

Abstract
Mycoplasma spp. are contagious bacteria, and mycoplasmal mastitis is a serious productivity problem on dairy farms. Bovine mammary epithelial cells (bMECs) have an important role in the elimination of pathogens, but the effect of Mycoplasma bovis on bMECs has not been fully described. To elucidate the immune response against intramammary infection by M. bovis, we undertook microarray analysis to examine and profile mRNA expression in bMECs after stimulation with M. bovis. We also compared the effects of M. bovis, Staphylococcus aureus, and Escherichia coli on immune-related mRNA expression in bMECs. Transcriptome analysis indicated a significant decrease in the level of mRNA-encoding lysine-specific demethylase 4D, suggesting that the immune response is suppressed by a decrease in histone demethylase activity. Interleukin (IL)-1β, IL-6, tumor necrosis factor alpha, toll-like receptor (TLR) 2, and TLR4 mRNA expression levels were significantly increased in bMECs stimulated with heat-killed M. bovis, but the expression levels were lower than those following stimulation by heat-killed S. aureus or E. coli. Our results suggest that M. bovis weakly affects mRNA expression in bMECs compared to the effects of E. coli or S. aureus. Moreover, live M. bovis may induce suppression of the immune response in bMECs.
Keywords: Mycoplasma bovis, bovine mammary epithelial cells, cytokines, innate immunity, microarray analysis
Introduction
Mycoplasma, a member of bacterial class Mollicutes, does not have a cell wall and is widespread in eukaryotes [19]. Among the Mycoplasma spp., Mycoplasma bovis is the most serious pathogen of cattle [14,16], contributing to mastitis, pneumonia, and arthritis [3,5,14]. Mycoplasma spp. are contagious bacteria on dairy farms and their ability to cause intramammary infection is a serious problem [17]. Because clinical mastitis caused by M. bovis is difficult to treat via antibiotic therapy, cows infected with M. bovis on farms must be culled to prevent outbreaks of mycoplasmal mastitis [17].
In mastitis, mammary epithelial cells (MECs) play an important role [18]. MECs have pattern recognition receptors (PRRs) that recognize pathogen-associated molecular patterns (PAMPs) that serve as the first line of defense against intramammary infections [18]. When MECs recognize PAMPs via PRRs, such as the toll-like receptor (TLR), the cells are induced to secrete chemokines, cytokines, and antimicrobial peptides, resulting in leukocyte recruitment and activation, helping in the fight against pathogens [18].
Escherichia coli and Staphylococcus aureus are well-studied major pathogens involved in mastitis and often cause different immune responses in bovine MECs (bMECs) [2,7,10,13]. It has previously been reported that the induction of tumor necrosis factor alpha (TNF-α), interleukin (IL)-1, and activated NF-κB in bMECs stimulated with E. coli was stronger than the induction in bMECs stimulated with S. aureus [6].
However, the immune response of bMECs to stimulation with M. bovis remains unclear. In the present study, we used microarray analysis to examine and profile mRNA expression in bMECs after stimulation with M. bovis; this work was expected to enhance the understanding of the immune response against intramammary infection by M. bovis. We evaluated the effect of M. bovis and compared that response to the effects of S. aureus and E. coli on proinflammatory cytokine-, chemokine- and antimicrobial peptide-related mRNA expression in bMECs.
Materials and Methods
Bacterial strains
M. bovis (PG45: ATCC 25523) was grown in modified PPLO medium (Kanto Kagaku, Japan) at 37℃ for 48 h. M. bovis was harvested by centrifugation (16,000 × g for 40 min) and then washed with phosphate-buffered saline. S. aureus (ATCC6538P) and E. coli (NBRC14237) were grown in brain-heart infusion medium (Nissui, Japan) at 37℃ for 24 h. The bacteria were heat-killed at 70℃ for 5 min (M. bovis) or at 75℃ for 30 min (S. aureus and E. coli). Live or heat-killed bacteria were suspended at 108 colony-forming unit/mL in RPMI 1640 medium with L-glutamine (RPMI 1640; Sigma-Aldrich, Japan) containing 10% fetal bovine serum and stored at −70℃ until required.
Bovine mammary epithelial cells
A clonal bMEC line established from a lactating Holstein cow was used as the model, as previously described [15]. Cells were stimulated with heat-killed or live M. bovis, S. aureus, or E. coli at multiplicities of infection (MOIs) of 10, 100 or 1,000 for 6 h, 12 h, and 24 h.
Total RNA isolation and cDNA synthesis
Total RNA (tRNA) extracted from the bMECs was obtained by using the PureLink RNA mini kit (Ambion, USA). Following DNAse digestion with TURBO DNA-free DNAse (Ambion), tRNA was quantified via spectrophotometry using a BioSpec-nano (Shimadzu, Japan). ReverTra Ace reverse transcriptase (Toyobo, Japan) and oligo dT primers (Toyobo) were used to synthesize cDNA from 1 µg of tRNA. For each reaction, a parallel negative control reaction was performed without reverse transcriptase. Polymerase chain reaction (PCR) was used to amplify the β-actin-encoding transcript, and bands were analyzed on 1.5% agarose gels stained with ethidium bromide and visualized on an ultraviolet transilluminator.
Microarray experiment and analysis
Data for six microarrays (3 stimuli, 3 control) of bMECs stimulated with M. bovis for 6 h were provided by Takara Bio (Japan). The gene expression dataset was obtained via Agilent single-color microarray platform (4 × 44K bovine gene expression array, grid ID 023647). Samples were processed for profiling via Agilent microarrays, and data were normalized as previously described [4]; t-tests were used to identify significant differences in gene expression (p<0.025) between samples. In a further filtering step, we selected only those genes exhibiting a fold change of ≥ 2. The whole dataset is publicly available from the ArrayExpress database (accession number E-MTAB-5306).
Quantitative PCR (qPCR) analysis
A Thunderbird SYBR qPCR mix (Toyobo) and a MyiQ-iCycler (Bio-Rad Laboratories, USA) were used for qPCR analysis. Primer sequences are depicted in Table 1. Melt curve analysis was used to confirm that each primer pair amplified only one product. We used a BLAST search (National Center for Biotechnology Information, USA) to confirm that only the target gene of interest was amplified by the primer sequences. The qPCR analyses were performed starting with denaturation at 95℃ for 5 min, followed by 40 cycles of denaturation at 95℃ for 15 sec, annealing at 60℃ for 30 sec, and extension at 72℃ for 30 sec. Melt curve analysis consisted of heating the PCR product from 55℃ to 95℃ and monitoring the fluorescence change at every increase of 0.5℃. We used the transcripts encoding β-actin and tryptophan 5-monooxygenase activation protein zeta polypeptide (YWHAZ) as reference genes [20,24].
Table 1. Sequences of oligonucleotide primers.

bp, base pair; F, forward; R, reverse; YWHAZ, tryptophan 5-monooxygenase activation protein zeta polypeptide; IL, interleukin; TNF, tumor necrosis factor; TLR, toll-like receptor; KDM4D, lysine-specific demethylase 4D.
Statistical analysis
Data from five bMECs were expressed as mean ± SE values. Inferential analysis was performed by using the Kruskal-Wallis test for comparison between groups, the Steel test for multiple comparisons, and the Welch's t-test for paired groups. These statistical analyses were performed by using Ekuseru-Toukei 2010 for Windows (Social Survey Research Information, Japan). In all cases, a p value of < 0.05 was considered to indicate statistical significance.
Results
Microarray analysis
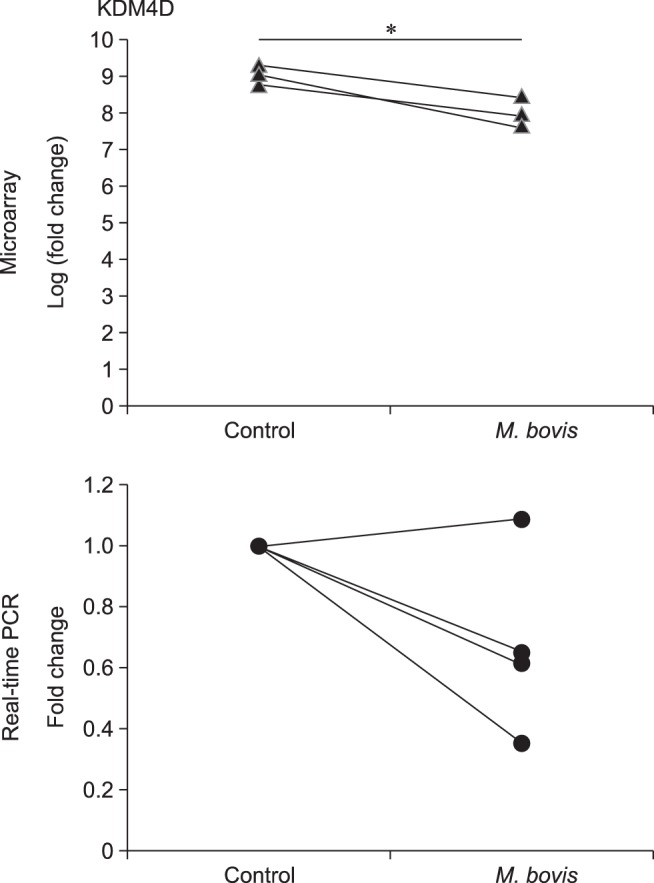
We investigated gene expression in bMECs stimulated with or without live M. bovis by using an Agilent Bovine Gene Expression Microarray. Statistical analysis revealed that expression of the gene encoding lysine-specific demethylase 4D (KDM4D; a protein with histone demethylase activity) was significantly decreased (p<0.025 with a fold increase>2) in bMECs stimulated with live M. bovis compared to expression in unstimulated bMECs. For validation of this result, KDM4D mRNA expression in bMECs stimulated with live M. bovis was quantified by using real-time PCR; the PCR result showed that expression of this gene was nominally decreased in bMECs stimulated with live M. bovis, although the data fell short of significance (p<0.061) (Fig. 1). In contrast, KDM4D gene expression was nominally increased (p<0.085) after stimulation with heat-killed M. bovis (data not shown).
Fig. 1. Microarray analysis and validation of KDM4D mRNA expression in bMECs stimulated with Mycoplasma bovis; bovine mammary epithelial cells (bMECs) were incubated with live M. bovis at an multiplicity of infection of 1,000 for 6 h. The data are expressed from three (microarray) or four (real-time polymerase chain reaction [PCR]) independent experiments. Significant difference (*p<0.05) compared with unstimulated bMECs.
Quantification of proinflammatory cytokine and chemokine mRNA expressions
The expression of genes encoding proinflammatory cytokines IL-1β, IL-6 and TNF-α in bMECs stimulated with M. bovis, S. aureus, or E. coli was evaluated by real-time PCR (Fig. 2). IL-1β, IL-6, and TNF-α mRNA expressions in bMECs stimulated with heat-killed M. bovis (MOIs of 1,000) for 12 h were significantly (p<0.01) increased compared with unstimulated cells. Stimulation with live M. bovis revealed a nominal increase that fell short of significance. In contrast, live S. aureus induced significant increases (p<0.01) in IL-1β (MOI of 10), IL-6 (MOI of 1,000), and TNF-α (MOI of 1,000) mRNA expression in bMECs after 24 h incubation. After 12 h of stimulation with E. coli, IL-1β (live: MOIs of 10, 100, and 1,000; heat-killed: MOIs of 10, 100, and 1,000), IL-6 (live: MOIs of 10, 100, and 1,000; heat-killed: MOIs of 100 and 1,000) and TNF-α (live: MOI of 100; heat-killed: MOI of 1,000) mRNA expressions in bMECs were significantly (p<0.01 or 0.05) increased.
Fig. 2. Expressions of cytokine mRNA in bovine mammary epithelial cells (bMECs) stimulated with Mycoplasma bovis, Staphylococcus aureus, or Escherichia coli; bMECs were incubated with M. bovis, S. aureus, or E. coli, live or heat-killed, at multiplicities of infection of 10 (△), 100 (□), and 1,000 (●) for 6, 12, and 24 h. Expressions of interleukin (IL)-1β, IL-6, and tumor necrosis factor alpha (TNF-α) mRNA determined by real-time polymerase chain reaction and expressed as fold increases, as described in Materials and Methods section. The data are expressed as mean ± SE values from five independent experiments. Significant differences (*p<0.05 or **p<0.01) compared with unstimulated bMECs.

Chemokine (IL-8) mRNA expressions in bMECs stimulated with M. bovis, S. aureus, or E. coli are shown in Fig. 3. There was no detectable effect of M. bovis on IL-8 mRNA expression in bMECs. However, significant increases in IL-8 mRNA expression in bMECs were detected after stimulation with live S. aureus at an MOI of 1,000 for 24 h (p<0.05), live E. coli at MOIs of 10 and 100 for 12 h (p<0.01 and 0.05, respectively), and heat-killed E. coli at an MOI of 1,000 for 12 h (p<0.05) and 24 h (p<0.05).
Fig. 3. Expressions of chemokine mRNA in bovine mammary epithelial cells (bMECs) stimulated with Mycoplasma bovis, Staphylococcus aureus, or Escherichia coli; bMECs were incubated with M. bovis, S. aureus, or E. coli, live or heat-killed, at multiplicities of infection of 10 (△), 100 (□), and 1,000 (●) for 6, 12, and 24 h. Expressions of interleukin (IL)-8 mRNA were determined by real-time polymerase chain reaction and are expressed as fold increases, as described in Materials and Methods section. The data are expressed as mean ± SE values from five independent experiments. Significant differences (*p<0.05 or **p<0.01) compared with unstimulated bMECs.

Quantification of antimicrobial peptide mRNA expression
The effects of M. bovis, S. aureus, and E. coli on the expression in bMECs of genes encoding antimicrobial peptides lactoferrin (Lf) and β-defensin were evaluated (Fig. 4). Changes in Lf and β-defensin mRNA expressions were not observed in bMECs stimulated with M. bovis. However, significant increases in Lf and β-defensin mRNA expression were found in bMECs stimulated with live S. aureus at an MOI of 1,000 for 24 h (Lf: p<0.05; β-defensin: p<0.01), live E. coli at MOIs of 10 for 24 h (Lf: p<0.05) and 1,000 for 24 h (β-defensin: p<0.05), and heat-killed E. coli at an MOI of 100 for 12 h (Lf: p<0.05) and 24 h (β-defensin: p<0.05) and at an MOI of 1,000 for 12 h (Lf: p<0.01; β-defensin: p<0.01).
Fig. 4. Expressions of antimicrobial peptide mRNA in bovine mammary epithelial cells (bMECs) stimulated with Mycoplasma bovis, Staphylococcus aureus, or Escherichia coli; bMECs were incubated with M. bovis, S. aureus, or E. coli, live or heat-killed, at multiplicities of infection of 10 (△), 100 (□), and 1,000 (●) for 6, 12, and 24 h. Expressions of lactoferrin (Lf) and β-defensin mRNA were determined by real-time polymerase chain reaction and expressed as fold increases, as described in Materials and Methods section. The data are expressed as mean ± SE values from five independent experiments. Significant differences (*p<0.05 or **p<0.01) compared with unstimulated bMECs.

Quantification of TLR mRNA expression
The effects of M. bovis, S. aureus, and E. coli on TLR mRNA expression in bMECs were evaluated (Fig. 5). Heat-killed M. bovis induced a significant (p<0.05) increase in TLR2 mRNA expression in bMECs at an MOI of 1,000 for 6 h. Significant increases in TLR2 mRNA expression were also detected in bMECs stimulated with live S. aureus at an MOI of 1,000 for 24 h (p<0.05), live E. coli at MOIs of 10 and 100 for 6 h (p<0.01), and heat-killed E. coli at MOIs of 10 and 100 for 6 h (p<0.01) and an MOI of 1,000 for 12 h (p<0.01).
Fig. 5. Expressions of TLR mRNA in bovine mammary epithelial cells (bMECs) stimulated with Mycoplasma bovis, Staphylococcus aureus, or Escherichia coli; bMECs were incubated with M. bovis, S. aureus, or E. coli, live or heat-killed, at multiplicities of infection of 10 (△), 100 (□), and 1,000 (●) for 6, 12, and 24 h. Expressions of toll-like receptor (TLR) 2 and TLR4 mRNA were determined by real-time polymerase chain reaction and expressed as fold increases, as described in Materials and Methods section. The data are expressed as mean ± SE values from five independent experiments. Significant differences (*p<0.05 or **p<0.01) compared with unstimulated bMECs.

Heat-killed M. bovis induced a significant (p<0.05) increase in TLR4 mRNA expression in bMECs at an MOI of 1,000 for 12 h. Significant increases in TLR4 mRNA expression were observed in bMECs stimulated with live S. aureus at an MOI of 10 for 24 h (p<0.01), live E. coli at MOIs of 10, 100, and 1,000 for 6 h (p<0.01, 0.01, and 0.05, respectively) and MOIs of 10 and 100 for 12 h (p<0.01), and heat-killed E. coli at MOIs of 10, 100, and 1,000 for 6 h (p<0.01, 0.01, and 0.05, respectively) and MOIs of 10, 100, and 1,000 for 12 h (p<0.05, 0.01, and 0.01, respectively).
Discussion
Transcriptome analysis revealed that KDM4D gene expression significantly decreased in bMECs stimulated with live M. bovis. KDM4D has been reported as one of a group of demethylases [12], but the protein's function in bMECs stimulated with M. bovis is unclear. Zhu et al. [26] reported that KDM4D regulates histone modification for activation of inflammatory gene transcription in dendritic cells and is required for NF-κB dependent inflammatory cytokine gene expression. In this study, we confirmed that live M. bovis did not affect mRNA expressions of proinflammatory cytokines (IL-1β, IL-6, and TNF-α) in bMECs. These results suggest that the decrease in KDM4D mRNA is related to the unchanged mRNA levels of proinflammatory cytokines in bMECs stimulated with live M. bovis. However, heat-killed M. bovis induced a significant increase in proinflammatory cytokine mRNA expression. In this study, we observed that KDM4D mRNA expression tended to increase following stimulation with heat-killed M. bovis. Although further studies are required to elucidate this difference in the response to live and heat-killed M. bovis, the difference may be related to a factor secreted by live M. bovis that suppresses the host's immune response. Our results suggest that KDM4D is an important factor in the regulation of proinflammatory cytokine mRNA expression in bMECs stimulated with M. bovis.
We did not observe changes in IL-8 mRNA expression in bMECs stimulated with M. bovis. We have already reported that IL-8 mRNA expression is increased in peripheral blood mononuclear cells (PBMCs) stimulated with live or heat-killed M. bovis and is similar to expression levels observed following stimulation of PBMC with S. aureus or E. coli [8]. Our results suggest that IL-8 mRNA expression is completely different in bMECs that it is in PBMCs following stimulation with M. bovis. IL-8 is one of the chemokines involved in recruitment of neutrophils from blood to the mammary gland and is related to the enhanced bactericidal activity of neutrophils [1]. It has been reported that a marked increase in somatic cell counts in milk was observed in mycoplasmal mastitis [11]. Our results suggest that bMECs are not a major factor in IL-8-regulated neutrophil recruitment or bactericidal function in mycoplasmal mastitis.
Lf and β-defensin mRNA expressions in bMECs stimulated with live or heat-killed M. bovis were similar to those of unstimulated bMECs. In contrast, expression levels in bMECs stimulated with live S. aureus, live E. coli, or heat-killed E. coli were clearly increased; results that are consistent with those in previous reports [6,9]. Both Lf and β-defensin have bactericidal activity against E. coli and S. aureus [21,25], but antimicrobial peptide activity against M. bovis has not been fully examined. Further study will be needed to clarify whether Lf and β-defensin are involved in the elimination of M. bovis.
Significant increases in TLR2 and TLR4 mRNA expression in bMECs were observed following stimulation with heat-killed M. bovis, but not with live M. bovis treatment. TLR2 and TLR4 are important receptors for recognition of Mycoplasma spp. in the immune response [22,23]. Our results suggest that live M. bovis inhibits TLR2 and TLR4 mRNA expressions in bMECs, an effect that may be related to immune evasion by M. bovis.
Interestingly, cytokine, chemokine, antimicrobial peptide, and TLR mRNA expressions in bMECs stimulated with S. aureus or E. coli were markedly higher than the expressions following stimulation with M. bovis. We have not yet clarified the mechanism responsible for these differences, but we do speculate that these differences are related to the characteristics of mycoplasmal mastitis.
In conclusion, M. bovis weakly affected transcriptional expressions in bMECs, and the observed effects on expression levels were different from those induced by E. coli or S. aureus. Live M. bovis may induce suppression of the immune response in bovine cells. For control of M. bovis infection, further studies of host immune responses will be needed.
Footnotes
Conflict of Interest: The authors declare no conflicts of interests.
References
- 1.Boulanger D, Bureau F, Mélotte D, Mainil J, Lekeux P. Increased nuclear factor kappaB activity in milk cells of mastitis-affected cows. J Dairy Sci. 2003;86:1259–1267. doi: 10.3168/jds.S0022-0302(03)73710-2. [DOI] [PubMed] [Google Scholar]
- 2.Brand B, Hartmann A, Repsilber D, Griesbeck-Zilch B, Wellnitz O, Kühn C, Ponsuksili S, Meyer HH, Schwerin M. Comparative expression profiling of E. coli and S. aureus inoculated primary mammary gland cells sampled from cows with different genetic predispositions for somatic cell score. Genet Sel Evol. 2011;43:24. doi: 10.1186/1297-9686-43-24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bürki S, Frey J, Pilo P. Virulence, persistence and dissemination of Mycoplasma bovis. Vet Microbiol. 2015;179:15–22. doi: 10.1016/j.vetmic.2015.02.024. [DOI] [PubMed] [Google Scholar]
- 4.Chain B, Bowen H, Hammond J, Posch W, Rasaiyaah J, Tsang J, Noursadeghi M. Error, reproducibility and sensitivity: a pipeline for data processing of Agilent oligonucleotide expression arrays. BMC Bioinformatics. 2010;11:344. doi: 10.1186/1471-2105-11-344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Fox LK. Mycoplasma mastitis: causes, transmission, and control. Vet Clin North Am Food Anim Pract. 2012;28:225–237. doi: 10.1016/j.cvfa.2012.03.007. [DOI] [PubMed] [Google Scholar]
- 6.Fu Y, Zhou E, Liu Z, Li F, Liang D, Liu B, Song X, Zhao F, Fen X, Li D, Cao Y, Zhang X, Zhang N, Yang Z. Staphylococcus aureus and Escherichia coli elicit different innate immune responses from bovine mammary epithelial cells. Vet Immunol Immunopathol. 2013;155:245–252. doi: 10.1016/j.vetimm.2013.08.003. [DOI] [PubMed] [Google Scholar]
- 7.Gilbert FB, Cunha P, Jensen K, Glass EJ, Foucras G, Robert-Granié C, Rupp R, Rainard P. Differential response of bovine mammary epithelial cells to Staphylococcus aureus or Escherichia coli agonists of the innate immune system. Vet Res. 2013;44:40. doi: 10.1186/1297-9716-44-40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gondaira S, Higuchi H, Iwano H, Nakajima K, Kawai K, Hashiguchi S, Konnai S, Nagahata H. Cytokine mRNA profiling and the proliferative response of bovine peripheral blood mononuclear cells to Mycoplasma bovis. Vet Immunol Immunopathol. 2015;165:45–53. doi: 10.1016/j.vetimm.2015.03.002. [DOI] [PubMed] [Google Scholar]
- 9.Griesbeck-Zilch B, Meyer HH, Kühn CH, Schwerin M, Wellnitz O. Staphylococcus aureus and Escherichia coli cause deviating expression profiles of cytokines and lactoferrin messenger ribonucleic acid in mammary epithelial cells. J Dairy Sci. 2008;91:2215–2224. doi: 10.3168/jds.2007-0752. [DOI] [PubMed] [Google Scholar]
- 10.Günther J, Esch K, Poschadel N, Petzl W, Zerbe H, Mitterhuemer S, Blum H, Seyfert HM. Comparative kinetics of Escherichia coli- and Staphylococcus aureus-specific activation of key immune pathways in mammary epithelial cells demonstrates that S. aureus elicits a delayed response dominated by interleukin-6 (IL-6) but not by IL-1A or tumor necrosis factor alpha. Infect Immun. 2011;79:695–707. doi: 10.1128/IAI.01071-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kauf AC, Rosenbusch RF, Paape MJ, Bannerman DD. Innate immune response to intramammary Mycoplasma bovis infection. J Dairy Sci. 2007;90:3336–3348. doi: 10.3168/jds.2007-0058. [DOI] [PubMed] [Google Scholar]
- 12.Krishnan S, Trievel RC. Structural and functional analysis of JMJD2D reveals molecular basis for site-specific demethylation among JMJD2 demethylases. Structure. 2013;21:98–108. doi: 10.1016/j.str.2012.10.018. [DOI] [PubMed] [Google Scholar]
- 13.Lahouassa H, Moussay E, Rainard P, Riollet C. Differential cytokine and chemokine responses of bovine mammary epithelial cells to Staphylococcus aureus and Escherichia coli. Cytokine. 2007;38:12–21. doi: 10.1016/j.cyto.2007.04.006. [DOI] [PubMed] [Google Scholar]
- 14.Maunsell FP, Woolums AR, Francoz D, Rosenbusch RF, Step DL, Wilson DJ, Janzen ED. Mycoplasma bovis infections in cattle. J Vet Intern Med. 2011;25:772–783. doi: 10.1111/j.1939-1676.2011.0750.x. [DOI] [PubMed] [Google Scholar]
- 15.Nakajima K, Nakamura M, Gao XD, Kozakai T. Possible involvement of prolactin in the synthesis of lactoferrin in bovine mammary epithelial cells. Biosci Biotechnol Biochem. 2008;72:1103–1106. doi: 10.1271/bbb.70713. [DOI] [PubMed] [Google Scholar]
- 16.Nicholas RA. Bovine mycoplasmosis: silent and deadly. Vet Rec. 2011;168:459–462. doi: 10.1136/vr.d2468. [DOI] [PubMed] [Google Scholar]
- 17.Nicholas RA, Ayling RD. Mycoplasma bovis: disease, diagnosis, and control. Res Vet Sci. 2003;74:105–112. doi: 10.1016/s0034-5288(02)00155-8. [DOI] [PubMed] [Google Scholar]
- 18.Rainard P, Riollet C. Innate immunity of the bovine mammary gland. Vet Res. 2006;37:369–400. doi: 10.1051/vetres:2006007. [DOI] [PubMed] [Google Scholar]
- 19.Razin S, Yogev D, Naot Y. Molecular biology and pathogenicity of mycoplasmas. Microbiol Mol Biol Rev. 1998;62:1094–1156. doi: 10.1128/mmbr.62.4.1094-1156.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Robinson TL, Sutherland IA, Sutherland J. Validation of candidate bovine reference genes for use with real-time PCR. Vet Immunol Immunopathol. 2007;115:160–165. doi: 10.1016/j.vetimm.2006.09.012. [DOI] [PubMed] [Google Scholar]
- 21.Selsted ME, Tang YQ, Morris WL, McGuire PA, Novotny MJ, Smith W, Henschen AH, Cullor JS. Purification, primary structures, and antibacterial activities of beta-defensins, a new family of antimicrobial peptides from bovine neutrophils. J Biol Chem. 1993;268:6641–6648. [PubMed] [Google Scholar]
- 22.Shimizu T. Inflammation-inducing factors of Mycoplasma pneumoniae. Front Microbiol. 2016;7:414. doi: 10.3389/fmicb.2016.00414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Shio MT, Hassan GS, Shah WA, Nadiri A, El Fakhry Y, Li H, Mourad W. Coexpression of TLR2 or TLR4 with HLA-DR potentiates the superantigenic activities of Mycoplasma arthritidis-derived mitogen. J Immunol. 2014;192:2543–2550. doi: 10.4049/jimmunol.1300591. [DOI] [PubMed] [Google Scholar]
- 24.Spalenza V, Girolami F, Bevilacqua C, Riondato F, Rasero R, Nebbia C, Sacchi P, Martin P. Identification of internal control genes for quantitative expression analysis by real-time PCR in bovine peripheral lymphocytes. Vet J. 2011;189:278–283. doi: 10.1016/j.tvjl.2010.11.017. [DOI] [PubMed] [Google Scholar]
- 25.Ulvatne H, Vorland LH. Bactericidal kinetics of 3 lactoferricins against Staphylococcus aureus and Escherichia coli. Scand J Infect Dis. 2001;33:507–511. doi: 10.1080/00365540110026692. [DOI] [PubMed] [Google Scholar]
- 26.Zhu Y, van Essen D, Saccani S. Cell-type-specific control of enhancer activity by H3K9 trimethylation. Mol Cell. 2012;46:408–423. doi: 10.1016/j.molcel.2012.05.011. [DOI] [PubMed] [Google Scholar]