

Abstract
Increased interest in fullerene C60 and derivatives in recent years implies an intensification of their environmental spread. Yet, the potential risks for living organisms are largely unknown, including the interaction of C60 with fungal organisms. This may be especially relevant for mycotoxigenic fungi since C60 may both scavenge and produce reactive oxygen species (ROS), and oxidative stress induces mycotoxin production in fungi. Therefore, this study examined effects of environmentally plausible concentrations of C60 (0, 10, 50, and 100 ng/mL) on Aspergillus flavus growth and aflatoxin production in culture media. In addition, ROS-dependent oxidative stress biomarkers—thiobarbituric acid reactive substances (TBARS), reduced and oxidised glutathione ratio, superoxide dismutase isoenzymes, catalase, glutathione peroxidase, and glutathione reductase were determined in mycelia. Nanoparticles of fullerene C60 (nC60) did not exhibit strong antifungal activity against A. flavus. At the same time, nC60 caused an antiaflatoxigenic effect at 10–100 ng/mL, and 50 ng/mL unexpectedly enhanced aflatoxin production. The TBARS content, reduced and oxidised glutathione ratio, and copper, zinc superoxide dismutase activity suggest that 10 ng/mL nC60 exerted antioxidative action and reduced aflatoxin B1 production within fungal cells. Detected prooxidative effects of 50 ng/mL fullerene exceeded cellular defenses and consequently enhanced aflatoxin B1 production. Finally, the results obtained with 100 ng/mL nC60 point to prooxidative effects, but the absence of increase in aflatoxin output may indicate additional, presumably cytotoxic effects of nC60. Thus, a range of rather low levels of nC60 in the environment has a potential to modify aflatoxin production in A. flavus. Due to possible implications, further studies should test these results in environmental conditions.
Electronic supplementary material
The online version of this article (10.1186/s13568-018-0544-0) contains supplementary material, which is available to authorized users.
Keywords: Fullerene C60, Nanoparticles, Aspergillus flavus, Aflatoxin B1, Oxidative stress
Introduction
The industry of nanomaterials has started bringing fullerene C60 containing commodities to the market, although their commercial applications are still rather modest (Hotze et al. 2008). Recent applications and research of fullerene C60 and derivatives include electronics, electrochemical sensors, medicine and cosmetics, textile and food products, etc. (Michalitsch et al. 2008; Duncan 2011; Pycke et al. 2012). Nevertheless, in addition to natural and industrial combustion processes, nanotechnology-related emissions of C60 in the environment are expected to rise in the future (Farré et al. 2011; Sanchís et al. 2012, 2013, 2015) despite the alert given by European Commission (European Commission Alert 2012). This underscores the importance of understanding environmental fate and behaviour of C60, as well as connected risks. All the potential benefits of fullerene C60, which were initially proposed, are shadowed by the possibility of its negative impact on environment and living organisms. For example, the most important environmental forms of C60 are stable aggregates mainly present as nano C60 or nC60 forms in water, were shown to interact with both prokaryotic and eukaryotic organisms (Lyon et al. 2008; Pycke et al. 2012).
Literature data concerning the nC60 effect on fungi is limited. Present data suggest that fungi are quite resistant to nC60 (Cherepanova et al. 1997; Aoshima et al. 2009; Hadduck et al. 2010). A recent review has similarly concluded that negative effects of carbon nanoparticles on soil microbial communities may be expected only at levels higher than 250 mg/kg (Simonin and Richaume 2015). Caution is, however, required due to differences in studies’ design and/or nC60 preparation methods (Lyon and Alvarez 2008; Chae et al. 2010). An obvious knowledge gap exists on fullerene nC60 and mycotoxigenic fungi interaction, despite the risks of this interaction ending with either positive or negative outcomes, as outlined below.
The most prominent fullerene properties are its ROS scavenging (Krusic et al. 1991; Gharbi et al. 2005; Trpkovic et al. 2012) and producing ability, with or without photosensitisation (Yamakoshi et al. 2003; Isakovic 2006a; Usenko et al. 2008; Chae et al. 2010). Based on this fact, nC60 possesses a great potential for interaction with Aspergillus flavus, a mycotoxigenic fungus which is the main producer of aflatoxin B1—the most potent natural carcinogen (Klich 2007; Amaike and Keller 2011; Yu 2012). Oxidative stress induces aflatoxin production in A. flavus (Narasaiah et al. 2006; Reverberi et al. 2010; Roze et al. 2013), thus highlighting the nC60 potential to modulate aflatoxin synthesis. According to Trpković et al. (2012) low C60 doses demonstrate antioxidant effects in contrast to extremely high concentrations, which are highly unlikely in the environment. Sanchis et al. (2015) reported ppq levels (pg/L) of fullerenes C60 and C70 in wastewaters, surface waters and river sediments while Farré et al. (2010) reported up to 19.1 µg/L of fullerene C60 in wastewaters. Sun et al. (2014) predicted annual increase of fullerene in sewage sludge-treated soil between 0.38 and 1.5 µg/kg. Therefore, the aim of present study is to examine the effect of low, environmentally plausible concentrations of nC60 water suspensions on A. flavus aflatoxin production ability and oxidative stress modulation. Another coincidental emerging issue that increases the relevance of this work revolves around anticipated higher contamination rates of crops with aflatoxins due to climate change (Battilani et al. 2012, 2016).
Materials and methods
Chemicals
Superoxide dismutase from bovine erythrocytes (3000 U/mg protein) (SOD), glutathione reductase from baker’s yeast (Saccharomyces cerevisiae) (100–300 U/mg protein) (GR), xanthine oxidase from bovine milk (0.4–1.0 U/mg protein) (XOD), xanthine, nitrotetrazolium blue chloride (NBT), ethylenediaminetetraacetic acid tetrasodium salt (EDTA-4Na), l-glutathione, reduced (GSH), l-glutathione oxidised disodium salt (GSSG), potassium cyanide, diethylenetriaminepentaacetic acid (DTPA), N-ethylmaleimide (NEM) and sodium dithionite (DT) were purchased by Sigma Aldrich (Germany). β-nicotinamide adenine dinucleotide 2′-phosphate reduced tetrasodium salt (NADPH) was purchased from Serva (Germany), while stabilized 3% solution of hydrogen peroxide (H2O2), o-phthalaldehyde (OPA) and formic acid were purchased from Fluka (Germany). Sodium azide and hydrochloric acid were obtained from Merck (Germany). Aflatoxin standard mix (B1, G1, B2, G2) was purchased from Biopure (Austria). Acetonitrile and methanol (both HPLC grade) were obtained from J. T. Baker (Italy), while yeast extract, potato dextrose agar and sucrose were purchased from Biolife (Italy).
Trichloroacetic acid (TCA) and ascorbic acid were from Kemika (Croatia). Ethylenediaminetetraacetic acid disodium salt 4 (EDTA-2Na) was from Pharmacia Biotech (Sweden), absolute ethanol was from Panreac (Spain), while butylated hydroxytoluene (BHT) and 2-thiobarbituric acid (TBA) were obtained from Acros Organics (USA). Fullerene C60 was purchased from Mer Corporation (USA).
Fullerene C60 nanoparticles (nC60) solution preparation and characterization
Fullerene C60 nanoparticle solution was prepared according to Lyon et al. (2006), with modifications. Briefly, 200 mg of bulk C60 was stirred in 500 mL of ultrapure water (Millipore Simplicity 185, Merck, Germany) for 10 min at 550 rpm (Rotamix 550 MMH, Tehtnica, Slovenia) and sonicated (Sonorex Super RK 100 H, Bandelin, Germany) for 60 min in two cycles with 30 min of cooling between cycles. Prepared brown suspension was additionally stirred at 550 rpm for 30 days and then filtered through Whatman No. 1 filter and through 0.45 µm polyamide syringe filters (Chromafil Xtra, Macherey–Nagel, Germany). Sterilization of nC60 solution was performed in laminar flow hood (Airstream AC2-4E1, Esco, Singapur) using 0.45 µm polyamide syringe filters (Chromafil Xtra, Macherey–Nagel, Germany).
Characterization of nC60 solution included determination of particle size distribution and zeta potential. Dynamic light scattering (DLS) was used for determination of hydrodynamic size, and electrophoretic light scattering (ELS) for measurements of surface charge (zeta potential) of analysed samples. The measurements were conducted on a Zetasizer Nano ZS instrument (Malvern Instruments, UK). All DLS measurements were performed in triplicate, at 633 nm wavelength and a measurement angle of 173° (backscatter detection) at room temperature. The zeta potential measurements were conducted in duplicate.
Aspergillus flavus growth and aflatoxin production in culture media
Suspension of A. flavus NRRL 3251 conidia preparation, inoculation as well as mycelia growth in aflatoxin-inducing YES medium were conducted as described by Kovač et al. (2017). Incubation was conducted in the dark at 29 °C, conditions which favour aflatoxin production (Yu 2012), for 168 h on a rotary shaker (KS 260 basic, IKA, Germany) set to 200 rpm in the presence of environmentally plausible nC60 concentrations (0, 10, 50 and 100 ng/mL). Samples were collected every 24 h from 48 to 168 h, following separation from YES media by filtration. Mycelia obtained at the same time-point were pooled and homogenised using pestle and mortar. The main part of homogenised mycelia was stored at – 80 °C until analysis of cell oxidative status, while 200 mg was dried until constant mass (24 h at 105 °C) to determine dry mycelial weight.
Quantitative analysis of aflatoxin content in culture filtrates was performed by a “dilute and shoot” method as described by Kovač et al. (2017). Recovery was assessed by spiking blank YES medium with aflatoxin standard solution at a concentration of 10 ng/mL, and it was 92% for aflatoxin B1. Instrumental limits of detection were 0.15 ng/mL, and limits of quantification were 0.5 ng/mL for all aflatoxins. All quantified aflatoxin concentrations were corrected for recovery.
Disintegration of A. flavus mycelia
Extracts of A. flavus mycelia used for analysis of cell oxidative status were prepared by glass bead homogenization using a Bead Bug Microtube homogenizer (Benchmark Scientific, USA). Disintegration mixture contained 0.1 g of mycelia, 1 g of precooled, acid washed glass beads (diameter 0.5 mm) (Sigma Aldrich, Germany) and 1 mL of ice cold extraction buffer. Disintegration was performed at 4000 rpm in three cycles consisting of 2 min of disruption and 2 min of sample cooling on ice. Extracts were clarified by centrifugation at 15000×g and 4 °C for 20 min (Heraeus, Germany), and immediately used for analysis. Depending on the type of analysis, extraction buffers slightly differed in composition. Extracts used for antioxidant enzyme assays were prepared using 50 mM potassium phosphate buffer (pH 7.0) containing 1 mM EDTA-2Na, while buffers used in TBARS and GSH and GSSG assays additionally contained TCA (100 and 50 mg/mL, respectively).
Oxidative status of A. flavus NRRL 3251
The non-enzymatic (TBARS concentration and GSH/GSSG ratio) and enzymatic ROS-dependent markers (Cu,Zn-SOD, Mn-SOD, catalase (CAT), glutathione peroxidase (GPX) and GR) of oxidative status were determined.
TBARS assay was performed according to Luschak and Gospodaryov (2005). TBARS concentration in mycelia extracts was evaluated spectrophotometrically (Helios γ, ThermoSpectronic, UK) at 535 nm and molar extinction coefficient of malondialdehyde (ε535 nm = 156 × 103 L/cm/mol) was used for calculation. GSH and GSSG concentrations were estimated spectrofluorometrically (Cary Eclipse, Varian, Australia) according to Senft et al. (2000) using fluorescence indicator OPA.
Xanthine/xanthine oxidase/NBT assay (Angelova et al. 2005) was used for estimation of superoxide dismutase (EC 1.15.1.1) activities at 505 nm. Activities of cyanide sensitive Cu,Zn-SOD and the cyanide resistant Mn-SOD isoenzyme were also estimated. Total SOD activity was measured without, while Mn-SOD in the presence of mM potassium cyanide. Cu,Zn-SOD activity was calculated by substracting Mn-SOD from total SOD activity. CAT (EC 1.11.1.6) and GR (EC 1.8.1.7) activities were measured using spectrophotometric methods described by Reverberi et al. (2005), while GPX (EC 1.11.1.9) activity was measured at 340 nm according to Esworthy et al. (2005).
Protein measurement
The Bradford assay was used to determine protein concentration in prepared extracts (Bradford 1976).
Statistical analysis
Data presented in this paper are expressed as the mean value ± SEM from three separate experiments. The datasets were pooled and checked for normality of distribution by Shapiro–Wilk test and compared by nonparametric statistics methods (Friedman ANOVA and Kendall coefficient of concordance; Kruskal–Wallis ANOVA). The programme package Statistica 12.0 (Dell, USA) was used and differences were considered significant when the p value was < 0.05.
Results
Characterisation of nC60
Most of the nC60 nanoparticles in suspension were within 50–190 nm in diameter, and 21% had a hydrodynamic radius of 78 nm (Additional file 1: Figure S1a). Mean zeta potential value of the analysed nC60 solution was − 24.5 mV (Additional file 1: Figure S1b). These results of nC60 characterization are in accordance with a study conducted by Lyon et al. (2006) who reported the diameter of nC60 ranging from 30 to 100 nm. Also, 62.5% of nC60 nanoparticles in the prepared sample had diameters below 100 nm, which is in accordance with the EC Recommendation for a definition of the term “nanomaterial” (Rauscher et al. 2015).
nC60 influence on mycelial growth, oxidative stress and aflatoxin production
The effect of fullerene nanoparticles on A. flavus growth is presented in Fig. 1. A measurable mycelium mass was reached after 48 h of incubation. Collectively, only 50 ng/mL nC60 affected mycelial growth, causing a statistically significant (H = 32.04; p = 0.0016) higher dry mycelial mass after 48 h of growth (by 54%), but also a slight reduction in dry mycelium weight during exponential growth phase (after 72 h) and between 96 and 120 h of stationary growth phase. These statistically significant mass reductions were in a range from 6 to 16%. On the other hand, mycelial dry weight after 120 h of growth was similar to control as well as to samples treated with 10 and 100 ng/mL concentration of nC60. Both control and nC60-treated samples’ mycelial biomass showed a decreasing trend after 120 h of growth.
Fig. 1.

Influence of fullerene C60 nanoparticles (nC60) on A. flavus mycelial growth (expressed as gram of dry weight (g.d.w.) per 50 mL) in YES medium during 168 h at 29 °C. Data are represented as the mean ± SEM from three separate experiments
Analysis of control and nC60-treated samples on aflatoxin presence revealed dominance of aflatoxin B1 in growth media, while aflatoxin B2 levels were below LOQ and aflatoxin G1 and G2 were below LOD. Accordingly, only aflatoxin B1 levels are presented in Fig. 2. Fullerene nanoparticles caused a strong antiaflatoxigenic effect at all time-points at concentrations of 10 and 100 ng/mL, whereas no aflatoxin B1 was detected in the media up to 48 h (10 ng/mL) and 72 h (100 ng/mL) of growth. Aflatoxin B1 production was reduced by an average 67% for 10 ng/mL concentration and by an average 90% for 100 ng/mL concentration of nC60. The medium nC60 concentration of 50 ng/mL slightly decreased aflatoxin B1 production only between 96 and 120 h of incubation period, while further incubation resulted in approximately twofold higher levels compared to controls.
Fig. 2.

Influence on fullerene C60 nanoparticles (nC60) on A. flavus aflatoxin B1 production in the culture filtrates during 168 h of incubation at 29 °C. Aflatoxin content is expressed in ng aflatoxin B1 per g mycelial dry weight and represented as the mean ± SEM from three separate experiments
Since, aflatoxin B1 production is a secondary defence mechanism of fungi against oxidative stress in the cell (Reverberi et al. 2012) ROS-dependent oxidative stress biomarkers TBARS (Fig. 3a), GSH/GSSG (Fig. 3b) and activities of antioxidative enzymes (Fig. 4a–e) were determined.
Fig. 3.

Influence of fullerene C60 nanoparticles (nC60) on: a lipid peroxidation and b reduced and oxidised glutathione ratio (GSH/GSSG) in A. flavus mycelia during 168 h growth period in YES medium at 29 °C. Lipid peroxides are expressed as pmols of thiobarbituric acid reactive substances (TBARS) per dry weight of mycelia and represented as the mean ± SEM from three separate experiments
Fig. 4.
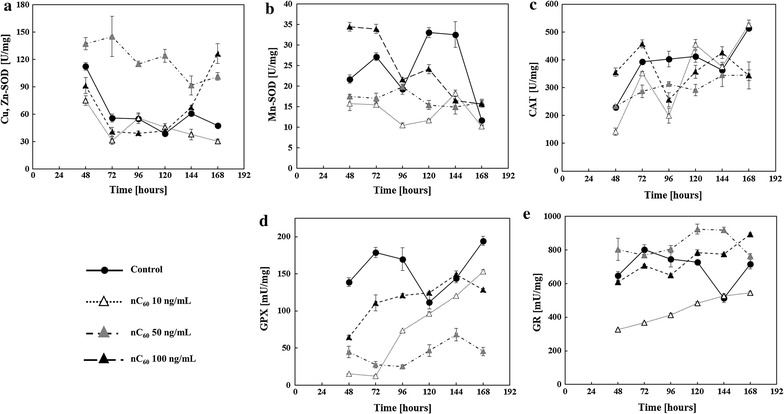
Modulation of antioxidant enzyme activities in A. flavus mycelium by fullerene C60 nanoparticles (nC60) during 168 h growth period in YES medium at 29 °C. Enzyme activity is expressed in U/mg of protein and represented as the mean ± SEM from three separate experiments. Enzymes: a copper, zinc superoxide dismutase (Cu,Zn-SOD), b manganese superoxid dismutase (Mn-SOD), c catalase (CAT), d glutathione peroxidase (GPX), and e glutathione reductase (GR)
The findings suggest that TBARS responded to nC60 treatments (Fig. 3a). Both 50 and 100 ng/mL nC60 led to somewhat elevated TBARS levels, especially during the final hours of incubation. The lowest dose had the opposite effect. In agreement with the TBARS results, the GSH/GSSG ratio (Fig. 3b) did not change significantly when treated with 10 ng/mL nC60. On the other hand, the GSH/GSSG ratio increased dramatically after 72 h of incubation with 50 and 100 ng/mL. GR activity displayed a similar pattern of responses to nC60 treatments (Fig. 4e).
Cu,Zn-SOD activity was found to be affected and increased only in the presence of 50 ng/mL nC60 (Fig. 4a), while Mn-SOD (Fig. 4b), CAT (Fig. 4c), and GPX activities (Fig. 4d) were decreased at almost all time points during incubation with all tested nC60 concentrations.
Discussion
The nC60 suspensions were prepared by long lasting stirring (30 days) of C60 powder in ultrapure water, as described above, in order to obtain environmentally most relevant nC60 form with no solvent or solubilizing agents involved. In this way, all residual solvent or solubilizing agent effects on observed nC60 interaction with A. flavus were excluded, and a potential scenario of C60 accidental release or disposal in the environment was mimicked in controlled laboratory conditions (Lyon et al. 2008; Lyon and Alvarez 2008). Noted minor differences in mycelial growth between treatments notwithstanding (Fig. 1), the overall impression is that this range of nC60 levels did not affect A. flavus growth substantially in either direction. This is in agreement with previous observations on eukaryotic microorganisms (Aoshima et al. 2009; Hadduck et al. 2010).
Quantification of aflatoxin B1 concentrations (Fig. 2 suggested general suppression of aflatoxin synthesis in the presence of nC60. Explaining the deviating results at the middle, 50 ng/mL nC60 level, is difficult but it could involve the distinct effects of C60 depending on concentration, medium, target organism, etc., as reported by other authors. For instance, the antioxidant effects of fullerene nanoparticles in rats were reported by Gharbi et al. (2005) and Baati et al. (2012). However, both in vitro (Isakovic 2006b) and in vivo (Oberdörster 2004) studies also determined prooxidative, ROS-related effects of fullerene nanoparticles. Moreover, Lyon et al. (2008) and Lyon and Alvarez (2008) demonstrated ROS-independent oxidative damage of bacterial cells by fullerene nanoparticles. Taking into account the relationship between A. flavus redox status and aflatoxin synthesis, and the versatility of nC60 action, an initial interpretation would assume antioxidative effects of nC60 at all tested levels except 50 ng/mL after 144 h of incubation.
Aflatoxin production is a secondary defence mechanism of fungi against oxidative stress in the cell (Reverberi et al. 2012; Hong et al. 2013; Grintzalis et al. 2014). Therefore, oxidative status of A. flavus cells was evaluated by determining the hallmarks of cellular ROS-inflicted damage and their elimination (Reverberi et al. 2005; Narasaiah et al. 2006): TBARS (Fig. 3a), GSH/GSSG ratio (Fig. 3b) and activities of antioxidative enzymes (Fig. 4).
The TBARS usually result from damage to lipids by hydroxyl radical which might be implicated in aflatoxin B1 production (Grintzalis et al. 2014). The present results on TBARS content (Fig. 3a) pointed to antioxidative action of C60 nanoparticles at the lowest concentration. The antioxidative effect was also observed in all other monitored oxidative status parameters (Figs. 2, 3, 4) in the presence of 10 ng/mL of nC60. Treatments with higher doses (50 and 100 ng/mL) exerted prooxidative effects in the fungus, leading to an increased formation of TBARS (Fig. 3a), and the GSH/GSSG ratio (Fig. 3b) which indicated an immense upregulation of GSH synthesis with the purpose of antioxidative defence. A congruous pattern of activity of GR, an enzyme involved in GSH maintenance in the cell (Li et al. 2009), was recorded for the selected range of nC60 doses (Fig. 4e).
An increase of Cu,Zn-SOD activity points to prooxidative effects of nC60, but curiously, only when A. flavus was treated with 50 ng/mL (Fig. 4a). Activity remained at control levels with the highest dose. Upregulation of Cu,Zn-SOD was reported to be a result of adaptive response of filamentous fungi to elevated ROS levels produced by a superoxide anion generator (Angelova et al. 2005). Presumably, nC60 generated superoxide anion, and such generation may have intensified in the YES medium which contains reducing agents as NADH or NADPH (Yamakoshi et al. 2003; Badireddy et al. 2007).
Since 100 ng/mL nC60 displayed prooxidative effects in terms of increased TBARS, GSH and GR, the absence of its influence on Cu,Zn-SOD activity may be an indication of cytotoxic effects of fullerene nanoparticles. The toxic effect of nC60 could be related to an interaction with cell membrane (Chen et al. 2010), absorption of nC60 and distribution in the organelles (Isaacson et al. 2009), etc. Additionally, inhibition of microtubule formation, mitochondrial dysfunction, disturbance of membrane transport, signal transduction, etc. (Johnson-Lyles et al. 2010; Ratnikova et al. 2011; Grebowski et al. 2013) reported for fullerenol nanoparticles should be considered. According to Chae et al. (2010), increased water solubility of fullerene nanoparticles results from surface hydroxylation. Thus, it seems quite possible that a part of the smallest fullerene nanoparticles acted as fullerol nanoparticles, whose effects on A. flavus were described by Kovač et al. (2017).
Fullerene nanoparticles could have, for example, either suppressed aflatoxin biosynthesis or interfered with aflatoxin release in the media (Holmes et al. 2008). In fact, the cytotoxic effects occurring at 100 ng/mL could reconcile prooxidative effects of nC60 (Fig. 3) with the lower concentration of aflatoxins in the growth media (Fig. 2).
The reduced activities of Mn-SOD, CAT and GPX (Fig. 4b–d) may be ascribed to antioxidative action of nC60, as mentioned above for the lowest (10 ng/mL) dose, to the inhibition of enzymes and cytotoxic effects, and/or the levels of peroxides were not large enough to upregulate CAT and GPX.
In conclusion, the present results clearly demonstrated volatility of nC60 effect on the mycotoxigenic potential of A. flavus. It may stimulate aflatoxin synthesis due to prooxidative action on the fungal cell, as detected with 50 ng/mL. An antiaflatoxigenic effect was observed at both 10–100 ng/mL nC60, with disparate mechanisms of action. An apparent antioxidative effect of 10 ng/mL nC60 reduced biosynthesis of aflatoxins (Reverberi et al. 2005), while 100 ng/mL nC60 exerted both prooxidative (measured) and cytotoxic (assumed) effects in fungal cells, thus effectively shutting down aflatoxin biosynthesis and/or their release from the cell. Further studies should extend the range of tested nC60 levels, investigate behaviour of photosensitised fullerene, and response of mycotoxigenic fungi in environmental conditions which are expected to favour aflatoxin synthesis due to climate change (Battilani et al. 2016; Medina et al. 2017).
Authors’ contributions
TK and IS conceived and designed experiments. TK, BŠ, MK, AN and IB carried out experiments. TK, IS, BŠ and TK analysed the data. TK wrote the manuscript. MK, BŠ, IB, IS and TK reviewed and revised manuscript. All authors read and approved the final manuscript.
Acknowledgements
Authors are grateful to Josip Juraj Strossmayer Osijek for providing research facilities.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
Most of the data supporting our findings are included in the manuscript.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Funding
The research was supported by Josip Juraj Strossmayer Osijek (Young scientist grant).
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- nC60
nanoparticles of fullerene C60
- ROS
reactive oxygen species
- GR
glutathione reductase
- XOD
xanthine oxidase
- NBT
nitrotetrazolium blue chloride
- EDTA-4Na
ethylenediaminetetraacetic acid tetrasodium salt
- GSH
glutathione reduced
- GSSG
glutathione oxidized
- OPA
o-phthalaldehyde
- NADPH
β-nicotinamide adenine dinucleotide 2′-phosphate reduced tetrasodium salt
- NEM
N-ethylmaleimide
- DT
sodium dithionite
- H2O2
hydrogen peroxide
- TCA
trichloroacetic acid
- EDTA-2Na
ethylenediaminetetraacetic acid disodium salt
- TBA
2-thiobarbituric acid
- YES
yeast extract sucrose
- TBARS
thiobarbituric acid reactive substances
- SOD
superoxide dismutase
- Cu,Zn-SOD
copper,zinc superoxide dismutase
- Mn-SOD
manganese superoxide dismutase
- GPX
glutathione peroxidase
- CAT
catalase
Additional file
Additional file 1. Size distributions by number (a) and zeta potential (b) of fullerene C60 nanoparticles.
Footnotes
Electronic supplementary material
The online version of this article (10.1186/s13568-018-0544-0) contains supplementary material, which is available to authorized users.
Contributor Information
Tihomir Kovač, Phone: +385 31 224 378, Email: tihomir.kovac@ptfos.hr.
Bojan Šarkanj, Email: bojan.sarkanj@ptfos.hr.
Tomislav Klapec, Email: tomi@ptfos.hr.
Ivana Borišev, Email: ivana.borisev@dh.uns.ac.rs.
Marija Kovač, Email: marija.kovac@inspecto.hr.
Ante Nevistić, Email: ante.nevistic@inspecto.hr.
Ivica Strelec, Email: ivica.strelec@ptfos.hr.
References
- Amaike S, Keller NP. Aspergillus flavus. Annu Rev Phytopathol. 2011;49:107–133. doi: 10.1146/annurev-phyto-072910-095221. [DOI] [PubMed] [Google Scholar]
- Angelova MB, Pashova SB, Spasova BK, Vassilev SV, Slokoska LS. Oxidative stress response of filamentous fungi induced by hydrogen peroxide and paraquat. Mycol Res. 2005;109:150–158. doi: 10.1017/S0953756204001352. [DOI] [PubMed] [Google Scholar]
- Aoshima H, Kokubo K, Shirakawa S, Ito M, Yamana S, Oshima T. Antimicrobial of fellerenes and their OH derivatives. Biocontrol Sci. 2009;14:69–72. doi: 10.4265/bio.14.69. [DOI] [PubMed] [Google Scholar]
- Baati T, Bourasset F, Gharbi N, Njim L, Abderrabba M, Kerkeni A, Szwarc H, Moussa F. The prolongation of the lifespan of rats by repeated oral administration of [60]fullerene. Biomaterials. 2012;33:4936–4946. doi: 10.1016/j.biomaterials.2012.03.036. [DOI] [PubMed] [Google Scholar]
- Badireddy AR, Hotze EM, Chellam S, Alvarez P, Wiesner MR. Inactivation of bacteriophages via photosensitization of fullerol nanoparticles. Environ Sci Technol. 2007;41:6627–6632. doi: 10.1021/es0708215. [DOI] [PubMed] [Google Scholar]
- Battilani P, Rossi V, Giorni P, Pietri A, Gualla A, van der Fels-Klerx H, Booij C, Moretti A, Logrieco A, Miglietta F, Toscano P, Miraglia M, De Santis B, Brera C. Scientific report submitted to EFSA. Modelling, predicting and mapping the emergence of aflatoxins in cereals in the EU due to climate change. EFSA. 2012;9(1):1–172. [Google Scholar]
- Battilani P, Toscano P, Van Der Fels-Klerx HJ, Moretti A, Leggieri MC, Brera C. Aflatoxin B1 contamination in maize in Europe increases due to climate change. Sci Rep. 2016;6:24328. doi: 10.1038/srep24328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- Chae SR, Badireddy AR, Farner Budarz J, Lin S, Xiao Y, Therezien M, Wiesner MR. Heterogeneities in fullerene nanoparticle aggregates affecting reactivity, bioactivity, and transport. ACS Nano. 2010;4:5011–5018. doi: 10.1021/nn100620d. [DOI] [PubMed] [Google Scholar]
- Chen R, Ratnikova TA, Stone MB, Lin S, Lard M, Huang G, Hudson JS, Ke PC. Differential uptake of carbon nanoparticles by plant and mammalian cells. Small. 2010;6:612–617. doi: 10.1002/smll.200901911. [DOI] [PubMed] [Google Scholar]
- Cherepanova NP, Panina LK, Bogomolova EV, Rybalchenko OV, Spitsina NG. The studying of fungi interactions with new carbon allotropic modification—Fullerens. Mikol Fitopatol. 1997;31:27–32. [Google Scholar]
- Duncan TV. Applications of nanotechnology in food packaging and food safety: barrier materials, antimicrobials and sensors. J Colloid Interface Sci. 2011;363:1–24. doi: 10.1016/j.jcis.2011.07.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Esworthy RS, CEsworthy RS, Chu FF DJ. Current protocols in Toxycology. New York: Wiley; 2005. pp. 7.1.1–7.1.32. [Google Scholar]
- European Commission DG (2012) “Science for Environment Policy”: European Commission DG Environment News Alert Service
- Farré M, Pérez S, Gajda-Schrantz K, Osorio V, Kantiani L, Ginebreda A, Barceló D. First determination of C60 and C70 fullerenes and N-methylfulleropyrrolidine C60 on the suspended material of wastewater effluents by liquid chromatography hybrid quadrupole linear ion trap tandem mass spectrometry. J Hydrol. 2010;383:44–51. doi: 10.1016/j.jhydrol.2009.08.016. [DOI] [Google Scholar]
- Farré M, Sanchís J, Barceló D. Analysis and assessment of the occurrence, the fate and the behavior of nanomaterials in the environment. TrAC Trends Anal Chem. 2011;30:517–527. doi: 10.1016/j.trac.2010.11.014. [DOI] [Google Scholar]
- Gharbi N, Pressac M, Hadchouel M, Szwarc H, Wilson SR, Moussa F. [60] Fullerene is a powerful antioxidant in vivo with no acute or subacute toxicity. Nano Lett. 2005;5:2578–2585. doi: 10.1021/nl051866b. [DOI] [PubMed] [Google Scholar]
- Grebowski J, Krokosz A, Puchala M. Membrane fluidity and activity of membrane ATPases in human erythrocytes under the influence of polyhydroxylated fullerene. Biochim Biophys Acta. 2013;1828:241–248. doi: 10.1016/j.bbamem.2012.09.008. [DOI] [PubMed] [Google Scholar]
- Grintzalis K, Vernardis SI, Klapa MI, Georgiou CD. Role of oxidative stress in sclerotial differentiation and aflatoxin B1 biosynthesis in Aspergillus flavus. Appl Environ Microbiol. 2014;80:5561–5571. doi: 10.1128/AEM.01282-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hadduck AN, Hindagolla V, Contreras AE, Li Q, Bakalinsky AT. Does aqueous fullerene inhibit the growth of Saccharomyces cerevisiae or Escherichia coli? Appl Environ Microbiol. 2010;76:8239–8242. doi: 10.1128/AEM.01925-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmes RA, Boston RS, Payne GA. Diverse inhibitors of aflatoxin biosynthesis. Appl Microbiol Biotechnol. 2008;78:559–572. doi: 10.1007/s00253-008-1362-0. [DOI] [PubMed] [Google Scholar]
- Hong S-Y, Roze L, Linz J. Oxidative stress-related transcription factors in the regulation of secondary metabolism. Toxins (Basel) 2013;5:683–702. doi: 10.3390/toxins5040683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hotze EM, Labille J, Alvarez P, Wiesner MR. Mechanisms of photochemistry and reactive oxygen production by fullerene suspensions in water. Environ Sci Technol. 2008;42:4175–4180. doi: 10.1021/es702172w. [DOI] [PubMed] [Google Scholar]
- Isaacson CW, Kleber M, Field JA. Quantitative analysis of fullerene nanomaterials in environmental systems: a critical review. Environ Sci Technol. 2009;43:6463–6474. doi: 10.1021/es900692e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Isakovic A. Distinct cytotoxic mechanisms of pristine versus hydroxylated fullerene. Toxicol Sci. 2006;91:173–183. doi: 10.1093/toxsci/kfj127. [DOI] [PubMed] [Google Scholar]
- Isakovic A. Distinct cytotoxic mechanisms of pristine versus hydroxylated fullerene. Toxicol Sci. 2006;91:173–183. doi: 10.1093/toxsci/kfj127. [DOI] [PubMed] [Google Scholar]
- Johnson-Lyles DN, Peifley K, Lockett S, Neun BW, Hansen M, Clogston J, Stern ST, McNeil SE. Fullerenol cytotoxicity in kidney cells is associated with cytoskeleton disruption, autophagic vacuole accumulation, and mitochondrial dysfunction. Toxicol Appl Pharmacol. 2010;248:249–258. doi: 10.1016/j.taap.2010.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klich MA. Aspergillus flavus: the major producer of aflatoxin. Mol Plant Pathol. 2007;8:713–722. doi: 10.1111/j.1364-3703.2007.00436.x. [DOI] [PubMed] [Google Scholar]
- Kovač T, Šarkanj B, Klapec T, Borišev I, Kovač M, Nevistić A, Strelec I. Fullerol C60(OH)24 nanoparticles and mycotoxigenic fungi: a preliminary investigation into modulation of mycotoxin production. Environ Sci Pollut Res. 2017;60:1–9. doi: 10.1007/s11356-017-9214-z. [DOI] [PubMed] [Google Scholar]
- Krusic PJ, Wasserman E, Keizer PN, Morton JR, Preston KF. Radical reactions of c60. Science. 1991;254:1183–1185. doi: 10.1126/science.254.5035.1183. [DOI] [PubMed] [Google Scholar]
- Li Q, Harvey LM, McNeil B. Oxidative stress in industrial fungi. Crit Rev Biotechnol. 2009;29:199–213. doi: 10.1080/07388550903004795. [DOI] [PubMed] [Google Scholar]
- Lushchak VI, Gospodaryov DV. Catalases protect cellular proteins from oxidative modification in Saccharomyces cerevisiae. Cell Biol Int. 2005;29:187–192. doi: 10.1016/j.cellbi.2004.11.001. [DOI] [PubMed] [Google Scholar]
- Lyon DY, Alvarez PJJ. Fullerene water suspension (nC) exerts antibacterial effects via ros-independent protein oxidation. Environ Sci Technol. 2008;42:8127–8132. doi: 10.1021/es801869m. [DOI] [PubMed] [Google Scholar]
- Lyon DY, Adams LK, Falkner JC, Alvarez PJJ. Antibacterial activity of fullerene water suspensions: effects of preparation method and particle size. Environ Sci Technol. 2006;40:4360–4366. doi: 10.1021/es0603655. [DOI] [PubMed] [Google Scholar]
- Lyon DY, Brunet L, Hinkal GW, Wiesner MR, Alvarez PJJ. Antibacterial activity of fullerene water suspensions (nC 60) is not due to ROS-mediated damage. Nano Lett. 2008;8:1539–1543. doi: 10.1021/nl0726398. [DOI] [PubMed] [Google Scholar]
- Medina A, Gilbert MK, Mack BM, OBrian GR, Rodríguez A, Bhatnagar D, Payne G, Magan N. Interactions between water activity and temperature on the Aspergillus flavus transcriptome and aflatoxin B1 production. Int J Food Microbiol. 2017;256:36–44. doi: 10.1016/j.ijfoodmicro.2017.05.020. [DOI] [PubMed] [Google Scholar]
- Michalitsch R, Kallinger C, Verbandt Y, Veefkind V, Huebner SR. The fullerene patent landscape in Europe. Nanotechnol Law Bus. 2008;5:85–94. [Google Scholar]
- Narasaiah KV, Sashidhar RB, Subramanyam C. Biochemical analysis of oxidative stress in the production of aflatoxin and its precursor intermediates. Mycopathologia. 2006;162:179–189. doi: 10.1007/s11046-006-0052-7. [DOI] [PubMed] [Google Scholar]
- Oberdörster E. Manufactured nanomaterials (fullerenes, C60) induce oxidative stress in the brain of juvenile largemouth bass. Environ Health Perspect. 2004;112:1058–1062. doi: 10.1289/ehp.7021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pycke BFG, Chao T-C, Herckes P, Westerhoff P, Halden RU. Beyond nC60: strategies for identification of transformation products of fullerene oxidation in aquatic and biological samples. Anal Bioanal Chem. 2012;404:2583–2595. doi: 10.1007/s00216-012-6090-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ratnikova TA, Nedumpully Govindan P, Salonen E, Ke PC. In vitro polymerization of microtubules with a fullerene derivative. ACS Nano. 2011;5:6306–6314. doi: 10.1021/nn201331n. [DOI] [PubMed] [Google Scholar]
- Rauscher H, Roebben G, Rauscher H, Roebben G, Sanfeliu AB, Emons H, Gibson N, Koeber R, Linsinger T, Rasmussen K, Sintes JR, Sokull-klüttgen B, Stamm H. Towards a review of the EC Recommendation for a definition of the term “nanomaterial”, Part 3: scientific-technical evaluation of options to clarify the definition and to facilitate its implementation. EC Joint Research Centre. 2015 [Google Scholar]
- Reverberi M, Fabbri AA, Zjalic S, Ricelli A, Punelli F, Fanelli C. Antioxidant enzymes stimulation in Aspergillus parasiticus by Lentinula edodes inhibits aflatoxin production. Appl Microbiol Biotechnol. 2005;69:207–215. doi: 10.1007/s00253-005-1979-1. [DOI] [PubMed] [Google Scholar]
- Reverberi M, Ricelli A, Zjalic S, Fabbri AA, Fanelli C. Natural functions of mycotoxins and control of their biosynthesis in fungi. Appl Microbiol Biotechnol. 2010;87:899–911. doi: 10.1007/s00253-010-2657-5. [DOI] [PubMed] [Google Scholar]
- Reverberi M, Punelli M, Smith CA, Zjalic S, Scarpari M, Scala V, Cardinali G, Aspite N, Pinzari F, Payne GA, Fabbri AA, Fanelli C. How peroxisomes affect aflatoxin biosynthesis in Aspergillus flavus. PLoS ONE. 2012;7:e48097. doi: 10.1371/journal.pone.0048097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roze LV, Hong S-Y, Linz JE. Aflatoxin biosynthesis: current frontiers. Annu Rev Food Sci Technol. 2013;4:293–311. doi: 10.1146/annurev-food-083012-123702. [DOI] [PubMed] [Google Scholar]
- Sanchís J, Berrojalbiz N, Caballero G, Dachs J, Farré M, Barceló D. Occurrence of aerosol-bound fullerenes in the mediterranean sea atmosphere. Environ Sci Technol. 2012;46:1335–1343. doi: 10.1021/es200758m. [DOI] [PubMed] [Google Scholar]
- Sanchís J, Božović D, Al-Harbi NA, Silva LF, Farré M, Barceló D. Quantitative trace analysis of fullerenes in river sediment from Spain and soils from Saudi Arabia. Anal Bioanal Chem. 2013;405:5915–5923. doi: 10.1007/s00216-013-6924-z. [DOI] [PubMed] [Google Scholar]
- Sanchís J, Bosch-Orea C, Farré M, Barceló D. Nanoparticle tracking analysis characterisation and parts-per-quadrillion determination of fullerenes in river samples from Barcelona catchment area. Anal Bioanal Chem. 2015;407:4261–4275. doi: 10.1007/s00216-014-8273-y. [DOI] [PubMed] [Google Scholar]
- Senft AP, Dalton TP, Shertzer HG. Determining glutathione and glutathione disulfide using the fluorescence probe o-phthalaldehyde. Anal Biochem. 2000;280:80–86. doi: 10.1006/abio.2000.4498. [DOI] [PubMed] [Google Scholar]
- Simonin M, Richaume A. Impact of engineered nanoparticles on the activity, abundance, and diversity of soil microbial communities: a review. Environ Sci Pollut Res. 2015;22:13710–13723. doi: 10.1007/s11356-015-4171-x. [DOI] [PubMed] [Google Scholar]
- Sun TY, Gottschalk F, Hungerbühler K, Nowack B. Comprehensive probabilistic modelling of environmental emissions of engineered nanomaterials. Environ Pollut. 2014;185:69–76. doi: 10.1016/j.envpol.2013.10.004. [DOI] [PubMed] [Google Scholar]
- Trpkovic A, Todorovic-Markovic B, Trajkovic V. Toxicity of pristine versus functionalized fullerenes: mechanisms of cell damage and the role of oxidative stress. Arch Toxicol. 2012;86:1809–1827. doi: 10.1007/s00204-012-0859-6. [DOI] [PubMed] [Google Scholar]
- Usenko CY, Harper SL, Tanguay RL. Fullerene C60 exposure elicits an oxidative stress response in embryonic zebrafish. Toxicol Appl Pharmacol. 2008;229:44–55. doi: 10.1016/j.taap.2007.12.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamakoshi Y, Umezawa N, Ryu A, Arakane K, Miyata N, Goda Y, Masumizu T, Nagano T. Active oxygen species generated from photoexcited fullerene (C-60) as potential medicines: O-2(-center dot) versus O-1(2) J Am Chem Soc. 2003;125:12803–12809. doi: 10.1021/ja0355574. [DOI] [PubMed] [Google Scholar]
- Yu J. Current understanding on aflatoxin biosynthesis and future perspective in reducing aflatoxin contamination. Toxins (Basel) 2012;4:1024–1057. doi: 10.3390/toxins4111024. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Most of the data supporting our findings are included in the manuscript.