

Abstract
Background:
Proof of the efficacy and safety of a xenogeneic mesenchymal stem cell (MSCs) transplant for spinal cord injury (SCI) may theoretically widen the spectrum of possible grafts for neuroregeneration.
Methods:
Twenty rats were submitted to complete spinal cord transection. Ovine bone marrow MSCs, retrovirally transfected with red fluorescent protein and not previously induced for neuroglial differentiation, were applied in 10 study rats (MSCG). Fibrin glue was injected in 10 control rats (FGG). All rats were evaluated on a weekly basis and scored using the Basso–Beattie–Bresnahan (BBB) locomotor scale for 10 weeks, when the collected data were statistically analyzed. The spinal cords were then harvested and analyzed with light microscopy, immunohistochemistry, and immunofluorescence.
Results:
Ovine MSCs culture showed positivity for Nestin. MSCG had a significant and durable recovery of motor functions (P <.001). Red fluorescence was found at the injury sites in MSCG. Positivity for Nestin, tubulin βIII, NG2 glia, neuron-specific enolase, vimentin, and 200 kD neurofilament were also found at the same sites.
Conclusions:
Xenogeneic ovine bone marrow MSCs proved capable of engrafting into the injured rat spinal cord. Transdifferentiation into a neuroglial phenotype was able to support partial functional recovery.
Keywords: Mesenchymal stem cells, neuroglial differentiation, neurological recovery, red fluorescent protein, spinal cord injury
INTRODUCTION
Spinal cord injury (SCI) is a devastating condition affecting millions of people every year worldwide, especially young males. The overall number of paraplegic and quadriplegic people living in the U.S. is about 1.5 million, with an annual global cost for the healthcare system exceeding $10 billion.[57,65] No cure for the catastrophic neurological sequelae correlated to SCI has materialized despite intensive investigation over several decades.
Mesenchymal stem cells (MSC) are potential candidates for a possible cell-based cure of SCI[2,13,15,20,23,25,28,30,31,32,33,36,41,44,45,47,51,52,64,65,68] due to their very low immunogenicity, lack of alloreactivity, easiness of isolation and culture, local immunosuppressive and angiogenetic potential, wide availability, lack of ethical concerns, and, above all, a well-known potential for tissue-specific differentiation. Studies up to now have focused on autologous or allogenic MSCs out of immunological concerns. However, the demonstration of safety and efficacy of a cross-species xenogeneic MSCs transplants could lead to an unlimited supply of replacement cells.
The present study aims at assessing the potential for engraftment, neurodifferentiation, and functional neuroglial regeneration following xenogeneic ovine bone marrow-derived MSCs in a rat model of complete spinal cord transection.
MATERIALS AND METHODS
Study design
All experiments were approved by the ethical committee of the University of Bari (Aldo Moro), Italy, in accordance with the national animal welfare legislation and in compliance with the guidelines outlined in the NRC Guide for the Care and Use of Laboratory Animals.
The first step of the study protocol involved the collection, isolation, and culture of bone marrow MSCs from healthy donor sheep and immunofluorescence analysis of immature neuroglial markers expression. MSCs were then retrovirally transfected with Red Fluorescent Protein (dsRFP) gene. Transfection with dsRFP would enable the cell to synthesize the marker as evidence of engraftment into the injured rat spinal cord. Subsequently, 20 Wistar rats underwent a pretreatment behavioral assessment by means of the Basso–Beattie–Bresnahan scoring system (BBB score)[3] and split in two groups, each consisting of ten rats. The first group was denoted as Mesenchymal Stem Cell Group (MSCG), whereas the second group as Fibrin Glue Group (FGG). All the rats were then anesthetized and submitted to a mid-thoracic laminectomy and complete spinal cord transection. During the same surgery, a solution of ovine bone marrow MSCs previously transfected with dsRFP gene (Red-MSCs) and fibrin glue was applied at the injury site in the MSCG. Conversely, a solution containing only fibrin glue was applied in the FGG. Careful monitoring of weight and bladder expression was performed postoperatively and antibiotic therapy was started in case of infection. Both groups were assessed on the BBB scale immediately after surgery and then weekly for 10 weeks; a short digital video of each rat was recorded at each follow-up. BBB scores collected in both groups were statistically analyzed. All continuous data were expressed as mean ± standard deviation. The comparison of BBB scores was done by repeated-measures analysis of variance (ANOVA) and post-hoc tests. The level of significance was set at P <.05. At the end of the follow-up, all the rats were sacrificed and their spinal cords were harvested. Three blocks, obtained at above and below the injury site of each spinal cord, were prepared for fluorescence and immunohistochemical examination. Fluorescence microscopy analysis defined the degree of engraftment of Red-MSCs and their spatial localization. Immunohistochemical analysis of the MSCG specimens aimed to assess immune-pattern of the cellular phenotypes at the injury sites. See study design in Figure 1.
Figure 1.

Diagram showing the design and experiments of the study. BM-MSCs: bone marrow mesenchymal stem cells; FACS: Fluorescence Activated Cell Sorting; Red-MSCs: mesenchymal stem cells retrovirally transfected with dsRFP gene; BBB: Basso-Beattie-Bresnahan score; MSCG: Mesenchymal Stem Cells Group; FGG: Fibrin Glue Group
Xenogeneic ovine bone marrow collection and isolation and culture of MSCs
A healthy donor sheep (Bergamasca breed, 2 year-old females, 45 kg in weight, not inbred) was selected as the source of MSCs. The donor sheep was sedated with diazepam (0.25 mg/kg) and the area of the tuber coxae was surgically prepared. Lidocaine (2%, 20 mL) was injected subcutaneously. A 14-gauge Jamshidi needle was inserted into the tuber coxae, and a 50 ml heparinized syringe was used to collect 30 ml of bone marrow. Bone marrow mononuclear cells were isolated on a Biocoll separating solution by centrifugation at 425 g for 30 minutes. The separated cells were counted with a nuclear stain (0.1% methyl violet in 0.1 M citric acid), rinsed twice with phosphate buffered saline (PBS), and suspended in an adequate amount of fibrin glue (Tisseel Baxter BioScience™, Munich, Germany). Cells were incubated at 37°C in a humidified 5% CO2 atmosphere for 2 weeks, replacing the medium twice a week. The cells were then washed with PBS, pH 7.2, fixed with buffered 4% formalin, and stained with 1% methylene blue in borate buffer (10 mM, pH 8.8). The mononuclear bone marrow cells were isolated as described above and seeded in flasks at a concentration of 4–5 × 106 cells/cm2 in complete Coon's medium at 37°C in a humidified 5% CO2 atmosphere. The medium was replaced twice a week until the cells reached 30% confluence.
Fluorescence activated cell sorting
Fluorescence activated cell sorting (FACS) was employed to assess the immunophenotype of MSCs in terms of cluster of differentiation (CD). The list of the CD targets selected to assess the native immunophenotype of the ovine bone marrow MSCs, with the relative percentage of positivity for each tested CD, is presented in Table 1.
Table 1.
CD immunophenotype of the ovine bone marrow MSCs’ colture

Immunofluorescence cytochemical study of native neuroglial markers expression in MSCs
The cells were fixed for 20 min in 4% paraformaldehyde in phosphate-buffered saline (pH 7·4), and then washed and incubated for 90 min at 37°C with PBS plus 0.3% Triton containing 10% normal goat serum and the appropriate antibody mixture. The specific antibodies used to detect the antigens were anti-human glial fibrillary acidic protein (GFAP), NG2, mouse anti-human tubulin βIII (b-tub-III), Nestin (Chemicon, Temecula, CA, USA), rabbit anti-Neurofilaments subunits 200 kD (NF-01), rabbit anti-choline acetyltransferase (ChAT) (Sigma-Aldrich, St. Louis, MO, USA), mouse anti-oligodendrocyte marker O4 (O4) (R&D Systems, MN, USA). The cells’ culture was then washed twice with PBS and incubated with an appropriate secondary antibody: anti-rabbit IgG (1:1000), anti-mouse IgG (1:500) (Cy2 and Cy3; Jackson Immunoresearch) for 45 min at room temperature. The cells were incubated with 4,6- diamidino-2-phenylinole dihydrochloride (DAPI) (1 g/L in methanol, 15 min at 37°C) (Sigma-Aldrich, St. Louis, MO, USA) and Fluorsave™ (Calbiochem, La Jolla, USA) and viewed under a Zeiss Axiophot-2 microscope.
The list of early neuronal and glial targets tested on MSCs before the transplant is shown in Table 2.
Table 2.
Native immunocytochemical neuro-glial phenotype of the ovine bone marrow MSCs’ colture

dsRFP cells labeling
dsRFP gene was retrovirally transfected into MSCs cultures by Dr. Andrea Levi at the Neurobiology and Molecular Medicine Branch of C.N.R. (Centro Nazionale delle Ricerche), Rome, Italy. The retrovirus coding for dsRFP (Retro V-RFP) was added along with 200 μL of serum-free medium, and the flask was shaken gently every 15 minutes for 2 hours. The passage 5 MSCs were transfected with the recombinant replication-defective retrovirus at different multiplicities of infection in the range of 0–200 units. After incubation with Retro V-RFP for 2 hours, culture medium containing 10% fetal bovine serum was added into the flask. The transfected MSCs were cultured for 72 hours and then microscopically observed. Three high-power fields of view were selected randomly to count the cells positive for retroviral transfection. The estimated transfection rate was 88.3%. The cells obtained by retroviral transfection of dsRFP gene were referred as Red-MSCs.
Pretreatment behavioral appraisal
Twenty rats (Wistar, 200–250 g) of either sex, sex not affecting the potential for neurologic recovery in SCI, were evaluated for behavioral analysis by the BBB open field locomotor scale.[3] A score was assigned to each rat beginning preoperatively (T pre). The rats were then divided in two Groups, each 10 strong (MSCG versus FGG).
Xenogeneic MSCs transplantation
The rats were anesthetized according to a standard protocol involving the intraperitoneal administration of Fentanyl/Fluanisone (0.3 mg/kg i.p., Hypnorm, Janssen, Belgium) and Midazolam (0.6 mg/kg i.p., Dormicum, Roche, Switzerland), then shaved, placed in a heating pad, achieving a body temperature at 37°C during the entire length of surgery, and underwent a standard mid-thoracic two-level laminectomy (T8-T9). The dura was carefully opened and the spinal cord was exposed. Under microscopic view (4×), a complete transection of the cord was performed making a single knife cut with a standard surgical scalpel blade no. 11 mounted on a Bard Parker surgical blade handle #3. All the cord transections were performed in an axial plane and according to the same standard technique in both groups. Thereafter, a solution containing 6 × 106 cells/mL Red-MSCs and fibrin glue (Tisseel, Baxter BioScience™) was applied at the cord transection level using a microsyringe in the MSCG group. Conversely, only fibrin glue was applied in the FGG rats. The dura was then closed in a running watertight fashion to avoid early or late central nervous system infections; muscles and skin were re-approximated with interrupted sutures [Figure 2].
Figure 2.

Rat spinal cord transection. (a) preparation and shaving of the rat; (b) skeletonization of the spine for mid-thoracic laminectomy; (c and d) spinal cord transection; (e) release of the solution containing Red-MSCs and fibrin glue at the injury site in MSCG. In some cases, the permeation of the fibrin glue caused a partial dissociation of the main bundles of the spinal cord
A standard postoperative analgesic protocol involving the administration of Buprenorphine (0.03 mg/kg, s.c., Temgesic, Reckitt & Colman, UK) was enacted during the early postoperative period. All the rats were evaluated for behavioral recovery within 24 hours of surgery (T-post), and housed under a 12:12-h light/dark cycle with water and food ad libitum. Careful monitoring of rats with daily weight recordings and bladder expression allowed for early detection of postoperative complications. Trimethoprim (0.83 ml/kg, i.p., Bactrim, Roche, Switzerland) was promptly started in case of infection.
Posttreatment behavioral follow-up
In addition to early postoperative evaluation (T-post), all the rats were evaluated weekly until the 70th posttreatment day. A short digital video was recorded for each rat at each behavioral evaluation. Data collected in both groups during the entire follow-up were statistically analyzed using a commercial available software (GraphPad Prism - La Jolla, CA, USA). All values were expressed as mean ± standard deviation. Differences between groups in terms of BBB scores were assessed by repeated-measure ANOVA and post-hoc test, and the level of significance was set at P <.05.
Light microscopy
Within 1 week from the last clinical and behavioral evaluation, all the rats were sacrificed with a pentobarbital overdose (50 mg/100 g), and all the previously injured spinal cords were harvested. Each cord was divided into three equal blocks of 5 mm in length. The first block was harvested at the maximal injury zone, whereas the other two blocks were sampled just above and below the cord transection.
Each block was fixed in 10% neutral buffered formalin for 48 hours. The cords were then washed, dehydrated in a series of passages through ethanol solutions at different concentrations (40% to 100%) and embedded in paraffin wax. The specimens were frozen in isopentane at −40°C and then cut in sections of 3 m thickness. The sections were then stained with hematoxylin-eosin (HE) for pathological examination. A series of digital photographs was obtained for both groups.
dsRFP cells tracking
In the MSCG, Red-MSCs tracking was achieved by fluorescence microscopy (Olympus BX61) at a 20 × magnification (absorption/emission peak 458 nm and 583 nm, respectively). The total amount of fluorescence was measured for each field at the same magnification by commercially available software (Image J, Windows Excel Microsoft Corporation). For each field, the background fluorescence was subtracted to avoid any potentially biasing factor and to obtain the real total fluorescence of the specimen. Images were digitally recorded.
Immunohistochemistry
Immunohistochemical study of the MSCG spinal cord specimens was performed to confirm the presence – and thus putative engraftment – of the xenogeneic bone marrow MSCs at the injury site. On the basis of the already reported immunohistochemical phenotype of ovine bone marrow MSCs,[12,14,35] a solution containing mouse monoclonal antibodies against CD34 (Zymed Inc., San Francisco, CA, USA), CD44-biotin, Nestin, fibroblast grow factor-1(FGF1), GFAP, NG2, neuron specific enolase (NSE), Vimentin, NF-01 (Abcam, Cambridge, MA, USA), CD45-APC, nerve grow factor receptor (NGFR), b-tub-III, ChAT, (Sigma-Aldrich, St. Louis, MO, USA), CD54-APC, CD73-PE, (BioLegend San Diego, CA, USA), CD90-Biotin (BD Bioscience, San Jose, CA, USA), CD166-PE (RDsystems, Minneapolis, MN, USA) was used for immunohistochemical staining. In case of biotin labeling, a second incubation with streptavidin–PerCP (BD Bioscience) was done (1:100 in blocking solution, 1 h RT). Tris-buffered solution or nonimmune sera were substituted for specific primary antibodies as negative controls. Sections were placed on pretreated slides (Bio-Optica, Milan, Italy) to promote adhesion and dried overnight at 37°C. After de-waxing, the sections were placed in ethylenediaminetetra-acetic acid buffer, pH 9.0, and processed in a microwave oven at 650 W for two cycles of 10 minutes each to enhance antigenicity. Slides were then allowed to cool at room temperature for at least 20 minutes before further processing for immunostaining, employing standard procedures. Tissue sections were incubated overnight in a moist chamber at 4°C with primary antibodies diluted 1:50 in TBS containing 0.1% crystalline bovine serum albumin. Binding of the antibodies was detected with ABC-peroxidase (Vector Laboratories Inc., Burlingame, CA, USA) techniques using 1:200 diluted biotin-conjugated goat anti-rabbit immunoglobulin G (Vector Laboratories Inc., Burlingame, CA, USA) and a 1:200 diluted biotinylated goat anti-mouse immunoglobulin (AO433: DAKO, Glostrup, Denmark) as secondary antibodies applied for 45 minutes at room temperature. The enzymatic reaction was developed using 3,1-diaminobenzidinel or Vector® VIPm as a substrate for the ABC-peroxidase and Mayer's hematoxylin as nuclear counterstain. For scoring puposes of each CD+ expression at different spinal cord sites, stained cells were quantified in different tissue areas around the injury sites. All cellular types were calculated using a light microscope (Carl Zeiss: Oberkochen, Germany), a 40 × objective, a 10× eyepiece, and a square eyepiece graticule (10 Å~ 10 squares, with a total area of 62,500 μm2). Five randomly selected sites were chosen to assess each marker and arithmetic means were calculated for each site. Results were expressed as immunohistochemical positive cells per 62,500 μm2. Multiple digital photographs were recorded. The list of neuroglial markers tested on the spinal cord specimens at the injury sites is shown in Table 3.
Table 3.
Neuro-glial pattern expression of spinal cord specimens at the injury sites

RESULTS
Evidence of stemness and native immunophenotype of MSCs
Immunostaining positivity for CD 44, CD 54, CD 73, CD 90 and CD 166 was found in ovine MSCs culture. A lack of expression of CD 34 and CD 45 confirmed the stemness of these cells [Figure 3]. MSCs resulted positive for the surface expression of Nestin and negative for GFAP, NG2, b-tubIII [Figure 4].
Figure 3.

FACS immunophenotype of MSCs for CD 34 (a), CD 44 (b), CD 45 (c), CD54 (d), CD90 (e), CD166 (f), CD73 (g). PE: Phycoerythrin; FITC: Fluorescein isothiocyanate
Figure 4.
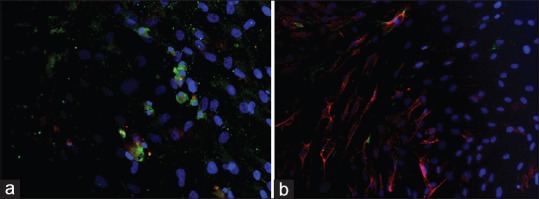
Native neuroglial immunophenotype of ovine bone marrow MSCs evaluated in fluorescence microscopy. (a) b-tub-III (red) + GFAP (green) + DAPI (10×); (b) Nestin (red) + NG2 (green) + DAPI (10×)
Surgery
All surgeries were performed successfully. Particularly, the use of a very sharp knife (blade no. 11), together with a microscopic detailed visualization of the surgical field, allowed micrometric axial cuts of the spinal cords: the gap between the proximal and distal stumps of the cord never exceed 4 mm in length. This aspect was considered paramount to minimize damage of the cord, both at the white matter and gray matter level, and, ultimately, to create the anatomical assumptions for the regrowth of neurites out of the gray matter core which is the vital component of the so-called cortico-trunco-reticulo-proprio-spinal pathway.[5–7] The assessment of the completeness of the spinal cord transection was achieved intraoperatively under microscopic vision by gentle elevation of the two stumps with a Penfield n. 2 periosteal elevator, as shown in Figure 2d. The two stumps were then re-approximated in their native position leaving a millimetric gap. No intraoperative or early postoperative complications were seen in both groups.
Posttreatment behavioral appraisal
Preoperatively, all rats showed a normal behavioral score; immediately after surgery, the BBB score dramatically decreased to a near zero level in both groups as an expression of the completeness of the lesional surgery.
In the MSCG, in the third week, one rat was lost due to severe self-mutilations. At the ninth week, two rats reached a score of 14 consisting of plantar stepping with full weight support and complete forelimb–hindlimb coordination[3] (see Supplemental Digital Content 1 (1.4MB, pdf) ).
STROBE Statement—Checklist of items that should be included in reports of case-control studies
In the FGG, two rats were lost in the second week and a another in the fourth week, all because of self-mutilations. The remaining rats did not experience a significant functional recovery [Graph 1].
Graph 1.
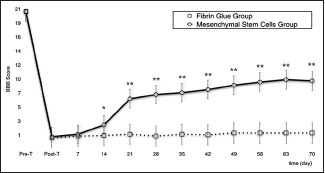
Line graph showing changes of BBB score in MSCG vs. FGG during the entire follow-up. *P <.05; **P <.001
Statistical analysis
In all treated rats, a difference between T-pre and T-post BBB scores was found (P <.001). Comparison between MSCG and FGG evinced a difference in terms of functional recovery of the MSCG over the FGG starting from the 14th day of behavioral assessment (P <.05). This difference between groups was confirmed throughout the follow-up (P <.001) [Graph 1].
Light microscopy
In the group of rats treated exclusively with fibrin glue injection, a general nonspecific pattern of severe glial scarring with scattered and necrotic large nucleated neurons was found. The scar tended to form bridges between the previously transected spinal cord stumps. It involved the entire axial plane of the cord up to 1 cm from the injury site [Figure 5a and b].
Figure 5.
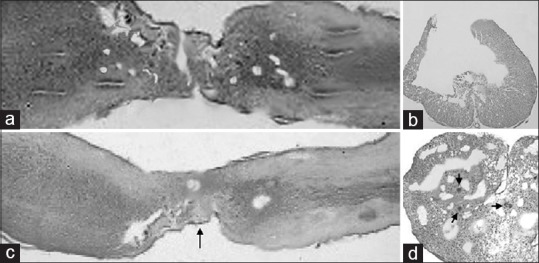
Light microscopy in FGG (a, b) vs. MSCG (c and d) at the injury site 10 weeks after injury. (a) FGG sagittal (HE scale bars = 400 μm); (b) FGG axial cut at the level of glial scar (HE scale bars = 200 μm); (c) MSCG sagittal (HE scale bars = 400 μm). Black arrow indicates the neuroglial bridges between the spinal cord stumps at the injury site. (d) MSCG axial (HE scale bars = 200 μm). Black arrows show a tight distribution of the neuroglial cells around the vessels
In the MSCG, HE staining revealed a tighter distribution of neuroglial cells, especially near the nerve stumps and around the vessels [Figure 5c and d]. The global cytoarchitecture at the injury sites appeared to be slightly less distorted compared to the FGG.
Fluorescence microscopy
Fluorescent microscopy allowed clear tracking of the presence of red fluorescence emission, attributable to a full engraftment of the transplanted Red-MSCs, at the cord injury sites in the MSCG. Fluorescence emission was generally arranged in the form of clusters of multiple red spots, which perfectly matched with the distribution of the MSCs at the injury site [Figure 6].
Figure 6.
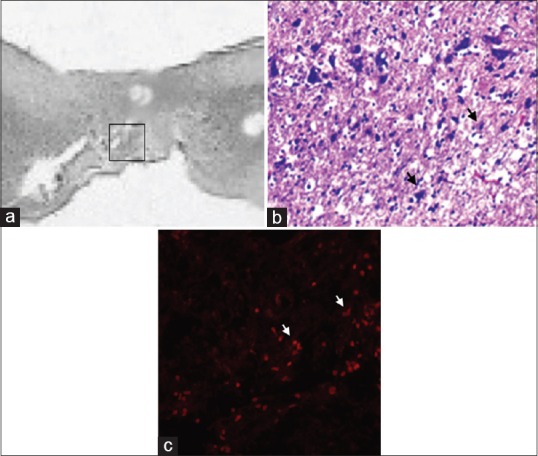
(a) Sagittal section (HE scale bars = 400 μm) of the harvested rat spinal cord in MSCG at 10th week after injury. Squared area indicates the site of sampling. (b) distribution of MSCs (black arrows) at the injury site in the MSCG (HE × 40); (c) co-localizzation of dsRFP MSCs (white arrows) within the injury site (fluorescence microscopy, excitation light 568 nm)
Immunohistochemistry
In the MSCG, positivity for Nestin, NG2, β-III tubulin, Vimentin, NF-01, and NSE immunostaining was found in all but two rats at the injury sites [Figure 7]. Interestingly, both rats, in which a lack of positivity for Nestin and β-III tubulin was found, had the worst BBB score in week 10.
Figure 7.
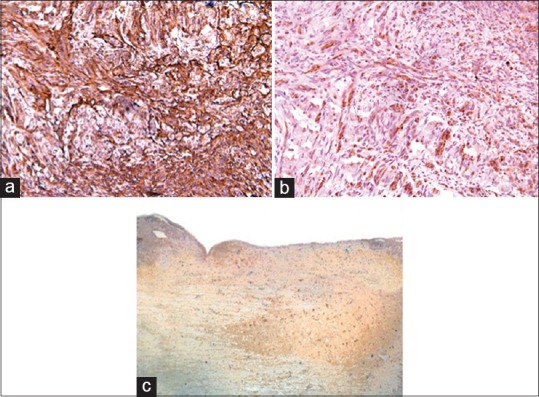
Immunostaining pattern for vimentin (a), NF-01 (b) and NSE (c) of the harvested spinal cords at the injury sites in MSCG
In the FGG, only very weak positivity for Vimentin was found probably associated with the fibroblastic component of the scar. No markers of neuroglial differentiation were detected in this group.
DISCUSSION
SCI involves both an acute and a chronic phase. The acute phase is characterized by blood vessels disruption, extensive posttraumatic cell death, recruitment of macrophages, neutrophils, and leukocytes tasked with cell debris phagocytosis and containment of further tissue damage.[38,46,55,60,63] Conversely, the chronic phase is dominated by glial scarring, sustained by reactive astrocytes at the injury site, with a subsequent local release of inhibitory proteins and axonal growth inhibitors able.[17,59,63] Many compounds have been tested in human SCI, targeting these mechanisms, including methylprednisolone sodium succinate, tirilazad mesylate, GM-1 ganglioside, thyrotropin-releasing hormone, gacyclidine, naloxone, and nimodipine, all with disappointing results.[11,18,21,22,26,27,37,58] Recently, the focus has shifted to myelin-associated and glial scar-associated inhibitors,[4,10,24,34,39,42,43,66] as these two classes of drugs seem to promote axonal growth and sprouting. However, initial human application so far has not been promising. The role of surgery is still limited to decompressive procedures in the acute phase of SCI aimed to restore the original volume of the spinal canal, to alleviate the compressive effects on the spinal cord, and to re-establish spinal alignment.
In 1980, Aguayo's group published a seminal work where they demonstrated, for the first time, the intrinsic ability the central nervous system (CNS) axons to regenerate and grow into the peripheral nerve system.[50] Twelve years later, two further pivotal works by Reynolds and Weiss demonstrated the existence of a neural stem cell line within the CNS with self-renewal properties.[49,67] The possibility that neurons can regenerate in the CNS took hold, especially following the discovery of the endless potential of stem cells, including the capability to differentiate into neurons and glial cells.[56,61] This ushered into “Cell-Based” neuroregeneration.
A cell-based regenerative approach involves the transplantation of specific stem cell lines into the injured CNS tissue, followed by integration and possible differentiation into actual neurons or glia: this process would promote tissue restoration directly and indirectly by stemming inhibitory influences. Bone marrow-derived MSCs are ideal candidates in this regard.[1,13,16,20,23,29,30,41,45,52,56,61,65,68] To standardize the methods of isolation, culture, expansion, and characterization of MSCs, the International Society for Cellular Therapy established minimal criteria for defining multipotent mesenchymal stromal cells in 2006.[16] MSCs are endowed with wide availability, minimal immunogenicity, lack of alloreactivity, easiness of isolation and culture, immunosuppressive and angiogenetic local effects, lack of ethical concerns and tissue-specific differentiation,[9,19,25,28,36,48,56,61,64] including neurons and glial cells.[15,40,56,61] Moreover, they can be found in several tissues.[1,8,40,54,62,69]
In the present study, we found that even the ovine xenogeneic bone marrow harvested MSCs proved to be extremely effective in promoting significant behavioral recovery. The red fluorescence emission found at the injury sites confirms the capacity of these cells to engraft within the damaged host neural tissue. Of relevance, we also found at the injury site immunopositivity for neural progenitors as Nestin, NG2 β-III tubulin, NSE, vimentin, and NF-01, undoubtedly an expression of an early “neurolike” and “glia-like” differentiation pattern.
Three important mechanisms seem to be responsible for the low immunogenicity of MSCs – lack of MHC-II and co-stimulatory molecule expression, modulation of dendritic cells, as well direct antagonization of NK, CD8+, and CD4+ T cell function, and induction of a local immunosuppressive microenvironment through the production of prostaglandins and interleukin-10.[53] These mechanisms are responsible for a very low or lacking immunogenic cell-mediated response against the transplanted xenogeneic MSCs, and of their intrinsic capacity of survival and engrafting at the target site. In transplanted rats, we found a significant behavioral improvement occurring within the 14th day after injury. It implies that not only the transplanted MSCs were able to survive and engraft at the injury site, but they also played a key role in an initial functional neuroregenerative process underlying the behavioral improvement.
Our findings suggest that MSCs, including xenogeneic ovine MSCs, favorably alter post-SCI neural tissue disruption. Ovine xenogeneic MSCs are known to migrate across the blood–brain barrier, reconstitute the neuroglial pool, and differentiate into neurons-like and microglia-like cells.[56,61] Apparently, MSCs affect the posttraumatic cord microenvironment through the secretion of a set of bioactive molecules, acting both paracrinely and autocrinely, to suppress local immune response, enhance angiogenesis, and inhibit scarring and cell death.[9,29,40,53] In the present study, no neuroglial differentiation of the ovine MSCs in vitro was induced to test the induction role on neuroglial differentiation played by the host environment.
In our study, we have selected a complete transection model rather than a contusion (incomplete) model of SCI. This choice is justified by the willingness to better assess the actual neuroregenerative potential of xenogeneic ovine bone marrow-derived MSCs, together with their capacity to survive, engraft, and form “de novo” neuron-like or glial-like cells, thereby leading to neural tissue reconstitution.
Our approach is best conceptualized within the boundaries of the GEMINI spinal cord fusion protocol.[5,6,7] A sharp, micrometric transection is entirely different from a contusive event and is key to local therapeutic manipulation with the aim of behavioral restoration, as seen in our study. A complete section allows direct assessment of therapeutic interventions in a “clean” environment, which is not possible after contusive SCI, where massive tissue damage, hemorrhage, necrosis, and cavitation interfere with an undisturbed assessment of proregenerative approaches. Importantly, a possible transdifferentiation into neuronal and glial cytotypes is easily incorporated into GEMINI “core” mechanism, i.e. the propriospinal neuronal network that sustains motor function in mammals, including man.[5,6,7] In this context, MSC would replenish the lost neuronal pool and restart transmission in the gray matter motor network. MSC may also provide a trophic environment for this so-called trunco-reticulo-prorpiospinal pathway: in this way, the two sides of the transected cord would be reconnected by regrowing fibers from gray-matter based cells across the plane of transection.[5,6,7]
MSC were applied immediately upon transection. As such they would perfectly fit inside the transection-reapposition treatment of chronic SCI (see 6) and could be applied in this context.
In conclusion, xenogeneic ovine bone marrow MSCs proved intrinsic, capable to survive, and engraft into the injured rat spinal cord; moreover, they showed signs of transdifferentiation into a neuroglial phenotype, and most importantly to support functional recovery.
Financial support and sponsorship
Nil.
Conflicts of interest
There are no conflicts of interest.
Video available on: www.surgicalneurologyint.com
Footnotes
Contributor Information
Luzzi Sabino, Email: sabino.luzzi@gmail.com.
Crovace Alberto Maria, Email: alberto.crovace@libero.it.
Lacitignola Luca, Email: luca.lacitignola@uniba.it.
Valentini Valerio, Email: valentini.valerio@libero.it.
Francioso Edda, Email: eddagiuseppina.francioso@uniba.it.
Rossi Giacomo, Email: giacomo.rossi@unicam.it.
Invernici Gloria, Email: gloria.invernici@gmail.com.
Galzio Renato Juan, Email: r.galzio@smatteo.pv.it.
Crovace Antonio, Email: antonio.crovace@uniba.it.
REFERENCES
- 1.Assunção-Silva RC, Gomes ED, Sousa N, Silva NA, Salgado AJ. Hydrogels and Cell Based Therapies in Spinal Cord Injury Regeneration. Stem Cells Int. 2015;2015:948040. doi: 10.1155/2015/948040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Baptiste DC, Fehlings MG. Update on the treatment of spinal cord injury. Prog Brain Res. 2007;161:217–33. doi: 10.1016/S0079-6123(06)61015-7. [DOI] [PubMed] [Google Scholar]
- 3.Basso DM, Beattie MS, Bresnahan JC. A sensitive and reliable locomotor rating scale for open field testing in rats. J Neurotrauma. 1995;12:1–21. doi: 10.1089/neu.1995.12.1. [DOI] [PubMed] [Google Scholar]
- 4.Benson MD, Romero MI, Lush ME, Lu QR, Henkemeyer M, Parada LF. Ephrin-B3 is a myelin-based inhibitor of neurite outgrowth. Proc Natl Acad Sci U S A. 2005;102:10694–9. doi: 10.1073/pnas.0504021102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Canavero S, Ren X-P. The Spark of Life: Engaging the Cortico-Truncoreticulo-Propriospinal Pathway by Electrical Stimulation. CNS Neurosci Ther. 2016;22:260–1. doi: 10.1111/cns.12520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Canavero S, Ren X. Houston, GEMINI has landed: Spinal cord fusion achieved. Surg Neurol Int. 2016;7:S626–8. doi: 10.4103/2152-7806.190473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Canavero S, Ren X, Kim C-Y, Rosati E. Neurologic foundations of spinal cord fusion (GEMINI) Surgery. 2016;160:11–9. doi: 10.1016/j.surg.2016.01.027. [DOI] [PubMed] [Google Scholar]
- 8.Cao F, Feng S. Human umbilical cord mesenchymal stem cells and the treatment of spinal cord injury. Chin Med J (Engl) 2009;122:225–31. [PubMed] [Google Scholar]
- 9.Caplan AI. Adult mesenchymal stem cells for tissue engineering versus regenerative medicine. J Cell Physiol. 2007;213:341–7. doi: 10.1002/jcp.21200. [DOI] [PubMed] [Google Scholar]
- 10.Chen MS, Huber AB, van der Haar ME, Frank M, Schnell L, Spillmann AA, et al. Nogo-A is a myelin-associated neurite outgrowth inhibitor and an antigen for monoclonal antibody IN-1. Nature. 2000;403:434–9. doi: 10.1038/35000219. [DOI] [PubMed] [Google Scholar]
- 11.Coleman WP, Benzel D, Cahill DW, Ducker T, Geisler F, Green B, et al. A critical appraisal of the reporting of the National Acute Spinal Cord Injury Studies (II and III) of methylprednisolone in acute spinal cord injury. J Spinal Disord. 2000;13:185–99. doi: 10.1097/00002517-200006000-00001. [DOI] [PubMed] [Google Scholar]
- 12.Czernik M, Fidanza A, Sardi M, Galli C, Brunetti D, Malatesta D, et al. Differentiation potential and GFP labeling of sheep bone marrow-derived mesenchymal stem cells. J Cell Biochem. 2013;114:134–43. doi: 10.1002/jcb.24310. [DOI] [PubMed] [Google Scholar]
- 13.Das AK, Gopurappilly R, Parhar I. Current status and prospective application of stem cell-based therapies for spinal cord injury. Curr Stem Cell Res Ther. 2011;6:93–104. doi: 10.2174/157488811795495477. [DOI] [PubMed] [Google Scholar]
- 14.Desantis S, Accogli G, Zizza S, Mastrodonato M, Blasi A, Francioso E, et al. Ultrastructural study of cultured ovine bone marrow-derived mesenchymal stromal cells. Ann Anat. 2015;201:43–9. doi: 10.1016/j.aanat.2015.06.002. [DOI] [PubMed] [Google Scholar]
- 15.Dezawa M, Kanno H, Hoshino M, Cho H, Matsumoto N, Itokazu Y, et al. Specific induction of neuronal cells from bone marrow stromal cells and application for autologous transplantation. J Clin Invest. 2004;113:1701–10. doi: 10.1172/JCI20935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F, Krause D, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8:315–7. doi: 10.1080/14653240600855905. [DOI] [PubMed] [Google Scholar]
- 17.Eftekharpour E, Karimi-Abdolrezaee S, Fehlings MG. Current status of experimental cell replacement approaches to spinal cord injury. Neurosurg Focus. 2008;24:E19. doi: 10.3171/FOC/2008/24/3-4/E18. [DOI] [PubMed] [Google Scholar]
- 18.Faden AI, Jacobs TP, Holaday JW. Comparison of early and late naloxone treatment in experimental spinal injury. Neurology. 1982;32:677–81. doi: 10.1212/wnl.32.6.677. [DOI] [PubMed] [Google Scholar]
- 19.Friedenstein AJ, Chailakhjan RK, Lalykina KS. The development of fibroblast colonies in monolayer cultures of guinea-pig bone marrow and spleen cells. Cell Tissue Kinet. 1970;3:393–403. doi: 10.1111/j.1365-2184.1970.tb00347.x. [DOI] [PubMed] [Google Scholar]
- 20.Goldschlager T, Oehme D, Ghosh P, Zannettino A, Rosenfeld JV, Jenkin G. Current and future applications for stem cell therapies in spine surgery. Curr Stem Cell Res Ther. 2013;8:381–93. doi: 10.2174/1574888x113089990048. [DOI] [PubMed] [Google Scholar]
- 21.Hall ED, Yonkers PA, Taylor BM, Sun FF. Lack of effect of postinjury treatment with methylprednisolone or tirilazad mesylate on the increase in eicosanoid levels in the acutely injured cat spinal cord. J Neurotrauma. 1995;12:245–56. doi: 10.1089/neu.1995.12.245. [DOI] [PubMed] [Google Scholar]
- 22.Hall ED, Springer JE. Neuroprotection and acute spinal cord injury: A reappraisal. NeuroRx. 2004;1:80–100. doi: 10.1602/neurorx.1.1.80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Harrop JS, Hashimoto R, Norvell D, Raich A, Aarabi B, Grossman RG, et al. Evaluation of clinical experience using cell-based therapies in patients with spinal cord injury: A systematic review. J Neurosurg Spine. 2012;17:230–46. doi: 10.3171/2012.5.AOSPINE12115. [DOI] [PubMed] [Google Scholar]
- 24.Hata K, Fujitani M, Yasuda Y, Doya H, Saito T, Yamagishi S, et al. RGMa inhibition promotes axonal growth and recovery after spinal cord injury. J Cell Biol. 2006;173:47–58. doi: 10.1083/jcb.200508143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Holden C, Vogel G. Stem cells? Plasticity: Time for a reappraisal. Science. 2002;296:2126–9. doi: 10.1126/science.296.5576.2126. [DOI] [PubMed] [Google Scholar]
- 26.Hurlbert RJ. Methylprednisolone for acute spinal cord injury: An inappropriate standard of care. J Neurosurg. 2000;93:1–7. doi: 10.3171/spi.2000.93.1.0001. [DOI] [PubMed] [Google Scholar]
- 27.Ito Y, Sugimoto Y, Tomioka M, Kai N, Tanaka M. Does high dose methylprednisolone sodium succinate really improve neurological status in patient with acute cervical cord injury.: A prospective study about neurological recovery and early complications? Spine (Phila Pa 1976) 2009;34:2121–4. doi: 10.1097/BRS.0b013e3181b613c7. [DOI] [PubMed] [Google Scholar]
- 28.Jiang Y, Jahagirdar BN, Reinhardt RL, Schwartz RE, Keene CD, Ortiz-Gonzalez XR, et al. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature. 2002;418:41–9. doi: 10.1038/nature00870. [DOI] [PubMed] [Google Scholar]
- 29.Joyce N, Annett G, Wirthlin L, Olson S, Bauer G, Nolta JA. Mesenchymal stem cells for the treatment of neurodegenerative disease. Regen Med. 2010;5:933–46. doi: 10.2217/rme.10.72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Karimi-Abdolrezaee S, Eftekharpour E. Stem cells and spinal cord injury repair. Adv Exp Med Biol. 2012;760:53–73. doi: 10.1007/978-1-4614-4090-1_4. [DOI] [PubMed] [Google Scholar]
- 31.Koda M, Okada S, Nakayama T, Koshizuka S, Kamada T, Nishio Y, et al. Hematopoietic stem cell and marrow stromal cell for spinal cord injury in mice. Neuroreport. 2005;16:1763–7. doi: 10.1097/01.wnr.0000183329.05994.d7. [DOI] [PubMed] [Google Scholar]
- 32.Koshizuka S, Okada S, Okawa A, Koda M, Murasawa M, Hashimoto M, et al. Transplanted hematopoietic stem cells from bone marrow differentiate into neural lineage cells and promote functional recovery after spinal cord injury in mice. J Neuropathol Exp Neurol. 2004;63:64–72. doi: 10.1093/jnen/63.1.64. [DOI] [PubMed] [Google Scholar]
- 33.Louro J, Pearse DD. Stem and progenitor cell therapies: Recent progress for spinal cord injury repair. Neurol Res. 2008;30:5–16. doi: 10.1179/174313208X284070. [DOI] [PubMed] [Google Scholar]
- 34.Löw K, Culbertson M, Bradke F, Tessier-Lavigne M, Tuszynski MH. Netrin-1 is a novel myelin-associated inhibitor to axon growth. J Neurosci. 2008;28:1099–108. doi: 10.1523/JNEUROSCI.4906-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lyahyai J, Mediano DR, Ranera B, Sanz A, Remacha AR, Bolea R, et al. Isolation and characterization of ovine mesenchymal stem cells derived from peripheral blood. BMC Vet Res. 2012;8:169. doi: 10.1186/1746-6148-8-169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Makino S, Fukuda K, Miyoshi S, Konishi F, Kodama H, Pan J, et al. Cardiomyocytes can be generated from marrow stromal cells in vitro . J Clin Invest. 1999;103:697–705. doi: 10.1172/JCI5298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Matsumoto T, Tamaki T, Kawakami M, Yoshida M, Ando M, Yamada H. Early complications of high-dose methylprednisolone sodium succinate treatment in the follow-up of acute cervical spinal cord injury. Spine (Phila Pa 1976) 2001;26:426–30. doi: 10.1097/00007632-200102150-00020. [DOI] [PubMed] [Google Scholar]
- 38.McCreedy DA, Sakiyama-Elbert SE. Combination therapies in the CNS: Engineering the environment. Neurosci Lett. 2012;519:115–21. doi: 10.1016/j.neulet.2012.02.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.McKerracher L, David S, Jackson DL, Kottis V, Dunn RJ, Braun PE. Identification of myelin-associated glycoprotein as a major myelin-derived inhibitor of neurite growth. Neuron. 1994;13:805–11. doi: 10.1016/0896-6273(94)90247-x. [DOI] [PubMed] [Google Scholar]
- 40.Meirelles L da S, Nardi NB. Methodology, biology and clinical applications of mesenchymal stem cells. Front Biosci. 2009;14:4281–98. doi: 10.2741/3528. [DOI] [PubMed] [Google Scholar]
- 41.Miller RH, Bai L. Translating stem cell therapies to the clinic. Neurosci Lett. 2012;519:87–92. doi: 10.1016/j.neulet.2012.01.043. [DOI] [PubMed] [Google Scholar]
- 42.Moreau-Fauvarque C, Kumanogoh A, Camand E, Jaillard C, Barbin G, Boquet I, et al. The transmembrane semaphorin Sema4D/CD100, an inhibitor of axonal growth, is expressed on oligodendrocytes and upregulated after CNS lesion. J Neurosci. 2003;23:9229–39. doi: 10.1523/JNEUROSCI.23-27-09229.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Mukhopadhyay G, Doherty P, Walsh FS, Crocker PR, Filbin MT. A novel role for myelin-associated glycoprotein as an inhibitor of axonal regeneration. Neuron. 1994;13:757–67. doi: 10.1016/0896-6273(94)90042-6. [DOI] [PubMed] [Google Scholar]
- 44.Nandoe Tewarie RS, Hurtado A, Bartels RH, Grotenhuis A, Oudega M. Stem cell-based therapies for spinal cord injury. J Spinal Cord Med. 2009;32:105–14. doi: 10.1080/10790268.2009.11760761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Noble M, Mayer-Pröschel M, Davies JE, Davies SJA, Pröschel C. Cell therapies for the central nervous system: How do we identify the best candidates? Curr Opin Neurol. 2011;24:570–6. doi: 10.1097/WCO.0b013e32834cd4c9. [DOI] [PubMed] [Google Scholar]
- 46.Obermair F-J, Schröter A, Thallmair M. Endogenous neural progenitor cells as therapeutic target after spinal cord injury. Physiology (Bethesda) 2008;23:296–304. doi: 10.1152/physiol.00017.2008. [DOI] [PubMed] [Google Scholar]
- 47.Phinney DG, Prockop DJ. Concise review: Mesenchymal stem/multipotent stromal cells: The state of transdifferentiation and modes of tissue repair--current views. Stem Cells. 2007;25:2896–902. doi: 10.1634/stemcells.2007-0637. [DOI] [PubMed] [Google Scholar]
- 48.Prockop DJ. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science. 1997;276:71–4. doi: 10.1126/science.276.5309.71. [DOI] [PubMed] [Google Scholar]
- 49.Reynolds BA, Weiss S. Generation of neurons and astrocytes from isolated cells of the adult mammalian central nervous system. Science. 1992;255:1707–10. doi: 10.1126/science.1553558. [DOI] [PubMed] [Google Scholar]
- 50.Richardson PM, McGuinness UM, Aguayo AJ. Axons from CNS neurons regenerate into PNS grafts. Nature. 1980;284:264–5. doi: 10.1038/284264a0. [DOI] [PubMed] [Google Scholar]
- 51.Rowland JW, Hawryluk GWJ, Kwon B, Fehlings MG. Current status of acute spinal cord injury pathophysiology and emerging therapies: Promise on the horizon. Neurosurg Focus. 2008;25:E2. doi: 10.3171/FOC.2008.25.11.E2. [DOI] [PubMed] [Google Scholar]
- 52.Ruff CA, Wilcox JT, Fehlings MG. Cell-based transplantation strategies to promote plasticity following spinal cord injury. Exp Neurol. 2012;235:78–90. doi: 10.1016/j.expneurol.2011.02.010. [DOI] [PubMed] [Google Scholar]
- 53.Ryan JM, Barry F, Murphy JM, Mahon BP. Interferon-gamma does not break, but promotes the immunosuppressive capacity of adult human mesenchymal stem cells. Clin Exp Immunol. 2007;149:353–63. doi: 10.1111/j.1365-2249.2007.03422.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Sakaguchi Y, Sekiya I, Yagishita K, Muneta T. Comparison of human stem cells derived from various mesenchymal tissues: Superiority of synovium as a cell source. Arthritis Rheum. 2005;52:2521–9. doi: 10.1002/art.21212. [DOI] [PubMed] [Google Scholar]
- 55.Sakiyama-Elbert SE, Panitch A, Hubbell JA. Development of growth factor fusion proteins for cell-triggered drug delivery. FASEB J. 2001;15:1300–2. doi: 10.1096/fj.00-0564fje. [DOI] [PubMed] [Google Scholar]
- 56.Sanchez-Ramos J, Song S, Cardozo-Pelaez F, Hazzi C, Stedeford T, Willing A, et al. Adult bone marrow stromal cells differentiate into neural cells in vitro . Exp Neurol. 2000;164:247–56. doi: 10.1006/exnr.2000.7389. [DOI] [PubMed] [Google Scholar]
- 57.Sekhon LH, Fehlings MG. Epidemiology, demographics, and pathophysiology of acute spinal cord injury. Spine (Phila 1976) 2001;26:S2–12. doi: 10.1097/00007632-200112151-00002. [DOI] [PubMed] [Google Scholar]
- 58.Short DJ, El Masry WS, Jones PW. High dose methylprednisolone in the management of acute spinal cord injury-a systematic review from a clinical perspective. Spinal Cord. 2000;38:273–86. doi: 10.1038/sj.sc.3100986. [DOI] [PubMed] [Google Scholar]
- 59.Silva NA, Sousa N, Reis RL, Salgado AJ. From basics to clinical: A comprehensive review on spinal cord injury. Prog Neurobiol. 2014;114:25–57. doi: 10.1016/j.pneurobio.2013.11.002. [DOI] [PubMed] [Google Scholar]
- 60.Silver J, Miller JH. Regeneration beyond the glial scar. Nat Rev Neurosci. 2004;5:146–56. doi: 10.1038/nrn1326. [DOI] [PubMed] [Google Scholar]
- 61.Simard AR, Rivest S. Bone marrow stem cells have the ability to populate the entire central nervous system into fully differentiated parenchymal microglia. FASEB J. 2004;18:998–1000. doi: 10.1096/fj.04-1517fje. [DOI] [PubMed] [Google Scholar]
- 62.Stenderup K, Justesen J, Clausen C, Kassem M. Aging is associated with decreased maximal life span and accelerated senescence of bone marrow stromal cells. Bone. 2003;33:919–26. doi: 10.1016/j.bone.2003.07.005. [DOI] [PubMed] [Google Scholar]
- 63.Stocum DL. Regenerative biology and medicine. J Musculoskelet Neuronal Interact. 2002;2:270–3. [PubMed] [Google Scholar]
- 64.Takayama T, Kondo T, Kobayashi M, Ohta K, Ishibashi Y, Kanemaru T, et al. Characteristic morphology and distribution of bone marrow derived cells in the cornea. Anat Rec (Hoboken) 2009;292:756–63. doi: 10.1002/ar.20851. [DOI] [PubMed] [Google Scholar]
- 65.Vawda R, Fehlings MG. Mesenchymal cells in the treatment of spinal cord injury: Current and future perspectives. Curr Stem Cell Res Ther. 2013;8:25–38. doi: 10.2174/1574888x11308010005. [DOI] [PubMed] [Google Scholar]
- 66.Wang KC, Koprivica V, Kim JA, Sivasankaran R, Guo Y, Neve RL, et al. Oligodendrocyte-myelin glycoprotein is a Nogo receptor ligand that inhibits neurite outgrowth. Nature. 2002;417:941–4. doi: 10.1038/nature00867. [DOI] [PubMed] [Google Scholar]
- 67.Weiss S, Dunne C, Hewson J, Wohl C, Wheatley M, Peterson AC, et al. Multipotent CNS stem cells are present in the adult mammalian spinal cord and ventricular neuroaxis. J Neurosci. 1996;16:7599–609. doi: 10.1523/JNEUROSCI.16-23-07599.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Wyatt LA, Keirstead HS. Stem cell-based treatments for spinal cord injury. Prog Brain Res. 2012;201:233–52. doi: 10.1016/B978-0-444-59544-7.00012-3. [DOI] [PubMed] [Google Scholar]
- 69.Zuk PA, Zhu M, Ashjian P, De Ugarte DA, Huang JI, Mizuno H, et al. Human adipose tissue is a source of multipotent stem cells. Mol Biol Cell. 2002;13:4279–95. doi: 10.1091/mbc.E02-02-0105. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
STROBE Statement—Checklist of items that should be included in reports of case-control studies