

Abstract
INTRODUCTION
Exposure of urothelial carcinoma (UC) cells to Bacille Calmette Guerin (BCG) affects cellular redox status and tumor cell biology but mechanism(s) remains unclear. This study examined free radical production by BCG, and in tumor cells in response to BCG, using global profiling of Reactive oxygen species/reactive nitrogen species (ROS/RNS). The relationship between free radical generation and downstream cellular events was evaluated.
MATERIALS AND METHODS
Using fluorescent probes, global profiling of ROS/RNS was carried out in Heat killed (hk) BCG, viable BCG, and in two UC cell lines post BCG exposure (253J and T24). Inhibition of BCG internalization and pharmacologic scavenging of H2O2 was studied for their effect on cellular ROS/RNS generation and various physiological end points.
RESULTS
Viable BCG produced H2O2 (Hydrogen peroxide) and O2− (Superoxides) but did not show NO (Nitric oxide) generation. Loss of viability decreased production of H2O2 by 50% compared to viable BCG. BCG internalization was necessary for BCG induced ROS/RNS generation in UC cells. Pharmacologic H2O2 scavenging reversed the ROS/RNS mediated signaling in UC cells. BCG dependent alterations in tumor biology including intracellular signaling, gene expression and cytotoxicity were dependent on free radical generation.
CONCLUSIONS
This study demonstrates the importance of free radical generation by BCG, and intracellular generation of Cellular oxidative stress (COS), on the UC cell response to BCG. Manipulation of the BCG induced COS represents a potential target for increasing BCG efficacy.
Keywords: Bladder cancer, BCG, H2O2, Oxidative stress, global profiling
Introduction
The intravesical administration of the attenuated Mycobacterium Bacille Calmette Guerin is standard of care treatment for high risk, non-muscle invasive bladder cancer. 1–2 Post BCG exposure, UC cells undergoes complex changes in redox status, intracellular signaling and gene expression. BCG internalization is associated with the downstream activation of multiple stress responsive pathways involving NF-κB, NrF2 and CEBP. These signaling pathways direct the transactivation of genes which subsequently contribute to alterations in tumor biology and phenotype. In addition to this direct effect on UC cells, internalization of BCG by UC cells triggers the expression of white blood cell receptors and the release of cytokines and chemokines. These effects combine to recruit and target various types of immune cells into the bladder. Activated phagocytes and cytokines leads to the differentiation of CD4+ T cells into TH1 and/or TH2 type immune response.3 Through a combination of both direct and systemic effects, the interaction between UC cell and BCG plays a critical role in the ultimate response to treatment.
Previous studies have shown that BCG, both live and lyophilized affects cellular ROS generation and oxidative stress in cancer cells.4–6 However, the precise mechanism through which BCG generates oxidative stress was not reported and the relationship between BCG induced oxidative stress and BCG dependent cellular events has not been investigated. This study evaluated ROS/RNS generation by BCG and BCG exposed tumor cells. The role of BCG internalization, BCG viability and intracellular H2O2 levels in mediating cellular changes in ROS/RNS and downstream cellular events were evaluated. Our results show that viable BCG acts as H2O2 generator. BCG internalization and generation of intracellular H2O2 is required for changes in cellular redox status, ROS/RNS production and the direct biologic response of UC cells to BCG. The identification of H2O2 and O2− production by BCG is a novel and previously unreported finding. BCG production of H2O2 and O2− contributes, if not playing a central role, to the known BCG-induced alterations in cellular redox status and the associated changes in UC cell biology.
Materials and Methods
2.1 Cell Lines
The human UC cell line T24 was obtained from American Type culture collection Culture (Rockville, MD). The 253J cell line was a kind gift of Dr. Richard Williams (University of Iowa). Cells were maintained as described earlier.7
2.2 Bacillus Calmette-Guerin (BCG)
TICE BCG and HkBCG were prepared and used as described earlier.8
2.3 Real time measurement of ROS and RNS in bacterial cells
Fluorescence probes were used for detection of ROS/RNS as described earlier.9 Coumarin 7-boronic acid (CBA) was synthesized as described earlier.9 Amplex® Red (AR) (10-acetyl-3,7-dihydroxyphenoxazine)/horseradish peroxidase were purchased from Cayman Chemical. CBA and AR were used for H2O2 detection. Hydroethidine (HE) was purchased from Invitrogen for O2− detection. Diaminofluorescein–2 diacetate (DAF-2DA) was purchased from EMD calbiochem for NO detection. For detection of ROS/RNS, equal number of reconstituted BCG and hkBCG were suspended in sterile DPBS. For measurements using the fluorescence plate reader, immediately after the addition of DPBS containing the probe CBA (25 μM), AR (50 μM), HE (40 μM) and DAF-2DA (10 μM), the plate with cells was placed in the plate reader pre warmed to 37°C and was read for 1h.
2.4 Fluorescent plate reader
Total fluorescence intensities were obtained using Beckman Coulter DTX-880 plate reader. The instrument was kept at 37°C during the measurements and fluorescence intensity read from bottom of each well was integrated over 0.5 s every 90 s. Fluorescence signal was detected at an excitation/emission wavelength for different probes as followed: 320/430 nm for CBA, 535/595 nm for AR, 485/574 nm fro HE, 485/535 nm for DAF-2DA.
2.5 Real Time global profiling of ROS/RNS in UC cells
For long term global profiling, 253J and T24 cells were plated at 1 × 105 cells/well in 24-well plates. 24 h later, BCG (50:1 BCG: cells) was added. Cells were incubated for 1, 3 and 6 days and washed twice with sterile PBS. Intracellular ROS/RNS levels were measured using fluorescence probes as described earlier. To measure ROS/RNS at earlier time points, cells were plated at 1 × 104 cells/well in 96-well plates. 24h later, BCG (50:1 BCG: cells) was added to the cultures. Cells were incubated up to 24h and washed twice with sterile PBS. Intracellular ROS/RNS levels were measured using probes as described earlier. A corresponding control (no BCG) was included at each time point and results were presented as difference (Δ) between corresponding “time correct” control and BCG treated group.
2.6 Effect BCG internalization on global profiling of ROS/RNS in UC cells
Inhibition of the cellular cytoskeleton using the spindle toxin cytochalasin b prevents BCG internalization.10 To investigate the role of BCG internalization on ROS/RNS generation; cells were plated at 1 × 104 cells/well in 96-well plates. 24h later, cells were pretreated with cytochalasin b (25 μg/ml) for 1h before BCG addition. The cells were incubated for 6h and washed twice with sterile PBS. Intracellular ROS/RNS levels were measured using fluorescence probes as described earlier.
2.7 Effect of ROS scavenger on global profiling of ROS/RNS in UC cells
Ebselen (2-phenyl-1, 2-benzisoselenazol-3(2H)-one) is a potent, cell permeable scavenger of H2O2.11 It functions as a mimic of glutathione peroxidase. To check effect of ebselen on ROS/RNS profiling, cells were plated at 1 × 104 cells/well in 96-well plate. 24h later, cells were pre-treated with ebselen (25 μM) for 1h followed by BCG addition. The cells were incubated for 6h and washed twice with sterile PBS. Intracellular ROS/RNS levels were measured using fluorescence probes as described earlier.
2.8 Effect of ROS scavenger on gene expression
Prior work has demonstrated that UC cell exposure to BCG increases the expression of cell cycle regulatory and immune response genes.12 The effect of ROS scavenger ebselen on gene expression in response to BCG was measured after 6 h using qRT-PCR and primers as described earlier.12
2.9 Effect of ROS scavenger on Intracellular signaling
The effect of ROS scavenger on signaling pathway activation was measured as described earlier.8 Cells were untreated or treated with BCG, ebselen and BCG with ebselen for 6h. Luciferase activity was measured using a luciferase assay system (Promega, Madison, WI) according to the manufacturer’s instructions. Luciferase activities were normalized to protein concentration measured by BCA protein assay kit (Pierce, Rockford, IL).
2.10 LDH release Assay
The effect of ROS scavenger on LDH release in response to BCG after 12h was measured by following manufacturer’s instruction (CytoTox 96® Non-Radioactive Cytotoxicity Assay Kit, Promega, Madison, WI).
2.11 Cell Proliferation
BCG is known to exert an antiproliferative effect on UC cells. The effect of ROS scavenger on BCG’s anti-proliferative effect was measured using the MTT assay as previously described.12. Cytotoxic effects of BCG were also confirmed by using Apo-tox-glo kit (Promega) and results were comparable with MTT.
2.12 Statistical Analysis
All experiments were performed in triplicate at three different time points. qRT PCR data were normalized using housekeeping gene β-actin expression. Data was analyzed using non parametric analysis and ANOVA using graphpad prism 6. Results were considered significant at p < 0.05. Graphical representation of the data is shown as the mean ± S.D.
Results
3.1 ROS/RNS generation by BCG
ROS/RNS production by BCG was monitored using fluorogenic probes. BCG showed H2O2 and O2− production as measured by increase in the fluorescent intensity but did not show NO production (figure 1). Compared to reconstituted BCG, hk BCG showed a 50% decrease in H2O2 generation (p < 0.001, figure 1). There was no significant difference in O2− generation by reconstituted and hk BCG (p = 0.4).
Figure 1.
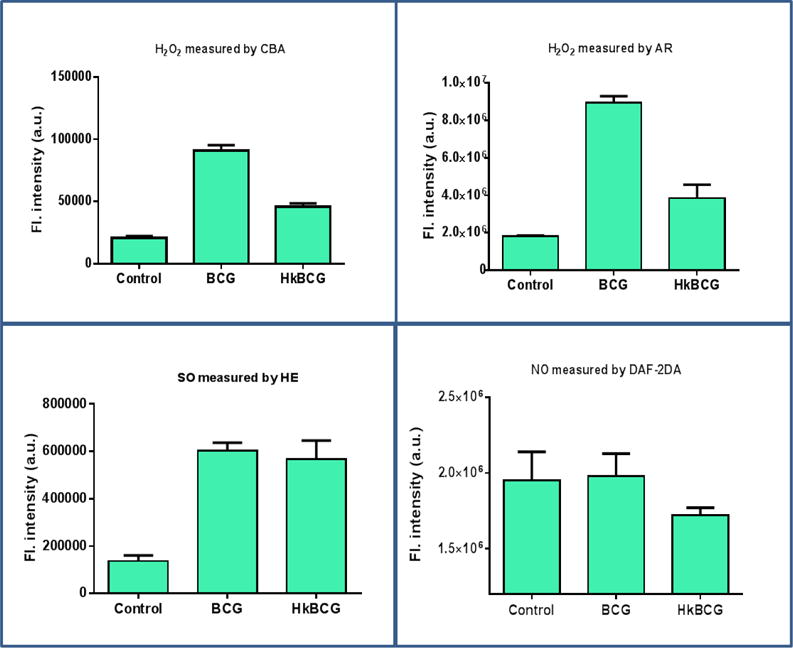
Global profiling of ROS/RNS generation by BCG. ROS/RNS generation was measured using different fluorescent probes a) H2O2 was measured by CBA and AR b) Superoxides were measured by HE c) NO was measured by DAF-2DA. BCG produces H2O2 and O2− as but do not generate NO. H2O2 production by BCG is affected by viability. Hk BCG produced decreased amount of H2O2 compared to freshly reconstituted BCG (p < 0.001). O2− production was not affected by viability as there was no significant difference in viable and hkBCG group (P = 0.1)
3.2 Intracellular real time global profiling of ROS/RNS
UC cells were treated with BCG up to 6 days. Real time global profiling of ROS/RNS generation was measured at 1, 3, 6, 12, and 18h and on days 1, 3 and 6. No significant changes in cellular ROS/RNS were observed beyond 24h (data not shown). When measured using Amplex red fluorogenic dye, both cell lines showed significant early increases in H2O2 levels. In T24 cells, the increase in H2O2 could be detected by 3h (p < 0.001) and remained significantly higher till 12h relative to controls (p < 0.001, Figure 2). In 253J cells, a significant increase in H2O2 was observed from 3h (p < 0.05) and significantly elevated levels were observed till 18h post BCG exposure (p < 0.01, Figure 2).
Figure 2.
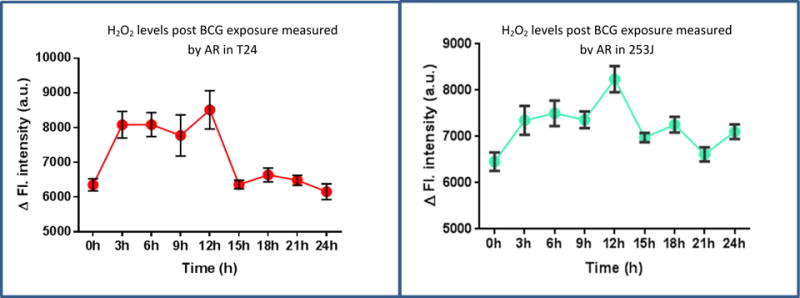
Real time global profiling of ROS post BCG exposure: T24 and 253J cell lines were exposed to BCG and H2O2 was measured using AR at different time points post BCG exposure. Exposure of UC cells to BCG significantly increased the levels of H2O2 generation. The increase could be detected from 3h till 12 h (p < 0.001) in T24. In 253J, increase in H2O2 could be detected from 3h onwards till 18h post BCG exposure h (p < 0.01). Values on Y axis show the difference between “time correct” control and BCG treated cells.
NO levels were measured using the DAF-2DA probe (Figure 3). BCG exposure resulted in higher levels of NO at 3h with a significant increase observed at 6h in both cell lines (p < 0.01) relative to untreated controls. No significant difference in NO levels between control and treated group were observed 6h onwards (T24, P = 0.57; 253J, p = 0.28).
Figure 3.
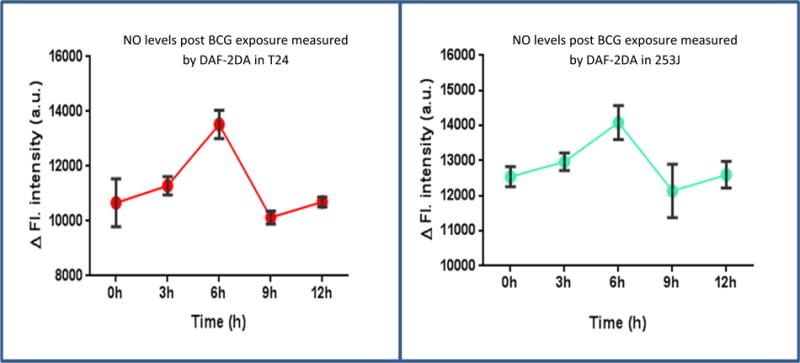
Effect of BCG on real time global profiling of RNS: T24 and 253J cell lines were exposed to BCG and NO levels were measured using DAF-2DA at different time points post BCG exposure. NO levels starts peaking up at 3h and reach to significantly higher levels at 6h in both T24 (p < 0.01) and 253J (p < 0.01) cell lines compared to control. No significant difference in NO levels between control and treated group were observed 6h onwards (T24, P = 0.57; 253J, p = 0.28). Values on Y axis show the difference between “time correct” control and BCG treated cells.
3.3 Effect of BCG internalization on real time ROS/RNS profiling
Figure 4 shows real time global profiling of ROS/RNS generation in control cells or cells treated with BCG or cytochalasin b and BCG. BCG treated cells demonstrated significantly increased levels of H2O2, O2− and NO compared to untreated controls. Inhibition of BCG internalization using cytochalasin b resulted in significant decreases in ROS/RNS levels. Relative to BCG alone, cytochalasin b with BCG treatment groups significantly decreased H2O2 (p < 0.05). O2− levels were significantly reduced in the cytochalasin b plus BCG treated group (p < 0.01), in both cell lines. NO levels in cytochalasin b plus BCG group showed significantly lower levels compared to BCG alone (p < 0.05). ROS/RNS generation profile in the cytochalasin b plus BCG group did not significantly differ from that in the corresponding cytochalasin b alone group (p = 0.4).
Figure 4.
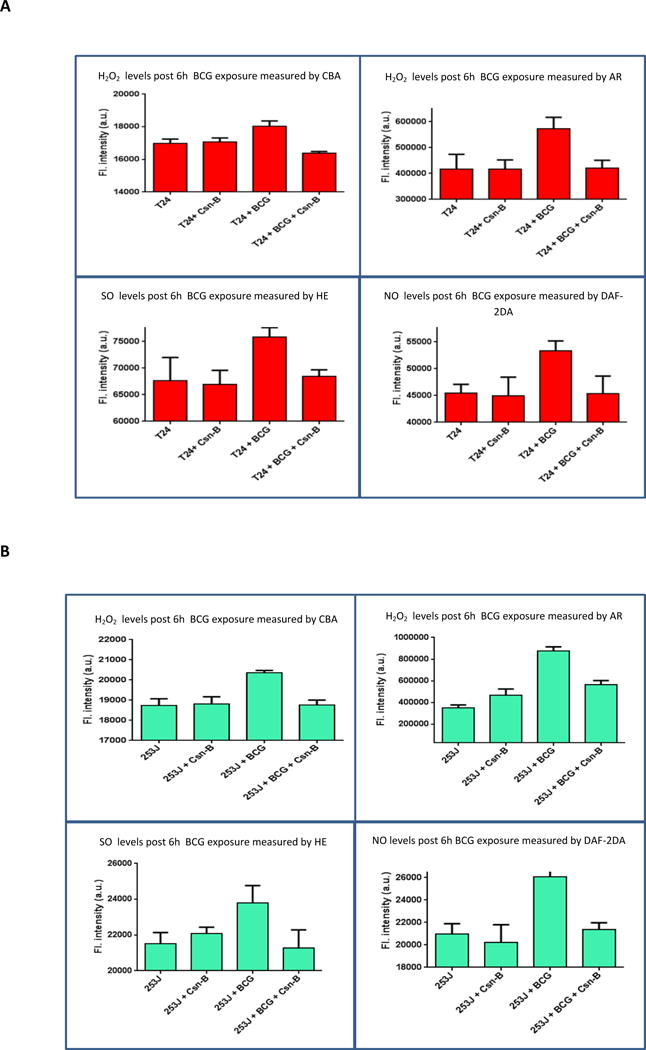
Effect of BCG internalization on real time ROS/RNS profiling: T24 and 253J cells were exposed to BCG for 6h. Post BCG exposure, H2O2 levels were measured using CBA and AR, O2− levels were measured using HE and NO levels were measured using DAF-2DA. Treatment groups included Untreated, Cytochalasin-B treated, BCG treated and BCG with cytochalasin-B. H2O2, O2− and NO levels significantly decreased in the BCG with cytochalasin-b treatment group in both cell lines, relative to BCG alone. (p < 0.05 for H2O2, p < 0.01 for O2− and p < 0.05 for NO). The difference in Cytochalasin-b control group and cytochalasin-b plus BCG group was statistically non-significant (p = 0.4).
3.4 Effect of free radical scavenger on real time ROS/RNS global profiling
Pretreatment of UC cells with the cell permeable H2O2 scavenger ebselen prior to BCG treatment significantly decreased ROS/RNS generation by UC cells. Figure 5 shows intracellular real time ROS/RNS levels in different treatment groups. H2O2, O2− and NO levels significantly decreased in the BCG plus ebselen treatment group in both cell lines, relative to BCG alone. (p < 0.05 for H2O2, p < 0.001 for O2− and NO). ROS/RNS generation profile in the ebselen plus BCG group did not significantly differ from that in the corresponding ebselen alone group (p = 0.7).
Figure 5.
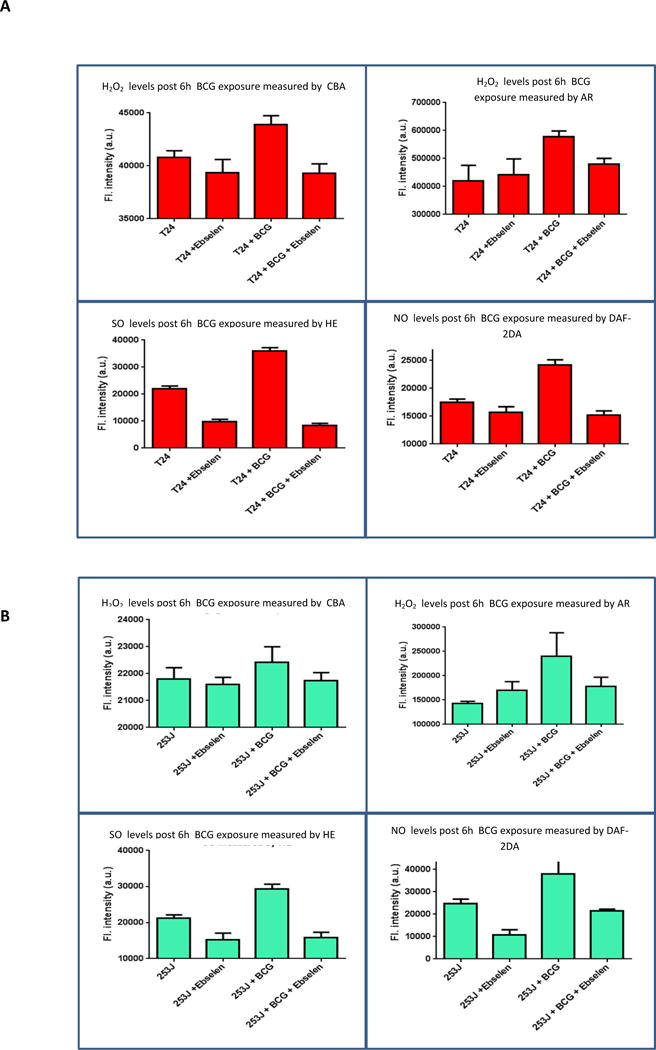
Effect of ebselen on real time ROS/RNS global profiling: T24 and 253J cells were exposed to BCG for 6h. Post BCG exposure. H2O2 levels were measured using CBA and AR, O2− levels were measured using HE and NO levels were measured using DAF-2DA. Treatment groups included Untreated, ebselen treated, BCG treated and BCG with ebselen. Compared to the BCG treated group, BCG with ebselen treated group had significantly reduced levels of ROS/RNS. (p < 0.05 for H2O2, p < 0.001 for O2− and NO). The difference in ebselen control group and ebselen plus BCG group was statistically non-significant (p = 0.7).
3.5 Effect of free radical scavenger on Gene Expression
Ebselen significantly decreased the response of UC cells to BCG as measured gene expression of several immune response genes. Figure 6 demonstrates the effect of ebselen on the UC cell expression of a panel of 8 BCG responsive genes as measured by q-rtPCR. Ebselen in combination with BCG reduced UC cell expression of all measured genes with 7 of 8 and 8 of 8 reaching the level of statistical significance in 253J and T24 cells respectively (p < 0.05 increase vs. control, p < 0.05 decrease vs. BCG).
Figure 6.
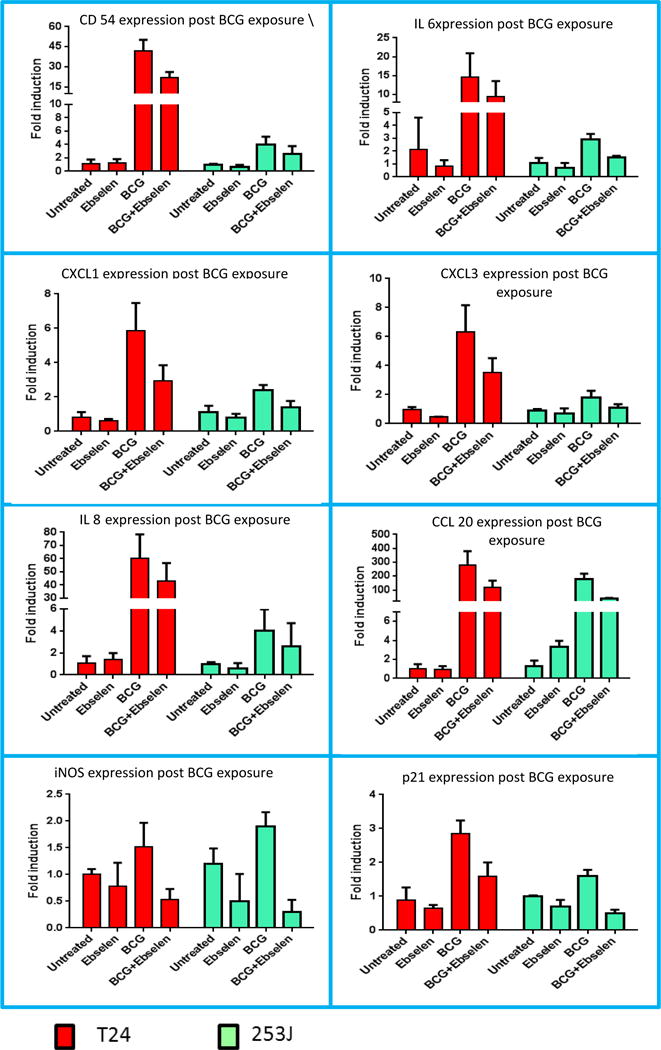
Effect of free radical scavenger on Gene Expression: T24 and 253J cells were exposed to BCG. Post BCG exposure, gene expression of various genes was measured using q-RT PCR. Treatment groups included untreated, ebselen treated, BCG treated and BCG with ebselen. BCG exposure resulted in significant increased the expression of 8 immune response genes as measured by qRTPCR in both cell lines (ANOVA, p < 0.05). Ebselen in combination with BCG reduced UC cell expression of all measured genes with 7 of 8 and 6 of 8 reaching the level of statistical significance in 253J and T24 cells respectively (ANOVA, P < 0.05).
3.6: Effect of free radical scavenger on intracellular signaling
Exposure of UC cells to BCG for 6 h resulted in the activation of the NF-κB, CEBP and NrF2 intracellular signaling pathways. BCG significantly increased activation of NF-κB, CEBP and NRF2 luciferase reporter construct in both cell lines (p < 0.01). Relative to BCG, pretreatment of cells with the ebselen significantly reduced NF-κB (p < 0.01), CEBP (p < 0.05) and NrF2 (p < 0.05) reporter activation. NF-κB, CEBP and NRF2 activation in response to BCG with ebselen treated cells was not significantly different than cells treated with ebselen alone (Figure 7).
Figure 7.
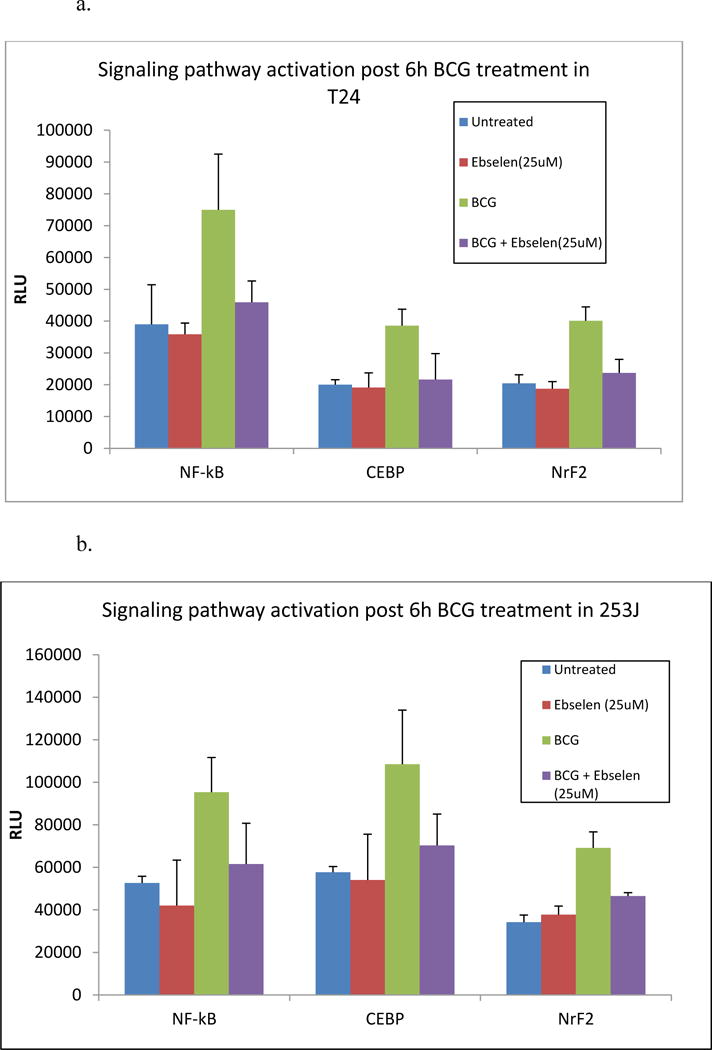
Effect of ebselen on signaling pathways: T24 and 253J cells were exposed to BCG for 6h. Treatment groups included Untreated, ebselen treated, BCG treated and BCG with ebselen. BCG exposure significantly increased activation of NF-κB, CEBP and NrF2 luciferase reporter construct in both cell lines (p < 0.01). Ebselen in combination with BCG significantly reduced NF-κB (p < 0.01), CEBP (p < 0.05) and NrF2 (p < 0.05) reporter activation.
3.7 Effect of free radical scavenger on BCG cytotoxicity
Exposure of UC cells to BCG significantly increased LDH release compared to controls in T24 and 253J cells, respectively (p < 0.001; figure 8a). Pretreatment with ebselen significantly decreased LDH release relative to BCG alone in both cell lines after 12 h (T24 p < 0.001; 253J p < 0.01). Of note, ebselen alone resulted in an increase in LDH relative to control cells. Relative to ebeslen alone, LDH release was significantly increased in the BCG + ebselen group (p < 0.05).
Figure 8.
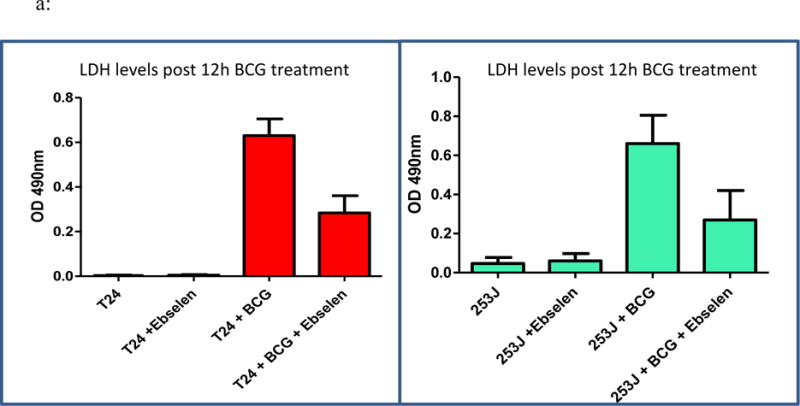
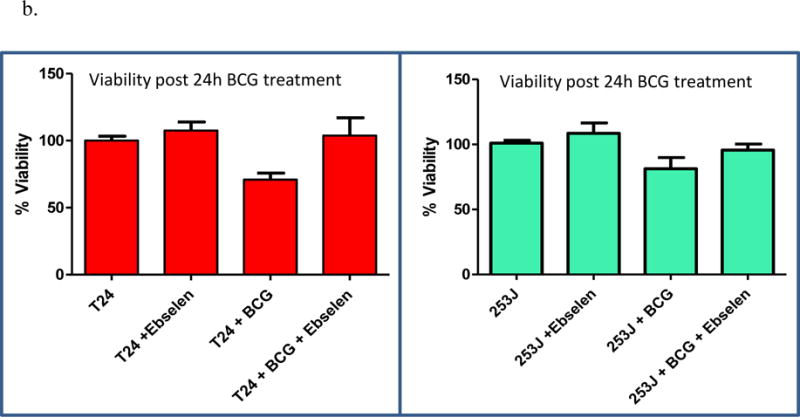
a: Effect of ebselen on BCG cytotoxicity: T24 and 253J cells were exposed to BCG for 12 or 24h. Treatment groups included Untreated, ebselen treated, BCG treated and BCG with ebselen. BCG exposure significantly increased LDH release in T24 and 253J cells (p < 0.001). Ebselen pretreatment significantly decreased LDH release relative to BCG alone in both cell lines after 12 h (T24 p < 0.001; 253J p < 0.01). b: BCG exposure for 24 h significantly inhibited cell proliferation in 253J and T24 cells, respectively (T24 p < 0.005; 253J p < 0.05). Treatment of cells with ebselen prior to BCG treatment inhibited BCG’s antiproliferative/cytotoxic effect in 253J and T24 cells (p < 0.05).
As measured by the MTT assay, exposure of UC cells to BCG for 24 h significantly inhibited cell proliferation compared to controls in 253J and T24 cells, respectively (T24 p < 0.005; 253J p < 0.05). Treatment of cells with ebselen prior to BCG treatment inhibited BCG’s antiproliferative/cytotoxic effect in 253J and T24 cells (p < 0.05, figure 8b).
Discussion
Multiple publications have reported on the direct effects of BCG on the biology of UC cells. BCG adherence to and internalization by UC cells, activation of cellular stress responsive signaling pathways, changes in gene expression and alterations in cellular viability and phenotype characterize the response of UC cells to BCG.13–15 iNOS activation and NO production in response to BCG are required for downstream cellular events.16–18 Loss of BCG viability is associated with a reduction in BCG’s direct effects.19–20 While important observations the mechanism through which BCG initiates these changes and how this is correlated with BCG viability, has here-to-fore been undefined.
In what to our knowledge is a novel finding, the current study demonstrates that BCG acts as an H2O2 generator. H2O2 is member of Reactive oxygen species (ROS) molecules which also includes super oxides and hydroxyl radicals. While H2O2 is a signaling molecule, it can also cause significant cellular damage particularly after reacting with reduced iron or copper to generate hydroxyl radicals which are strong oxidizing agents.21 H2O2 production has been reported in many bacterial species including E.coli, Streptoccous spp., Pneumococcus spp., lactic acid bacteria etc.22–25 While H2O2 is continuously produced in aerobically growing cells as by product of respiration, the main source of hydrogen peroxide production in bacteria is still unclear. In case of E.coli, respiratory enzymes were suggested to be source but recently it has been reported that it may be formed by a non-respiratory enzyme or by a cellular metabolite which reacts with oxygen. 22, 26 In case of Streptococcus sanguinis, pyruvate oxidase and acetate kinase enzymes have been shown to affect H2O2 production.24 In case of Streptococcus pyogens, lactate oxidase was reported to be involved.24
It has been previously reported that BCG exposure results in oxidative stress and lipid peroxidation in UC cells.6 However, the exact mechanism or the source of the oxidative stress was not studied in these prior publications. Based upon the results of this study H2O2 production by viable BCG appears to serve as a trigger for cellular oxidative stress and cell damage. Post BCG exposure, UC cells showed increased levels of H2O2 till 12-18h. This increase is dependent upon the internalization of viable BCG and the “effective” (non-scavenged) intracellular release of H2O2 by BCG. Increased levels of H2O2 could set the stage for secondary generation of oxidants involving iNOS, NO and peroxynitrite. Exposure of cells to H2O2 can up regulate iNOS expression and NO production.27, 28 Several studies have shown that increased iNOS/NO levels in carcinoma cells and laboratory models following intravesical instillation of BCG.18,19 Prior reports by our group have demonstrated that BCG activates UC cell iNOS expression and NO generation in vitro.13 The direct effects of BCG on UC biology were dependent upon iNOS expression and NO generation. In the current study intracellular NO levels were temporally related to cellular H2O2 production. Along with bacteria induced oxidative stress, cancer cells may also contribute to oxidative stress in response to bacterial infection 29. It has been postulated that H2O2 production by BCG together with eukaryotic NO production can promote peroxynitrite generation leading to further cellular oxidative stress.30 Oxidative stress also activates signaling pathways involving NF-κB, NrF2 and CEBP, which results in expression of several immune response genes. Previously we have shown that the inhibition of BCG internalization prevents BCG induced UC cell gene expression. Treatment with ebselen which acts as free radical scavenger for H2O2, nullified the increase in expression of immune response genes, signaling pathway activation and cytotoxicity, further indicating that the free radical generation post BCG internalization serves as a source of cellular oxidative stress. The relative decrease in H2O2 generation by non-viable BCG may explain the loss of therapeutic efficacy associated with the loss of BCG viability. In combination, these findings provide strong evidence that the intracellular generation of H2O2 by internalized, viable BCG, is a requisite and early event in initiating the UC response to BCG in vitro. It is possible that in the in vivo settings, there are additional contributors to cellular oxidative stress (COS), such as activated immune cells are known to produce ROS. Immune cells migrating to sites of BCG therapy may contribute to in vivo COS.
H2O2 generation by internalized, viable BCG is a potential unifying element in BCG induced oxidative stress and downstream, BCG dependent alterations in tumor biology.
Conclusions
This study demonstrates the importance of free radical generation by BCG, and intracellular generation of Cellular oxidative stress (COS), on the UC cell response to BCG. Pharmacological manipulation of COS in combination with viable or hkBCG provides an alternative strategy for treatment of NMIBC.
Acknowledgments
This work was supported by a grant from the Department of Veterans Affairs and the Milwaukee Veterans Affairs Medical Center. We would like to thank Daniel Eastwood, MS, Division of Biostatistics, Medical College of Wisconsin for his help in statistical analysis of the data.
Abbreviations
- ANOVA
analysis of variance
- BCG
Bacillus Calmette Guérin
- hkBCG
Heat killed BCG
- CD54
intercellular adhesion molecule 1
- CXCL1
chemokine (C-X-C motif) ligand 1
- CXCL3
chemokine (C-X-C motif) ligand 3
- CCL20
chemokine (C-C motif) ligand 20
- DNA
deoxyribonucleic acid
- FBS
fetal bovine serum
- IL6
interleukin 6
- IL8
interleukin 8
- iNOS
inducible nitric oxide synthase
- mRNA
messenger ribonucleic acid
- NF-κB
nuclear factor kappa B
- NO
nitric oxide
- NRF2
nuclear factor (erythroid-derived 2)-like 2
- CEBP
CCAAT-enhancer-binding proteins
- P21
cyclin dependent kinase inhibitor p21 (cip1; waf1)
- PBS
phosphate buffered saline
- RNA
ribonucleic acid
- rtPCR
reverse transcriptase polymerase chain reaction
- q rtPCR
quantitative reverse transcriptase polymerase chain reaction
- UC
urothelial carcinoma
- ROS
reactive oxygen species
- RNS
reactive nitrogen species
- H2O2
hydrogen peroxide
- O2−
superoxides
- CBA
Coumarin 7-boronic acid
- HE
Hydorethidine
- AR
amplex red; 10-acetyl-3,7-dihydroxyphenoxazine
- DAF-2DA
Diaminofluorescein–2 diacetate
- COS
cellular oxidative stress
References
- 1.Babjuk M, Oosterlinck W, Sylvester R, et al. EAU guidelines on non-muscle-invasive urothelial carcinoma of the bladder, the 2011 update. Eur Urol. 2011;59:997. doi: 10.1016/j.eururo.2011.03.017. [DOI] [PubMed] [Google Scholar]
- 2.Kassouf W, Kamat AM, Zlotta A, et al. Canadian guidelines for treatment of non-muscle invasive bladder cancer: a focus on intravesical therapy. Can Urol Assoc J. 2010;4:168. doi: 10.5489/cuaj.10051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Luo Y, Askeland EJ, Newton M, et al. Immunotherapy of Urinary Bladder Carcinoma: BCG and Beyond. Advances in Urology. 2012:181987. doi: 10.1155/2012/181987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Pook SH, Esuvaranathan K, Mahendran R. N-acetylcysteine augments the cellular redox changes and cytotoxic activity of internalized Mycobacterium bovis in human bladder cancer cells. J Urol. 2002;168:780. [PubMed] [Google Scholar]
- 5.Méndez-Samperio P, Pérez A, Torres L. Role of reactive oxygen species (ROS) in Mycobacterium bovis Bacillus Calmette-Guerin mediated up-regulation of the human cathelicidin LL-37 in A549 cells. Microb Pathog. 2009;47:252. doi: 10.1016/j.micpath.2009.08.006. [DOI] [PubMed] [Google Scholar]
- 6.Rahmat JN, Esuvaranathan K, Mahendran R. Bacillus Calmette-Guérin induces cellular reactive oxygen species and lipid peroxidation in cancer cells. Urology. 2012;79:1411.e15. doi: 10.1016/j.urology.2012.01.017. [DOI] [PubMed] [Google Scholar]
- 7.Zhang G, Chen F, Cao Y, et al. Contributors to HMGB1 Release by Urothelial Carcinoma Cells in Response to BCG. J Urol. 2013 Apr 11; doi: 10.1016/j.juro.2013.03.123. pii: S0022-5347(13)04055-X. [DOI] [PubMed] [Google Scholar]
- 8.Shah G, Zhang G, Chen F, et al. Loss of BCG Viability Adversely Affects the Direct Response of Urothelial Carcinoma Cells to BCG Exposure. J Urol. 2014;191:823. doi: 10.1016/j.juro.2013.09.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zielonka J, Zielonka M, Sikora A, et al. Global profiling of reactive oxygen and nitrogen species in biological systems: high-throughput real-time analyses. J Biol Chem. 2012;287:2984. doi: 10.1074/jbc.M111.309062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Luo Y, Szilvasi A, Chen X, et al. A novel method for monitoring Mycobacterium bovis BCG trafficking with recombinant BCG expressing green fluorescent protein. Clin Diagn Lab Immunol. 1996;3:761. doi: 10.1128/cdli.3.6.761-768.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Schewe T. Molecular actions of ebselen- an anti-inflammatory antioxidant. Gen Pharmacol. 1995;26:1153. doi: 10.1016/0306-3623(95)00003-j. [DOI] [PubMed] [Google Scholar]
- 12.Chen F, Zhang G, Cao Y, et al. A Synthetic Polyvalent Ligand for α5β1 Integrin Activates Components of the Urothelial Carcinoma Cell Response to Bacillus Calmette-Guérin. J Urol. 2013;189:1104. doi: 10.1016/j.juro.2012.08.218. [DOI] [PubMed] [Google Scholar]
- 13.Bevers RF, Kurth KH, Schamhart DH. Role of urothelial cells in BCG immunotherapyfor superficial bladder cancer. Br J Cancer. 2004;91:607. doi: 10.1038/sj.bjc.6602026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Pook SH, Rahmat JN, Esuvaranathan K, et al. Internalization of Mycobacterium bovis, Bacillus Calmette Guerin, by bladder cancer cells is cytotoxic. Oncol Rep. 2007;18:1315. [PubMed] [Google Scholar]
- 15.Chen FH, Crist SA, Zhang GJ, et al. Interleukin-6 production by human bladder tumor cell lines is up-regulated by bacillus Calmette-Guérin through nuclear factor-kappaB and Ap-1 via an immediate early pathway. J Urol. 2002;168:786. [PubMed] [Google Scholar]
- 16.Mitropoulos D, Petsis D, Kyroudi-Voulgari A, et al. The effect of intravesical Bacillus Calmette- Guérin instillations on the expression of inducible nitric oxide synthase in humans. Nitric Oxide. 2005;13:36. doi: 10.1016/j.niox.2005.04.012. [DOI] [PubMed] [Google Scholar]
- 17.Hosseini A, Koskela LR, Ehren I, et al. Enhanced formation of nitric oxide in bladder carcinoma in situ and in BCG treated bladder cancer. Nitric Oxide. 2006;15:337. doi: 10.1016/j.niox.2006.01.007. [DOI] [PubMed] [Google Scholar]
- 18.Yamada H, Matsumoto S, Matsumoto T, et al. Enhancing effect of an inhibitor of nitric oxide synthesis on bacillus Calmette- Guérin-induced macrophage cytotoxicity against murine bladder cancer cell line MBT-2 in vitro. Jpn J Cancer Res. 2000;91:534. doi: 10.1111/j.1349-7006.2000.tb00978.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Shapiro A, Ratliff TL, Oakley DM, et al. Reduction of bladder tumor growth in mice treated with intravesical Bacillus Calmette-Guerin and its correlation with Bacillus Calmette-Guerin viability and natural killer cell activity. Cancer Res. 1983;43:1611. [PubMed] [Google Scholar]
- 20.Kelley DR, Ratliff TL, Catalona WJ, et al. Intravesical bacillus Calmette-Guerin therapy for superficial bladder cancer: effect of bacillus Calmette-Guerin viability on treatment results. J Urol. 1985;134:48. doi: 10.1016/s0022-5347(17)46976-x. [DOI] [PubMed] [Google Scholar]
- 21.Veal E, Day A. Hydrogen peroxide as a signaling molecule. Antioxid Redox Signal. 2011;15:147. doi: 10.1089/ars.2011.3968. [DOI] [PubMed] [Google Scholar]
- 22.Costa Seaver L, Imlay JA. Are respiratory enzymes the primary source of intracellular hydrogen peroxide. J Biol Chem. 2004;279:48742. doi: 10.1074/jbc.M408754200. [DOI] [PubMed] [Google Scholar]
- 23.Chen L, Ge X, Dou Y, et al. Identification of hydrogen peroxide production-related genes in Streptococcus sanguinis and their functional relationship with pyruvate oxidase. Microbiol. 2011;157:13. doi: 10.1099/mic.0.039669-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Seki M, Iida K, Saito M, et al. Hydrogen Peroxide Production in Streptococcus pyogenes: Involvement of Lactate Oxidase and Coupling with Aerobic Utilization of Lactate. J Bacteriol. 2004;186:2046. doi: 10.1128/JB.186.7.2046-2051.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Marty-Teysset C. Torre DL and Garel JR: Increased production of hydrogen peroxide by Lactobacillus delbrueckii subsp. Bulgaricus upon activation aeration: involvement of an NADH oxidase in oxidative stress. Appl Environ Microbiol. 2000;66:262. doi: 10.1128/aem.66.1.262-267.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gonzalez-Flecha B, Demple B. Metabolic sources of hydrogen peroxide in aerobically growing Escherichia coli. J Biol Chem. 1995;270:13681. doi: 10.1074/jbc.270.23.13681. [DOI] [PubMed] [Google Scholar]
- 27.Thomas S, Kotamraju S, Zielonka J, et al. Hydrogen peroxide induces nitric oxide and proteosome activity in endothelial cells: a bell-shaped signaling response. Free Radic Biol Med. 2007;42:1049. doi: 10.1016/j.freeradbiomed.2007.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Eguchi H, Fujiwara N, Sakiyama H, et al. Hydrogen peroxide enhances LPS-induced nitric oxide production via the expression of interferon beta in BV02 microglian cells. Neurosci Lett. 2011;494:29. doi: 10.1016/j.neulet.2011.02.047. [DOI] [PubMed] [Google Scholar]
- 29.Strickertsson JA, Desler C, Martin-Bertelsen T, et al. Enterococcus faecalis infection causes inflammation, intracellular oxphos-independent ROS production, and DNA damage in human gastric cancer cells. PLoS One. 2013 Apr 30;8(4):e63147. doi: 10.1371/journal.pone.0063147. doi: 10.1371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hoffmann O, Zweigner J, Smith SH, et al. Interplay of pneumococcal hydrogen peroxide and host-derived nitric oxide. Infect Immun. 2006;74:5058. doi: 10.1128/IAI.01932-05. [DOI] [PMC free article] [PubMed] [Google Scholar]