

Abstract
Background
There is evidence that suggests that undernutrition has a detrimental effect on malarial immunity in children. The aim of the study was to discover whether nutrient supplementation improved development of malarial antibody immunity in children up to 18 months of age.
Methods
The study was conducted with a subset of 432 Malawian children from a randomized controlled trial of nutritional supplements. The arms included pre- and postnatal small-quantity lipid-based nutrient supplements for both mother and child; prenatal supplementation with iron and folic acid; and pre- and postnatal supplementation with multiple micronutrients. Paired plasma samples were collected at 6 and 18 months of age. The levels of antibodies against merozoite surface protein 1 (MSP1 19kD) and MSP2, erythrocyte binding antigen 175 (EBA175), reticulocyte binding protein homologue 2A (Rh2A9), schizont extract and variant antigens expressed on the surface of infected erythrocytes were measured.
Results
At 18 months of age, 5.4% of children were parasitaemic by microscopy and 49.1% were anaemic. Antibodies to the tested merozoite antigens and schizont extract increased between 6 and 18 months and this increase was statistically significant for MSP1, MSP2 and EBA175 (p < 0.0001) whereas IgG to variant surface antigens decreased with increasing age (p < 0.0001). However, the supplementation type did not have any impact on the prevalence or levels of antibodies at either 6 or 18 months of age to any of the tested malaria antigens in either univariate analysis or multivariate analysis after adjusting for covariates.
Conclusions
Pre- and postnatal lipid-based nutrient supplementation did not alter malaria antibody acquisition during infancy, compared to prenatal supplementation with iron and folic acid or pre- and postnatal supplementation with multiple micronutrients.
Trail registeration Clinicaltrials.gov registration number NCT01239693
Keywords: Malarial immunity in children, Nutrient supplements, Randomized controlled trial, Merozoite antigens, Variant surface antigens, Seroprevalence
Background
Malaria is one of the leading causes of death in children and pregnant women with an estimated 214 million new cases and 438,000 deaths worldwide in 2015. The disease can be caused by five different species of the genus Plasmodium, of which Plasmodium falciparum causes the highest rates of mortality and morbidity and is particularly prominent in young children of sub-Saharan Africa, with an estimated 292,000 deaths in 2015 [1].
In sub-Saharan Africa, malaria and malnutrition often co-exist, and both contribute significantly to deaths in young children. However, studies of possible synergistic clinical effects of malaria and malnutrition have given conflicting results, indicating the need for further studies in this area. For example, in a cross-sectional study among pre-school Kenyan children [2] and a longitudinal study in Gambian children under 5 years of age [3], stunting was associated with increased malarial risk, but in Papua New Guinea it was reported that stunting might protect children against clinical malaria episodes [4]. Some other studies noted no significant association between anthropometric measurements [5], stunting [6] or undernutrition [7] and altered susceptibility to malaria.
A limited number of studies have examined the impact of nutrient supplementation on malaria susceptibility in children. Zinc and vitamin A supplementation reduced clinical malaria episodes caused by P. falciparum in young children [8–10]. In a high malaria transmission setting, iron supplementation was associated with increased parasitaemia [11] and increased mortality [12] in iron-sufficient children, whereas the provision of iron with micronutrients was associated with reduced risk of malaria in iron-deficient children [13]. Other studies have found evidence of associations between acute malaria and deficiency of thiamine [14] and antioxidants including vitamin E [15], which suggests they have roles in protection against malaria. While there is limited evidence that supplementation with micronutrients such as zinc or vitamin B12 can improve antibody response to vaccination [16, 17], the ability of micro- or macronutrient supplementation to affect the acquisition of antibody to pathogens following natural exposure is unknown.
The aim of this study was to identify whether pre- and postnatal nutritional supplements could improve malarial immunity in young children. The study was part of a nutrient supplementation clinical trial, the International Lipid-based Nutrient Supplement (iLiNS) Project DYAD-Malawi trial (clinicaltrials.gov registration number NCT01239693). For this report, the level and prevalence of antibody to merozoite antigens, schizont extract and variant surface antigens (VSA) expressed by P. falciparum-infected erythrocytes (IEs) were determined in infants aged 6 and 18 months as antibodies to merozoite antigens and VSAs are believed to play important roles in mediating acquired immunity against malaria [18, 19].
Methods
Study location and participants
The study participants were a cohort of 432 infants residing in Lungwena, Malindi and Mangochi from rural Malawi who participated in the iLiNS Project DYAD-Malawi nutrient supplementation trial, part of the iLiNS Project [20]. The details of the trial design and supplements have been published elsewhere [21]. In brief, participating pregnant women were randomly allocated to receive iron and folic acid (IFA), multiple micronutrients (MMN) or a small-quantity (20 g) of lipid based nutrient supplement (LNS) daily. After delivery, women in the IFA group received placebo tablets, while MMN and LNS supplementation was continued during the first 6 months of lactation. Children of mothers in the LNS group also received LNS 10 g twice daily from 6 to 18 months of age. At 18 months of age, anthropometric assessments revealed no significant differences in the children’s mean length, mean weight, the prevalence of stunting, or head or mid-upper arm circumference between the intervention groups for all participants in the iLiNS project [22]. Plasma samples were collected from infants at 6 and 18 months of age. At these time points blood haemoglobin concentration was measured with a Hemo-Cue® haemoglobinometer from venous blood samples. Malaria parasitaemia was sought by microscopic examination of thick blood film and by rapid diagnostic test (RDT) using Clearview® Malaria Combo (British Biocell International Ltd., Dundee, UK).
Plasma samples preparation
Blood from participants was separated by centrifugation shortly after collection and plasma was stored at − 80 °C before shipment on dry ice to Australia. Plasma samples were heat-inactivated for 45 min at 57 °C to inactivate complement proteins. The heat-inactivated samples were then stored at − 80 °C.
Culturing and maintaining parasites
The P. falciparum lines used were E8B-ICAM, R29 and 3D7 varA over-expressing parasite line. E8B-ICAM adheres to ICAM-1 and CD36 [23], and expresses group B/C var genes whereas R29 expresses group A var genes and forms rosettes [24]. The 3D7 line spontaneously expressed a group A var gene as its dominant transcript [25]; its binding ligands have not been characterized. The parasites were grown and maintained in culture as described previously [26]. IEs were synchronized with 5% sorbitol and subject to gelatin flotation regularly [27]. To select R29 for rosetting, gelatin flotation without then with heparin lithium salt, 0.05 mg/ml Sigma Aldrich), was performed.
Measuring IgG to malaria merozoite antigens and schizont extract
Recombinant merozoite protein 1 (MSP-1 19 kD, 3D7 clone), region III-V of erythrocyte binding antigen 175 (EBA 175), and P. falciparum reticulocyte binding homologue 2 (PfRh2, construct PfRh2-2030) were expressed in Escherichia coli as previously reported [28–30]. Full-length MSP-2 (FC27 clone) expressed in E. coli was kindly provided by Robin Anders (La Trobe University, Australia). The schizont extract was prepared according to a previously published method [31]. Briefly, magnetic-activated cell sorting (MACS) purified schizont pellet was mixed with three times the volume of the pellet with PBS. Cell-lysis was done by freeze-thawing for 6 times, then it was spun down to clarify the supernatant and this extract was used after optimizing the coating concentration.
Each merozoite antigen was coated at 0.5–2 µg/ml and schizont extract at 1:8000 dilution on to 384 well NUNC MaxiSorp™ plates (Thermo Fisher Scientific Inc, MA, USA) and left overnight at 4 °C. The plates were washed with PBS/Tween 20 and non-specific binding was blocked with 0.1% casein (Thermo Fisher Scientific) on the following day. The plates were then incubated with participant sera diluted at 1:250 in 0.1% casein in triplicates for 1 h and washed. Horseradish peroxidase-conjugated goat anti-human IgG (Life Technologies, Australia) was added at 1:2500 dilution for 1 h and plates were again washed. ABTS [2,2′-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid)] was added as enzyme–substrate for 15 min and absorbance at 405 nm was measured using a BMG POLARstar Omega fluorimeter (BMG Labtech, Germany).
Measurement of total IgG levels against VSA
Total IgG antibody levels against VSAs expressed on the surface of IEs were measured by flow cytometry as previously described [32] with slight modifications. These antibodies are believed to primarily target P. falciparum erythrocyte membrane antigen 1 (PfEMP1) [33]. In brief, 2.5 μl of patient sera (1:20 dilution) were co-incubated with IEs at 0.2% haematocrit and at approximately 7–8% parasitaemia, diluted in PBS solution with 1% HI-FBS (Heat inactivated fetal bovine serum) for 30 min. Following incubation the cells were washed 3 times with PBS/1% HI-FBS and incubated with 25 μl of 1:100 rabbit anti-human IgG (Dako, Australia) diluted in PBS/1% HI-FBS. The cells were washed again in PBS/1% HI-FBS and incubated with Alexa Fluor 647 donkey anti-rabbit IgG in 1:500 dilution (Life Technologies, Australia) and 10 μg/ml ethidium bromide (EtBr) in PBS/1% HI-FBS for 30 min. Following incubation the cells were washed in PBS/1% HI-FBS and resuspended in ice-cold 2% paraformaldehyde fixative solution (prepared in PBS). The fixed IEs were then run through a HyperCyt® system with a plate reader adapter (Intellicyt®, NM, USA) connected to a Cyan flow cytometer (Beckman Coulter Inc., CA, USA) where the cells were acquired. The flow cytometry data was analysed according to a previously published method [32].
Data analysis
Statistical analyses were performed using Stata version 13.0 (StataCorp, Texas, USA). Statistical analyses were performed according to a pre-planned and approved analysis plan available at [34]. Measured antibody levels (in optical density [OD] for schizont and merozoite antigens and geometric mean fluorescence intensity [MFI] for VSA) were presented as a percentage of the positive control. The positive control came from a pool of plasma from malaria immune African adults whereas the negative control came from plasma samples from 3 Melbourne donors for the ELISA and 8 Melbourne donors for the flow cytometry assay.
Seroprevalence was defined as the percentage of the cohort having antibody responses greater than the mean antibody response plus three standard deviations for negative controls, which were malaria naïve samples from Melbourne blood donors. Socioeconomic status (SES) was calculated on the basis of a scoring system for household assets (HHA) adapted from [35].
Participant characteristics including demographic and basic clinical characteristics were categorized by intervention groups and the median and interquartile range for each characteristic were tabulated. Differences in characteristics across the groups were determined by Mann–Whitney test (non-parametric continuous variables with two groups), Kruskal–Wallis (non-parametric continuous variables with more than two groups) or Chi2 test (for categorical variables) where applicable. Differences in the antibody level and the seropositivity between 6 and 18 months old children were determined by Wilcoxon matched-pairs signed-ranks test and McNemar’s test, respectively. Statistical differences between the groups were reported as p < 0.05 and 95% confidence intervals were also reported for the analyses.
Antibody levels at 6 and 18 months of age were reported as the median percentage of the positive control and the interquartile range (IQR). The Kruskal–Wallis test was performed to compare antibody levels across supplementation groups. Regression analyses were carried out using natural logarithmically transformed antibody levels which were back transformed for reporting descriptive results. Linear regression univariate analysis was performed between LNS versus IFA, LNS versus MMN and MMN versus IFA to determine the antibody level differences between supplementation groups. Multivariate regression was also performed adjusting for the following covariates: maternal BMI at enrolment, duration of gestation (from enrolment to delivery), number of pregnancies, sex of the child, maternal education, proxy for SES, study site, maternal anaemic status at enrolment, maternal HIV status and bed net use by children and these covariates were uniformly included in all the adjusted analyses according to the pre-specified analysis plan. For both univariate and multivariate analyses, coefficients and 95% confidence intervals (CI) were reported. The number and the percentage of children who were seropositive for each malaria antigen were reported by supplementation groups, and chi2 test was performed to determine the differences across the supplementation groups. Univariate logistic regression was performed between LNS versus IFA, LNS versus MMN and MMN versus IFA to determine the differences in seropositivity between different supplementation groups. Multivariate logistic regression was performed adjusting for the above-mentioned covariates, reporting odds ratios (OR) and 95% CI.
Results
Study population characteristics
From a total of 1391 enrolled pregnant women recruited to the trial, 869 completed the intervention and follow-up to 18 months after delivery. Four hundred and thirty-two singleton children from these women with sample availability at both 6 and 18 months were tested for this study (Fig. 1).
Fig. 1.
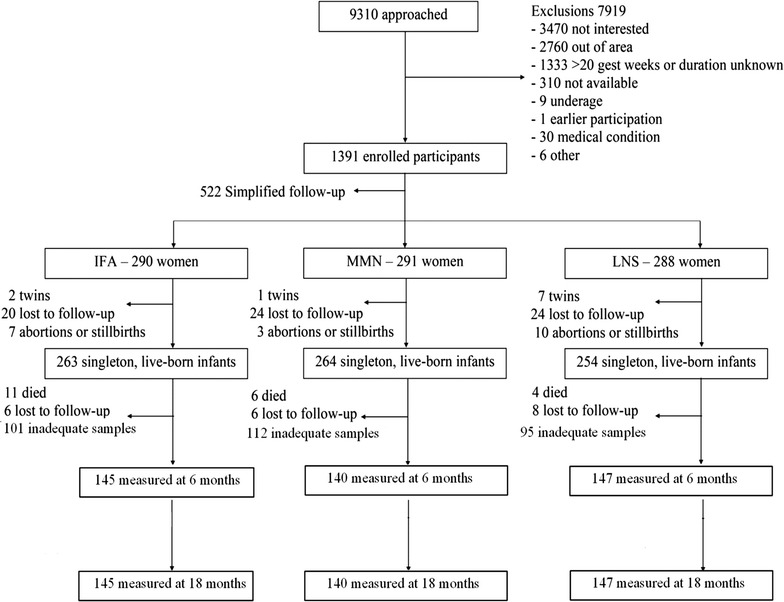
Participant flow in Consolidated Standards of Reporting Trials recommended format; adapted and modified from [22]. gest gestation, IFA iron and folic acid, LNS lipid-based nutrient supplement, MMN multiple micronutrients
Of the 432 children (47.9% male and 52.1% female) tested at 6 and 18 months of age, 33.6% were from the IFA group, 32.4% from the MMN group and 34.0% in the LNS group. Table 1 summarizes the participant characteristics according to different supplementation groups. The characteristics did not differ substantially between the three intervention groups. The percentages of children who were parasitaemic at 6 months were higher in children from the IFA group (8.3% by microscopy and 7.6% by RDT) compared to MMN (3.6% by microscopy and 7.1% by RDT) or LNS (3.4% by microscopy and 6.8% by RDT) but the differences were non-significant.
Table 1.
Participant characteristics according to supplementation groups
| Characteristics | IFAa n (%) |
MMNb n (%) |
LNSc n (%) |
P valued |
|---|---|---|---|---|
| Number of children | 145 (33.6) |
140 (32.4) |
147 (34.0) |
0.87 |
| Male | 69 (47.6) |
61 (43.6) |
77 (52.4) |
0.32 |
| Parasitaemia by microscopy at 6 months | 12 (8.3) |
5 (3.6) |
5 (3.4) |
0.10 |
| Parasitaemia by RDTe at 6 months | 11 (7.6) |
10 (7.1) |
10 (6.8) |
0.97 |
| Haemoglobin level at 6 months, mean ± SD, g/l | 102.7 ± 16.7 | 103.6 ± 16.0 | 103.7 ± 14.7 | 0.74 |
| Anaemiaf at 6 months | 86 (59.3) |
81 (57.9) |
84 (57.1) |
0.93 |
| Low socioeconomic status | 95 (65.5) |
75 (53.6) |
81 (55.1) |
0.08 |
| Mother’s education below median | 79 (54.5) |
66 (47.1) |
80 (54.4) |
0.34 |
aIron and folic acid supplementation
bMultiple micronutrient supplementation
cLipid-based nutrient supplements
dP value obtained by Kruskal–Wallis test (continuous variables) or Chi square test
eRapid diagnostic test
fAnaemia defined as haemoglobin level < 110 g/l (World Health Organization [1])
The characteristics of the included and excluded children at 6 months are illustrated in Table 2. Significantly more included children had parasitaemia by microscopy (P = 0.01), but results of RDT showed no such difference. The percentage of anaemia at 6 months was significantly higher (P < 0.0001) in the excluded participants (71.7%) than the included group (58.1%).
Table 2.
Comparative characteristics of the included and excluded participants at 6 months of age
| Characteristics | Included (432) | Excluded (349) | P valuea |
|---|---|---|---|
| Parasitaemia by microscopy | 22 (5.1) | 6 (1.7) | 0.01 |
| Parasitaemia by RDTb | 31 (7.17) | 35 (10.0) | 0.15 |
| Haemoglobin level, mean ± SD, g/l | 103.4 ± 15.8 | 101.4 ± 15.9 | 0.09 |
| Anaemiac | 251 (58.1) | 250 (71.7) | < 0.0001 |
| Low socioeconomic status | 251 (58.1) | 215 (61.6) |
0.32 |
| Mother’s education below median | 225 (52.1) | 193 (55.3) | 0.37 |
Values are number (%) or mean ± SD
aP value obtained by Chi square test or Mann–Whitney test (continuous variables)
bRapid diagnostic test
cAnaemia defined as haemoglobin level < 110 g/l (World Health Organization [1])
The magnitude and prevalence of antibodies in different age groups
The levels of antibodies and seroprevalence for the antibodies against merozoite antigens, schizont extract and VSA for three different parasite lines were measured at 6 months and 18 months of age (Table 3). The antibody levels to the merozoite antigens and schizont extract were significantly higher at 18 months compared to the levels at 6 months and it was statistically significant (P < 0.0001) for MSP1, MSP2 and EBA175.
Table 3.
Magnitude and prevalence of antibodies in 432 children at 6 and 18 months
| Antigen tested/Pfa isolate | Antibody levelb | P valued | Antibody seropositivityc (%) | P valuee | ||
|---|---|---|---|---|---|---|
| 6 months | 18 months | 6 months | 18 months | |||
| MSP1f | 0.97 [0.38, 2.89] | 2.80 [0.83, 6.35] | < 0.0001 | 128 (29.6) | 236 (54.6) | < 0.0001 |
| MSP2g | 2.14 [1.31, 3.59] | 2.95 [0.92, 9.12] | < 0.0001 | 114 (26.3) | 117 (27.0) | 0.1004 |
| EBA175h | 2.50 [1.44, 4.13] | 3.73 [2.19, 18.73] | < 0.0001 | 48 (11.1) | 51 (11.8) | 0.5764 |
| Rh2A9i | 5.49 [2.01, 33.78] | 8.5 [4.68, 14.29] | 0.2829 | 14 (3.2) | 122 (28.2) | < 0.0001 |
| Schizont | 1.92 [0.84, 6.99] | 3.32 [1.44, 6.89] | 0.2522 | 202 (46.7) | 233 (53.9) | 0.0314 |
| E8B | 0.20 [0.03, 0.45] | 0 [0, 0.21] | < 0.0001 | 105 (25.2) | 34 (7.8) | < 0.0001 |
| R29 | 0.18 [0, 0.63] | 0 [0, 0.11] | < 0.0001 | 37 (9.1) | 16 (3.7) | 0.0016 |
| 3D7 | 0.23 [0.01, 0.53] | 0 [0, 0.29] | < 0.0001 | 94 (22.2) | 36 (8.3) | < 0.0001 |
a Plasmodium falciparum
bAntibody level presented as a percentage of the positive control showing the median and interquartile range
cSeropositivity defined as sample mean optical density or fluorescence intensity > mean + 3 standard deviations of the negative controls
dP value calculated using Wilcoxon matched-pairs signed-ranks test
eP value calculated using McNemar’s test
fMerozoite surface protein 1
gMerozoite surface protein 2
hErythrocyte binding antigen 175
iReticulocyte binding protein homologue 2A
Significant P values <0.05 are indicated in italics
However, the levels of naturally acquired IgG against VSAs were significantly lower in children at 18 months of age compared to the same children at 6 months. This difference was significant (< 0.0001) for all the tested parasite lines.
As with the antibody level data, seroprevalence of antibodies against the tested merozoite antigens and schizont extract were also significantly higher at 18 months compared to the seroprevalence at 6 months. The highest percentage of seropositivity was observed against MSP1 in 18 month old children; 54.6% compared to 29.6% seropositives at 6 months (P < 0.0001). Significant increases in seropositivity with increasing age were also observed for Rh2A9 (P < 0.0001) and schizont extract (P = 0.0314). However, the seroprevalence of IgG against VSAs of different parasite lines declined significantly between 6 and 18 months of age for the three tested parasite lines (P < 0.0001 for IgG against E8B and 3D7 and P = 0.0016 for R29).
Association between nutrient supplementation and antibody seroprevalence in 6 months old children
Seroprevalence of the malaria antigens in 6 months old children was not significantly different between any of the treatment arms for any of the tested antigens except Rh2A9 (P = 0.044) (Table 4). Adjustment of the analysis for the selected covariates did not significantly change the results of the analysis, and the only significant difference was observed for the odds ratio of Rh2A9 between LNS and IFA group in the univariate analysis (P = 0.047), with the odds of being seropositive at 6 months of age in LNS group being 54% less than in the IFA group. This association did not remain significant in the adjusted analysis.
Table 4.
Association between nutrient supplementation and seroprevalence in 6 months old children
| Outcome | Number of children seropositive/total number of children | Comparison between LNS and IFA group | Comparison between LNS and MMN group | Comparison between MMN and IFA group | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| IFAa | MMNb | LNSc | P valued | OR (95% CI) | P valuee | OR (95% CI) | P valuee | OR (95% CI) | P valuee | |
| MSP-1f 19kD | 43/145 (29.7%) |
44/140 (31.4%) |
41/147 (27.9%) |
0.806 | 0.96 (0.74, 1.23) | 0.739 | 0.84 (0.51, 1.40) | 0.512 | 1.07 (0.64, 1.77) | 0.804 |
| Adjusted modelg | 0.97 (0.73, 1.28) | 0.808 | 0.86 (0.49, 1.49) | 0.583 | 1.01 (0.57, 1.76) | 0.981 | ||||
| MSP-2h | 38/145 (26.2%) |
40/140 (28.6%) |
36/147 (24.5%) |
0.734 | 0.96 (0.73, 1.24) | 0.736 | 0.81 (0.48, 1.37) | 0.434 | 1.07 (0.63, 1.80) | 0.807 |
| Adjusted modelg | 0.98 (0.74, 1.31) | 0.893 | 0.80 (0.45, 1.42) | 0.447 | 1.09 (0.62, 1.94) | 0.764 | ||||
| EBA-175i | 15/145 (10.3%) |
20/140 (14.3%) |
13/147 (8.9%) |
0.320 | 0.92 (0.62, 1.35) | 0.663 | 0.58 (0.28, 1.22) | 0.152 | 1.34 (0.65, 2.76) | 0.426 |
| Adjusted modelg | 0.96 (0.63, 1.47) | 0.857 | 0.54 (0.25, 1.18) | 0.120 | 1.53 (0.70, 3.37) | 0.289 | ||||
| Rh2A9j | 9/145 (6.21%) |
3/140 (2.14%) |
2/147 (1.4%) |
0.044 | 0.46 (0.21, 0.99) | 0.047 | 0.63 (0.10, 3.83) | 0.616 | 0.33 (0.09, 1.23) | 0.098 |
| Adjusted modelg | 0.48 (0.21, 1.06) | 0.070 | 0.79 (0.12, 5.42) | 0.814 | 0.34 (0.09, 1.30) | 0.114 | ||||
| Schizont extract | 68/145 (46.9%) |
67/140 (47.86%) |
67/147 (45.6%) |
0.927 | 0.97 (0.77, 1.23) | 0.821 | 0.91 (0.57, 1.45) | 0.699 | 1.07 (0.67, 1.71) | 0.774 |
| Adjusted modelg | 0.91 (0.70, 1.18) | 0.468 | 0.89 (0.53, 1.51) | 0.677 | 0.97 (0.58, 1.63) | 0.923 | ||||
| VSAk of E8B parasite line | 31/145 (21.4%) |
39/140 (27.9%) |
35/147 (23.8%) |
0.437 | 1.09 (0.83, 1.43) | 0.552 | 0.80 (0.47, 1.36) | 0.402 | 1.52 (0.88, 2.64) | 0.134 |
| Adjusted modelg | 1.01 (0.74, 1.37) | 0.957 | 0.70 (0.38, 1.26) | 0.233 | 1.55 (0.86, 2.80) | 0.146 | ||||
| VSA of R29 parasite line | 14/145 (9.7%) |
12/140 (8.6%) |
11/147 (7.5%) |
0.803 | 0.86 (0.57, 1.30) | 0.470 | 0.83 (0.35, 1.94) | 0.659 | 0.88 (0.39, 1.98) | 0.756 |
| Adjusted modelg | 0.76 (0.48, 1.20) | 0.234 | 0.69 (0.27, 1.74) | 0.427 | 0.91 (0.38, 2.14) | 0.822 | ||||
| VSA of 3D7 parasite line | 27/145 (18.6%) |
33/140 (23.6%) |
34/147 (23.1%) |
0.530 | 1.16 (0.87, 1.54) | 0.313 | 0.97 (0.56, 1.69) | 0.924 | 1.35 (0.76, 2.41) | 0.301 |
| Adjusted modelg | 1.04 (0.75, 1.43) | 0.829 | 0.89 (0.48, 1.65) | 0.721 | 1.24 (0.66, 2.31) | 0.501 | ||||
aIron and folic acid
bMultiple micronutrients
clipid based nutrient supplements
d P value calculated using the Chi2 test
e P value calculated using logistic regression reporting Odds Ratios (OR) and 95% Confidence intervals (CI)
fMerozoite surface protein 1
gP value calculated using multivariate logistic regression reporting odds ratios (OR) while adjusting for maternal BMI at enrolment, duration of gestation (from enrolment to delivery), number of pregnancies, sex of the child, maternal education, socioeconomic status, study site, maternal anaemic status at enrolment, maternal HIV status and bed net use by children
hMerozoite surface protein 2
iErythrocyte binding antigen 175
jReticulocyte binding protein homologue 2A
kVariant surface antigens
Significant P values <0.05 are indicated in italics
Association between nutrient supplementation and antibody seroprevalence in 18 months old children
Seroprevalence of the tested malaria antigens in 18 months old children was not significantly different between any of the treatment arms for any of the antigens tested (Table 5). Adjustment of the analysis for the selected covariates did not significantly change the results of the analysis, and no significant differences in antibody seroprevalence were observed between the different supplementation groups for any of tested antigens.
Table 5.
Association between nutrient supplementation and seroprevalence in 18 months old children
| Outcome | Number of children seropositive/total number of children | Comparison between LNS and IFA group | Comparison between LNS and MMN group | Comparison between MMN and IFA group | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| IFAa | MMNb | LNSc | P valued | OR (95% CI) | P valuee | OR (95% CI) | P valuee | OR (95% CI) | P valuee | |
| MSP-1 19kDf | 82/145 (56.5%) |
72/140 (51.4%) |
82/147 (55.7%) |
0.646 | 0.98 (0.78, 1.24) | 0.895 | 1.19 (0.75, 1.90) | 0.460 | 0.83 (0.52, 1.33) | 0.444 |
| Adjusted modelg | 0.98 (0.77, 1.25) | 0.865 | 1.10 (0.67, 1.82) | 0.708 | 0.80 (0.48, 1.34) | 0.398 | ||||
| MSP-2h | 42/145 (28.9%) |
39/140 (27.8%) |
36/147 (24.4%) |
0.669 | 0.88 (0.66, 1.16) | 0.359 | 0.79 (0.45, 1.40) | 0.428 | 0.95 (0.54, 1.66) | 0.851 |
| Adjusted modelg | 0.82 (0.60, 1.13) | 0.226 | 0.63 (0.34, 1.18) | 0.149 | 0.96 (0.53, 1.76) | 0.906 | ||||
| EBA-175i | 19/145 (13.1%) |
14/140 (10.0%) |
18/147 (12.2%) |
0.705 | 0.99 (0.67, 1.43) | 0.967 | 1.59 (0.69, 3.63) | 0.273 | 0.61 (0.27, 1.40) | 0.242 |
| Adjusted modelg | 0.91 (0.60, 1.37) | 0.642 | 1.19 (0.48, 2.95) | 0.710 | 0.65 (0.27, 1.58) | 0.342 | ||||
| Rh2A9j | 42/145 (28.9%) |
40/140 (28.5%) |
40/147 (27.2%) |
0.941 | 0.96 (0.74, 1.24) | 0.739 | 0.93 (0.56, 1.57) | 0.797 | 0.96 (0.57, 1.61) | 0.880 |
| Adjusted modelg | 0.96 (0.73, 1.26) | 0.763 | 0.84 (0.48, 1.47) | 0.540 | 0.98 (0.56, 1.71) | 0.931 | ||||
| Schizont extract | 73/145 (50.3%) |
78/140 (55.7%) |
82/147 (55.7%) |
0.568 | 1.12 (0.89, 1.40) | 0.352 | 1.003 (0.63, 1.60) | 0.991 | 1.24 (0.78, 1.99) | 0.365 |
| Adjusted modelg | 1.18 (0.90, 1.54) | 0.223 | 0.87 (0.52, 1.46) | 0.600 | 1.40 (0.83, 2.35) | 0.208 | ||||
| VSAk of E8B parasite line | 14/145 (9.6%) |
11/140 (7.9%) |
9/147 (6.1%) |
0.534 | 0.78 (0.51, 1.21) | 0.266 | 0.76 (0.30, 1.89) | 0.554 | 0.78 (0.33, 1.83) | 0.562 |
| Adjusted modelg | 0.80 (0.50, 1.29) | 0.360 | 0.72 (0.27, 1.89) | 0.504 | 0.75 (0.29, 1.93) | 0.557 | ||||
| VSA of R29 parasite line | 7/145 (4.8%) |
5/140 (3.5%) |
4/147 (2.7%) |
0.632 | 0.74 (0.40, 1.39) | 0.351 | 0.76 (0.20, 2.87) |
0.680 | 0.72 (0.22, 2.32) | 0.582 |
| Adjusted modelg | 0.79 (0.39, 1.60) | 0.506 | 0.95 (0.22, 4.07) | 0.946 | 0.85 (0.23, 3.14) | 0.808 | ||||
| VSA of 3D7 parasite line | 15/145 (10.3%) |
11/140 (7.8%) |
10/147 (6.8%) |
0.532 | 0.80 (0.52, 1.21) | 0.283 | 0.86 (0.35, 2.08) | 0.732 | 0.85 (0.37, 1.97) | 0.710 |
| Adjusted modelg | 0.74 (0.46, 1.22) | 0.241 | 0.55 (0.20, 1.52) | 0.250 | 1.11(0.43, 2.83) | 0.830 | ||||
aIron and folic acid
bMultiple micronutrients
clipid based nutrient supplements
d P-value calculated using the Chi2 test
e P-value calculated using logistic regression reporting Odds Ratios (OR) and 95% Confidence intervals (CI)
fMerozoite surface protein 1
gP-value calculated using multivariate logistic regression reporting odds ratios (OR) while adjusting for maternal BMI at enrolment, duration of gestation (from enrolment to delivery), number of pregnancies, sex of the child, maternal education, socioeconomic status, study site, maternal anaemic status at enrolment, maternal HIV status and bed net use by children
hMerozoite surface protein 2
iErythrocyte binding antigen 175
jReticulocyte binding protein homologue 2A
kVariant surface antigens
Association between nutrient supplementation and antibody levels in 6 month old children
The level of antibodies did not differ significantly according to different nutrient supplementation groups for any of the tested antigens at 6 months of age. Moreover, multivariate linear regression showed no significant differences in the levels of antibodies against any of the tested antigens when they were categorized by different supplementation groups (Table 6).
Table 6.
Association between nutrient supplementation and antibody levels in 6 months old children
| Outcome | Antibody levels by study group, median (IQR) | LNS and IFA | LNS and MMN | MMN and IFA | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| IFAa | MMNb | LNSc | P valued | Coeff (95% CI) | P valuee | Coeff (95% CI) | P valuee | Coeff (95% CI) | P valuee | |
| Number of participants | N = 145 | N = 140 | N = 147 | |||||||
| MSP-1 19kDf | 0.81 (0.38, 2.45) | 1.13 (0.37, 3.02) | 1.03 (0.40, 0.72) | 0.430 | 1.03 (0.85, 1.25) | 0.773 | 0.88 (0.60, 1.28) | 0.498 | 1.15 (0.78, 1.70) | 0.467 |
| Adjusted modelg | 1.06 (0.87, 1.30) | 0.542 | 0.94 (0.63, 1.39) | 0.755 | 1.11 (0.75, 1.64) | 0.614 | ||||
| MSP-2h | 2.36 (1.43, 3.57) | 2.02 (1.33, 4.13) | 1.98 (1.19, 3.27) | 0.313 | 0.99 (0.87, 1.12) | 0.833 | 0.87 (0.66, 1.15) | 0.334 | 1.08 (0.82, 1.42) | 0.592 |
| Adjusted modelg | 0.99 (0.86, 1.13) | 0.834 | 0.88 (0.66, 1.19) | 0.417 | 1.06 (0.80, 1.40) | 0.703 | ||||
| EBA-175i | 2.37 (1.34, 3.85) | 2.60 (1.50, 4.50) | 2.50 (1.42, 4.02) | 0.335 | 1.03 (0.92, 1.14) | 0.610 | 0.91 (0.74, 1.13) | 0.390 | 1.14 (0.91, 1.43) | 0.240 |
| Adjusted modelg | 1.04 (0.93, 1.16) | 0.502 | 0.86 (0.69, 1.08) | 0.195 | 1.24 (0.99, 1.54) | 0.061 | ||||
| Rh2A9j | 4.93 (1.53,18.55) | 5.97 (2.12, 39.95) | 5.86 (2.45, 33.70) | 0.245 | 1.11 (0.89, 1.38) | 0.364 | 1.00 (0.66, 1.53) | 0.986 | 1.23 (0.77, 1.95) | 0.382 |
| Adjusted modelg | 1.08 (0.85, 1.37) | 0.532 | 0.99 (0.63, 1.57) | 0.990 | 1.16 (0.70, 1.93) | 0.559 | ||||
| Schizont extract | 1.85 (0.80, 4.93) | 2.11 (0.82, 8.08) | 2.06 (0.89, 7.23) | 0.569 | 1.07 (0.89, 1.28) | 0.465 | 0.97 (0.68, 1.40) | 0.875 | 1.21 (0.85, 1.73) | 0.282 |
| Adjusted modelg | 1.06 (0.89, 1.27) | 0.481 | 0.88 (0.61, 1.26) | 0.475 | 1.30 (0.91, 1.85) | 0.155 | ||||
| VSAk of E8B parasite line | 0.17 (0.03, 0.46) | 0.26 (0.06, 0.47) | 0.19 (0.02, 0.44) | 0.453 | 1.04 (0.87, 1.24) | 0.688 | 1.00 (0.70, 1.42) | 0.995 | 1.07 (0.76, 1.51) | 0.709 |
| Adjusted modelg | 1.00 (0.82, 1.21) | 0.996 | 0.99 (0.68, 1.47) | 0.987 | 1.02 (0.71, 1.47) | 0.925 | ||||
| VSA of R29 parasite line | 0.16 (0, 0.59) | 0.25 (0, 0.81) | 0.12 (0, 0.55) | 0.452 | 0.93 (0.75, 1.15) | 0.496 | 0.70 (0.46, 1.07) | 0.098 | 1.21 (0.80, 1.82) | 0.358 |
| Adjusted modelg | 0.91 (0.73, 1.13) | 0.407 | 0.63 (0.41, 0.97) | 0.063 | 1.30 (0.88, 1.91) | 0.187 | ||||
| VSA of 3D7 parasite line | 0.19 (0.01, 0.48) | 0.24 (0.02, 0.64) | 0.26 (0.01,0.59) | 0.544 | 1.19 (0.98, 1.46) | 0.081 | 1.06 (0.72, 1.55) | 0.774 | 1.34 (0.91, 1.99) | 0.138 |
| Adjustedg | 1.13 (0.92, 1.39) | 0.245 | 1.05 (0.69, 1.61) | 0.810 | 1.18 (0.81, 1.73) | 0.382 | ||||
aIron and folic acid
bMultiple micronutrients
clipid based nutrient supplements
d P value calculated using Kruskal–Wallis test
e P value calculated using linear regression of antibody levels between supplementation groups reporting coefficient and 95% Confidence intervals (CI)
fMerozoite surface protein 1
gP value calculated using multivariate regression adjusting for maternal BMI at enrolment, duration of gestation (from enrolment to delivery), number of pregnancies, sex of the child, maternal education, socioeconomic status, study site, maternal anaemic status at enrolment, maternal HIV status and bed net use by children
hMerozoite surface protein 2
iErythrocyte binding antigen 175
jReticulocyte binding protein homologue 2A
kVariant surface antigens
Association between nutrient supplementation and antibody levels in 18 month old children
The level of antibodies did not differ significantly between different nutrient supplementation groups for any of the tested antigens except for IgG against E8B parasite line (P = 0.043) in 18 months old children. Multivariate linear regression showed no significant differences in the levels of antibodies against any of the tested antigens between the different supplementation groups (Table 7).
Table 7.
Association between nutrient supplementation and antibody levels in 18 months old children
| Outcome | Antibody levels by study group, median (IQR) | LNS and IFA | LNS and MMN | MMN and IFA | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| IFAa | MMNb | LNSc | P valued | Coeff (95% CI) | P valuee | Coeff (95% CI) | P valuee | Coeff (95% CI) | P valuee | |
| Number of participants | N = 145 | N = 140 | N = 147 | |||||||
| MSP-1 19kDf | 2.59 (0.83, 6.09) | 2.95 (0.75, 6.37) | 2.64 (1.00, 6.59) | 0.877 | 1.05 (0.87, 1.26) | 0.603 | 1.02 (0.70, 1.49) | 0.909 | 1.09 (0.75, 1.58) | 0.663 |
| Adjusted modelg | 1.04 (0.86, 1.26) | 0.681 | 0.97 (0.65, 1.42) | 0.857 | 1.07 (0.72, 1.58) | 0.745 | ||||
| MSP-2h | 3.10 (0.97, 9.81) | 2.74 (0.64, 8.57) | 3.21 (1.38, 9.05) | 0.334 | 1.01 (0.83, 1.22) | 0.938 | 1.12 (0.77, 1.64) | 0.548 | 0.92 (0.61, 1.38) | 0.683 |
| Adjusted modelg | 0.99 (0.82, 1.22) | 0.984 | 1.17 (0.79, 1.74) | 0.434 | 0.86 (0.56, 1.31) | 0.477 | ||||
| EBA-175i | 4.06 (2.32, 19.29) | 3.39 (1.88, 20.11) | 3.62 (2.29, 18.47) | 0.425 | 0.94 (0.80, 1.10) | 0.427 | 0.99 (0.07, 1.38) | 0.973 | 0.89 (0.64, 1.24) | 0.489 |
| Adjusted modelg | 0.90 (0.77, 1.06) | 0.217 | 0.98 (0.70, 1.36) | 0.894 | 0.84 (0.60, 1.17) | 0.294 | ||||
| Rh2A9j | 8.96 (4.66,13.50) | 8.38 (4.77, 14.46) | 8.76 (4.86,15.17) | 0.952 | 1.02 (0.92, 1.12) | 0.723 | 1.02 (0.85, 1.22) | 0.848 | 1.02 (0.83, 1.24) | 0.869 |
| Adjusted modelg | 1.03 (0.93, 1.14) | 0.566 | 1.03 (0.84, 1.25) | 0.790 | 1.05 (0.85, 1.29) | 0.679 | ||||
| Schizont extract | 3.13 (1.69, 6.19) | 3.94 (1.40, 8.18) | 2.96 (1.28, 6.35) | 0.407 | 0.94 (0.80, 1.10) | 0.433 | 0.79 (0.56, 1.12) | 0.185 | 1.10 (0.80, 1.49) | 0.564 |
| Adjusted modelg | 0.91 (0.78, 1.09) | 0.332 | 0.78 (0.54, 1.12) | 0.179 | 1.10 (0.80, 1.51) | 0.559 | ||||
| VSAk of E8B parasite line | 0.00 (0.00, 0.27) | 0.00 (0.00, 0.25) | 0.00 (0.00, 0.12) | 0.043 | 0.82 (0.60, 1.11) | 0.189 | 0.62 (0.35, 1.10) | 0.099 | 0.99 (0.60, 1.61) | 0.953 |
| Adjusted modelg | 0.83 (0.61, 1.14) | 0.250 | 0.61 (0.34, 1.11) | 0.102 | 0.95 (0.60, 1.51) | 0.818 | ||||
| VSA of R29 parasite line | 0.00 (0.00, 0.07) | 0.00 (0.00, 0.24) | 0.00 (0.00, 0.004) | 0.223 | 0.86 (0.58, 1.29) | 0.469 | 0.89 (0.39, 1.98) | 0.756 | 0.85 (0.39, 1.84) | 0.671 |
| Adjusted modelg | 0.82 (0.53, 1.28) | 0.382 | 0.73 (0.32, 1.70) | 0.465 | 0.95 (0.43, 2.06) | 0.889 | ||||
| VSA of 3D7 parasite line | 0.002 (0.00, 0.39) | 0.00 (0.00, 0.24) | 0.00 (0.00, 0.28) | 0.150 | 1.05 (0.78, 1.43) | 0.731 | 1.18 (0.64, 2.19) | 0.592 | 0.98 (0.56, 1.71) | 0.930 |
| Adjustedg | 1.14 (0.82, 1.59) | 0.441 | 1.11 (0.55, 2.25) | 0.765 | 0.94 (0.50, 1.75) | 0.834 | ||||
aIron and folic acid
bMultiple micronutrients
clipid based nutrient supplements
d P value calculated using Kruskal–Wallis test
e P value calculated using linear regression of antibody levels between supplementation groups reporting coefficient and 95% Confidence intervals (CI)
fMerozoite surface protein 1
gP value calculated using multivariate regression adjusting for maternal BMI at enrolment, duration of gestation (from enrolment to delivery), number of pregnancies, sex of the child, maternal education, socioeconomic status, study site, maternal anaemic status at enrolment, maternal HIV status and bed net use by children
hMerozoite surface protein 2
iErythrocyte binding antigen 175
jReticulocyte binding protein homologue 2A
kVariant surface antigens
Significant P values <0.05 are indicated in italics
Discussion
This study investigated the impact of nutritional supplementation on malarial immunity in a subset of young children from a randomized controlled trial of pre-natal nutrient supplementation with IFA or pre- and postnatal MMN or LNS. Antibodies to several important merozoite antigens (MSP1, MSP2, EBA175, Rh2A9), schizont extract and VSA for three different parasite lines (E8B-ICAM, R29 and 3D7 overexpressing var A) were measured in order to determine whether nutrient supplementation improves the acquisition of malaria antibody in young children.
In malaria endemic areas, antibodies to malaria antigens increase with age, being higher in adults than in children, but the dynamics of antibody production in infants are less well studied [36]. The observations that the levels and seroprevalence of antibody to most of the merozoite antigens tested increased between 6 and 18 months of age are in agreement with observations in a case control study conducted in Kenyan children, in which the level of antibodies against MSP1 and PfRh2 increased steadily with increasing age from birth to 2 years [37], and a cohort study from Benin [38], in which MSP1 and MSP2 antibody levels showed a constant increase until 18 months of age. In a higher transmission setting, the seroprevalence of antibody to MSP1 reached a peak after 6 months of age, whereas prevalence of antibody to MSP2 peaked at 9 months of age [39].
In contrast to the increase in antibodies to merozoite antigens, the levels of antibodies against VSA decreased from 6 to 18 months, suggesting an ongoing loss of maternal anti-VSA antibodies with little or no development of the child’s own VSA antibody responses. Consistent with this, in a recent study anti-VSA antibodies waned by 6–9 months of age and did not reappear during infancy and early childhood [40]. It is possible that the anti-VSA antibodies require more infections than the merozoite antigens to develop, and anti-VSA responses are known to often be short-lived following infection [41]. This may be related to the highly variant nature of VSA [42], requiring repeated exposure to develop cross-reactive antibodies to multiple VSA variants, whereas merozoite antigens are more conserved.
The study found no evidence that nutrient supplementation altered the development of antibody responses to either merozoite antigens or VSA in young children, in accordance with a study conducted on Beninese children of similar age group in whom there was no association between nutritional intake and anti-malarial antibody levels [38]. Antibody levels or seroprevalence of antibodies to the tested antigens did not differ between the supplementation groups in either unadjusted or adjusted analysis (P > 0.05) except for the seroprevalence in 6 months old children for Rh2A9 (P = 0.044), and that significance was lost in the adjusted analysis. The trial did not demonstrate differences in growth or morbidity events between the different supplementation arms and suggested that the nutritional supplements provided were not sufficient by themselves to promote infant growth [22]. This may also explain why no differences were seen in anti-malaria antibody levels between the intervention groups.
Clinical studies have suggested that under-nutrition has an impact on early childhood mortality and morbidity [2, 43]. However, other studies did not find any relationship between protein energy malnutrition or stunting and clinical malaria occurrence [7, 44]. The interaction between nutrition and malarial immunity has been relatively little studied. Among Senegalese pre-school children, those children who were stunted had lower levels of IgG to schizont extract compared to non-stunted children [45], and Papua New Guinean children with wasting were reported to have lower levels of specific IgG to blood stage malaria antigens [4]. But these previous studies of the impact of nutritional status on malarial immunity were observational and therefore subjected to confounding. The only statistically significant difference between the supplement groups seen in the present study in 6 months old children was the seroprevalence against Rh2A9 between three treatment arms (P = 0.044); the univariate analysis also showed significant differences in the odds ratio between LNS and IFA group. There was no significant association in the adjusted analysis. Although IgG against VSA in E8B parasite line in 18 months old children was significantly different in unadjusted analyses between groups (P = 0.043), this, together with the Rh2A9 observations, were most probably chance findings, in the absence of similar findings in other assays.
Strengths of the study include the substantial sample size of children from a randomized controlled trial with paired samples at 6 and 18 months of age and the number of different antibody measurements performed. A high proportion of iLiNS study participants were stunted at 18 months of age, indicating a high prevalence of nutritional deprivation, frequent infection or inflammation in infants. Possible study weaknesses include the relatively low prevalence of severe nutritional deficits in the mothers [21] as a previous study showed that with protein energy supplementation, improved birth outcome was more prominently observed in malnourished mothers than marginally nourished or well-nourished mothers [46]. Other limitations could be the inclusion of only a subset of all children followed in the parent study [21], the potential selection bias due to differences in the included and excluded children, different assay platforms for measuring merozoite immunity (ELISA) compared to VSA immunity (flow cytometry), and the lack of functional assays of malaria immunity [36].
Conclusions
In conclusion, this study reports the acquisition of malaria antibody in young children with different pre- and postnatal nutrient supplements and shows no significant changes in antibody acquisition among different supplement groups. It addresses the knowledge gap regarding the effect of nutrient supplementation on malarial immunity in young children. Further studies may be required to determine whether malaria immunity is significantly impaired in groups with severe nutritional depletion, or whether more substantial protein energy supplementation might help the development of malaria immunity, especially in high risk groups.
Authors’ contributions
SJR and PA designed the laboratory studies. KGD, PA and KM designed the intervention trial. PB and UPC performed laboratory work. PB, SJR, PA and JGB analysed the data. PB and SJR drafted the manuscript. All authors read and approved the final manuscript.
Acknowledgements
We acknowledge all the study participants, the local communities, the health services staff and our research personnel at the study sites as well as members of the trial’s data safety and monitoring board, and the International Lipid-based Nutrient Supplementation (iLiNS)-Project Steering Committee (http://www.ilins.org/) for their positive attitude, support and help in all stages of the study. In addition, we would like to thank Jack Richards and Wina Hasang for advice and technical support. We also acknowledge the independent statistical consultancy of Sabine Braat from the Melbourne School of Population and Global Health and Melbourne Clinical and Translational Sciences Platform, University of Melbourne.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The datasets supporting the conclusions of this article are included within the article and its additional files.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Informed consent was obtained from the participants of the study. Ethical approval for the trial and the laboratory work was granted by the College of Medicine Research and Ethics Committee of Malawi, and by Tampere University Hospital Ethics Committee, Finland. The Melbourne Health Human Research Ethics Committee approved the laboratory work performed for the study.
Funding
This publication is based on research funded in part by a grant from the Office of Health, Infectious Diseases, and Nutrition, Bureau for Global Health, U.S. Agency for International Development (USAID) under terms of Cooperative Agreement No. AID-OAA-A-12-00005, through the Food and Nutrition Technical Assistance III Project (FANTA), managed by FHI 360. Additional funding was obtained from a grant to the University of California, Davis from the Bill & Melinda Gates Foundation and the Academy of Finland. SJR and JGB are supported by a Program Grant from the National Health and Medical Research Council of Australia (NHMRC) and JGB is supported by an NHMRC Senior Research Fellowship. The Burnet Institute is supported by the NHMRC IRIIS Scheme and Victorian State Government Operational Infrastructure Support.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- EBA
erythrocyte binding antigen
- HHA
household assets
- IE
infected erythrocytes
- IFA
iron and folic acid
- iLiNS
International Lipid-based Nutrient Supplement
- IPTp
intermittent preventive treatment in pregnancy
- IQR
interquartile range
- ITN
insecticide-treated net
- LNS
lipid-rich nutrient supplement
- MACS
magnetic-activated cell sorting
- MMN
multiple micronutrients
- MSP
merozoite surface protein
- OR
odds ratio
- PCR
polymerase chain reaction
- PfEMP1
P. falciparum erythrocyte membrane protein-1
- PfRh2
Plasmodium falciparum reticulocyte binding homologue 2
- Rh2A9
reticulocyte binding protein homologue 2A
- SES
socioeconomic status
- VSA
variant surface antigen
References
- 1.WHO . World malaria report, 2015. Geneva: World Health Organization; 2015. [Google Scholar]
- 2.Friedman JF, Kwena AM, Mrel LB, Kariuki SK, Terlouw DJ, Phillips-Howard PA, et al. Malaria and nutritional status among pre-school children: results from cross-sectional surveys in western Kenya. Am J Trop Med Hyg. 2005;73:698–704. [PubMed] [Google Scholar]
- 3.Deen JL, Walraven GE, Seidlein L. Increased risk for malaria in chronically malnourished children; under 5 years of age in rural Gambia. J Trop Pediatr. 2002;48:78–83. doi: 10.1093/tropej/48.2.78. [DOI] [PubMed] [Google Scholar]
- 4.Genton B, Al-Yaman F, Ginny M, Taraika J, Alpers MP. Relation of anthropometry to malaria morbidity and immunity in Papua New Guinean children. Am J Clin Nutr. 1998;68:734–741. doi: 10.1093/ajcn/68.3.734. [DOI] [PubMed] [Google Scholar]
- 5.Snow RW, Byass P, Shenton FC, Greenwood BM. The relationship between anthropometric measurements and measurements of iron status and susceptibility to malaria in Gambian children. Trans R Soc Trop Med Hyg. 1991;85:584–589. doi: 10.1016/0035-9203(91)90351-X. [DOI] [PubMed] [Google Scholar]
- 6.Fillol F, Cournil A, Boulanger D, Cisse B, Sokhna C, Targett G, et al. Influence of wasting and stunting at the onset of the rainy season on subsequent malaria morbidity among rural preschool children in Senegal. Am J Trop Med Hyg. 2009;80:202–208. [PubMed] [Google Scholar]
- 7.Deribew A, Alemseged F, Tessema F, Sena L, Birhanu Z, Zeynudin A, et al. Malaria and under-nutrition: a community based study among under-five children at risk of malaria South-West Ethiopia. Plos One. 2010;5:e10775. doi: 10.1371/journal.pone.0010775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Owusu-Agyei S, Newton S, Mahama E, Febir LG, Ali M, Adjei K, et al. Impact of vitamin A with zinc supplementation on malaria morbidity in Ghana. Nutr J. 2013;12:131. doi: 10.1186/1475-2891-12-131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Shankar AH, Genton B, Baisor M, Paino J, Tamja S, Adiguma T, et al. The influence of zinc supplementation on morbidity due to Plasmodium falciparum: a randomized trial in preschool children in Papua New Guinea. Am J Trop Med Hyg. 2000;62:663–669. doi: 10.4269/ajtmh.2000.62.663. [DOI] [PubMed] [Google Scholar]
- 10.Shankar AH, Genton B, Semba RD, Baisor M, Paino J, Tamja S, et al. Effect of vitamin A supplementation on morbidity due to Plasmodium falciparum in young children in Papua New Guinea: a randomised trial. Lancet. 1999;354:203–209. doi: 10.1016/S0140-6736(98)08293-2. [DOI] [PubMed] [Google Scholar]
- 11.Prasetyani MA, de Mast Q, Afeworki R, Kaisar MMM, Stefanie D, Sartono E, et al. Effect of a short course of iron polymaltose on acquisition of malarial parasitaemia in anaemic Indonesian schoolchildren: a randomized trial. Malar J. 2017;16:50. doi: 10.1186/s12936-017-1691-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sazawal S, Black RE, Ramsan M, Chwaya HM, Stoltzfus RJ, Dutta A, et al. Effects of routine prophylactic supplementation with iron and folic acid on admission to hospital and mortality in preschool children in a high malaria transmission setting: community-based, randomised, placebo-controlled trial. Lancet. 2006;367:133–143. doi: 10.1016/S0140-6736(06)67962-2. [DOI] [PubMed] [Google Scholar]
- 13.Zlotkin S, Newton S, Aimone AM, Azindow I, Amenga-Etego S, Tchum K, et al. Effect of iron fortification on malaria incidence in infants and young children in Ghana: a randomized trial. JAMA. 2013;310:938–947. doi: 10.1001/jama.2013.277129. [DOI] [PubMed] [Google Scholar]
- 14.Krishna S, Taylor AM, Supanaranond W, Pukrittayakamee S, ter Kuile F, Tawfiq KM, et al. Thiamine deficiency and malaria in adults from southeast Asia. Lancet. 1999;353:546–549. doi: 10.1016/S0140-6736(98)06316-8. [DOI] [PubMed] [Google Scholar]
- 15.Metzger A, Mukasa G, Shankar AH, Ndeezi G, Melikian G, Semba RD. Antioxidant status and acute malaria in children in Kampala Uganda. Am J Trop Med Hyg. 2001;65:115–119. doi: 10.4269/ajtmh.2001.65.115. [DOI] [PubMed] [Google Scholar]
- 16.Osendarp SJM, Prabhakar H, Fuchs GJ, van Raaij JMA, Mahmud H, Tofail F, et al. Immunization with the heptavalent pneumococcal conjugate vaccine in Bangladeshi infants and effects of zinc supplementation. Vaccine. 2007;25:3347–3354. doi: 10.1016/j.vaccine.2007.01.001. [DOI] [PubMed] [Google Scholar]
- 17.Siddiqua TJ, Ahmad SM, Ahsan KB, Rashid M, Roy A, Rahman SM, et al. Vitamin B12 supplementation during pregnancy and postpartum improves B12 status of both mothers and infants but vaccine response in mothers only: a randomized clinical trial in Bangladesh. Eur J Nutr. 2016;55:281–293. doi: 10.1007/s00394-015-0845-x. [DOI] [PubMed] [Google Scholar]
- 18.Teo A, Feng G, Brown GV, Beeson JG, Rogerson SJ. Functional antibodies and protection against blood-stage malaria. Trends Parasitol. 2016;32:887–898. doi: 10.1016/j.pt.2016.07.003. [DOI] [PubMed] [Google Scholar]
- 19.Chan JA, Fowkes FJI, Beeson JG. Surface antigens of Plasmodium falciparum-infected erythrocytes as immune targets and malaria vaccine candidates. Cell Mol Life Sci. 2014;71:3633–3657. doi: 10.1007/s00018-014-1614-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.iLiNS project. [www.iLiNS.org]. Accessed 4 Feb 2018.
- 21.Ashorn P, Alho L, Ashorn U, Cheung YB, Dewey KG, Harjunmaa U, et al. The impact of lipid-based nutrient supplement provision to pregnant women on newborn size in rural Malawi: a randomized controlled trial. Am J Clin Nutr. 2015;101:387–397. doi: 10.3945/ajcn.114.088617. [DOI] [PubMed] [Google Scholar]
- 22.Ashorn P, Alho L, Ashorn U, Cheung YB, Dewey KG, Gondwe A, et al. Supplementation of maternal diets during pregnancy and for 6 months postpartum and infant diets thereafter with small-quantity lipid-based nutrient supplements does not promote child growth by 18 months of age in rural Malawi: a Randomized Controlled Trial. J Nutr. 2015;145:1345–1353. doi: 10.3945/jn.114.207225. [DOI] [PubMed] [Google Scholar]
- 23.Biggs BA, Anders RF, Dillon HE, Davern KM, Martin M, Petersen C, et al. Adherence of infected erythrocytes to venular endothelium selects for antigenic variants of Plasmodium falciparum. J Immunol. 1992;149:2047–2054. [PubMed] [Google Scholar]
- 24.Rowe JA, Moulds JM, Newbold CI, Miller LH. P. falciparum rosetting mediated by a parasite-variant erythrocyte membrane protein and complement-receptor 1. Nature. 1997;388:292–295. doi: 10.1038/40888. [DOI] [PubMed] [Google Scholar]
- 25.Chan JA, Stanisic DI, Duffy MF, Robinson LJ, Lin E, Kazura JW, et al. Patterns of protective associations differ for antibodies to P. falciparum-infected erythrocytes and merozoites in immunity against malaria in children. Eur J Immunol. 2017;47(12):2124–2136. doi: 10.1002/eji.201747032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Chandrasiri UP, Randall LM, Saad AA, Bashir AM, Rogerson SJ, Adam I. Low antibody levels to pregnancy-specific malaria antigens and heightened cytokine responses associated with severe malaria in pregnancy. J Infect Dis. 2014;209:1408–1417. doi: 10.1093/infdis/jit646. [DOI] [PubMed] [Google Scholar]
- 27.Goodyer ID, Johnson J, Eisenthal R, Hayes DJ. Purification of mature-stage Plasmodium falciparum by gelatine flotation. Ann Trop Med Parasitol. 1994;88:209–211. doi: 10.1080/00034983.1994.11812859. [DOI] [PubMed] [Google Scholar]
- 28.Stanisic DI, Richards JS, McCallum FJ, Michon P, King CL, Schoepflin S, et al. Immunoglobulin G subclass-specific responses against Plasmodium falciparum merozoite antigens are associated with control of parasitemia and protection from symptomatic illness. Infect Immun. 2009;77:1165–1174. doi: 10.1128/IAI.01129-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Richards JS, Stanisic DI, Fowkes FJI, Tavul L, Dabod E, Thompson JK, et al. Association between naturally acquired antibodies to erythrocyte-binding antigens of Plasmodium falciparum and protection from malaria and high-density parasitemia. Clin Infect Dis. 2010;51:E50–E60. doi: 10.1086/656413. [DOI] [PubMed] [Google Scholar]
- 30.Reiling L, Richards JS, Fowkes FJI, Barry AE, Triglia T, Chokejindachai W, et al. Evidence that the erythrocyte invasion ligand PfRh2 is a target of protective immunity against Plasmodium falciparum malaria. J Immunol. 2010;185:6157–6167. doi: 10.4049/jimmunol.1001555. [DOI] [PubMed] [Google Scholar]
- 31.Persson KEM, McCallum FJ, Reiling L, Lister NA, Stubbs J, Cowman AF, et al. Variation in use of erythrocyte invasion pathways by Plasmodium falciparum mediates evasion of human inhibitory antibodies. J Clin Invest. 2008;118:342–351. doi: 10.1172/JCI32138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Teo A, Hasang W, Rogerson S. Evaluating IgG antibody to variant surface antigens expressed on Plasmodium falciparum infected erythrocytes using flow cytometry. Methods Mol Biol. 2015;1325:207–213. doi: 10.1007/978-1-4939-2815-6_17. [DOI] [PubMed] [Google Scholar]
- 33.Chan JA, Howell KB, Reiling L, Ataide R, Mackintosh CL, Fowkes FJ, et al. Targets of antibodies against Plasmodium falciparum-infected erythrocytes in malaria immunity. J Clin Invest. 2012;122(9):3227–3238. doi: 10.1172/JCI62182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.iLiNS-DYAD-M analysis plan version 27.0 with appendices. [http://www.ilins.org/ilins-project-research/data-analysis/ilins-dyad-m-analysis-plan-version-27.0-with-appendices-2017-02-12/view]. Accessed on 12 Feb 2017.
- 35.Filmer D, Pritchett LH. Estimating wealth effects without expenditure data—Or tears: an application to educational enrollments in states of India. Demography. 2001;38:115–132. doi: 10.1353/dem.2001.0003. [DOI] [PubMed] [Google Scholar]
- 36.Dobbs KR, Dent AE. Plasmodium malaria and antimalarial antibodies in the first year of life. Parasitology. 2016;143:129–138. doi: 10.1017/S0031182015001626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Murungi LM, Sonden K, Llewellyn D, Rono J, Guleid F, Williams AR, et al. Targets and mechanisms associated with protection from severe Plasmodium falciparum malaria in Kenyan children. Infect Immun. 2016;84:950–963. doi: 10.1128/IAI.01120-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Dechavanne C, Sadissou I, Bouraima A, Ahouangninou C, Amoussa R, Milet J, et al. Acquisition of natural humoral immunity to P. falciparum in early life in Benin: impact of clinical, environmental and host factors. Sci Rep. 2016;6:33961. doi: 10.1038/srep33961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kangoye DT, Mensah VA, Murungi LM, Nkumama I, Nebie I, Marsh K, et al. Dynamics and role of antibodies to Plasmodium falciparum merozoite antigens in children living in two settings with differing malaria transmission intensity. Vaccine. 2016;34:160–166. doi: 10.1016/j.vaccine.2015.10.058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Dent AE, Malhotra I, Wang X, Babineau D, Yeo KT, Anderson T, et al. Contrasting patterns of serologic and functional antibody dynamics to Plasmodium falciparum antigens in a Kenyan birth cohort. Clin Vaccine Immunol. 2015;23:104–116. doi: 10.1128/CVI.00452-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kinyanjui SM, Bull P, Newbold CI, Marsh K. Kinetics of antibody responses to Plasmodium falciparum-infected erythrocyte variant surface antigens. J Infect Dis. 2003;187:667–674. doi: 10.1086/373994. [DOI] [PubMed] [Google Scholar]
- 42.Marsh K, Howard RJ. Antigens induced on erythrocytes by P. falciparum: expression of diverse and conserved determinants. Science. 1986;231:150–153. doi: 10.1126/science.2417315. [DOI] [PubMed] [Google Scholar]
- 43.Caulfield LE, Richard SA, Black RE. Undernutrition as an underlying cause of malaria morbidity and mortality in children less than 5 years old. Am J Trop Med Hyg. 2004;71:55–63. [PubMed] [Google Scholar]
- 44.Muller O, Garenne M, Kouyate B, Becher H. The association between protein-energy malnutrition, malaria morbidity and all-cause mortality in West African children. Trop Med Int Health. 2003;8:507–511. doi: 10.1046/j.1365-3156.2003.01043.x. [DOI] [PubMed] [Google Scholar]
- 45.Fillol F, Sarr JB, Boulanger D, Cisse B, Sokhna C, Riveau G, et al. Impact of child malnutrition on the specific anti-Plasmodium falciparum antibody response. Malar J. 2009;8:116. doi: 10.1186/1475-2875-8-116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Winkvist A, Habicht JP, Rasmussen KM. Linking maternal and infant benefits of a nutritional supplement during pregnancy and lactation. Am J Clin Nutr. 1998;68:656–661. doi: 10.1093/ajcn/68.3.656. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets supporting the conclusions of this article are included within the article and its additional files.