

Abstract
Objectives
Selective targeting of cancer-associated fibroblasts (CAFs) has been proposed to synergize with immune-checkpoint inhibitors. While the roles of CAFs in cancer development are well described, their immune-regulatory properties remain incompletely understood. This study investigates correlations between CAF and immune-markers in tumor stroma from non-small cell lung cancer (NSCLC) patients, and examines whether a combination of CAF and immune cell scores impact patient prognosis.
Methods
Tumor specimens from 536 primary operable stage I-III NSCLC patients were organized in tissue microarrays. Expression of protein-markers was evaluated by immunohistochemistry.
Results
Fibroblast and stromal markers PDGFRα, PDGFRβ, FAP-1 and vimentin showed weak correlations while αSMA, and Masson’s trichrome did not correlate with any of the investigated markers. Hierarchical clustering indicated the existence of different CAF-subsets. No relevant correlations were found between any CAF-marker and the immune-markers CD3, CD4, CD8, CD20, CD68, CD1a, CD56, FoxP3 and CD45RO. High density of fibroblast-activation protein positive mesenchymal cells (CAFFAP) was associated with better prognosis in tumors with high infiltration of CD8 and CD3 T-lymphocytes.
Conclusions
The presented data suggest that CAFs, irrespective of identity, have low influence on the degree of tumor infiltration by inflammatory- and/or immune-cells. However, CAFFAP may exert immuno-adjuvant roles in NSCLC, and targeting CAFs should be cautiously considered.
Introduction
In solid tumors, complex and reciprocal interactions between neoplastic cells and surrounding cells lead to a tumor tissue compartment often referred to as reactive stroma, desmoplastic stroma or tumor microenvironment. A dominant component of the tumor stroma are fibroblasts, which are known to play determinant roles in tumor initiation, expansion, dissemination and metastasis [1]. Cancer-associated fibroblasts (CAFs) is a generic name given to a heterogeneous group of non-epithelial, non-immune cells with a likely mesenchymal lineage, located within tumors or at the tumor borders [2]. Quiescent connective tissue fibroblasts are generally considered indolent, exhibiting rather low metabolic and transcriptomic activity, and expressing classical mesenchymal markers such as vimentin, integrin α1β1 or FSP-1 [3]. Fibroblasts associated with tumors normally display an activated phenotype, and depending on their origin, morphology or spatial distribution, they may receive different names such as myofibroblasts, activated tumor fibroblasts, activated stellate cells, bone marrow-derived mesenchymal stromal cells or pericytes [4, 5]. Several markers such as αSMA, FAP-1, desmin, podoplanin, neuron-glial antigen 2 (NG2) and PDGF receptors-α and -β are used to identify CAFs. However, due to the great plasticity of this cell population, none of these markers can be used as a universal marker for all CAFs as their expression is likely to be temporal and context dependent [6]. Different CAF subsets, expressing overlapping and non-overlapping markers, can be identified in a single tumor. However, it remains unknown whether the various CAF subtypes in tumors have different functions. In non-small cell lung cancer (NSCLC), several studies have explored the prognostic significance of established CAF markers such as podoplanin, vimentin, FAP-1, αSMA or PDGFRβ. In most cases, these markers have demonstrated unfavorable outcomes related to survival (Table 1).
Table 1. Prognostic role of different CAF markers in lung cancer cohorts.
| Fibroblast marker | Tumor subtype | Samples (n) | Technique | Prognostic impact | Ref. |
|---|---|---|---|---|---|
| αSMA | NSCLC Stage I-III | 78 | IHC whole tissue/PCR | Unfavorable | Chen [7] |
| NSCLC Stage I-IIIA | 536 | IHC/TMA | No significance | Kilvaer [8] | |
| αSMA/ Vimentin | ADC | 102 | IHC whole tissue | Unfavorable | Shu [9] |
| Vimentin | NSCLC Stage I-IIIA | 335 | IHC/TMA | No significance | Al-Saad [10] |
| Podoplanin | ADC Stage-I | 304 | IHC whole tissue | Unfavorable | Ito [11] |
| ADC | 177 | IHC whole tissue | Unfavorable | Kawase [12] | |
| ADC (post chemotherapy) | 87 | IHC whole tissue | Unfavorable | Koriyama [13] | |
| ADC N2 Stage-III | 112 | IHC Lymph nodes | Unfavorable | Neri [14] | |
| SCLC | 36 | IHC whole tissue | Favorable | Takahashi [15] | |
| NSCLC | 400 | IHC/TMA | Unfavorable | Kitano [16] | |
| SCC Stage-I | 142 | IHC whole tissue | Unfavorable | Ono [17] | |
| FAP-1 | NSCLC | 59 | IHC whole tissue | Unfavorable | Liao [18] |
| NSCLC Stage I-III | 536 | IHC/TMA | Favorable in SCC | Kilvaer [8] | |
| MMP-2 | NSCLC Stage I-IIIA | 218 | IHC whole tissue | Unfavorable | Ishikawa [19] |
| NSCLC | 212 | IHC whole tissue | Unfavorable | Leinonen [20] | |
| CD99 | NSCLC | 190/240 (2cohorts) | IHC/TMA | Favorable | Edlund [21] |
| PDGFRβ | NSCLC | 190 | IHC/TMA | No significance | Edlund [21] |
Abbreviations: NSCLC, non-small cell lung cancer; IHC, immunohistochemistry; PCR, polymerase chain reaction; ADC, adenocarcinoma; SCLC, small-cell lung cancer; SCC, squamous cell carcioma
Despite the widely acknowledged role of CAFs in malignant progression, the understanding of their relationship with tumor-infiltrating lymphocytes (TILs) is incomplete. Emerging evidence propose that the tumor stroma influence tumor immunity and response to immunotherapy [22]. CAFs are suggested to interfere with tumor immunity in multiple ways, including remodeling of extracellular matrix, shaping the phenotype of vessels and endothelial cells and directly influencing the migration and function of inflammatory and immune cells by the release of paracrine signals. In general, CAFs are believed to exert pro-inflammatory and immunosuppressive functions in tumors [23, 24]. However, some studies have challenged this view, arguing that different CAF subsets may exert opposite functions, and that in a context-dependent manner, CAFs may aid the antitumor immune responses [25]. In the context of cancer immunotherapy, selective depletion of CAFFAP has been shown to synergize with immune-checkpoint inhibitors in pre-clinical models [26], thus proposing that targeting immunosuppressive elements of the tumor stroma may aid to the efficacy of immuno-therapeutic interventions.
We have previously explored the prognostic significance of CAF-markers in the stroma of resected NSCLC tumors [8], with the intriguing finding that high CAFFAP levels were associated with an improved prognosis in patients with squamous cell carcinoma. Earlier studies on NSCLC have also highlighted the prognostic relevance of other stromal markers such as CD99 [21], Forkhead Box F1 [27], and Cox-2 [28]. In the present study, we explore associations between CAFs and immune cell infiltrates in NSCLC tumor tissues. Additionally, we investigate whether CAFs impact patient survival according to different levels of immune cell infiltration.
Materials and methods
Patients and clinical samples
This study presents data on a cohort of 536 unselected stage IA-IIIA NSCLC patients. A detailed description of the cohort was published previously by our group [29]. Briefly, the included patients were staged after the 7th edition of the UICC TNM classification [30], and histologically classified according to the 2013 edition of the pathological classification of lung cancer [31]. Of the 536 patients, 289 were squamous cell carcinomas, 201 were adenocarcinomas and 46 were classified as NOS. Most patients classified as NOS were previously considered as large cell carcinoma, while a few patients were too undifferentiated to be classified. The Regional Committee for Medical and Health Research Ethics (REK-Nord) approved the use of human material for this study (Project-ID: 2016/2307/REK-Nord). Due to the retrospective nature of the study, and the fact that two thirds of the study population was deceased at time of study initiation, the need of written informed consent was waivered. All methods involving human material were performed in accordance with relevant guidelines and regulations.
Tissue micro-array construction, immunohistochemistry and scoring
The work-flow for sample preparation, tissue micro-array (TMA) construction, immunohistochemistry (IHC) and scoring of TMA-slides is extensively documented [8, 29, 32]. In brief, representative areas of primary lung tumor stroma specimens were identified on H&E slides. Two duplicate tissue cores were collected from the primary tumor blocks based on the overlay of the H&E stained slides. TMAs were assembled using a Beecher Instruments tissue-arraying instrument (Beecher Instruments, Silver Springs, MD, USA). Blocks were sectioned with 4 μm thickness and heated over-night at 60°C. The IHC staining procedures- including validations of antibodies—for FAP-1 and αSMA [8], CD3 (pan T-cell marker), CD4 (T-helper cells), CD8 (cytotoxic T-cells), CD20 (B-cells) and CD45RO (T-memory cells) and CD1A (Dendritic cells), CD56 (Natural Killer cells) and CD68 (macrophages) [33] were previously reported. The S1 Table includes a summary of staining procedures for the CAF markers [29]. Expert pathologists established semi-quantitative cut-offs for each marker. Immune- and CAF markers were scored as percentages of positive cells from the total number of cells in the stromal area, using the following thresholds: CD3, CD4, CD8, CD20 and CD45RO 0, = <1%, 1 = 1–5%, 2 = 6–25%, 3 = 26–50%, 4 = >50% (29); CD1A and CD56, 0 = < 1%, 1 = > 1%; CD68, 0 = <25%, 1 = > 25% [33]; FAP1, [SMA, PDGFRα and PDGFRβ 0 = <1%, 1 = 1–10%, 2 = 11–50%,d 3 = > 50% [8]. Vimentin was scored based on intensity and density with the following thresholds: intensity 1 = weak, 2 = intermediate and 3 = strong [10]. Micro vessels density and lymphatic micro vessel density was scored based on the total number of vessels found per core, using the following thresholds: CD34, 0 = negative; 1 = (1–10 vessels per core); 2 = (11–20 vessels per core); 3 = (>20 vessels per core) [30]; D240, 0 = negative; 1 = (one vessel per core); 2 = (2–5 vessels per core); 3 = (>5 vessels per core) [34]. All TMA cores were scored independently by two investigators, blinded to each other and to patient data. High expression of CAF-markers were defined as: > 0.5 (FAP-1), >2 (αSMA), >1,75 (PDGFRα) and > 1.5 (PDGFRβ) utilizing an optimal cut-off approach (Fig 1). The cut-offs used for FAP1 and αSMA were previously defined [8]. In order to evenly distribute CAF high and low patients in subgroup analyses based on immune cell infiltration, high expression of immune markers (CD3 and CD8) was decided bases on a median cut-off approach (S1 Fig)
Fig 1. Immunostaining of TMA cores showing low vs high score of the four main CAFs markers.

(A) PDGFRβ; (B) PDGFRα; (C) FAP-1; (D) αSMA. Abbreviations: PDGFR, platelet-derived growth factor receptor; FAP-1, Fibroblast activating protein 1; αSMA, alpha-smooth muscle actin.
Statistical methods
All statistical analyses were conducted in RStudio, version 1.0.136 with R version 3.3.2 and packages "sjmisc", "Hmisc", "survival", "ggplot2", "reshape2", "grid", "gridExtra" and "cowplot". Associations between the dichotomized markers and clinicopathological variables were tested with Chi-square or Fisher's exact tests. Due to the semi-quantitative scoring of the investigated markers, Spearman`s rank-correlation was used to examine the between-marker correlations and visualized utilizing heat-maps and hierarchical clustering. Survival analyses were conducted and visualized using the log-rank test and the Kaplan-Meier method To allow the investigation of markers in the presence of other co-variables from the same dataset, Cox proportional hazards models were fitted to the data. A supervised iterative process, based on significance levels in each step, was used to select the variables included in the final models. The significance level used for all statistical tests was P < 0.05.
Results
Associations between markers and clinicopathological variables
No significant associations were observed between neither PDGFRα, nor vimentin expression and clinicopathological variables (Table 2). Low expression of PDGFRβ was associated with normal ECOG status (P = 0.003) and high expression of αSMA was associated with vascular invasion (P = 0.012). No associations between FAP-1 and clinicopathological variables was observed, as previously reported by our group [8].
Table 2. Correlations between marker expressions and clinicopathological variables with stromal PDGFRα, PDGFRβ, αSMA and vimentin (chi-square and Fisher's exact tests).
| PDGFRα S | PDGFRβ S | αSMA | Vimentin | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Low | High | P | Low | High | P | Low | High | P | Low | High | P | |
| Age | 0.431 | 0.443 | 0.341 | 0.594 | ||||||||
| <65 | 146 | 68 | 124 | 90 | 148 | 79 | 100 | 35 | ||||
| ≥65 | 207 | 81 | 176 | 109 | 214 | 94 | 133 | 55 | ||||
| Gender | 0.225 | 1.000 | 0.138 | 0.331 | ||||||||
| Female | 109 | 55 | 97 | 64 | 123 | 47 | 51 | 25 | ||||
| Male | 244 | 94 | 203 | 135 | 239 | 126 | 182 | 65 | ||||
| Weight-loss | 0.659 | 0.119 | 0.602 | 0.808 | ||||||||
| <10% | 319 | 132 | 275 | 173 | 327 | 153 | 211 | 80 | ||||
| >10% | 34 | 17 | 25 | 26 | 35 | 20 | 22 | 10 | ||||
| Smoking status | 0.842 | 0.100 | 0.726 | 0.041 | ||||||||
| Never | 10 | 4 | 12 | 2 | 10 | 7 | 13 | 0 | ||||
| Present | 222 | 98 | 184 | 132 | 233 | 109 | 146 | 63 | ||||
| Previous | 121 | 47 | 104 | 65 | 119 | 57 | 74 | 27 | ||||
| ECOG | 0.756 | 0.003 | 0.704 | 0.860 | ||||||||
| Normal | 203 | 91 | 193 | 98 | 211 | 98 | 136 | 51 | ||||
| Slightly | 125 | 48 | 91 | 82 | 125 | 65 | 85 | 33 | ||||
| In bed<50% | 25 | 10 | 16 | 19 | 26 | 10 | 12 | 6 | ||||
| Histology | 0.069 | 0.093 | 0.393 | 0.303 | ||||||||
| SCC | 190 | 83 | 152 | 120 | 198 | 90 | 137 | 53 | ||||
| ADC | 127 | 60 | 124 | 64 | 137 | 64 | 77 | 25 | ||||
| NOS | 36 | 6 | 24 | 15 | 27 | 19 | 19 | 12 | ||||
| T-stage | 0.159 | 0.840 | 0.505 | 0.888 | ||||||||
| T1a | 49 | 22 | 45 | 26 | 53 | 21 | 25 | 11 | ||||
| T1b | 68 | 20 | 54 | 34 | 63 | 31 | 42 | 15 | ||||
| T2a | 127 | 49 | 101 | 71 | 128 | 61 | 86 | 36 | ||||
| T2b | 42 | 29 | 46 | 25 | 48 | 27 | 33 | 14 | ||||
| T3 | 64 | 26 | 51 | 40 | 68 | 29 | 47 | 14 | ||||
| T4 | 3 | 3 | 3 | 3 | 2 | 4 | 0 | 0 | ||||
| N-stage | 0.273 | 0.911 | 0.541 | 0.227 | ||||||||
| N0 | 237 | 105 | 202 | 136 | 248 | 116 | 159 | 64 | ||||
| N1 | 85 | 27 | 68 | 42 | 75 | 42 | 58 | 16 | ||||
| N2 | 31 | 17 | 30 | 21 | 39 | 15 | 16 | 10 | ||||
| P-stage | 0.769 | 0.938 | 0.875 | 0.954 | ||||||||
| IA | 93 | 34 | 79 | 47 | 94 | 40 | 53 | 21 | ||||
| IB | 79 | 33 | 67 | 43 | 81 | 41 | 52 | 23 | ||||
| IIA | 82 | 32 | 65 | 45 | 76 | 42 | 63 | 21 | ||||
| IIB | 48 | 23 | 43 | 29 | 51 | 24 | 35 | 14 | ||||
| IIIA | 51 | 27 | 46 | 35 | 60 | 26 | 30 | 11 | ||||
| Differentiation | 0.129 | 0.250 | 0.688 | 0.917 | ||||||||
| Poor | 153 | 59 | 131 | 78 | 160 | 70 | 98 | 39 | ||||
| Moderate | 152 | 77 | 138 | 91 | 158 | 82 | 104 | 38 | ||||
| Well | 48 | 13 | 31 | 30 | 44 | 21 | 31 | 13 | ||||
| Vascular invasion | 0.520 | 0.906 | 0.012 | 0.236 | ||||||||
| No | 292 | 119 | 244 | 163 | 306 | 130 | 200 | 72 | ||||
| Yes | 60 | 29 | 55 | 35 | 55 | 42 | 33 | 18 | ||||
Abbreviations: PDGFR, platelet derived growth factor; αSMA, alpha-smooth muscle actin; S, stroma; ECOG, Eastern cooperative oncology group; SCC, squamous cell carcinoma; ADC, adenocarcinoma; NOS, not otherwise specified
Correlations between CAF markers
Fig 2A and S2A and S2B Fig summarizes the correlations between different CAF-markers investigated in the overall cohort and the two main histological subgroups of NSCLC, respectively. In the overall cohort, hierarchical clustering suggested a CAF phenotype in patients expressing vimentin, FAP-1 and PDGFRα and -β. On the other hand αSMA was only weakly associated with the other CAF-markers (-0.2<R<0.2). Considering correlations with R-values < -0.2 and > 0.2: 1) PDGFRα was associated with FAP-1 (R = 0.28) and PDGFRβ (R = 0.33); 2) PDGFRβ was associated with vimentin (R = 0.24) and FAP-1 (R = 0.32); and 3) αSMA was associated with Masson's trichrome (MT) (R = 0.20). In addition, correlations of CAF-markers with angio- and lymphangigenic markers CD34 and D240 had low R-values (< 0.2). Other significant, but weak (-0.2 < R < 0.2) associations were also observed. Correlations in histological subgroups were mostly similar to those observed in the overall cohort (S2A and S2B Fig). Distinct expression patterns of FAP-1 and αSMA (both considered hallmarks of activated CAFs) in the same tissue specimens are shown in Fig 3.
Fig 2.

Spearman's rank correlations between A) Different CAF markers and B) CAF markers and markers of leukocyte subsets. *P < 0.05, **P < 0.01, ***P < 0.001. Abbreviations: CAF, cancer-associated fibroblast; Vim, vimentin; FAP-1, Fibroblast activating protein 1; PDGFR, platelet-derived growth factor receptor; αSMA, alpha-smooth muscle actin; MT, Masson's trichrome; CD, cluster of differentiation.
Fig 3. Double staining of FAP-1 & αSMA showing distinct expression patterns: in three randomly selected TMA cores corresponding to three NSCLC patients co-stained with FAP-1(purple) and αSMA(yellow).

Correlations between CAFs and immune-cell markers
Fig 2B and S2C and S2D Fig summarizes the correlations between CAF- and immune-cell markers in the overall cohort and the two main histological subtypes of NSCLC, respectively. No clear strong correlations between CAF- and immune-markers were observed. Considering correlations with R-values <-0.2 and > 0.2; 1) FAP-1 was positively associated with FOXP3 (R = 0.231); 2) PDGFRα was positively associated with CD3 (R = 0.236) and CD20 (R = 0.201) and 3) PDGFRβ was positively associated with FOXP3 (R = 0.221). Other significant, but weak (R < 0.2) associations between CAF- and immune-markers was observed. Correlations in histological subgroups were mostly similar and similar to those of the overall cohort (S2C and S2D Fig). Images of TMA cores representing tumors with different scores of CD3+ and CD8+ cells are shown in S1 Fig.
Survival analyses
Fig 4 and S3 Fig shows the survival curves of PDGFRα, PDGFRβ, FAP-1 and αSMA in patients expressing high and low levels of CD8 and CD3 respectively. In univariate analyses, high expression of FAP-1 was a significant positive marker for survival in patients with high expression of CD8 (P = 0.013) and CD3 (P = 0.042), while high expression of PDGFRβ was a significant negative marker for survival in patients with low expression of CD8 (P = 0.005) and near significant in patients with low expression of CD3 (P = 0.052).
Fig 4.
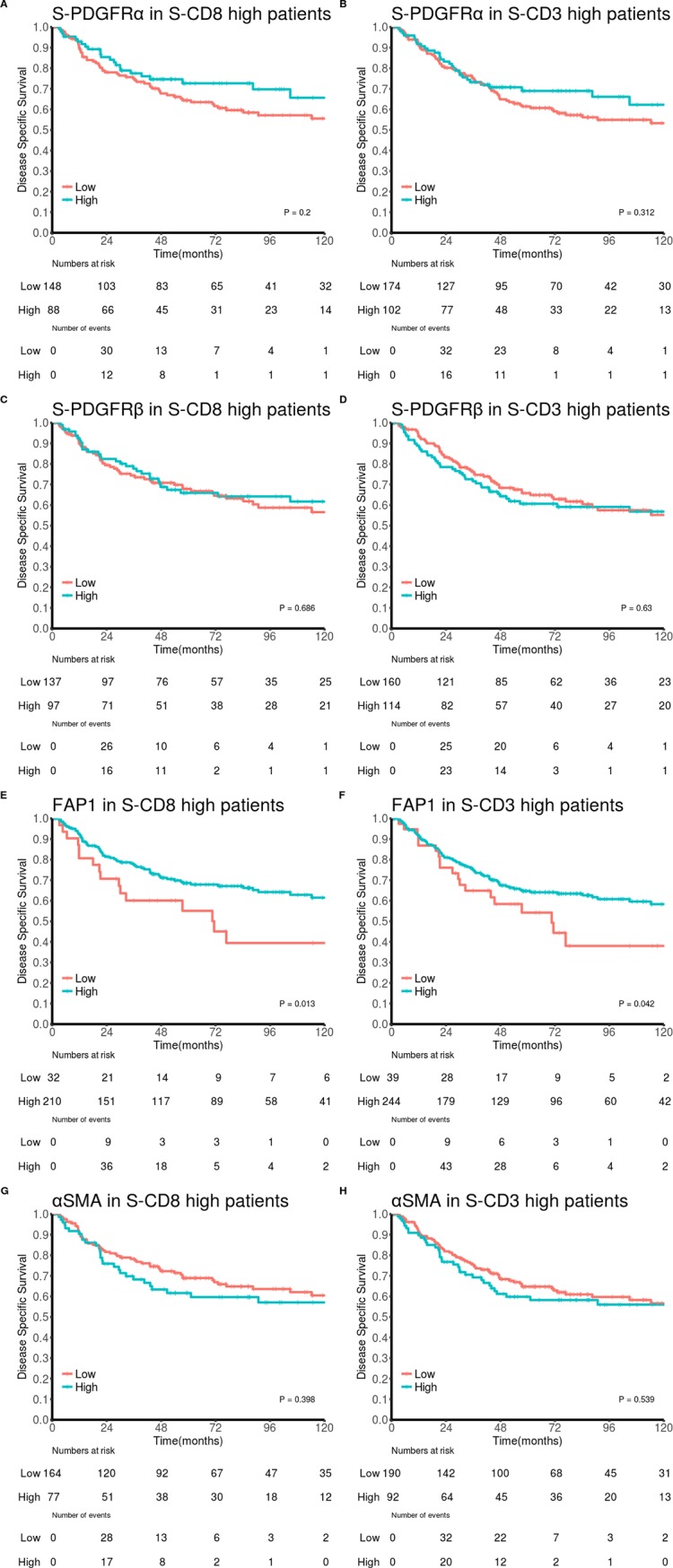
Disease-specific survival curves for: PDGFRα in patients expressing high levels of A) CD8 and B) CD3, PDGFRβ in patients expressing high levels of C) CD8 and D) CD3, FAP-1 in patients expressing high levels of E) CD8 and F) CD3 and αSMA in patients expressing high levels of G) CD8 and H) CD3. Abbreviations: FAP-1, Fibroblast activating protein 1; PDGFR, platelet-derived growth factor receptor; αSMA, alpha-smooth muscle actin.
In multivariable analyses, summarized in Table 3, FAP-1 was an independent positive marker for survival in patients with high, but not low, expression of CD8 (HR 0.42, 95% CI 0.24–0.74, P = 0.003, Table 3A) and PDGFRβ was an independent negative marker of survival in patients with low, but not high, expression of CD8 (HR 1.85, 95% CI 1.28–2.66, P < 0.001, Table 3E). Neither FAP-1, nor PDGFRβ were independent markers for survival in patients with high or low expression of CD3 (Table 3C, 3D, 3G and 3H).
Table 3. Multivariable models of FAP1 (A-D) and PDGFRβ (E-H) predicting survival of NSCLC patients with high and low expression of CD8 (A, C, E and G) and CD3 (B D, F and H, Cox regression analysis).
| (A) CD8 High | (B) CD3 High | (C) CD8 Low | (D) CD3 Low | |||||
|
HR (95% CI) |
P |
HR (95% CI) |
P |
HR (95% CI) |
P |
HR (95% CI) |
P | |
| FAP1 | ||||||||
| Low | 1 | 1 | 1 | 1 | ||||
| High | 0.42(0.24–0.74) | 0.003 | 0.75(0.45–1.25) | 0.266 | 0.99(0.65–1.49) | 0.954 | 0.84(0.54–1.31) | 0.452 |
| Gender | ||||||||
| Female | 1 | 1 | 1 | 1 | ||||
| Male | 2.3(1.37–3.85) | 0.002 | 1.84(1.18–2.84) | 0.007 | 1.48(0.98–2.23) | 0.063 | 1.3(0.79–2.12) | 0.302 |
| Histology | ||||||||
| SCC | 1 | 1 | 1 | 1 | ||||
| ADC | 3.13(1.91–5.16) | <0.001 | 2.25(1.45–3.48) | <0.001 | 1.16(0.78–1.73) | 0.451 | 1.11(0.72–1.71) | 0.647 |
| NOS | 1.92(0.76–4.88) | 0.170 | 2.24(1.12–4.48) | 0.022 | 0.96(0.51–1.8) | 0.894 | 0.56(0.24–1.27) | 0.166 |
| ECOG | ||||||||
| Normal | 1 | 1 | 1 | 1 | ||||
| Slightly reduced | 1.98(1.21–3.23) | 0.006 | 1.36(0.89–2.08) | 0.157 | 1.29(0.9–1.87) | 0.171 | 1.53(1.02–2.29) | 0.040 |
| In bed <50% | 2.92(1.11–7.65) | 0.029 | 1.86(0.83–4.2) | 0.133 | 1.36(0.62–2.99) | 0.447 | 1.08(0.41–2.82) | 0.880 |
| P-stage | ||||||||
| IA | 1 | 1 | 1 | 1 | ||||
| IB | 1.25(0.55–2.83) | 0.596 | 1.39(0.7–2.74) | 0.345 | 1.16(0.64–2.11) | 0.616 | 1.08(0.54–2.15) | 0.836 |
| IIA | 2.16(1.05–4.45) | 0.036 | 1.95(1.03–3.7) | 0.041 | 1.37(0.74–2.57) | 0.320 | 1.69(0.86–3.33) | 0.130 |
| IIB | 1.6(0.58–4.4) | 0.361 | 1.48(0.64–3.41) | 0.354 | 2.93(1.69–5.09) | <0.001 | 4.17(2.29–7.59) | <0.001 |
| IIIA | 5.3(2.49–11.28) | <0.001 | 4.36(2.28–8.3) | <0.001 | 4.2(2.34–7.53) | <0.001 | 6.68(3.43–12.98) | <0.001 |
| Vascular invasion | ||||||||
| No | 1 | 1 | 1 | 1 | ||||
| Yes | 2.84(1.61–5) | <0.001 | 2.14(1.34–3.44) | 0.002 | 1.57(1.03–2.41) | 0.037 | 1.67(1–2.78) | 0.048 |
| (E) CD8 High | (F) CD3 High | (G) CD8 Low | (H) CD3 Low | |||||
|
HR (95% CI) |
P |
HR (95% CI) |
P |
HR (95% CI) |
P |
HR (95% CI) |
P | |
| PDGFRβ | ||||||||
| Low | 1 | 1 | 1 | 1 | ||||
| High | 0.89(0.55–1.44) | 0.626 | 1.17(0.77–1.79) | 0.464 | 1.85(1.28–2.66) | <0.001 | 1.45(0.97–2.17) | 0.071 |
| Gender | ||||||||
| Female | 1 | 1 | ||||||
| Male | 2.19(1.29–3.74) | 0.004 | 1.8(1.15–2.8) | 0.010 | ||||
| Histology | ||||||||
| SCC | 1 | 1 | ||||||
| ADC | 2.84(1.73–4.67) | <0.001 | 2.22(1.42–3.46) | <0.001 | ||||
| NOS | 1.36(0.5–3.68) | 0.546 | 1.93(0.95–3.93) | 0.069 | ||||
| ECOG | ||||||||
| Normal | 1 | 1 | ||||||
| Slightly reduced | 1.88 (1.12–3.15) | 0.017 | 1.29(0.83–2.02) | 0.254 | ||||
| In bed <50% | 2.51 (0.97–6.52) | 0.059 | 1.76(0.77–4) | 0.179 | ||||
| P-stage | ||||||||
| IA | 1 | 1 | 1 | 1 | ||||
| IB | 1.33 (0.59–3.04) | 0.493 | 1.45 (0.73–2.89) | 0.292 | 1.17 (0.64–2.14) | 0.610 | 1.09 (0.55–2.2) | 0.798 |
| IIA | 2.36 (1.15–4.85) | 0.020 | 2.05 (1.08–3.91) | 0.029 | 1.42 (0.77–2.64) | 0.261 | 1.75 (0.9–3.43) | 0.102 |
| IIB | 1.92(0.69–5.31) | 0.212 | 1.67(0.72–3.87) | 0.228 | 2.91(1.67–5.05) | <0.001 | 3.92(2.12–7.26) | <0.001 |
| IIIA | 5.63(2.64–12.02) | <0.001 | 4.52(2.36–8.66) | <0.001 | 4.19(2.35–7.45) | <0.001 | 6.3(3.29–12.06) | <0.001 |
| Vascular invasion | ||||||||
| No | ||||||||
| Yes | 2.36(1.31–4.26) | 0.004 | 2.01(1.23–3.31) | 0.006 | 1.47(0.96–2.27) | 0.078 | 1.53(0.93–2.53) | 0.097 |
Abbreviations: FAP-1, Fibroblast activating protein 1; NSCLC, non small-cell lung cancer; CD8, cluster of differentiation 8; ECOG, Eastern Cooperative Oncology Group.
Discussion
A major finding from this in situ study is that the levels of CAFs did not correlate markedly with the infiltration of major leukocyte subsets into NSCLC tumor tissue (Fig 2B), indicating that CAFs may not play a dominant role in the regulation of leukocyte recruitment/infiltration in these tumors. Interestingly, survival analyses show that high levels of CAFFAP in CD3/CD8 infiltrated tumors correlate with increased patient survival. This finding may suggest that CAFFAP positively influence the effector function of cytotoxic tumor infiltrating lymphocytes.
The utilization of a TMA approach, rather than whole tissue slides, may partly explain the lack of correlations between CAFs and leukocyte subsets. However, the potential negative effect of using TMA in this context should be negated by the sheer number of patients included in the study and the inclusion of duplicate cores from each patient leading to increased representativeness. In addition, the presence of phenotypically different subsets of CAFs may differ between tumor regions due to heterogeneity. Neither TMA, nor whole slide studies are able to fully address this issue. Double, triple or quadruple IHC staining would ensure intra-patient sample homogeneity and would allow co-localization of CAF markers on the same cells. However, the issue of inter-patient sample heterogeneity, due to structurally and functionally different tumor areas being investigated in each patient, will remain a problem. Moreover, the spatial organization of the immune cell infiltrate, which discriminates between peritumoral and intratumoral areas is not addressed in TMA-based analyses. This is a relevant point that should be considered at the time of interpreting the results from this study. In this work, we lack information on peritumoral areas or the invasive front of tumors. However, we have considered the distinction between stromal and intratumoral areas.
Tumor fibroblasts, or CAFs, refers to a heterogeneous population of mesenchymal cells occurring during tumor development. Each subclass express different sets of cellular markers, and probably exert different regulatory functions in the context of cancer [1]. Despite the well-defined set of markers to identify mesenchymal cells, it is not possible to differentiate CAF subtypes based merely on the overlapping or non-overlapping expression of these markers [35]. Hence, we selected a panel of frequently used markers of fibroblast activation, and analyzed them separately. Initially, we checked for correlations between CAF-markers to study CAF diversity in NSCLC (Table 1, Fig 2A). Notably, vimentin, PDGFRα, PDGFRβ and FAP-1 showed modest correlation and did not correlate with αSMA and MT (collagen). Hierarchical clustering indicate the existence of different CAF subsets. Intriguingly the expression patterns of FAP-1 and αSMA, two of the most frequently used markers of CAF activation, showed distinct expression patterns when the staining was conducted in the same tumor specimen (Fig 3), clearly demonstrating the existence of phenotypically different CAF subsets in NSCLC tumors. Additionally, FAP-1 was not exclusively expressed by CAFs in tumors; since positive expression was also found in macrophage like cells, as previously observed for other tumor types [36]. However, our scoring was restricted to FAP-1 expression in fibroblast-like cells. In addition, PDGFRα expression by neoplastic epithelial cells was observed in about 15% of the cases.
In our cohort of unselected stage I-IIIA NSCLC patients, we have previously studied more than 100 prognostic markers related to in situ immunology [37–39], angiogenesis [30, 34], and epithelial-mesenchymal transition [10]. In this study, we have capitalized on these previous analyses and checked for correlations between CAF-markers and a panel of different leukocyte markers in the same patient cohort. Overall, the different CAF-markers do not show strong correlations with any of the selected myeloid or lymphoid immune markers, which included CD3, CD4, CD8 (T-lymphocytes), CD20 (B-lymphocytes), CD68 (TAMs), CD1a (dendritic cells DCs), CD56 (NK cells), FoxP3 (Treg) and CD45RO (T-memory cells). Moderate positive correlations were observed between CAFFAP and CAFPDGFRβ with FoxP3, and between CAFPDGFRα and CD3 or CD20 (Fig 2B). The positive associations of CAF subsets with T regulatory cells highlight a potential CAF-mediated immuno-regulatory mechanism connected to the chemotaxis of immuno-suppressive cells.
The interplay between CAFs and immune cells has been recognized as a major contributor in cancer development as summarized in recent reviews [23, 40]. CAFs may influence trafficking and function of effector immune cells indirectly by modulating ECM deposition and stiffness, via the synthesis of ECM components such as collagens, fibronectin and laminin, or by the release of matrix remodeling agents including MMPs, cathepsins or proteases of the uPA system [41, 42]. In our study we have identified correlations between CAFαSMA and collagen deposition in tumors (by MT), indicating that CAFs expressing αSMA (myofibroblasts) may be the main cells responsible for ECM formation in NSCLC tumors. However, no clear correlations were found between MT or CAFαSMA patterns and the infiltration of immune cells.
According to numerous pre-clinical studies, the pleiotropic immune-modulatory functions of CAFs are orchestrated primarily through the production of a plethora of cytokines, chemokines and small molecules functioning in a paracrine fashion [43, 44]. Moreover, CAFs have been shown to regulate recruitment and polarization of innate immune cells such as macrophages and neutrophils via secretion of factors like CCL5, MCP-1/CCL2, IL-6, IL-8, CXCL10 and CXCL14 [18, 45]. In the presented study, results do not reveal clear associations between CAFs and intratumoral levels of either TAMs (CD68) or DCs (CD1a). Whether CAFs influence the functional polarization of TAMs or DCs in vivo, remains to be elucidated. In future studies, the phenotypic stage of these cells should be established by the use of differentiation markers such as CD206 (M2 macrophages), iNOS (M1 macrophages) or CD303 (plasmacytoid DC).
Furthermore, CAFs regulate recruitment and activation of adaptive immune cells via the release of peptide signals such as transforming growth factor β1 (TGFβ1), tumor necrosis factor-α (TNFα), stromal-derived factor 1 (SDF-1), or thymic stromal lymphopoietin (TSLP) [46–49]. In our NSCLC patient cohort, we have investigated associations between CAF subtypes and lymphoid cells including T-cells (CD3+), CD4+ T-helper cells, CD8+ cytotoxic lymphocytes, memory T-cells (CD45RO), T-regulatory cells (FoxP3+), B-lymphocytes (CD20+) and NK cells (CD56+). No relevant correlations were observed between CAFs and adaptive immune cells. However, with the intention of checking CAF influence on T-cell effector functions, survival rates in patients with high or low infiltration rates of CD3+ and CD8+ T-cells were compared. Intriguingly, in the subgroup of patients with tumors highly infiltrated by lymphocytes, the presence of CAFFAP was a positive prognostic factor. This observation suggests that CAFFAP may—directly or indirectly—exert positive immuno-adjuvant effects. On the contrary, increased CAFPDGFRβ levels were associated with adverse prognosis in patients with tumors poorly infiltrated by T-lymphocytes (S3 Fig). This latter finding indicate that CAFPDGFRβ exerts no influence on the tumor immune response. Combined, the results of CAFFAP and CAFPDGFRβ support the theory of functionally different and/or context-dependent CAF subclasses exerting divergent immune-modulatory effects in NSCLC, despite the assumption that both FAP-1 and PDGFRβ are considered specific markers for activated CAFs. FAP-1, also known as seprase, is a ubiquitously expressed membrane bound serine protease that has both dipeptidyl peptidase and endopeptidase activities, cleaving substrates at post-proline bonds. FAP-1 is essentially undetectable in non-disease tissues, but greatly enhanced at sites of tissue remodeling, including fibrotic tissue reactions, chronically inflamed tissues, epithelial cancers and embryonic tissues. The role of this protease as regulator of the immune system is uncertain. However, selective depletion of FAP-1-positive fibroblasts in murine models of pancreatic cancer has been shown to enhance anti-tumor immunity and delay tumor growth [50].
Generally, CAFs have been considered to promote an immunosuppressive microenvironment, but this may be context dependent rather than a specific feature of CAFs. Most of the accumulated evidence in this regard are limited to in vitro experiments, or animal experiments with admixed fibroblasts previously expanded in vitro which do not recapitulate the CAF heterogeneity observed in vivo. Only a handful of studies, with diverging outcomes, have investigated immunoregulatory effects of CAFs in vivo by selective depletion of CAF subsets. In genetically engineered murine models, selective depletion of CAFFAP showed antitumor effects via intratumoral recruitment of CD8+ T cells [50]. On the contrary, specific depletion of CAFαSMA led to invasive tumors associated with an immunosuppressive adaptive response [25]. Besides, clinical studies in pancreas, breast and lung cancer that correlate ECM or CAF-markers with disease outcomes have shown that patients with high desmoplasia or CAF infiltration can have improved prognosis and overall survival [51]. The presented explorative study propose a beneficial prognostic role played by CAFFAP in immune infiltrated tumors, and underscores the need of caution in targeting CAFFAP as a therapeutic strategy in lung cancer. However, additional functional studies should be considered to conclusively demonstrate causal effects of CAF subtypes on the anti-tumor immune responses.
Supporting information
Immunostaining of TMA cores showing different scores for CD3 (A) and CD8 (B). Abbreviations: CD, Cluster of differentiation.
(TIFF)
Spearman's rank correlations between (A and B). Different CAF markers and (C and D) CAF markers and markers of leukocyte subsets in the adenocarcinoma (A and C) and squamous cell carcinoma (B and D) subgroups. *P < 0.05, **P < 0.01, ***P < 0.001. Abbreviations: CAF, cancer-associated fibroblast; Vim, vimentin; FAP-1, Fibroblast activation protein 1; PDGFR, platelet-derived growth factor receptor; αSMA, alpha-smooth muscle actin; MT, Masson's trichrome; CD, cluster of differentiation.
(TIFF)
Disease-specific survival curves for: PDGFRα in patients expressing low levels of A) CD8 and B) CD3, PDGFRβ in patients expressing low levels of C) CD8 and D) CD3, FAP-1 in patients expressing low levels of E) CD8 and F) CD3 and αSMA in patients expressing low levels of G) CD8 and H) CD3. Abbreviations: FAP-1, Fibroblast activating protein 1; PDGFR, platelet-derived growth factor receptor; αSMA, alpha-smooth muscle actin.
(TIFF)
(DOCX)
(CSV)
Acknowledgments
The authors thank pathologists Samer Al-Saad, Khalid Al-Shibli and Elin Richardsen for providing human lung tissue samples. The publication charges for this article have been funded by a grant from the publication fund of UiT The Arctic University of Norway.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by the Northern Norway Regional Health Authority with URL: https://helse-nord.no/ (grant no: SPP1137-13 to TH and SFP1138-13), the Norwegian Cancer Society with URL: https://kreftforeningen.no; (grant no: 9223 to IMZ) and Erna & Olav Aakre Foundation for Cancer Research to IMZ. The publication charges for this article have been funded by a grant from the publication fund of UiT The Arctic University of Norway. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Ostman A, Augsten M. Cancer-associated fibroblasts and tumor growth—bystanders turning into key players. Curr Opin Genet Dev. 2009;19(1):67–73. doi: 10.1016/j.gde.2009.01.003 [DOI] [PubMed] [Google Scholar]
- 2.Kalluri R, Zeisberg M. Fibroblasts in cancer. Nat Rev Cancer. 2006;6(5):392–401. doi: 10.1038/nrc1877 [DOI] [PubMed] [Google Scholar]
- 3.Darby IA, Laverdet B, Bonte F, Desmouliere A. Fibroblasts and myofibroblasts in wound healing. Clinical, cosmetic and investigational dermatology. 2014;7:301–11. doi: 10.2147/CCID.S50046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ohlund D, Elyada E, Tuveson D. Fibroblast heterogeneity in the cancer wound. J Exp Med. 2014;211(8):1503–23. doi: 10.1084/jem.20140692 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kalluri R. The biology and function of fibroblasts in cancer. Nat Rev Cancer. 2016;16(9):582–98. doi: 10.1038/nrc.2016.73 [DOI] [PubMed] [Google Scholar]
- 6.Cortez E, Roswall P, Pietras K. Functional subsets of mesenchymal cell types in the tumor microenvironment. Seminars in cancer biology. 2014;25:3–9. doi: 10.1016/j.semcancer.2013.12.010 [DOI] [PubMed] [Google Scholar]
- 7.Chen Y, Zou L, Zhang Y, Chen Y, Xing P, Yang W, et al. Transforming growth factor-beta1 and alpha-smooth muscle actin in stromal fibroblasts are associated with a poor prognosis in patients with clinical stage I-IIIA nonsmall cell lung cancer after curative resection. Tumour Biol. 2014;35(7):6707–13. doi: 10.1007/s13277-014-1908-y [DOI] [PubMed] [Google Scholar]
- 8.Kilvaer TK, Khanehkenari MR, Hellevik T, Al-Saad S, Paulsen EE, Bremnes RM, et al. Cancer Associated Fibroblasts in Stage I-IIIA NSCLC: Prognostic Impact and Their Correlations with Tumor Molecular Markers. PLoS One. 2015;10(8):e0134965 doi: 10.1371/journal.pone.0134965 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Shu H, Li HF. Prognostic effect of stromal myofibroblasts in lung adenocarcinoma. Neoplasma. 2012;59(6):658–61. doi: 10.4149/neo_2012_083 [DOI] [PubMed] [Google Scholar]
- 10.Al-Saad S, Al-Shibli K, Donnem T, Persson M, Bremnes RM, Busund LT. The prognostic impact of NF-kappaB p105, vimentin, E-cadherin and Par6 expression in epithelial and stromal compartment in non-small-cell lung cancer. Br J Cancer. 2008;99(9):1476–83. doi: 10.1038/sj.bjc.6604713 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ito M, Ishii G, Nagai K, Maeda R, Nakano Y, Ochiai A. Prognostic impact of cancer-associated stromal cells in patients with stage I lung adenocarcinoma. Chest. 2012;142(1):151–8. doi: 10.1378/chest.11-2458 [DOI] [PubMed] [Google Scholar]
- 12.Kawase A, Ishii G, Nagai K, Ito T, Nagano T, Murata Y, et al. Podoplanin expression by cancer associated fibroblasts predicts poor prognosis of lung adenocarcinoma. Int J Cancer. 2008;123(5):1053–9. doi: 10.1002/ijc.23611 [DOI] [PubMed] [Google Scholar]
- 13.Koriyama H, Ishii G, Yoh K, Neri S, Morise M, Umemura S, et al. Presence of podoplanin-positive cancer-associated fibroblasts in surgically resected primary lung adenocarcinoma predicts a shorter progression-free survival period in patients with recurrences who received platinum-based chemotherapy. Journal of cancer research and clinical oncology. 2015;141(7):1163–70. doi: 10.1007/s00432-014-1891-0 [DOI] [PubMed] [Google Scholar]
- 14.Neri S, Ishii G, Taira T, Hishida T, Yoshida J, Nishimura M, et al. Recruitment of podoplanin positive cancer-associated fibroblasts in metastatic lymph nodes predicts poor prognosis in pathological N2 stage III lung adenocarcinoma. Ann Surg Oncol. 2012;19(12):3953–62. doi: 10.1245/s10434-012-2421-4 [DOI] [PubMed] [Google Scholar]
- 15.Takahashi A, Ishii G, Neri S, Yoshida T, Hashimoto H, Suzuki S, et al. Podoplanin-expressing cancer-associated fibroblasts inhibit small cell lung cancer growth. Oncotarget. 2015;6(11):9531–41. doi: 10.18632/oncotarget.3371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kitano H, Kageyama S, Hewitt SM, Hayashi R, Doki Y, Ozaki Y, et al. Podoplanin expression in cancerous stroma induces lymphangiogenesis and predicts lymphatic spread and patient survival. Arch Pathol Lab Med. 2010;134(10):1520–7. doi: 10.1043/2009-0114-OA.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ono S, Ishii G, Nagai K, Takuwa T, Yoshida J, Nishimura M, et al. Podoplanin-positive cancer-associated fibroblasts could have prognostic value independent of cancer cell phenotype in stage I lung squamous cell carcinoma: usefulness of combining analysis of both cancer cell phenotype and cancer-associated fibroblast phenotype. Chest. 2013;143(4):963–70. doi: 10.1378/chest.12-0913 [DOI] [PubMed] [Google Scholar]
- 18.Liao D, Luo Y, Markowitz D, Xiang R, Reisfeld RA. Cancer associated fibroblasts promote tumor growth and metastasis by modulating the tumor immune microenvironment in a 4T1 murine breast cancer model. PLoS One. 2009;4(11):e7965 doi: 10.1371/journal.pone.0007965 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ishikawa S, Takenaka K, Yanagihara K, Miyahara R, Kawano Y, Otake Y, et al. Matrix metalloproteinase-2 status in stromal fibroblasts, not in tumor cells, is a significant prognostic factor in non-small-cell lung cancer. Clin Cancer Res. 2004;10(19):6579–85. doi: 10.1158/1078-0432.CCR-04-0272 [DOI] [PubMed] [Google Scholar]
- 20.Leinonen T, Pirinen R, Bohm J, Johansson R, Kosma VM. Increased expression of matrix metalloproteinase-2 (MMP-2) predicts tumour recurrence and unfavourable outcome in non-small cell lung cancer. Histol Histopathol. 2008;23(6):693–700. doi: 10.14670/HH-23.693 [DOI] [PubMed] [Google Scholar]
- 21.Edlund K, Lindskog C, Saito A, Berglund A, Ponten F, Goransson-Kultima H, et al. CD99 is a novel prognostic stromal marker in non-small cell lung cancer. Int J Cancer. 2012;131(10):2264–73. doi: 10.1002/ijc.27518 [DOI] [PubMed] [Google Scholar]
- 22.Turley SJ, Cremasco V, Astarita JL. Immunological hallmarks of stromal cells in the tumour microenvironment. Nature reviews Immunology. 2015;15(11):669–82. doi: 10.1038/nri3902 [DOI] [PubMed] [Google Scholar]
- 23.Servais C, Erez N. From sentinel cells to inflammatory culprits: cancer-associated fibroblasts in tumour-related inflammation. The Journal of pathology. 2013;229(2):198–207. doi: 10.1002/path.4103 [DOI] [PubMed] [Google Scholar]
- 24.Silzle T, Randolph GJ, Kreutz M, Kunz-Schughart LA. The fibroblast: sentinel cell and local immune modulator in tumor tissue. Int J Cancer. 2004;108(2):173–80. doi: 10.1002/ijc.11542 [DOI] [PubMed] [Google Scholar]
- 25.Ozdemir BC, Pentcheva-Hoang T, Carstens JL, Zheng X, Wu CC, Simpson TR, et al. Depletion of carcinoma-associated fibroblasts and fibrosis induces immunosuppression and accelerates pancreas cancer with reduced survival. Cancer Cell. 2014;25(6):719–34. doi: 10.1016/j.ccr.2014.04.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Feig C, Jones JO, Kraman M, Wells RJ, Deonarine A, Chan DS, et al. Targeting CXCL12 from FAP-expressing carcinoma-associated fibroblasts synergizes with anti-PD-L1 immunotherapy in pancreatic cancer. Proc Natl Acad Sci U S A. 2013;110(50):20212–7. doi: 10.1073/pnas.1320318110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Saito RA, Micke P, Paulsson J, Augsten M, Pena C, Jonsson P, et al. Forkhead box F1 regulates tumor-promoting properties of cancer-associated fibroblasts in lung cancer. Cancer Res. 2010;70(7):2644–54. doi: 10.1158/0008-5472.CAN-09-3644 [DOI] [PubMed] [Google Scholar]
- 28.Mattsson JS, Bergman B, Grinberg M, Edlund K, Marincevic M, Jirstrom K, et al. Prognostic impact of COX-2 in non-small cell lung cancer: a comprehensive compartment-specific evaluation of tumor and stromal cell expression. Cancer letters. 2015;356(2 Pt B):837–45. doi: 10.1016/j.canlet.2014.10.032 [DOI] [PubMed] [Google Scholar]
- 29.Kilvaer TK, Paulsen EE, Khanehkenari MR, Al-Saad S, Johansen RM, Al-Shibli K, et al. The presence of intraepithelial CD45RO+ cells in resected lymph nodes with metastases from NSCLC patients is an independent predictor of disease-specific survival. Br J Cancer. 2016;114(10):1145–51. doi: 10.1038/bjc.2016.92 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Donnem T, Al-Saad S, Al-Shibli K, Delghandi MP, Persson M, Nilsen MN, et al. Inverse prognostic impact of angiogenic marker expression in tumor cells versus stromal cells in non small cell lung cancer. Clin Cancer Res. 2007;13(22 Pt 1):6649–57. doi: 10.1158/1078-0432.CCR-07-0414 [DOI] [PubMed] [Google Scholar]
- 31.Detterbeck FC, Postmus PE, Tanoue LT. The stage classification of lung cancer: Diagnosis and management of lung cancer, 3rd ed: American College of Chest Physicians evidence-based clinical practice guidelines. Chest. 2013;143(5 Suppl):e191S–e210S. [DOI] [PubMed] [Google Scholar]
- 32.Bremnes RM, Veve R, Gabrielson E, Hirsch FR, Baron A, Bemis L, et al. High-throughput tissue microarray analysis used to evaluate biology and prognostic significance of the E-cadherin pathway in non-small-cell lung cancer. J Clin Oncol. 2002;20(10):2417–28. doi: 10.1200/JCO.2002.08.159 [DOI] [PubMed] [Google Scholar]
- 33.Al-Shibli K, Al-Saad S, Donnem T, Persson M, Bremnes RM, Busund LT. The prognostic value of intraepithelial and stromal innate immune system cells in non-small cell lung carcinoma. Histopathology. 2009;55(3):301–12. doi: 10.1111/j.1365-2559.2009.03379.x [DOI] [PubMed] [Google Scholar]
- 34.Donnem T, Al-Saad S, Al-Shibli K, Busund LT, Bremnes RM. Co-expression of PDGF-B and VEGFR-3 strongly correlates with lymph node metastasis and poor survival in non-small-cell lung cancer. Annals of oncology: official journal of the European Society for Medical Oncology / ESMO. 2010;21(2):223–31. [DOI] [PubMed] [Google Scholar]
- 35.Sugimoto H, Mundel TM, Kieran MW, Kalluri R. Identification of fibroblast heterogeneity in the tumor microenvironment. Cancer Biol Ther. 2006;5(12):1640–6. [DOI] [PubMed] [Google Scholar]
- 36.Tchou J, Zhang PJ, Bi Y, Satija C, Marjumdar R, Stephen TL, et al. Fibroblast activation protein expression by stromal cells and tumor-associated macrophages in human breast cancer. Human pathology. 2013;44(11):2549–57. doi: 10.1016/j.humpath.2013.06.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Paulsen EE, Kilvaer T, Khanehkenari MR, Maurseth RJ, Al-Saad S, Hald SM, et al. CD45RO(+) Memory T Lymphocytes—a Candidate Marker for TNM-Immunoscore in Squamous Non-Small Cell Lung Cancer. Neoplasia. 2015;17(11):839–48. doi: 10.1016/j.neo.2015.11.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Al-Shibli KI, Donnem T, Al-Saad S, Persson M, Bremnes RM, Busund LT. Prognostic effect of epithelial and stromal lymphocyte infiltration in non-small cell lung cancer. Clin Cancer Res. 2008;14(16):5220–7. doi: 10.1158/1078-0432.CCR-08-0133 [DOI] [PubMed] [Google Scholar]
- 39.Rakaee M, Busund LT, Paulsen EE, Richardsen E, Al-Saad S, Andersen S, et al. Prognostic effect of intratumoral neutrophils across histological subtypes of non-small cell lung cancer. Oncotarget. 2016;7(44):72184–96. doi: 10.18632/oncotarget.12360 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Harper J, Sainson RC. Regulation of the anti-tumour immune response by cancer-associated fibroblasts. Seminars in cancer biology. 2014;25:69–77. doi: 10.1016/j.semcancer.2013.12.005 [DOI] [PubMed] [Google Scholar]
- 41.Salmon H, Franciszkiewicz K, Damotte D, Dieu-Nosjean MC, Validire P, Trautmann A, et al. Matrix architecture defines the preferential localization and migration of T cells into the stroma of human lung tumors. J Clin Invest. 2012;122(3):899–910. doi: 10.1172/JCI45817 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kunz-Schughart LA, Knuechel R. Tumor-associated fibroblasts (part II): Functional impact on tumor tissue. Histol Histopathol. 2002;17(2):623–37. doi: 10.14670/HH-17.623 [DOI] [PubMed] [Google Scholar]
- 43.Erez N, Truitt M, Olson P, Arron ST, Hanahan D. Cancer-Associated Fibroblasts Are Activated in Incipient Neoplasia to Orchestrate Tumor-Promoting Inflammation in an NF-kappaB-Dependent Manner. Cancer Cell. 2010;17(2):135–47. doi: 10.1016/j.ccr.2009.12.041 [DOI] [PubMed] [Google Scholar]
- 44.Hellevik T, Pettersen I, Berg V, Bruun J, Bartnes K, Busund LT, et al. Changes in the Secretory Profile of NSCLC-Associated Fibroblasts after Ablative Radiotherapy: Potential Impact on Angiogenesis and Tumor Growth. Translational oncology. 2013;6(1):66–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.De Boeck A, Hendrix A, Maynard D, Van Bockstal M, Daniels A, Pauwels P, et al. Differential secretome analysis of cancer-associated fibroblasts and bone marrow-derived precursors to identify microenvironmental regulators of colon cancer progression. Proteomics. 2013;13(2):379–88. doi: 10.1002/pmic.201200179 [DOI] [PubMed] [Google Scholar]
- 46.Barnas JL, Simpson-Abelson MR, Yokota SJ, Kelleher RJ, Bankert RB. T cells and stromal fibroblasts in human tumor microenvironments represent potential therapeutic targets. Cancer microenvironment: official journal of the International Cancer Microenvironment Society. 2010;3(1):29–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kojima Y, Acar A, Eaton EN, Mellody KT, Scheel C, Ben-Porath I, et al. Autocrine TGF-beta and stromal cell-derived factor-1 (SDF-1) signaling drives the evolution of tumor-promoting mammary stromal myofibroblasts. Proc Natl Acad Sci U S A. 2010;107(46):20009–14. doi: 10.1073/pnas.1013805107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Nazareth MR, Broderick L, Simpson-Abelson MR, Kelleher RJ Jr., Yokota SJ, Bankert RB. Characterization of human lung tumor-associated fibroblasts and their ability to modulate the activation of tumor-associated T cells. Journal of immunology. 2007;178(9):5552–62. [DOI] [PubMed] [Google Scholar]
- 49.De Monte L, Reni M, Tassi E, Clavenna D, Papa I, Recalde H, et al. Intratumor T helper type 2 cell infiltrate correlates with cancer-associated fibroblast thymic stromal lymphopoietin production and reduced survival in pancreatic cancer. J Exp Med. 2011;208(3):469–78. doi: 10.1084/jem.20101876 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kraman M, Bambrough PJ, Arnold JN, Roberts EW, Magiera L, Jones JO, et al. Suppression of antitumor immunity by stromal cells expressing fibroblast activation protein-alpha. Science. 2010;330(6005):827–30. doi: 10.1126/science.1195300 [DOI] [PubMed] [Google Scholar]
- 51.Paulsson J, Micke P. Prognostic relevance of cancer-associated fibroblasts in human cancer. Seminars in cancer biology. 2014;25:61–8. doi: 10.1016/j.semcancer.2014.02.006 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Immunostaining of TMA cores showing different scores for CD3 (A) and CD8 (B). Abbreviations: CD, Cluster of differentiation.
(TIFF)
Spearman's rank correlations between (A and B). Different CAF markers and (C and D) CAF markers and markers of leukocyte subsets in the adenocarcinoma (A and C) and squamous cell carcinoma (B and D) subgroups. *P < 0.05, **P < 0.01, ***P < 0.001. Abbreviations: CAF, cancer-associated fibroblast; Vim, vimentin; FAP-1, Fibroblast activation protein 1; PDGFR, platelet-derived growth factor receptor; αSMA, alpha-smooth muscle actin; MT, Masson's trichrome; CD, cluster of differentiation.
(TIFF)
Disease-specific survival curves for: PDGFRα in patients expressing low levels of A) CD8 and B) CD3, PDGFRβ in patients expressing low levels of C) CD8 and D) CD3, FAP-1 in patients expressing low levels of E) CD8 and F) CD3 and αSMA in patients expressing low levels of G) CD8 and H) CD3. Abbreviations: FAP-1, Fibroblast activating protein 1; PDGFR, platelet-derived growth factor receptor; αSMA, alpha-smooth muscle actin.
(TIFF)
(DOCX)
(CSV)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.