exosome content and properties. It is increasingly evident that viruses, such as EBV, can hijack and utilize the host cell exosome pathway to modulate cell‐cell signaling and control the tumor microenvironment.
6. INTERACTION WITH STROMAL CELLS
A dense infiltration of lymphocytes, called lymphoepithelioma, is a well‐known pathological characteristic of NPC.47 In addition to exosomal secretion of LMP1, the close association of EBV infection with lymphoepithelioma‐like carcinoma implies that the poorly differentiated properties of epithelial cells and an inflammatory environment are involved in viral oncogenesis. To develop and maintain this coexistence of tumor infiltrates lymphocytes (TIL) with EBV‐infected cells, clever immune evasion systems from the early phase of EBV infection is necessary. Infiltrating cells consist of different subsets of T‐lymphocytes, including CD8+ CTLs, which are capable of killing tumor cells.48 The CD8+ T‐cell responses to LMP1 are generally low and rarely detected in healthy virus carriers. In addition to exosome‐mediated immune modification, EBV has multiple ways to escape from immune attack.49, 50
Latent membrane protein 1 localizes to lipid rafts in the cell membrane when aggregated.51 An LMP1 mutant that cannot aggregate at lipid rafts is recognized by HLA epitope‐sensitized CTLs, but WT and LMP mutants that retain aggregation and localization to lipid rafts are not. These results suggest that aggregation of LMP1 at lipid rafts protects LMP1 from being processed and presented with MHC class I. In addition, LMP‐1 limits self‐presentation without compromising its ability to modulate trans‐presentation of CD8+ T‐cell epitopes.52
Programmed cell death protein‐1 is a negative checkpoint molecule. Programmed cell death protein‐1, expressed by T‐lymphocytes, and programmed cell death ligand 1 (PD‐L1), expressed by tumor cells, contribute to T‐cell inhibition, and thus are correlated with the prognosis of various cancers including NPC.53, 54, 55, 56 The LMP1 and γ‐interferon pathways co‐operate to regulate PD‐L1 (Figure 2). Expression of PD‐L1 was more abundant in EBV‐positive NPC cell lines than in EBV‐negative cell lines. Knockdown of LMP1 expression suppressed PD‐L1 expression in EBV‐positive cell lines. In addition, γ‐interferon upregulates PD‐L1 expression independently and synergistically with LMP1 in NPC tissue, and PD‐L1 expression is associated with worse disease‐free survival in patients with NPC.57
Figure 2.
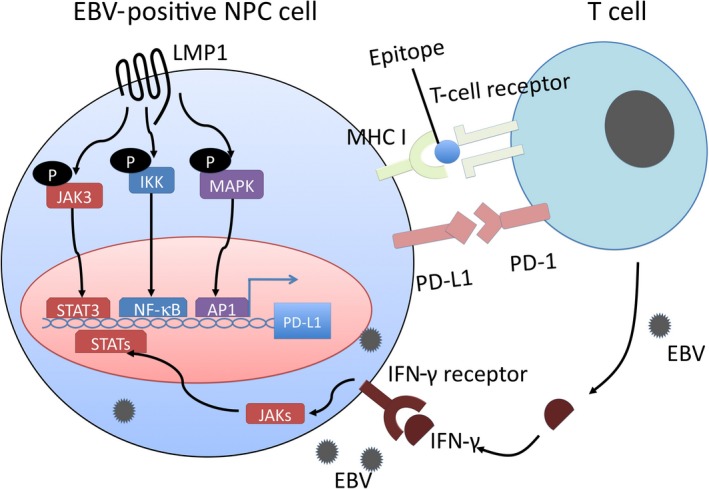
Two possible mechanisms of programmed cell death ligand 1 (PD‐L1) regulation in Epstein‐Barr virus (EBV)‐positive nasopharyngeal carcinoma (NPC). Upregulation of PD‐L1 is mediated by 2 mechanisms. One is the latent membrane protein 1 (LMP1)‐mediated oncogenic pathway, which is independent of inflammatory signals in the tumor microenvironment. The other is the γ‐interferon (IFN‐γ) mediated inflammatory pathway, which depends on an antiviral and antitumor immune response. IKK, IκB kinase; NF‐κB, nuclear factor‐κB; PD‐1, programmed cell death protein‐1; STAT, signal transducer and activator of transcription
Latent membrane protein 1 promotes myeloid‐derived suppressor cell expansion in the tumor microenvironment by promoting extra‐mitochondrial glycolysis in malignant cells. In addition to RAGE, LMP1 promotes the expression of multiple glycolytic genes such as GLUT1. This metabolic reprogramming induces the Nod‐like receptor family protein 3 inflammasome, activates the arachidonic cascade, and consequently various cytokines such as IL‐1β, IL‐6, and granulocyte/macrophage colony‐stimulating factor. These changes in the tumor environment result in NPC tumor‐derived myeloid‐derived suppressor cell induction.58
7. PERSISTENT EBV INFECTION IN NPC TUMOR CELLS
In contrast to EBV‐associated lymphomagenesis, a major obstacle to elucidate the mechanisms of EBV‐mediated NPC initiation is that the genomes of EBV in NPC cell lines are gradually lost after prolonged in vitro culture.59, 60 This suggests that EBV is not essential for in vitro proliferation of NPC tumor cells. The copy number of the EBV episome decreases without drug selection or cell sorting. Analysis of the distributions of EBV‐derived plasmids in single live cells throughout the cell cycle revealed that 16% of plasmids failed to duplicate, leading to a net loss of 8% of the plasmids from a population of cells.61 Nevertheless, a high proportion of NPC tumors retain the EBV genome regardless of the endemic or non‐endemic area of NPC. The histological feature of EBV‐positive NPC that obviously differs from EBV‐negative NPC is the undifferentiated state of tumor cells and heavy lymphoid cell infiltration in stroma.47 Expression of LMP1 prevents epithelial cell differentiation.62 Interaction of NPC tumor cells with TIL is thought to be maintained through cytokine production and exosome excretion. The ubiquitous existence of EBV in advanced NPC tumors, recurrent NPC tumors, and passaged NPC xenografts in nude mice suggests that EBV infection in epithelial cells provides an advantage for EBV‐infected epithelial cells grown in vivo.63, 64 Similarly, inflammation induced by EBV infection and expression of EBV genes may enhance initiation and progression of NPC in vivo.65 In addition, LMP1‐mediated cytokine production might attract migration of inflammatory cells, including activated macrophages and T regulatory cells, to support EBV‐infected cell growth and immune avoidance. Moreover, stromal lymphoid cells in NPC tissues secrete IL‐6 to stimulate cell growth of EBV‐infected NPC cells.66, 67 This mutually dependent relationship between cancer cells and stroma cells suggests that LMP1 expression levels and the LMP1‐positive NPC tumor cell population is also regulated by the tumor‐stroma interaction.
8. CONCLUDING REMARKS
Although EBV products, including LMP1, enhance the invasive and metastatic capabilities of EBV‐infected malignant cells, the overall impact of EBV for the prognosis of NPC patients is somewhat limited.67 Similar to human papillomavirus (HPV)‐associated oropharyngeal carcinoma, which has a better prognosis than non‐HPV‐associated oropharyngeal carcinoma, patients with EBV‐associated NPC have a better prognosis compared to patients with non‐EBV‐associated NPC.47, 68, 69 This might seem contradictory considering the results discussed in this review, but EBV‐associated NPC is sensitive to chemoradiotherapy treatment and immune checkpoint inhibitors.
LMP1 is not the only EBV gene expressed in NPC. Emerging evidence has revealed that EBV‐encoded non‐coding RNAs, such as EBERs, BARTs, and miR‐BARTs, also contribute directly to NPC development. For example, EBERs contribute to immune evasion by inducing IL‐10 through protein kinase R‐independent pathways.70 Thus, the function of these abundantly expressed RNAs suggests an interplay with tumor cells and the microenvironment.11, 12, 71, 72 In addition, for Hodgkin's lymphoma and nasal NK/T‐cell lymphoma, which are distinct type II latency tumors originating in B cells and NK/T cells, LMP1 may have different roles in a cell type‐specific manner.73, 74, 75
Finally, investigation into the role of heavily infiltrated lymphoid stroma will be important for understanding the pathogenic role of EBV in epithelial malignancies, including NPC. Hopefully, this will provide clues to develop new therapeutic targets in addition to emerging immune checkpoint inhibitors.
CONFLICT OF INTEREST
The authors have no conflict of interest.
ACKNOWLEDGMENTS
This work was supported by the Japan Society for the Promotion of Science (KAKENHI Grant Nos. JP26293367, JP26670739, 17K19718, and 17H01590).
Yoshizaki T, Kondo S, Endo K, et al. Modulation of the tumor microenvironment by Epstein‐Barr virus latent membrane protein 1 in nasopharyngeal carcinoma. Cancer Sci. 2018;109:272–278. https://doi.org/10.1111/cas.13473
REFERENCES
- 1. Epstein MA, Barr YM. Cultivation in vitro of human lymphoblasts from Burkitt's lymphoma. Lancet. 1964;1:252‐253. [DOI] [PubMed] [Google Scholar]
- 2. Rickinson AB. Co‐infections, inflammation and oncogenesis: future directions for EBV research. Semin Cancer Biol. 2014;26:99‐115. [DOI] [PubMed] [Google Scholar]
- 3. Young LS, Rickinson AB. Epstein–Barr virus: 40 years on. Nat Rev Cancer. 2004;4:757‐768. [DOI] [PubMed] [Google Scholar]
- 4. Vockerodt M, Yap L‐F, Shannon‐Lowe C, et al. The Epstein–Barr virus and the pathogenesis of lymphoma. J Pathol. 2015;235:312‐322. [DOI] [PubMed] [Google Scholar]
- 5. Pathmanathan R, Prasad U, Sadler R, Flynn K, Raab‐Traub N. Clonal proliferations of cells infected with Epstein‐Barr virus in preinvasive lesions related to nasopharyngeal carcinoma. N Engl J Med. 1995;333:693‐698. [DOI] [PubMed] [Google Scholar]
- 6. Thorley‐Lawson DA, Hawkins JB, Tracy SI, et al. The pathogenesis of Epstein–Barr virus persistent infection. Curr Opin Virol. 2013;3:227‐232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Lo KW, To KF, Huang DP. Focus on nasopharyngeal carcinoma. Cancer Cell. 2004;5:423‐428. [DOI] [PubMed] [Google Scholar]
- 8. Tsang CM, Yip YL, Lo KW, et al. Cyclin D1 overexpression supports stable EBV infection in nasopharyngeal epithelial cells. Proc Natl Acad Sci USA. 2012;109:E3473‐E3482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Tsang CM, Zhang G, Seto E, et al. Epstein–Barr virus infection in immortalized nasopharyngeal epithelial cells: regulation of infection and phenotypic characterization. Int J Cancer. 2010;127:1570‐1583. [DOI] [PubMed] [Google Scholar]
- 10. Wang D, Liebowitz D, Kieff E. An EBV membrane protein expressed in immortalized lymphocytes transforms established rodent cells. Cell. 1985;43:831‐840. [DOI] [PubMed] [Google Scholar]
- 11. Yoshizaki T, Kondo S, Wakisaka N, et al. Pathogenic role of Epstein‐Barr virus latent membrane protein‐1 in the development of nasopharyngeal carcinoma. Cancer Lett. 2013;337:1‐7. [DOI] [PubMed] [Google Scholar]
- 12. Tsao SW, Tsang CM, To KF, Lo KW. The role of Epstein–Barr virus in epithelial malignancies. J Pathol. 2015;235:323‐333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Lu QL, Elia G, Lucas S, et al. Bcl‐2 proto‐oncogene expression in Epstein–Barr‐virus‐associated nasopharyngeal carcinoma. Int J Cancer. 1993;53:29‐35. [DOI] [PubMed] [Google Scholar]
- 14. Benders AA, Tang W, Middeldorp JM, et al. Epstein–Barr virus latent membrane protein 1 is not associated with vessel density nor with hypoxia inducible factor‐1α expression in nasopharyngeal carcinoma tissue. Head Neck Pathol. 2009;3:276‐282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Tsao SW, Tramoutanis G, Dawson CW, et al. The significance of LMP1 expression in nasopharyngeal carcinoma. Semin Cancer Biol. 2002;12:473‐487. [DOI] [PubMed] [Google Scholar]
- 16. Lun SW, Cheung ST, Cheung PF, et al. CD44+ cancer stem‐like cells in EBV‐associated nasopharyngeal carcinoma. PLoS One. 2012;7:e52426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Kondo S, Wakisaka N, Muramatsu M, et al. Epstein–Barr virus latent membrane protein 1 induces cancer stem/progenitor‐like cells in nasopharyngeal epithelial cell lines. J Virol. 2011;85:11255‐11264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Yoshizaki T, Sato H, Furukawa M, Pagano JS. The expression of matrix metalloproteinase 9 is enhanced by Epstein‐Barr virus latent membrane protein 1. Proc Natl Acad Sci USA. 1998;95:3621‐3626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Yoshizaki T, Wakisaka N, Pagano JS. Chapter 12, Epstein‐Barr virus, Invasion and Metastasis. London: Caister Academic Press; 2005. [Google Scholar]
- 20. Zhao Y, Wang Y, Zeng S, Hu X. LMP1 expression is positively associated with metastasis of nasopharyngeal carcinoma: evidence from a meta‐analysis. J Clin Pathol. 2012;65:41‐45. [DOI] [PubMed] [Google Scholar]
- 21. Mani SA, Guo W, Liao MJ, et al. The epithelial‐mesenchymal transition generates cells with properties of stem cells. Cell. 2008;133:704‐715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Polyak K, Weinberg RA. Transitions between epithelial and mesenchymal states: acquisition of malignant and stem cell traits. Nat Rev Cancer. 2009;9:265‐273. [DOI] [PubMed] [Google Scholar]
- 23. Horikawa T, Yang J, Kondo S, et al. Twist and epithelial‐mesenchymal transition are induced by the EBV oncoprotein latent membrane protein 1 and are associated with metastatic nasopharyngeal carcinoma. Cancer Res. 2007;67:1970‐1978. [DOI] [PubMed] [Google Scholar]
- 24. Horikawa T, Yoshizaki T, Kondo S, Furukawa M, Kaizaki Y, Pagano JS. Epstein‐Barr virus latent membrane protein 1 induces Snail and epithelial‐mesenchymal transition in metastatic nasopharyngeal carcinoma. Br J Cancer. 2011;104:1160‐1167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Endo K, Shackelford J, Aga M, Yoshizaki T, Pagano JS. Up‐regulation of SATB1 by EBV LMP1 in human nasopharyngeal cells and nasopharyngeal cancer. J Gen Virol. 2013;94:507‐513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Semenza GL. Defining the role of hypoxia‐inducible factor 1 in cancer biology and therapeutics. Oncogene. 2010;29:625‐634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Cuninghame S, Jackson R, Zehbe I. Hypoxia‐inducible factor 1 and its role in viral carcinogenesis. Virology. 2014;456–457:370‐383. [DOI] [PubMed] [Google Scholar]
- 28. Wakisaka N, Kondo S, Yoshizaki T, Murono S, Furukawa M, Pagano JS. Epstein‐Barr virus latent membrane protein 1 induces synthesis of hypoxia‐inducible factor 1 alpha. Mol Cell Biol. 2004;24:5223‐5234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Murono S, Inoue H, Tanabe T, et al. Induction of cyclooxygenase‐2 by Epstein‐Barr virus latent membrane protein 1 is involved in vascular endothelial growth factor production in nasopharyngeal carcinoma cells. Proc Natl Acad Sci USA. 2001;98:6905‐6910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Kondo S, Seo SY, Yoshizaki T, et al. EBV latent membrane protein 1 up‐regulates hypoxia‐inducible factor 1alpha through Siah1‐mediated down‐regulation of prolyl hydroxylases 1 and 3 in nasopharyngeal epithelial cells. Cancer Res. 2006;66:9870‐9877. [DOI] [PubMed] [Google Scholar]
- 31. Wakisaka N, Hirota K, Kondo S, et al. Induction of lymphangiogenesis through vascular endothelial growth factor‐C/vascular endothelial growth factor receptor 3 axis and its correlation with lymph node metastasis in nasopharyngeal carcinoma. Oral Oncol. 2012;48:703‐708. [DOI] [PubMed] [Google Scholar]
- 32. Wakisaka N, Murono S, Yoshizaki T, Furukawa M, Pagano JS. Epstein‐Barr virus latent membrane protein 1 induces and causes release of fibroblast growth factor‐2. Cancer Res. 2002;62:6337‐6344. [PubMed] [Google Scholar]
- 33. Yoshizaki T, Horikawa T, Qing‐Chun R, et al. Induction of interleukin‐8 by Epstein‐Barr virus latent membrane protein‐1 and its correlation to angiogenesis in nasopharyngeal carcinoma. Clin Cancer Res. 2001;7:1946‐1951. [PubMed] [Google Scholar]
- 34. Vander Heiden MG, Cantley LC, Thompson CB. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science. 2009;324:1029‐1033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Xiao L, Hu ZY, Dong X, et al. Targeting Epstein‐Barr virus oncoprotein LMP1‐mediated glycolysis sensitizes nasopharyngeal carcinoma to radiation therapy. Oncogene. 2014;33:4568‐4578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Mathupala SP, Ko YH, Pedersen PL. Hexokinase‐2 bound to mitochondria: cancer's stygian link to the ‘Warburg effect’ and a pivotal target for effective therapy. Semin Cancer Biol. 2009;19:17‐24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Tsuji A, Wakisaka N, Kondo S, Murono S, Furukawa M, Yoshizaki T. Induction of receptor for advanced glycation end products by EBV latent membrane protein 1 and its correlation with angiogenesis and cervical lymph node metastasis in nasopharyngeal carcinoma. Clin Cancer Res. 2008;14:5368‐5375. [DOI] [PubMed] [Google Scholar]
- 38. Schorey JS, Bhatnagar S. Exosome function: from tumor immunology to pathogen biology. Traffic. 2008;9:871‐881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Simons M, Raposo G. Exosomes–vesicular carriers for intercellular communication. Curr Opin Cell Biol. 2009;21:575‐581. [DOI] [PubMed] [Google Scholar]
- 40. Hurwitz SN, Nkosi D, Conlon MM, et al. CD63 regulates Epstein‐Barr virus LMP1 exosomal packaging, enhancement of vesicle production, and noncanonical NF‐κB signaling. J Virol. 2017;91:e02251–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Meckes DG Jr, Shair KH, Marquitz AR, Kung CP, Edwards RH, Raab‐Traub N. Human tumor virus utilizes exosomes for intercellular communication. Proc Natl Acad Sci USA. 2010;107:20370‐20375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Flanagan J, Middeldorp J, Sculley T. Localization of the Epstein‐Barr virus protein LMP 1 to exosomes. J Gen Virol. 2003;84:1871‐1879. [DOI] [PubMed] [Google Scholar]
- 43. Dukers DF, Meij P, Vervoort MB, et al. Middeldorp, Direct immunosuppressive effects of EBV‐encoded latent membrane protein 1. J Immunol. 2000;165:663‐670. [DOI] [PubMed] [Google Scholar]
- 44. Middeldorp JM, Pegtel DM. Multiple roles of LMP1 in Epstein‐Barr virus induced immune escape. Semin Cancer Biol. 2008;18:388‐396. [DOI] [PubMed] [Google Scholar]
- 45. Staudt LM. Oncogenic activation of NF‐kappaB. Cold Spring Harb Perspect Biol. 2010;2:a000109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Aga M, Bentz GL, Raffa S, et al. Exosomal HIF1α supports invasive potential of nasopharyngeal carcinoma‐associated LMP1‐positive exosomes. Oncogene. 2014;33:4613‐4622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Petersson F. Nasopharyngeal carcinoma: a review. Semin Diagn Pathol. 2015;32:54‐73. [DOI] [PubMed] [Google Scholar]
- 48. Yao M, Ohshima K, Suzumiya J, Kume T, Shiroshita T, Kikuchi M. Interleukin‐10 expression and cytotoxic‐T‐cell response in Epstein‐Barr‐virus‐associated nasopharyngeal carcinoma. Int J Cancer. 1997;72:398‐402. [DOI] [PubMed] [Google Scholar]
- 49. Li J, Zhang XS, Xie D, et al. Expression of immune‐related molecules in primary EBV‐positive Chinese nasopharyngeal carcinoma: associated with latent membrane protein 1 (LMP1) expression. Cancer Biol Ther. 2007;6:1997‐2004. [DOI] [PubMed] [Google Scholar]
- 50. Gourzones C, Barjon C, Busson P. Host‐tumor interactions in nasopharyngeal carcinomas. Semin Cancer Biol. 2012;22:127‐136. [DOI] [PubMed] [Google Scholar]
- 51. Clausse B, Fizazi K, Walczak V, et al. High concentration of the EBV latent membrane protein 1 in glycosphingolipid‐rich complexes from both epithelial and lymphoid cells. Virolgy. 1997;228:285‐293. [DOI] [PubMed] [Google Scholar]
- 52. Smith C, Wakisaka N, Crough T, et al. Discerning regulation of cis‐ and trans‐presentation of CD8+ T‐cell epitopes by EBV‐encoded oncogene LMP‐1 through self‐aggregation. Blood. 2009;113:6148‐6152. [DOI] [PubMed] [Google Scholar]
- 53. Zhang J, Fang W, Qin T, et al. Co‐expression of PD‐1 and PD‐L1 predicts poor outcome in nasopharyngeal carcinoma. Med Oncol. 2015;32:86. [DOI] [PubMed] [Google Scholar]
- 54. Hsu MC, Hsiao JR, Chang KC, et al. Increase of programmed death‐1‐expressing intratumoral CD8 T cells predicts a poor prognosis for nasopharyngeal carcinoma. Mod Pathol. 2010;23:1393‐1403. [DOI] [PubMed] [Google Scholar]
- 55. Badoual C, Hans S, Merillon N, et al. PD‐1‐expressing tumor‐infiltrating T cells are a favorable prognostic biomarker in HPV‐associated head and neck cancer. Cancer Res. 2013;73:128‐138. [DOI] [PubMed] [Google Scholar]
- 56. Derks S, Liao X, Chiaravalli AM, et al. Abundant PD‐L1 expression in Epstein‐Barr virus‐infected gastric cancers. Oncotarget. 2016;7:32925‐32932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Fang W, Zhang J, Hong S, et al. EBV‐driven LMP1 and IFN‐γ up‐regulate PD‐L1 in nasopharyngeal carcinoma: implications for oncotargeted therapy. Oncotarget. 2014;5:12189‐12202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Cai TT, Ye SB, Liu YN, et al. LMP1‐mediated glycolysis induces myeloid‐derived suppressor cell expansion in nasopharyngeal carcinoma. PLoS Pathog. 2017;13:e1006503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Yoshizaki T, Takimoto T, Takeshita H, et al. Epstein‐Barr virus lytic cycle spreads via cell fusion in a nasopharyngeal carcinoma hybrid cell line. Laryngoscope. 1994;104:91‐94. [DOI] [PubMed] [Google Scholar]
- 60. Dittmer DP, Hilscher CJ, Gulley ML, Yang EV, Chen M, Glaser R. Multiple pathways for Epstein‐Barr virus episome loss from nasopharyngeal carcinoma. Int J Cancer. 2008;123:2105‐2112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Nanbo A, Sugden A, Sugden B. The coupling of synthesis and partitioning of EBV's plasmid replicon is revealed in live cells. EMBO J. 2007;26:252‐262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Dawson CW, Rickinson AB, Young LS. Epstein‐Barr virus latent membrane protein inhibits human epithelial cell differentiation. Nature. 1990;344:777‐780. [DOI] [PubMed] [Google Scholar]
- 63. Kulwichit W, Edwards RH, Devenport EM, Baskar JF, Godfrey V, Raab‐Traub N. Expression of the Epstein‐Barr virus latent membrane protein 1 induces B cell lymphoma in transgenic mice. Proc Natl Acad Sci USA. 1998;95:11963‐11968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Yoshizaki T, Ito M, Murono S, Wakisaka N, Kondo S, Endo K. Current understanding and management of nasopharyngeal carcinoma. Auris Nasus Larynx. 2012;39:137‐144. [DOI] [PubMed] [Google Scholar]
- 65. Ooft ML, van Ipenburg JA, Braunius WW, Zuur CI, Koljenović S, Willems SM. Prognostic role of tumor infiltrating lymphocytes in EBV positive and EBV negative nasopharyngeal carcinoma. Oral Oncol. 2017;71:16‐25. [DOI] [PubMed] [Google Scholar]
- 66. Zhang G, Tsang CM, Deng W, et al. Enhanced IL‐6/IL‐6R signaling promotes growth and malignant properties in EBV‐infected premalignant and cancerous nasopharyngeal epithelial cells. PLoS One. 2013;8:e62284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Hannigan A, Qureshi AM, Nixon C, et al. Lymphocyte deficiency limits Epstein‐Barr virus latent membrane protein 1 induced chronic inflammation and carcinogenic pathology in vivo. Mol Cancer. 2011;10:11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Nakanishi Y, Wakisaka N, Kondo S, et al. Progression of understanding for the role of Epstein‐Barr virus and management of nasopharyngeal carcinoma. Cancer Metastasis Rev. 2017;36:435‐447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Ang KK, Harris J, Wheeler R, et al. Human papillomavirus and survival of patients with oropharyngeal cancer. N Engl J Med. 2010;363:24‐35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Kitagawa N, Goto M, Kurozumi K, et al. Epstein‐Barr virus‐encoded poly(A)(‐) RNA supports Burkitt's lymphoma growth through interleukin‐10 induction. EMBO J. 2000;19:6742‐6750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Duan Y, Li Z, Cheng S, et al. Nasopharyngeal carcinoma progression is mediated by EBER‐triggered inflammation via the RIG‐I pathway. Cancer Lett. 2015;361:67‐74. [DOI] [PubMed] [Google Scholar]
- 72. Kanda T, Miyata M, Kano M, Kondo S, Yoshizaki T, Iizasa H. Clustered microRNAs of the Epstein‐Barr virus cooperatively downregulate an epithelial cell‐specific metastasis suppressor. J Virol. 2015;89:2684‐2697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Harabuchi Y, Takahara M, Kishibe K, et al. Nasal natural killer (NK)/T‐cell lymphoma: clinical, histological, virological, and genetic features. Int J Clin Oncol. 2009;14:181‐190. [DOI] [PubMed] [Google Scholar]
- 74. Komabayashi Y, Kishibe K, Nagato T, Ueda S, Takahara M, Harabuchi Y. Downregulation of miR‐15a due to LMP1 promotes cell proliferation and predicts poor prognosis in nasal NK/T‐cell lymphoma. Am J Hematol. 2014;89:25‐33. [DOI] [PubMed] [Google Scholar]
- 75. Chen YP, Zhang WN, Chen L, et al. Effect of latent membrane protein 1 expression on overall survival in Epstein‐Barr virus‐associated cancers: a literature‐based meta‐analysis. Oncotarget. 2015;6:29311‐29323. [DOI] [PMC free article] [PubMed] [Google Scholar]