

Abstract
Understanding the factors that facilitate the emergence of cooperation among organisms is central to the study of social evolution. Spotted hyenas Crocuta crocuta frequently cooperate to mob lions Panthera leo, approaching the lions as a tightknit group while vocalizing loudly in an attempt to overwhelm them and drive them away. Whereas cooperative mobbing behavior has been well documented in birds and some mammals, to our knowledge it has never been described during interactions between 2 apex predators. Using a 27-year dataset, we characterize lion–hyena encounters, assess rates of mobbing behavior observed during these interactions, and inquire whether mobbing results in successful acquisition of food. Lions and hyenas interacted most often at fresh kills, especially as prey size and the number of hyenas present increased. Possession of food at the beginning of an interaction positively affected retention of that food by each predator species. The presence of male lions increased the probability of an interspecific interaction but decreased the likelihood of hyenas obtaining or retaining possession of the food. Hyena mobbing rates were highest at fresh kills, but lower when adult male lions were present. The occurrence of mobbing was predicted by an increase in the number of hyenas present. Whether or not mobbing resulted in acquisition of food from lions was predicted by an increase in the number of mobs formed by the hyenas present, suggesting that cooperation among hyenas enhances their fitness.
Keywords: competition, cooperation, hyena, lion, mobbing, sociality.
A central focus in the study of social evolution is the emergence of cooperation among organisms, including the factors that facilitate or impede cooperation. We define cooperation as collective action among individuals for mutual benefit (Dugatkin et al. 1992a). In many animal species, cooperation involves coordinated action by multiple individuals (Stephens and Anderson 1996). Cooperative actions must, on average, increase the fitness of the individuals involved, even though the direct or indirect benefits accruing to any specific individual may not be obvious (Noë 2006; Brosnan and Bshary 2010). Cooperative behaviors function importantly in animal societies to allow groups of individuals to accomplish objectives that would not be achievable by any individual acting alone (Hammerstein 2003).
After cooperative breeding (e.g., Moehlman and Hofer 1997), most research on cooperation among mammalian carnivores has focused on group hunting, which occurs in many gregarious species (Bailey et al. 2013). However, mobbing is another important form of cooperative behavior, which occurs when 2 or more individuals in a high state of arousal synchronously approach or attack a threatening stimulus in the environment. Mobbing behavior is commonly observed in many groups of animals, including birds (Altmann 1956; Sandoval and Wilson 2012), ground squirrels (Owings and Coss 1977), primates (Gursky-Doyen and Nekaris 2007), and cetaceans (De Stephanis et al. 2015). In these species, mobbing functions to deter or harass potential predators. Among mammalian carnivores, mobbing behavior has been observed in gregarious mongooses (Rood 1975) coatis (Nasua narica; Janzen 1970), meerkats (Suricata suricatta; Graw and Manser 2007), and spotted hyenas (Crocuta crocuta; Kruuk 1972; Mills 1990). Mobbing by spotted hyenas is observed most frequently during their interactions with lions (Panthera leo; Kruuk 1972).
Lions and spotted hyenas are the dominant large carnivores in most African ecosystems, and these 2 species are one another’s main competitors for resources (Périquet et al. 2015). When interacting with lions, hyenas frequently mob them by making synchronous coalitionary attacks on the lions. Hyenas live in fission–fusion societies (Smith et al. 2008) and often recruit group-mates to locations where they encounter lions by emitting long-distance vocalizations (Gersick et al. 2015). Once multiple hyenas are present, individuals may cooperate to mob the lions, approaching them as a cohesive group and vocalizing loudly together, a behavior that can enable the hyenas to overwhelm the lions and drive them away (Kruuk 1972; Mills 1990). However, there are considerable fitness costs associated with this form of cooperation. Lions are significantly larger and stronger than hyenas, and they represent the leading cause of mortality in many hyena populations (Cooper 1991; Hofer and East 1995; Trinkel and Kastberger 2005). An attack from a lion can result in serious injury or death for a mobbing hyena (Kruuk 1972).
Here, we focus on interspecific interactions between lions and spotted hyenas, and the intraspecific cooperative mobbing behavior that occurs among hyenas during many of these encounters. Because lion–hyena interactions are complex and highly variable (Kruuk 1972; Mills 1990), their analysis requires a large sample size, which can only be obtained from detailed long-term observational data. We utilize a dataset spanning 27 years to characterize the lion–hyena interactions taking place within the territories of 7 hyena clans at 2 study sites in Kenya. Our first goal is simply to describe lion–hyena encounters, asking where they occur and under what circumstances, and when they result in interspecific encounters. We next focus on mobbing behavior, inquiring about specific variables recorded during lion–hyena encounters that predict whether or not this cooperative behavior will occur. Finally, we test a hypothesis suggesting that mobbing behavior increases the probability that hyenas will obtain food from lion-controlled kills or carcasses, thus likely enhancing the fitness of the hyenas participating in those mobs.
Materials and Methods
Study animals
Spotted hyenas are gregarious, long-lived predators that live in social groups called clans. Clans in east Africa are comprised of multiple matrilines of adult females and their offspring, and several adult immigrant males (Kruuk 1972; Frank 1986). Clans can contain up to 130 individuals (Holekamp et al. 2015), and all female clan-mates concurrently nursing young cubs rear them together at a communal den (Kruuk 1972). Each clan is structured by a linear dominance hierarchy that determines the priority of access to resources by individual group members (Frank 1986). Spotted hyenas cooperate in coalitionary aggression against clan-mates, a behavior that serves to enforce rank-relationships and defend resources within their societies (Engh et al. 2000; Smith et al. 2010). Although mean relatedness among clan-mates is very low (Van Horn et al. 2004), clan-mates also frequently join forces to defend a common territory against conspecifics, to guard their kills, and to secure resources from sympatric carnivores (Cooper 1991; Henschel and Skinner 1991). Here all hyenas were identified individually by their unique spot patterns and sexed based on the morphology of the erect phallus (Frank et al. 1990).
Study populations
Between 1988 and 2014, we monitored 7 clans of free-living spotted hyenas in 2 different national parks in Kenya, 5 in the Masai Mara National Reserve and 2 in Amboseli National Park (Supplementary Table S1). Both of these national parks are areas of open tropical grassland that support large herds of resident and seasonally migrant herbivores, which serve as prey for the resident carnivores (Kolowski et al. 2007; Watts and Holekamp 2008). Prey availability was recorded during biweekly surveys by counting all herbivores within 100 m of 2–4 line transects of 1–5.4 km in each territory (Holekamp et al. 1999; Watts and Holekamp 2008; Green 2015; Supplementary Table S1). We calculated the prey density within the territories of each of our study clans on a monthly basis, and used the monthly number of standard deviations above or below the yearly mean to determine prey availability for each clan during each month of study.
Observation sessions
All methods of data collection were identical among populations. We monitored clans daily during 2 observation periods, in the morning from 6 to 10 AM and in the evening from 4 to 8 PM. When we encountered a subgroup of one or more hyenas, we initiated an observation session, and all hyenas within 200 m of that subgroup were considered to be present in that session. Observation sessions (hereafter, “sessions”) lasted from 5 min to several hours and ended when interactions ceased and observers left that individual or group. Session length was recorded as the duration of each session in minutes. “Den” sessions occurred within 200 m of an active hyena den, “kill” sessions occurred within 200 m of a fresh kill (made less than 24 h previously), and “carcass” sessions occurred within 200 m of a kill older than 24 h (Boydston et al. 2003). Carcass age was determined by observers based on its odor, appearance, and the presence or absence of fresh blood. Locations of all remaining sessions were categorized as “other” sessions, which usually involved animals resting or travelling far from a den, kill, or carcass.
We defined a lion–hyena “encounter” as occurring whenever we found members of the 2 species within 200 m of one another. We identified an interspecific “interaction” as occurring whenever hyenas and lions approached within 10 m of one another. At each encounter, we recorded the total number and identities of all hyenas present, as well as the total number, age class, and sex of all lions present (Whitman and Packer 2006). All encounters occurred within or on the boundaries of the territory of a study clan.
Feeding behavior
Many lion–hyena interactions involve contests over food resources. Here “food” included both fresh kills and older carcasses. In each kill or carcass session, we recorded all observed feeding behavior by any lions or hyenas using one-zero sampling (Altmann 1974). A predator species was considered to be feeding if at least 1 member of that species obtained at least 4 kg of food, which represents the spotted hyena’s typical daily food intake (Green et al. 1984; Henschel and Skinner 1990). Food mass consumed was estimated from archived field notes in which our observations of lion–hyena encounters were recorded. These notes contained detailed descriptions of the kill or carcass, how this changed over the course of the session, which individuals fed, and which specific body parts were consumed. Whenever possible, we also recorded which prey species had been killed. Kill size was categorized by prey species weight (Sachs 1967) as small (<100 kg; e.g., gazelles, impala), medium (100–300 kg; e.g., zebra, wildebeest, topi), or large (>300 kg; e.g., eland, buffalo, giraffe).
Interspecific competition between lions and hyenas
Both the prevention of kleptoparasitism of food by lions and the usurpation of food resources from lions are achieved primarily through synchronous mobbing behavior by hyenas (Trinkel and Kastberger 2005). Throughout each session involving both lions and hyenas, we recorded all mobbing events using all-occurrence sampling (Altmann 1974). We defined “mobbing” as a group of 2 or more hyenas, usually side-by-side and within 1 m of one another, with tails bristled over their backs, approaching within 10 m of at least 1 lion. In association with each mobbing event, we recorded the identities of all participating hyenas and the age/sex classes of the lions being approached. We counted all mobs that occurred during each session and calculated an hourly mobbing rate for each session as the total number of observed mobbing events divided by the total number of minutes in that session, multiplied by 60.
At sessions where kills or carcasses were present, we recorded which species controlled the food throughout the session, including which predator species was in control when observers arrived on the scene and which controlled the food, if any remained, when observers left. When observers arrived after a kill had been made, we determined which predator species had made the kill based on 2 criteria. A predator species was assigned credit for killing the prey animal if members of that species were considerably bloodier than members of the competing species when the session began (Cooper 1991), or if no members of the competing predator species were present when observers arrived on the scene of a very fresh kill (Watts and Holekamp 2008). If both predators were present when observers arrived, but neither species was obviously bloodier than the other, the predator species making the kill was assigned as unknown.
Statistical analyses
Four trained research assistants (S.M.M., J.M.P., O.S.S., and K.J.V) extracted data on lion–hyena encounters from detailed written field notes describing these interactions. To ensure consistency, all work was overseen by T.M.M. and K.D.S.L. 13% of all sessions were randomly reviewed by T.M.M. and K.D.S.L., and this subset did not differ from the remaining sessions with respect to hyena count, lion count, male lion count, or number of mobs (Wilcoxon–Mann–Whitney P > 0.05). Mean inter-observer reliability was calculated by determining the research assistants’ agreement with a “correct dataset” generated by T.M.M. and K.D.S.L. on 12 complex lion–hyena interaction sessions. The agreement score between the 4 individuals extracting data from field notes averaged 83% for all 7 variables of interest: hyena count, lion count, male lion count, number of mobs, who controlled the food at the start and end of the session, and whether hyenas fed.
All datasets and their criteria are described in Table 1, as well as below. We used nonparametric statistical tests for between-group comparisons, as all datasets failed to meet the assumption of normality for parametric tests.
Table 1.
The criteria for inclusion in datasets analyzed in Results
| Dataset | Criteria | Number of Sessions | Complete Cases Modeled |
|---|---|---|---|
| Lion–Hyena Sessions | Sessions in which a known number of lions and hyenas occurred within 200 m of each other (“encounters”) | 935 | 903 |
| Food Sessions | Lion–Hyena Sessions in which a kill or carcass was present with known food possession | 394 | – |
| Known Killer Sessions | Food Sessions in which the species that made the kill was known | 221 | 216 |
| Potential Mobbing Sessions | Lion–Hyena Sessions in which more than 1 hyena was present | 761 | 736 |
| Potential Feeding Sessions | Potential Mobbing Sessions in which the session began with lions in control of the food | 235 | 227 |
| Talek Kill Sessions | Talek clan sessions in which a fresh, identifiable kill was present | 2,558 | 2,239 |
All datasets include both “encounters” (within 200 m) and “interactions” (within 10 m) between lions and hyenas. All sample sizes represent the number of observation sessions meeting inclusion criteria. Nonparametric statistical tests and other calculations were conducted using the relevant full “number of sessions” dataset. Any sessions for which prey availability could not be calculated were excluded from modeling analyses, which used the reduced “complete cases modeled” dataset.
Generalized linear mixed models (GLMMs) were built to include all biologically relevant predictors as well as our predictors of interest. Sessions for which prey availability could not be calculated were excluded from modeling analyses; thus, modeling was conducted on datasets comprised exclusively of “complete cases.” We included year nested within clan as random effects in all models using data from more than 1 clan, and we included year as a random effect in all models using data from a single clan. This allowed us to determine whether our predictors of interest were significant while controlling for variation in territory size, territory quality, and predator and prey density. Session length in minutes was included in all models to control for variation in observation time. Prey availability was included in all models to control for likely variation in hunger levels and motivation in both predator species due to seasonal fluctuations in prey abundance. All numeric model predictors were z-score standardized using the scale function in R to simplify comparing coefficients. Coefficients were estimated via maximum likelihood and Laplace approximation. All model predictors were tested for multicollinearity that would justify their elimination from the model and none were found to be collinear. All model residuals were visually inspected to confirm assumptions of homoscedasticity and comparable variance between groups. All groups and observations were inspected for disproportionate influence on the models and none warranted exclusion. All responses were modeled via logistic regression and the logit link function.
All analyses were conducted using RStudio Version 0.99.489 and R Version 3.2.3 utilizing the lme4 package for all GLMMs, the car package to test for multicollinearity in model predictors, and the popbio package to create Figures 3 and 4.
Figure 3.
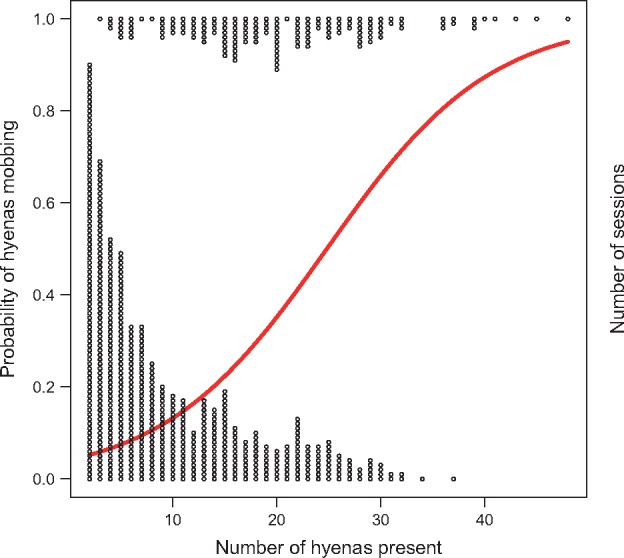
Probability of hyenas mobbing. The probability of mobbing function (red) plotted against the number of hyenas present in each session using complete cases in dataset “Potential Mobbing Sessions.” The open circles represent the number of sessions in which mobbing does (top) or does not (bottom) occur, with sessions binned by the number of hyenas present. As the number of hyenas present increased, the log odds of mobbing also increased (Table 5).
Figure 4.
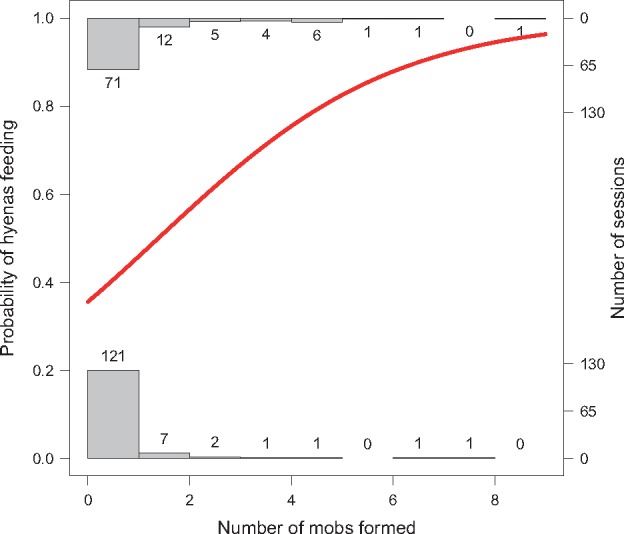
Probability of hyenas feeding. The probability of hyenas feeding function (red) plotted against the number of mobs formed during each session using complete cases in dataset “Potential Feeding Sessions.” Histogram bars and sample sizes indicate the number of sessions in which mobbing occurred when hyenas fed (top) or did not feed (bottom). As the number of mobs increased, the log odds of hyenas feeding also increased (Table 6).
Characterizing lion–hyena interactions
We created dataset “Lion–Hyena Sessions” using all sessions where known numbers of lions and hyenas occurred within 200 m of each other (n = 935). Using the subset of “Lion–Hyena Sessions” where the 2 species approached within 10 m of one another (n = 410), we calculated the median and mean numbers of hyenas and lions present at sessions where the 2 predators actually interacted. We used “Lion–Hyena Sessions” with known prey availability to build a GLMM of the log odds of lions and hyenas interacting (n = 903 complete cases), with presence or absence of an interspecies interaction as the binary response variable and session location, hyena count, lion count, presence of male lions, the interaction between session location and session length, the interaction between lion count and hyena count, and the interaction between hyena count and presence of male lions as fixed effects.
To determine how often hyenas encountered lions at a food source, we used all fresh kill sessions with an identifiable prey species in the Talek clan territory (n = 2,558; hereafter, “Talek Kill Sessions”) to calculate the percentage of kills at which lions were present. We restricted this analysis to the Talek clan because killed prey species' identities were reliably recorded there. We used “Talek Kill Sessions” with known prey availability to build a GLMM to inquire whether lions were present more or less often at kills of prey of different sizes (n = 2,239 complete cases). This GLMM modeled lion presence or absence as the binary response variable, with prey size and the interaction of prey size and session length as fixed effects.
We created dataset “Food Sessions” using “Lion–Hyena Sessions” with a kill or carcass present in which control of the carcass was unambiguous at both the beginning and end of the session (n = 394). These sessions were used to calculate the percentage of fresh kills made by hyenas versus lions, as well as the number and percentage of sessions in which each species started and ended with food. We used “Food Sessions” where lions had possession of the food at the start of the session (n = 255) to calculate the mean hyena:lion ratio in sessions where hyenas either won or lost contests over kills and carcasses against groups of lions with and without adult male lions present.
Using the subset of “Food Sessions” where the predator species making the kill was known (n = 221; hereafter “Known Killer Sessions”), we built a GLMM to determine whether the predator species that killed the prey animal, or controlled its carcass at the beginning of the session, affected that species’ retention of the food until the end of the session (n = 216 complete cases). We modeled control of the food at the end of the session as the binary response variable (hyenas = 1, lions = 0), and included the following variables as fixed effects: which species started with food, which species killed the prey animal, hyena count, lion count, presence of male lions, number of mobs formed, the interaction between session length and which species started with food, the interaction between hyena count and lion count, the interaction between hyena count and presence of male lions, and the interaction between session length and number of mobs formed.
Variables predicting whether or not mobbing occurs
We used “Lion–Hyena Sessions” with multiple hyenas present (n = 761; hereafter “Potential Mobbing Sessions”) to calculate the percentage of all lion–hyena encounters in which hyenas mobbed the lions. Using the subset of “Potential Mobbing Sessions” containing mobs (n = 157), we calculated the total number of mobs observed, the median and mean number of hyenas per mob, and the median and mean number of mobs per session.
Using “Potential Mobbing Sessions,” we compared mobbing rates between session locations using Kruskal–Wallis and Wilcoxon–Mann–Whitney tests with Bonferroni corrections. We also counted the number of sessions in which hyenas mobbed lions when no discernable resource was present. Using the subset of “Food Sessions” where multiple hyenas were present (n = 353), we calculated the percentage of encounters involving mobbing when male lions were either present or absent. We also used Wilcoxon–Mann–Whitney tests on this subset to compare mobbing rates between kills with and without male lions present. Using “Talek Kill Sessions” at which both lions and multiple hyenas were present (n = 209) and a Kruskal–Wallis test, we compared mobbing rates among kills of known sizes.
We built a GLMM using “Potential Mobbing Sessions” to identify variables that increased the probability of hyenas forming mobs against lions (n = 736 complete cases). Here our response variable was binary, whether or not at least 1 mobbing event occurred during the session. Potential predictors included as fixed effects were session location, hyena count, lion count, presence of male lions, the interaction between session length and session location, the interaction between hyena count and lion count, and the interaction between hyena count and presence of male lions.
Does mobbing affect the probability that hyenas will feed?
Using a subset of “Potential Mobbing Sessions” in which the session began with lions in control of the food (n = 235; hereafter “Potential Feeding Sessions”), we built a GLMM to identify variables that increased the probability of hyenas feeding (n = 227 complete cases). This allowed us to test the hypothesis that cooperation, in the form of mobbing, increased the probability of hyenas obtaining food. Here our response variable was binary, whether or not any hyena obtained at least 4 kg of food during the session. Potential predictors included as fixed effects were hyena count, lion count, presence of male lions, number of mobs formed, the interaction between hyena count and lion count, the interaction between hyena count and presence of male lions, and the interaction between session length and the number of mobs formed.
Results
In our 7 study clans, lions and hyenas were observed within 200 m of one another in 1,038 different observation sessions. Of these, observers could accurately determine the number of lions and hyenas present in 935 sessions that lasted on average 37 min (median 25, range 5–240). Data from these “Lion–Hyena Sessions” were used to characterize lion–hyena encounters and interactions, and the mobbing behavior occurring therein.
Characterizing lion–hyena encounters
Lions and hyenas interacted in 43.9% (n = 410) of the 935 interspecific encounters. In these 410 interaction sessions, the median number of hyenas present was 14 (mean 14.9, range 1–48), and the median number of lions present was 3 (mean 3.6, range 1–20). In sessions where lions and hyenas co-occurred without interacting, the 2 species were either resting or traveling in proximity to one another, but the only relevant behavior we observed was watchfulness.
In our model of the likelihood of lions and hyenas interacting using complete cases (n = 903) from “Lion–Hyena Sessions,” the 2 species were more likely to interact at kill sessions than at any other session type (P = 0.02; Table 2). There were also 2 significant interactions between session location and session length (P < 0.02; Table 2; Supplementary Figure S1). In shorter sessions, den and kill locations were more likely to have interactions than carcass or “other” locations. A larger number of hyenas (P < 0.0001) and the presence of male lions (P < 0.04) increased the log odds of lions and hyenas interacting, although the number of lions present did not (P > 0.7; Table 2; Figure 1). In addition, a significant interaction between hyena count and lion count indicated that, as the number of hyenas present increased, so did the positive effect of the number of lions present on the probability that the 2 species would interact (P < 0.02; Table 2).
Table 2.
GLMM of the log odds of lions and hyenas interacting using complete cases in dataset “Lion–Hyena Sessions”
| Modeling log odds of lions and hyenas interacting (n = 903) | B | SE | z | P |
|---|---|---|---|---|
| Full Model | ||||
| Intercept | −0.46 | 0.18 | −2.58 | 0.0100 |
| Session length | 1.14 | 0.24 | 4.83 | <0.0001 |
| Prey availability | 0.16 | 0.09 | 1.72 | 0.0847 |
| Location – carcass | 0.03 | 0.35 | 0.09 | 0.9263 |
| Location – den | −0.07 | 0.33 | −0.20 | 0.8437 |
| Location – kill | 0.48 | 0.21 | 2.33 | 0.0200 |
| Hyena count | 0.76 | 0.14 | 5.45 | <0.0001 |
| Lion count | −0.03 | 0.09 | −0.31 | 0.7543 |
| Male lions present | 0.42 | 0.20 | 2.11 | 0.0346 |
| Session length × Location – carcass | −0.10 | 0.52 | −0.20 | 0.8424 |
| Session length × Location – den | −1.14 | 0.38 | −3.01 | 0.0026 |
| Session length × Location – kill | −0.63 | 0.26 | −2.38 | 0.0173 |
| Hyena count × Lion count | 0.32 | 0.13 | 2.53 | 0.0115 |
| Hyena count × Male lions present | 0.49 | 0.25 | 1.95 | 0.0507 |
The response is binary: lions and hyenas interact (1) or not (0). Year is nested within clan, with both included as random effects. All numeric predictors are standardized. SE: standard error. Bolded rows indicate P-value<0.05.
Figure 1.
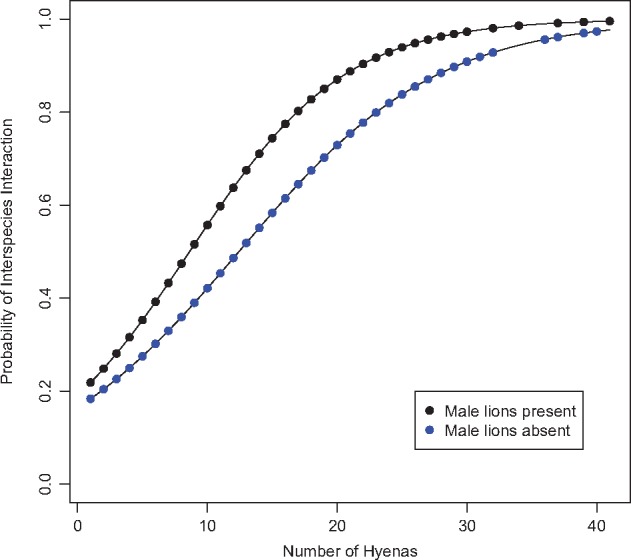
The probability of lions and hyenas interacting based on number of hyenas present and the presence or absence of adult male lions using complete cases in dataset “Lion–Hyena Sessions.” Each line plots the modeled log odds that lions and hyenas would interact in sessions in which they were within 200 m of one another. Data points are true hyena counts plotted on the curves predicted by the model. Hyenas and lions were more likely to interact when male lions and more hyenas were present (Table 2).
Lion–hyena encounters occurred at 10.3% (n = 263) of all 2,558 “Talek Kill Sessions.” While controlling for session length, the size of the prey animal had a significant effect on the log odds of a lion–hyena encounter at a kill (Table 3; Supplementary Figure S2). Small prey reduced the probability of lions being present (P < 0.0001), whereas large prey increased the probability of lion presence (P < 0.04; Table 3).
Table 3.
GLMM of the log odds of lions encountering hyenas at a kill session using complete cases in dataset “Talek Kill Sessions”
| Modeling log odds of lion presence with Talek hyenas at a kill (n = 2,239) | B | SE | z | P |
|---|---|---|---|---|
| Full Model | ||||
| Intercept | −2.02 | 0.11 | −18.62 | < 0.0001 |
| Session length | 0.62 | 0.09 | 6.70 | <0 .0001 |
| Prey availability | −0.11 | 0.07 | −1.60 | 0.1092 |
| Prey size – small | −0.71 | 0.17 | −4.17 | < 0.0001 |
| Prey size – large | 0.69 | 0.33 | 2.07 | 0.0387 |
| Session length × Prey size - small | −0.03 | 0.19 | −0.18 | 0.8604 |
| Session length × Prey size - large | −0.06 | 0.27 | −0.23 | 0.8175 |
The response is binary: lions are present (1) or not (0). Year is included as a random effect. All numeric predictors are standardized. Bolded rows indicate P-value<0.05.
In analyzing “Food Sessions” (n = 394), we found that hyenas made the majority of kills to which a predator species could be assigned (Supplementary Table S2); this was unsurprising given that hyenas are our primary study animals. In our model of food possession at lion–hyena sessions using complete cases from “Known Killer Sessions” (n = 216), the species in possession of the food when the session began positively affected retention of the food by both hyenas and lions regardless of session length (P < 0.003; Table 4; Supplementary Figure S3). However, which species actually made the kill in the first place had no effect on retention of the food by either species (P > 0.1; Table 4). Neither hyena count (P > 0.7) nor lion count (P > 0.1) affected the log odds of hyenas controlling food at the end of a session (Table 4), although hyena:lion ratios were higher at sessions in which hyenas successfully usurped food than when lions retained control of it throughout (n = 255, W = 4,525, P < 0.0001; Supplementary Table S3). Unsurprisingly, the presence of male lions significantly decreased the log odds of hyenas possessing the food at the end of a session (P = 0.0002; Table 4; Figure 2), and the hyena: lion ratio required to usurp the food was higher when male lions were present than absent, although this trend was not significant (n = 98, W = 1,042.5, P > 0.3; Supplementary Table S3). Somewhat surprisingly, the number of mobs did not significantly increase the log odds of hyenas controlling the food at the end of the session (P > 0.1).
Table 4.
GLMM of the log odds of lions or hyenas controlling the carcass or kill at the end of a session using complete cases in dataset “known killer sessions”
| Modeling log odds of lions (0) vs. hyenas (1) possessing the food at the end of a session (n = 216) | B | SE | z | P |
|---|---|---|---|---|
| Full Model | ||||
| Intercept | 1.06 | 0.43 | 3.76 | 0.0002 |
| Session length | 0.25 | 0.36 | 0.70 | 0.4834 |
| Prey availability | −0.04 | 0.18 | −0.20 | 0.8403 |
| Lions start with food | −1.22 | 0.40 | −3.08 | 0.0021 |
| Lions killed food | −0.81 | 0.52 | −1.57 | 0.1173 |
| Hyena count | 0.08 | 0.28 | 0.27 | 0.7851 |
| Lion count | −0.32 | 0.23 | −1.35 | 0.1763 |
| Male lions present | −1.57 | 0.42 | −3.71 | 0.0002 |
| Number of mobs | 0.40 | 0.26 | 1.54 | 0.1240 |
| Session length × Lions start with food | 0.21 | 0.40 | 0.53 | 0.5986 |
| Hyena count × Lion count | 0.26 | 0.25 | 1.07 | 0.2827 |
| Hyena count × Male lions present | −0.43 | 0.42 | −1.01 | 0.3118 |
| Session length × Number of mobs | −0.10 | 0.14 | −0.70 | 0.4860 |
The response variable is binary: lions (0) or hyenas (1) end the session in possession of the carcass. Year is nested within clan, with both included as random effects. All numeric predictors are standardized. Bolded rows indicate P-value<0.05.
Figure 2.
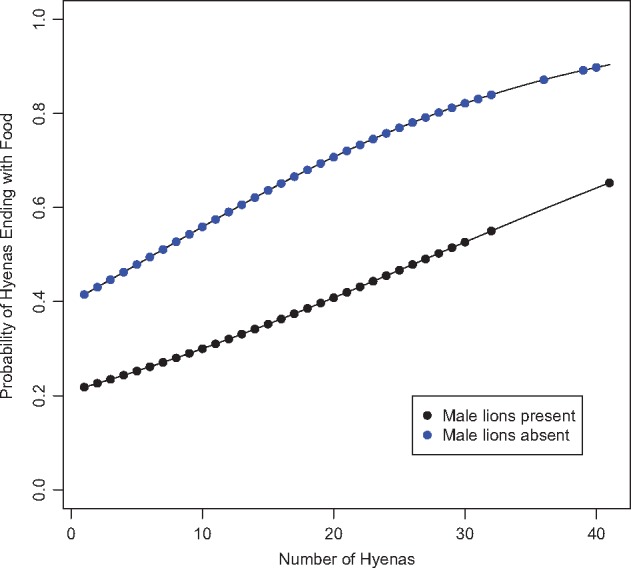
The probability of hyenas possessing the carcass at the end of the session based on number of hyenas present and the presence or absence of adult male lions using complete cases in dataset “Known Killer Sessions.” Each line plots the modeled log odds that hyenas would have possession of the carcass at the end of the session. Data points represent true hyena counts plotted on curves predicted by the model. Hyenas were more likely to end the session with food when male lions were absent and more hyenas were present (Table 4).
Variables predicting whether or not mobbing will occur
Hyenas formed 1 to 9 mobs (median 2, mean 2.5) against lions in 20.6% (n = 157) of 761 “Potential Mobbing Sessions” for a total of 394 mobs observed. The median size of these hyena mobs was 4 individuals (mean 6.0, range 2–28). Both adult male and female hyenas participated in mobbing, as did both high and low-ranking individuals. Mobbing hyenas often approached within 1 or 2 m of the lions, which put them within the lions’ reach and thus at considerable risk.
In analyzing “Potential Mobbing Sessions,” we found that hyena mobbing rates were highest at fresh kills, followed by carcass, den and then “other” locations, but only kill sessions and “other” sessions differed significantly from one another (Bonferroni corrected critical value for multiple pairwise comparisons = 0.0083; U = 40,567, P < 0.0001). Nonetheless, hyenas did mob lions in 29 “other” sessions where neither food nor den-dependent cubs were present. In sessions with multiple hyenas, mobbing occurred in 20.7% (n = 25) of 121 “Food Sessions” with male lions present, and in 31.9% (n = 74) of 232 “Food Sessions” at which male lions were absent. In these same sessions, mobbing rates were significantly higher when male lions were absent than when they were present (“Food Sessions” with multiple hyenas: n = 353, W = 12,312, P < 0.02). At kills, mobbing rates did not differ based on variation in prey size (“Talek Kill Sessions” with lions and multiple hyenas: n = 209, U = 1.846, df = 2, P > 0.3).
Our model of mobbing probability using complete cases from “Potential Mobbing Sessions” (n = 736) determined that hyenas were more likely to mob when larger numbers of hyenas were present (P < 0.0001; Table 5; Figure 3). Location (P > 0.3), lion count (P > 0.2), and presence of male lions (P > 0.1) were not significant predictors of mobbing (Table 5).
Table 5.
GLMM of the log odds of hyenas mobbing lions using complete cases in dataset “potential mobbing sessions”
| Modeling log odds of hyenas mobbing lions (n = 736) | B | SE | z | P |
|---|---|---|---|---|
| Full Model | ||||
| Intercept | −1.80 | 0.25 | −7.27 | <0.0001 |
| Session length | 0.66 | 0.25 | 2.58 | 0.0098 |
| Prey availability | 0.19 | 0.11 | 1.78 | 0.0756 |
| Location – carcass | −0.25 | 0.54 | −0.47 | 0.6353 |
| Location – den | 0.30 | 0.45 | 0.66 | 0.5070 |
| Location – kill | 0.28 | 0.28 | 1.02 | 0.3096 |
| Hyena count | 0.92 | 0.15 | 6.14 | <0.0001 |
| Lion count | −0.18 | 0.14 | −1.27 | 0.2056 |
| Male lions present | −0.45 | 0.30 | −1.52 | 0.1293 |
| Session length × Location – carcass | 0.83 | 0.62 | 1.34 | 0.1807 |
| Session length × Location – den | −0.83 | 0.47 | −1.77 | 0.0775 |
| Session length × Location – kill | −0.43 | 0.27 | −1.60 | 0.1097 |
| Hyena count × Lion count | 0.11 | 0.12 | 0.90 | 0.3660 |
| Hyena count × Male lions present | 0.27 | 0.26 | 1.05 | 0.2940 |
The response variable is binary: hyenas mob lions (1) or not (0). Year is nested within clan, with both included as random effects. All numeric predictors are standardized. Bolded rows indicate P-value<0.05.
Does mobbing affect the probability that hyenas will feed?
Our model of the probability of hyenas feeding using complete cases from “Potential Feeding Sessions” (n = 227) found that hyenas were more likely to feed when the number of mobs in the session was higher (P = 0.0003). A significant interaction between session length and number of mobs indicated that the shorter the session, the more influential the number of mobs were on feeding occurrence (P < 0.002; Table 6; Figure 4). Hyena count (P > 0.6), lion count (P > 0.1), and presence of male lions (P > 0.1) were not significant predictors of feeding (Table 6). These data support the hypothesis that mobbing behavior increases the likelihood that hyenas will obtain food from lion-controlled kills or carcasses.
Table 6.
GLMM of the log odds of hyenas feeding using complete cases in dataset “potential feeding sessions”
| Modeling log odds of hyenas feeding (n = 227) | B | SE | z | P |
|---|---|---|---|---|
| Full Model | ||||
| Intercept | −0.33 | 0.24 | −1.36 | 0.1741 |
| Session length | 0.58 | 0.21 | 2.78 | 0.0055 |
| Prey availability | 0.17 | 0.17 | 1.03 | 0.3014 |
| Hyena count | −0.11 | 0.23 | −0.47 | 0.6369 |
| Lion count | −0.29 | 0.19 | −1.55 | 0.1212 |
| Male lions present | 0.58 | 0.37 | 1.58 | 0.1140 |
| Number of mobs | 1.15 | 0.32 | 3.63 | 0.0003 |
| Hyena count × Lion count | 0.27 | 0.18 | 1.56 | 0.1191 |
| Hyena count × Male lions present | 0.51 | 0.39 | 1.30 | 0.1946 |
| Session length × Number of mobs | −0.50 | 0.16 | −3.23 | 0.0012 |
The response variable is binary: hyenas feed (1) or not (0). Year is nested within clan, with both included as random effects. All numeric predictors are standardized. Bolded rows indicate P-value<0.05.
Discussion
Interactions between spotted hyenas and lions offer a particularly interesting case study compared with other species considered in the literature on risk-taking behavior in shifting cost/benefit landscapes. Although hyenas are top predators themselves, they are frequently killed or wounded by lions (Périquet et al. 2015). Hyenas and lions have a high degree of dietary overlap and often compete directly for the same food resources (Kruuk and Turner 1967; Hayward 2006). Lions are larger and more powerful than hyenas, which puts the hyenas at risk of injury or death during competitive interactions over food (Kruuk 1972; Schaller 1972; Hofer and East 1995). However, the possible benefits of acquiring food may outweigh the risk of injury from lions (Watts and Holekamp 2008). Here we document some important situational variables, such as the relative numbers of lions and hyenas present, the size of the contested carcass, the presence of male lions, and whether or not hyenas form mobs against the lions, that affect the probability that the hyenas will benefit from their risky interactions with lions. Our data enhance the understanding of the role of cooperation in mediating interspecific competition between these 2 competitors.
Characterizing lion–hyena interactions
Where the 2 species co-occurred, lions and hyenas were most likely to interact over fresh kills (Table 2), which is unsurprising for 2 predators whose diets overlap by more than 68% (Périquet et al. 2015). Fresh kills are extremely rich but highly ephemeral resources in the Mara ecosystem (Jones et al. 2015) and are thus sites of intense interspecific feeding competition. Lions and hyenas also had a high probability of interacting at the communal dens of hyenas (Supplementary Figure S1). The communal den is a valuable resource for hyenas to guard, as it contains the young offspring of all females in the clan (Kruuk 1972). Furthermore, lions are known to kill den-dwelling cubs when the opportunity arises (Mills 1990; Hofer and East 1995; Watts and Holekamp 2009), so hyena fitness should be enhanced by driving lions away from dens.
Lions and hyenas were more likely to interact as the number of hyenas increased (Table 2; Figure 1), which is not surprising given that the risks involved in interacting with lions can be more widely distributed when more hyenas are present. The probability of successfully defending or obtaining food from lions also likely increases with the number of allies present (Supplementary Table S3).
Lions and hyenas encountered each other more often as prey size increased (Table 3; Supplementary Figure S2). This is consistent with the notion that acceptable fight costs should increase with resource value (Enquist and Leimar 1987). Furthermore, hyenas can completely consume smaller kills in a few minutes (Kruuk 1972), leaving little or nothing for lions to kleptoparasitize. A large group of hyenas can ingest even medium-sized prey, such as an entire fresh topi or wildebeest, in as little as 13 min (Holekamp and Smale 1998). Local prey availability, however, did not significantly affect the probability of lions and hyenas interacting (Table 2), indicating that the intensity of interspecific competition is more situational (i.e., a fresh kill within hearing distance of both species) than dependent on larger scale environmental factors such as low prey abundance.
When lions and hyenas did clash over food, the species with control of the carcass when observers arrived was most likely to maintain possession of the food until the end of the session (Table 4). This is consistent with the hypothesis that the resource holder enjoys an inherent advantage over potential usurpers (Parker 1974). Interestingly, when a session began with hyenas in possession of the food, session length did not appear to affect their likelihood of retaining the food. In contrast, the longer the session, the more likely lions were to lose control of the food to hyenas (Supplementary Figure S3). This may indicate the hyena’s persistence in remaining near the food (Kruuk 1972), the lions’ willingness to abandon food once satiated (Kissui and Packer 2004), or the hyenas’ ability to recruit more allies, given enough time (Gersick et al. 2015).
Presence of male lions
Several past studies of lion–hyena interactions found that the presence of male lions either partially (Mills 1990) or completely (Elliott and Cowan 1978; Cooper 1991; Honer et al. 2002) prevented hyenas from obtaining food from lion-controlled carcasses. Here, although the presence of male lions increased the probability that lions and hyenas would interact (Table 2; Figure 1), it decreased the probability that hyenas would possess the food at the end of the session (Table 4; Figure 2). Male lions may increase the probability of interspecies interaction by instigating the interactions themselves, as male lions are more likely to approach feeding hyenas on a kill than are females (Elliott and Cowan 1978). Furthermore, males are known to stalk hyenas and even detour in attempts to kill them (Frank et al. 1995).
Once in possession of the carcass, male lions, due to their larger size, are far more effective than female lions at keeping hyenas at bay and preventing hyenas from obtaining food (Cooper 1991; Kissui and Packer 2004). Accordingly, our descriptive data revealed that hyenas were only able to obtain food from lion groups containing adult males at sessions with high hyena:lion ratios (Supplementary Table S3). Surprisingly, hyenas were nonetheless able to secure food in the presence of male lions on 26 of 93 occasions (Supplementary Table S3). We believe that our large sample of observations enabled us to document this otherwise rare behavior. We also documented mobbing of male lions in 25 of 121 sessions involving a kill or carcass, although mobbing rates were much lower when male lions were present than when they were absent. This suggests that, although hyenas view male lions as a source of added danger, they are nonetheless willing to risk approaching male lions when the potential benefits warrant it.
Variables predicting whether or not mobbing occurs
The number of hyenas present, rather than the presence of food, was the key variable predicting the occurrence of mobbing (Table 5; Figure 3). Mobbing in large groups may reduce the risk of injury, either by distributing it among participants or by making it more difficult for lions to select an individual to pursue (Miller 1922). This supports the notion that availability of potential support from conspecifics has a stronger effect on mobbing behavior than do either lions or the presence of food.
Nevertheless, mobbing rates were highest around food resources, especially fresh kills. However, mobbing rates did not vary based on the size of the prey animal killed, indicating that mobbing depends on more than just the resource value. Mobbing rates were next highest at hyena communal dens, where hyenas presumably mobbed to protect their young. Additionally, hyenas mobbed lions in 29 sessions without any discernable resource present to fight over, which is surprising due to the danger inherent in mobbing behavior. One possible explanation for this is that hyenas use mobbing to gather information about potential threats, as also occurs in meerkats (Graw and Manser 2007). Lions, like hyenas, are territorial, so mobbing hyenas can expect to re-encounter the same individual lions within their lifetimes (Packer et al. 1990). Hyena mobbing may thus have a broader function beyond deterrence of lions, in that it may facilitate situational risk assessment on which subsequent decisions by group-members are based.
Mobbing affects the probability that hyenas will feed
Cooperative mobbing significantly increased the hyenas’ ability to feed from lion-controlled carcasses, as the number of mobbing events observed in a session was a significant predictor of whether or not hyenas fed under these circumstances (Table 6; Figure 4). Thus, cooperation appears to enhance fitness in spotted hyenas by increasing their probability of feeding when competing with lions for control of a food resource. The ephemeral nature of kills and carcasses (Jones et al. 2015), and the lions’ ability to steal a significant proportion of hyena kills (Périquet et al. 2015), should theoretically have imposed strong selection pressure in the past, promoting the evolution of cooperative mobbing in this species.
Future directions
Given that cooperative mobbing does indeed increase access to food resources by hyenas, several further investigations now seem warranted. First, we will ask whether all group members who participate in mobbing events realize comparable direct benefits, and whether cheating occurs with individuals who fail to participate in mobs yet feed when food is usurped from lions. Our future work will also inquire about the individuals who participate in mobs, their age, sex, and rank, and the relationships among members of individual mobs. In intragroup aggression, adult females provide coalitionary support most frequently to their close kin regardless of intensity of aggression or risk of counterattack (Smith et al. 2010). However, evidence from previous work suggests that relatedness within a clan is not high enough to select for cooperation without additional direct benefits (Van Horn et al. 2004). Unless hyena mobs are composed of related individuals, a mechanism other than kin selection must maintain cooperative mobbing behavior. For example, ritualized greeting ceremonies reinforce social bonds in hyenas (Smith et al. 2011), and can provide a mechanism to assess and strengthen bonds with potential mobbing allies. Cooperative mobbing against lions might also serve a social bonding function through quality advertisement to potential mates and social allies (Dugatkin and Godin 1992b).
We are also interested in the mechanisms mediating collective movement during mobbing in spotted hyenas. Social facilitation is a predominant feature of hyena behavior (Glickman et al. 1997), and promotes coalition formation among captive juveniles (Zabel et al. 1992). Social facilitation could function to synchronize movement in mobbing behavior (Zabel et al. 1992) and has been hypothesized to play a role in the development of cooperative behavior (Glickman et al. 1997; Drea and Carter 2009). Furthermore, in contrast with intraspecific coalitionary aggression, lion–hyena interactions are accompanied by raucous signaling. It is unclear whether these vocalizations function to facilitate cooperation or to intimidate the mobbed lions. One of these, the lowing vocalization, is usually emitted during mobbing and may help individuals synchronize their movements (Kruuk 1972).
Given that risk-taking behavior has been correlated with hormone concentrations in a variety of species (e.g., Martins et al. 2007; Stanton et al. 2011), future work should focus on the hormones modulating behavior in participating hyenas before and during these cooperative encounters. Understanding the mechanisms maintaining cooperation, including the relevant communication signals and endocrine mediation of mobbing, will undoubtedly enhance our understanding of the relationships among communication, cognition, and cooperation.
Supplementary Material
Acknowledgments
We are indebted to Marc Wiseman for his insights early in project development. Nazli Uygen provided generous statistical support. We thank H.E. Watts & W.W. Dowd for collection of Amboseli data. We thank the Kenyan National Commission for Science, Technology and Innovation, the Kenya Wildlife Service, the Mara Conservancy, and the Narok County Government for permission to conduct this work.
Funding
This work was funded by National Institute of Health Grant 1R01GM105042 and National Science FoundationF grants IOS1121474 and DEB1353110 to K.E.H., and by National Science Foundation Cooperative Agreement DBI 0939454 supporting the BEACON Center for the Study of Evolution in Action. K.D.S.L. and T.M.M. were generously supported by National Science Foundation Graduate Research Fellowships, and T.M.M. was also supported by a University Distinguished Fellowship from MSU.
Supplementary Material
Supplementary material can be found at http://www.cz.oxfordjournals.org/.
References
- Altmann SA, 1956. Avian mobbing behavior and predator recognition. Condor 58:241–253. [Google Scholar]
- Altmann J, 1974. Observational study of behavior: sampling methods. Behaviour 49:227–265. [DOI] [PubMed] [Google Scholar]
- Bailey I, Myatt JP, Wilson AM, 2013. Group hunting within the Carnivora: physiological, cognitive and environmental influences on strategy and cooperation. Behav Ecol Sociobiol 67:1–17. [Google Scholar]
- Boydston EE, Kapheim KM, Szykman M, Holekamp KE, 2003. Individual variation in space use by female spotted hyenas. J Mammal 84:1006–1018. [Google Scholar]
- Brosnan SF, Bshary R, 2010. Cooperation and deception: from evolution to mechanisms. Philos Trans R Soc London B Biol Sci 365:2593–2598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooper SM, 1991. Optimal hunting group size: the need for lions to defend their kills against loss to spotted hyaenas. Afr J Ecol 29:130–136. [Google Scholar]
- De Stephanis R, Giménez J, Esteban R, Gauffier P, García-Tiscar S. et al. , 2015. Mobbing-like behavior by pilot whales towards killer whales: a response to resource competition or perceived predation risk? Acta Ethol 18:69–78. [Google Scholar]
- Drea CM, Carter AN, 2009. Cooperative problem solving in a social carnivore. Anim Behav 78:967–977. [Google Scholar]
- Dugatkin LA, Mesterton-Gibbons M, Houston AI, 1992a. Beyond the prisoner’s dilemma: toward models to discriminate among mechanisms of cooperation in nature. Tren Ecol Evol 7:202–205. [DOI] [PubMed] [Google Scholar]
- Dugatkin LA, Godin JJ, 1992b. Prey approaching predators: a cost-benefit perspective. Ann Zool Fenn 29:233–252. [Google Scholar]
- Elliott JP, Cowan IM, 1978. Territoriality, density, and prey of lion in Ngorongoro-Crater, Tanzania. Can J Zool 56:1726–1734. [Google Scholar]
- Engh AL, Esch K, Smale L, Holekamp KE, 2000. Mechanisms of maternal rank “inheritance” in the spotted hyaena Crocuta crocuta. Anim Behav 60:323–332. [DOI] [PubMed] [Google Scholar]
- Enquist M, Leimar O, 1987. Evolution of fighting behaviour: the effect of variation in resource value. J Theor Biol 127:187–205. [Google Scholar]
- Frank LG, 1986. Social organization of the spotted hyaena Crocuta crocuta. II. Dominance and reproduction. Anim Behav 151:1510–1527. [Google Scholar]
- Frank LG, Glickman SE, Powch I, 1990. Sexual dimorphism in the spotted hyaena Crocuta crocuta. J Zool 221:308–313. [Google Scholar]
- Frank LG, Holekamp KE, Smale L, 1995. Dominance, demography, and reproductive success of female spotted hyenas In: Sinclair ARE, Arcese P, editors. Serengeti II: Dynamics, Management, and Conservation of an Ecosystem. Chicago: University of Chicago Press, 364–384. [Google Scholar]
- Gersick AS, Cheney DL, Schneider JM, Seyfarth RM, Holekamp KE, 2015. Long-distance communication facilitates cooperation among wild spotted hyaenas Crocuta crocuta. Anim Behav 103:107–116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glickman SE, Zabel CJ, Yoerg SI, Weldele ML, Drea CM. et al. , 1997. Social facilitation, affiliation, and dominance in the social life of spotted hyenas In: Carter CS, Lederhendler II, Kirkpatrick B, editors. The Integrative Neurobiology of Affiliation. Cambridge: MIT Press, 131–140. [DOI] [PubMed] [Google Scholar]
- Graw B, Manser MB, 2007. The function of mobbing in cooperative meerkats. Anim Behav 74:507–517. [Google Scholar]
- Green B, Anderson J, Whateley T, 1984. Water and sodium turnover and estimated food consumption in free-living lions Panthera leo and spotted hyaenas Crocuta crocuta. J Mammal 65:593–599. [Google Scholar]
- Green DS, 2015. Anthropogenic Disturbance, Ecological Change, and Wildlife Conservation at the Edge of the Mara-Serengeti Ecosystem [Doctoral Thesis]. East Lansing (MI): Michigan State University.
- Gursky-Doyen S, Nekaris KA-I, 2007. Primate Anti-Predator Strategies. New York (NY): Springer Science & Business Media. [Google Scholar]
- Hammerstein P, 2003. Genetic and Cultural Evolution of Cooperation. Cambridge: MIT Press. [Google Scholar]
- Hayward MW, 2006. Prey preferences of the spotted hyaena Crocuta crocuta and degree of dietary overlap with the lion Panthera leo. J Zool 270:606–614. [Google Scholar]
- Henschel JR, Skinner JD, 1990. The diet of the spotted hyaenas Crocuta crocuta in Kruger National Park. Afr J Ecol 28:69–82. [Google Scholar]
- Henschel JR, Skinner JD, 1991. Territorial behaviour by a clan of spotted hyaenas Crocuta crocuta. Ethology 88:223–235. [Google Scholar]
- Hofer H, East M, 1995. Population dynamics, population size, and the commuting system of Serengeti spotted hyenas In: Sinclair ARE, Arcese P, editors. Serengeti II: Dynamics, Management, and Conservation of an Ecosystem. Chicago: University of Chicago Press, 332–363. [Google Scholar]
- Holekamp KE, Smale L, 1998. Behavioral development in the spotted hyena. BioScience 48:997–1005. [Google Scholar]
- Holekamp KE, Szykman M, Boydston EE, Smale L, 1999. Association of seasonal reproductive patterns with changing food availability in an equatorial carnivore, the spotted hyaena Crocuta crocuta. J Reprod Fertil 116:87–93. [DOI] [PubMed] [Google Scholar]
- Holekamp KE, Dantzer B, Stricker G, Yoshida KCS, Benson-Amram S, 2015. Brains, brawn and sociality: a hyaena’s tale. Anim Behav 103:237–248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Honer OP, Wachter B, East ML, Hofer H, 2002. The response of spotted hyaenas to long-term changes in prey populations: functional response and interspecific kleptoparasitism. J Anim Ecol 71:236–246. [Google Scholar]
- Janzen DH, 1970. Altruism by coatis in the face of predation by Boa constrictor. J Mammal 51:387–389. [Google Scholar]
- Jones SC, Strauss ED, Holekamp KE, 2015. Ecology of African carrion In: Benbow ME, Tomberlin JK, Tarone AM, editors. Carrion Ecology, Evolution and Their Applications. Boca Raton (FL): CRC Press, 461–491. [Google Scholar]
- Kissui BM, Packer C, 2004. Top-down population regulation of a top predator: lions in the Ngorongoro Crater. Proc R Soc B Biol Sci 271:1867–1874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kolowski JM, Katan D, Theis KR, Holekamp KE, 2007. Daily patterns of activity in the spotted hyena. J Mammal 88:1017–1028. [Google Scholar]
- Kruuk H, Turner M, 1967. Comparative notes on predation by lion, leopard, cheetah and wild dog in the Serengeti area, East Africa. Mammalia 31:1–27. [Google Scholar]
- Kruuk H, 1972. The Spotted Hyena: A Study of Predation and Social Behavior. Chicago: University of Chicago Press. [Google Scholar]
- Martins TLF, Roberts ML, Giblin I, Huxham R, Evans MR, 2007. Speed of exploration and risk-taking behavior are linked to corticosterone titres in zebra finches. Horm Behav 52:445–453. [DOI] [PubMed] [Google Scholar]
- Miller RC, 1922. The significance of the gregarious habit. Ecology 3:122–126. [Google Scholar]
- Mills MGL, 1990. Kalahari Hyenas: Comparative Behavioral Ecology of Two Species. London: Unwin Hyman Ltd. [Google Scholar]
- Moehlman PD, Hofer H, 1997. Cooperative breeding, reproductive suppression, and body mass in canids In: Solomon NG, French JA, editors. Cooperative Breeding in Mammals. Cambridge: Cambridge University Press, 76–128. [Google Scholar]
- Noë R, 2006. Cooperation experiments: coordination through communication versus acting apart together. Anim Behav 71:1–18. [Google Scholar]
- Owings DH, Coss RG, 1977. Snake mobbing by California ground squirrels: adaptive variation and ontogeny. Behaviour 62:50–68. [Google Scholar]
- Packer C, Scheel D, Pusey AE, 1990. Why lions form groups: food is not enough. Am Nat 135:1–19. [Google Scholar]
- Parker GA, 1974. Assessment strategy and the evolution of fighting behaviour. J Theor Biol 47:223–243. [DOI] [PubMed] [Google Scholar]
- Périquet S, Fritz H, Revilla E, 2015. The lion king and the hyaena queen: large carnivore interactions and coexistence. Biol Rev 90:1197–1214. [DOI] [PubMed] [Google Scholar]
- Rood JP, 1975. Population dynamics and food habits of the banded mongoose. Afr J Ecol 13:89–111. [Google Scholar]
- Sachs R, 1967. Liveweights and body measurements of Serengeti game animals. Afr J Ecol 5:24–36. [Google Scholar]
- Sandoval L, Wilson DR, 2012. Local predation pressure predicts the strength of mobbing responses in tropical birds. Curr Zool 58:781–790. [Google Scholar]
- Schaller GB, 1972. The Serengeti Lion: A Study of Predator-Prey Relations. Chicago: University of Chicago Press. [Google Scholar]
- Smith JE, Kolowski JM, Graham KE, Dawes SE, Holekamp KE, 2008. Social and ecological determinants of fission - fusion dynamics in the spotted hyaena. Anim Behav 76:619–636. [Google Scholar]
- Smith JE, Van Horn RC, Powning KS, Cole AR, Graham KE. et al. , 2010. Evolutionary forces favoring intragroup coalitions among spotted hyenas and other animals. Behav Ecol 21:284–303. [Google Scholar]
- Smith JE, Powning KS, Dawes SE, Estrada JR, Hopper AL. et al. , 2011. Greetings promote cooperation and reinforce social bonds among spotted hyaenas. Anim Behav 81:401–415. [Google Scholar]
- Stanton SJ, Liening SH, Schultheiss OC, 2011. Testosterone is positively associated with risk taking in the Iowa Gambling Task. Horm Behav 59:252–256. [DOI] [PubMed] [Google Scholar]
- Stephens D, Anderson J, 1996. Reply to Roberts: cooperation is an outcome, not a mechanism. Anim Behav 53:1363–1364. [DOI] [PubMed] [Google Scholar]
- Trinkel M, Kastberger G, 2005. Competitive interactions between spotted hyenas and lions in the Etosha National Park, Namibia. Afr J Ecol 43:220–224. [Google Scholar]
- Van Horn RC, Engh AL, Scribner KT, Funk SM, Holekamp KE, 2004. Behavioural structuring of relatedness in the spotted hyena Crocuta crocuta suggests direct fitness benefits of clan-level cooperation. Mol Ecol 13:449–458. [DOI] [PubMed] [Google Scholar]
- Watts HE, Holekamp KE, 2008. Interspecific competition influences reproduction in spotted hyenas. J Zool 276:402–410. [Google Scholar]
- Watts HE, Holekamp KE, 2009. Ecological determinants of survival and reproduction in the spotted hyena. J Mammal 90:461–471. [Google Scholar]
- Whitman KL, Packer C, 2006. A Hunter’s Guide to Aging Lions in Eastern and Southern Africa. Huntington Beach, CA: Conservation Force. [Google Scholar]
- Zabel CJ, Glickman SE, Frank LG, Woodmansee KB, Keppel G, 1992. Coalition formation in a colony of prepubertal spotted hyenas In: Harcourt AH, De Waal FBM, editors. Coalitions and Alliances in Humans and Other Animals. New York: Oxford University Press, 113–135. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.