

Abstract
Mesenchymal stem cells (MSC) exert immune modulatory properties and previous studies demonstrated suppressive effects of MSC treatment in animal models of allergic airway inflammation. However, the underlying mechanisms have not been fully elucidated. We studied the role of MSC in immune activation and subsequent recruitment of monocytes in suppressing airway hyperresponsiveness and airway inflammation using a mouse model of allergic airway inflammation. MSC administration prior to or after allergen challenge inhibited the development of airway inflammation in allergen-sensitized mice. This was accompanied by an influx of CCR2-positive monocytes, which were localized around injected MSC in the lungs. Notably, IL-10-producing monocytes and/or macrophages were also increased in the lungs. Systemic administration of liposomal clodronate or a CCR2 antagonist significantly prevented the suppressive effects of MSC. Activation of MSC by IFN-γ leading to the upregulation of CCL2 expression was essential for the suppressive effects, as administration of wild-type (WT) MSC into IFN-γ–deficient recipients, or IFN-γ receptor- or CCL2-deficient MSC into WT mice failed to suppress airway inflammation. These results suggest that MSC activation by IFN-γ, followed by increased expression of CCL2 and recruitment of monocytes to the lungs, is essential for suppression by MSC in allergen-induced airway hyperresponsiveness and airway inflammation.
Keywords: asthma, immune, chemokine, leukocyte
INTRODUCTION
Asthma is the most common chronic respiratory disease and the prevalence is increasing in most areas of the world [1]. Many severe asthma patients are resistant to available treatments regardless of significant advances in care [2]. This has prompted the search for newer and more creative ways to manage allergic airway disease and asthma, including treatment with mesenchymal stem cells (MSC).
MSC are known to exert potent immune modulatory properties, both in vitro and in vivo [3–6]. The in vivo effects of these immune modulatory properties of MSC have been demonstrated in a number of different disease models, including inflammatory bowel disease, arthritis, allergic encephalitis, and respiratory diseases including asthma [4, 5, 7, 8]. In addition, MSC have also been observed clinically to suppress immune responses in patients with severe graft-versus-host disease and in patients with inflammatory bowel disease [9, 10]. Thus, immunomodulation with MSC has been viewed as a potential alternative or complement to conventional immunosuppressive therapy.
A number of mechanisms have been advanced to explain the immunomodulatory activities of MSC. These include production of immunoregulatory cytokines such as TGF-β or IL-10-, release of biochemical mediators (e.g., PGE2), and release of extracellular vesicles that contain miRNA or mRNA [3–6, 11]. Activities of MSC on immune cells such as induction of T regulatory cells (Treg) and monocytes and/or macrophages have also been described [4, 6].
Based on these findings, numerous studies following MSC treatment in rodent models of asthma have been reported [7, 12–16]. Recently, we demonstrated the ameliorative effects of MSC treatment in a large animal (feline) model of asthma [17, 18]. When MSC were injected intravenously, most of the injected cells initially lodged in lung capillaries via filtration effects [6]. The interaction of administered MSC with lung monocytes and/or macrophages as regulators of inflammatory responses in lung disease models, including asthma, has been previously investigated [19–23].
In the present study, we used an experimental model of allergen-induced airway inflammation to investigate changes in monocyte and macrophage populations following interaction with systemically administered MSC. We determined the mechanism underlying activation of MSC to initiate monocyte recruitment through chemokine (C–C motif) legend 2 (CCL2), which is a potent monocyte chemoattractant and shown to be secreted by MSC [5, 11]. These studies revealed previously unreported pathways of interaction between MSC activation by IFN-γ, MSC production of CCL2, and recruitment of IL-10-expressing monocytes and/or macrophages as negative regulators of airway inflammation.
MATERIALS AND METHODS
Animals
Adult C57Bl/6 (wild type, WT) mice (female, age 8–12 weeks) were purchased from Jackson Laboratory (Bar Harbor, ME) and were housed in micro-isolator cages in the animal facilities at National Jewish Health and Colorado State University. Mice on the C57Bl/6 background lacking a functional IFN-γ gene (IFN-γ−/−) or IFN-γ receptor gene (IFN-γR−/−) were purchased from Jackson Laboratory as were mice lacking the CCL2 gene (CCL2−/−). CCR2-GFP reporter mice (on the C57Bl/6 background) were provided by Dr. Eric Pamer (Memorial Sloan Kettering, NY). Foxp3-GFP reporter mice (C57Bl/6 background) were provided by Dr. A. Rudensky (Washington University, St. Louis, MO).
Animal model of asthma protocol
All animal protocols used in these studies were approved by the Institutional Animal Care and Use Committees at National Jewish Health and Colorado State University. Airway inflammation was induced in mice using ovalbumin (OVA) as previously described [24]. Briefly, mice were immunized twice, 14 days apart via intraperitoneal (i.p.) injection with 20 μg OVA with 1.0 mg of alum (AlumImuject: Pierce, Rockford, IL) as an adjuvant. Fourteen days later, mice were challenged with 20 minutes of inhalation of OVA (1% in saline) for 3 consecutive days by ultrasonic nebulizer (model NE-U07, Omron Healthcare, Vernon Hills, IL). Forty-eight hours after the last OVA nebulization, the mice were anesthetized and airway hyperresponsiveness (AHR) following methacholine challenge was determined in anesthetized, tracheostomized, mechanically ventilated mice. The mice were then euthanized and bronchoalveolar lavage (BAL) fluid with 1 ml of HBSS and lung tissues for histology were collected.
Generation of adipose tissue-derived MSC and treatment protocol
Adipose tissue-derived MSC were generated from abdominal adipose tissues. Adipose tissue-derived MSC were chosen for this study as we previously demonstrated that these cells were easier to propagate in vitro and shared a similar phenotype with bone marrow-derived MSC [25]. Adipose tissue-derived MSC were generated from abdominal adipose tissues collected from WT or CCL2−/− mice. To prepare MSC cultures, adipose tissues were minced under sterile conditions and incubated in a 1 mg/ml solution of collagenase (Sigma-Aldrich, St. Louis, MO) at 37°C for 30 minutes to generate single cell suspensions in PBS. The cell suspension was then centrifuged at 1,050 x G to pellet the stromal vascular fraction, rinsed twice, and resuspended in MSC culture medium. Culture medium consisted of low glucose DMEM (InVitrogen/Gibco, Carlsbad, CA) supplemented with essential and non-essential amino acids (InVitrogen/Gibco) and 10% heat-inactivated fetal bovine serum (FBS) (Cell Generation, Fort Collins, CO), plus penicillin and streptomycin solution (InVitrogen/Gibco). Cells in medium were allowed to adhere in tissue culture flasks (BD Falcon, Bedford, MA) for 72 hours, after which the non-adherent cells were removed and fresh medium was added. When cells reached 70–80% confluence, they were passaged using trypsin-EDTA solution (InVitrogen/Gibco). All cells for in vivo experiments were used at passage 2–3. The cell surface phenotype of MSC was determined by flow cytometry and the ability of the MSC to undergo tri-lineage differentiation was assessed as described previously [17]. MSC (2x105) were administered intravenously (i.v.) 24 hours prior to the first OVA challenge or 2 hours after the last OVA challenge in sensitized mice.
Histological studies
After BAL fluid was collected, lungs were inflated with 1 ml 10% formalin through the trachea and fixed in formalin by immersion. Blocks of lung tissue were cut from around the main bronchus and embedded in paraffin; 2–3 tissue sections (4 μm) per mouse were then affixed to microscope slides and deparaffinized. The slides were then stained with hematoxylin and eosin (H/E) or periodic acid-Schiff (PAS). The pictures of slides were taken with a microscope (BX40; Olympus America Inc., Melville, NY) equipped with a digital camera (Q-color 3; Olympus America Inc.). Goblet cell metaplasia was quantified from PAS+ areas along the airway epithelium using NIH ImageJ software (version 1.38), available on the Internet at http://rsb.info.nih.gov/ij/download.html. Four different fields per slide in 5 samples from each group of mice were examined in a blinded manner.
Biochemicals
OVA was purchased from Thermo-Fisher Scientific (Hampton, NH). Liposomal clodronate was administered by i.v. injection 24 hours before and after MSC administration. Macrophage depletion activity was assessed using in vitro assays prior to in vivo experiments. To track injected MSC in the lung tissue, MSC were labeled with Cell Tracker Orange CMTMR dye (Thermo-Fisher Scientific). The small molecule CCR2 antagonist RS102895 was purchased from Tocris Bioscience (Bristol, United Kingdom) and administered (5 mg/kg BW) by i.p. injection 24 hours prior to the first OVA challenge and 2 hours prior to each OVA challenge.
Tissue processing and flow cytometry
Lung tissue was collected, placed in tissue culture medium on ice and then minced using scissors. Tissue was then digested in a 1% solution of collagenase (Sigma-Aldrich) in MEM medium with 2% FBS and DNAse for 30 minutes at 37°C, erythrocytes lysed with ACK solution, and single cell suspensions were filtered through cell strainers and washed twice with FACs buffer (PBS plus 2% FBS and 0.05% sodium azide). Cells were incubated with antibodies in FACs buffer in 96-well round bottom plates for 30 minutes at room temperature, then washed and incubated with streptavidin conjugates when necessary.
Intracellular cytokine staining was performed to identify IL-10-producing monocytes and macrophages. For these experiments, OVA sensitized and challenged mice received MSC treatment after the last OVA challenge. Lung tissues were removed 7 hours after MSC treatment. Lung cells were isolated as described above and the cells were cultured with 10 μg/mL of LPS and brefeldin A followed by surface staining [19]. Cells were then fixed and permeabilized prior to intracellular IL-10- staining with anti-mouse IL-10- (allophycocyanin; APC, clone JES5-16E3) or rat IgG2b K APC as an isotype control following surface staining with anti-CD11b (FITC, clone M1/70) and anti-F4/80(PE-Cyanine5, clone BM8). All of the antibodies were purchased from eBioscience (San Diego, CA). Directly-labeled antibodies for flow cytometry and/or immunofluorescence staining were purchased from either eBiosciences or Becton-Dickinson (Bedford, MA) except as noted, and included the following clones: mouse Ly6G (clone 1A8), mouse B220 (clone RA3-6B2), mouse CD20 (clone eBioL31), mouse CCR2 (clone 475301, R&D Systems, Minneapolis, MN), mouse CD4 (clone RM4-5), mouse CD8 (clone 53.6.7), and mouse NK1.1 (clone PK 136). Antibodies used for characterization of mouse MSC included mouse CD44 (clone 1M7), mouse CD73 (clone TY/11.8), mouse CD34 (clone RAM34), mouse CD90 (clone HI551), mouse CD105 (clone 8A1), and mouse CD45 (clone 30-F11).
Flow cytometry was carried out using a Beckman Coulter Cyan ADP flow cytometer running Summit software (Fort Collins, CO) or LSRFortessa (Becton-Dickinson). Analysis was performed using FlowJo software (Ashland, OR). Cell counts were performed with an automated cell counter (Nexcelom Bioscience, Lawrence, MA). Confocal microscopy was performed using an Olympus IX83 instrument, with CellSens software.
Immunohistochemistry
Lung tissues were snap-frozen in OCT solution (VWR Scientific, Radnor, PA), and then crysosectioned to 4mm thickness onto Superfrost slides (VWR Scientific). Tissues were fixed in acetone, non-specific peroxidase activity blocked, then incubated with primary antibodies diluted in PBS with 2% normal serum overnight at 4°C, rinsed and incubated with biotinylated secondary antibody, followed by incubation with streptavidin-HRP (Vector Laboratories, Velencia, CA), then incubated with amino-ethyl carbazole substrate followed by hematoxylin counterstain. Sections were sealed using cryomount (PolyScientific, Bay Shore, NY) and examined using a Zeiss microscope.
For immunofluorescence evaluation of MSC and monocyte trafficking to lungs, immediately following dissection lung tissues were immersion fixed in 1% paraformaldehyde-lysine-periodate fixative (1% paraformaldehyde in 0.2 M lysine-HCL, 0.1 M anhydrous dibasic sodium phosphate, with 0.21% sodium periodate) for 24 hours at 4°C. Following fixation, lungs were placed in a 30% w/v sucrose solution for 24 hours at 4°C, prior to embedding and freezing in Tissue Tek OCT compound. Embedded tissues were sectioned at 5 μm for immunostaining. Tissue sections were immunostained with a rabbit antibody against GFP (Life Technologies, Carlsbad, CA) to enhance GFP signal intensity, followed by incubation with a donkey anti-mouse IgG antibody, conjugated to AF647 (Jackson ImmunoResearch, West Grove, PA), and then pseudo-colored green after image analysis.
Cytokine assays
Cytokine concentrations in BAL fluid and in supernatants from MSC cultures were assayed using commercial ELISA kits and performed according to manufacturers’ directions. ELISA kits for IL-4, IL-5, IL-10-, IL-13, and IFN-γ were purchased from eBioscience and the kit for CCL2 was purchased from R&D Systems (Minneapolis, MN). Recombinant murine IFN-γ was obtained from Peprotech (Rocky Hill, NJ).
Statistical analyses
Statistical comparisons between the data sets with two treatment groups were performed using nonparametric t-tests (Mann-Whitney test). Comparisons between 3 or more groups were performed using ANOVA, followed by Tukey multiple means post-test. Analyses were performed using Prism5 software (GraphPad, La Jolla, CA). For all analyses, statistical significance was determined for p<0.05. All results were expressed as the mean±standard error of mean from at least 2 independent experiments.
RESULTS
MSC suppress airway inflammation following i.v. injection
MSC were administered by i.v. injection prior to allergen challenge or after completion of the 3 days of inhaled allergen challenge in allergen sensitized mice. We observed that i.v. administration of MSC either prior to or following allergen challenge significantly suppressed development of AHR and airway eosinophilia in allergen sensitized and challenged C57Bl/6 (WT) mice, compared to sensitized and challenged mice not injected with MSC (Figures 1A and 1B). Reduction of eosinophil numbers in BALF was more prominent in mice that received MSC prior to allergen challenge compared with mice that received MSC following allergen challenge. However, following both treatments the suppressive effects on AHR, airway inflammation and goblet cell metaplasia were comparable (Figures 1A, 1C and 1D). In addition, the administration of MSC prior to allergen challenge reduced BAL fluid levels of IL-4, IL-5, and IL-13 in sensitized mice (Figure 1E). Further, IL-10- levels in BAL fluid were increased following MSC administration compared with vehicle-injected mice (Figure 1E).
Figure 1. Effects of MSC Injection on Allergen-Induced AHR and Inflammation.

C57Bl/6 mice (n=8 per group) were sensitized to OVA and then exposed to aerosol challenge with OVA, as described in Methods. In (A), the effects of a single i.v. injection of 2x105 adipose-derived MSC immediately prior to first aerosolized OVA challenge on AHR were assessed in allergen sensitized mice (OVA
 MSC/OVA) and compared to AHR in OVA sensitized mice injected with PBS (OVA
PBS/OVA) and to AHR in sham sensitized mice (PBS/OVA). In some group of mice, MSC or PBS was injected following allergen challenge in sensitized mice (OVA/OVA
MSC and OVA/OVA
PBS respectively). Injection of MSC prior to or following allergen challenge significantly reduced AHR in OVA sensitized and challenged mice (*p<0.05 versus OVA/OVA+PBS and +p<0.05 versus PBS/OVA). In (B), effects of MSC injection on numbers of eosinophils in BAL fluids were assessed. Mice injected with MSC had significantly fewer eosinophils in their airways than PBS-injected mice (*p<0.05). In (C), representative photomicrographs of cellular responses in the lung tissue sections of sham-sensitized but OVA challenged mice (a and f), OVA sensitized and challenged mice received vehicle (b and g) or MSC injection (c and h) prior to OVA challenge, and vehicle (d and i) or MSC injection (e and j) following the last OVA challenge were assessed using routine histology. Panels a, b, c, d and e: H&E staining; panels f, g, h, i, and j: PAS staining. In (D), PAS+ areas were quantified and analyzed. PAS+ areas were significantly decreased in mice that received MSC (n=5 per group) (*p<0.05). In (E), effects of MSC injection on BAL concentrations of IL-4, IL-5, and IL-13 were assessed. Concentrations of IL-4, IL-5, and IL-13 were significantly decreased in MSC-treated mice compared to untreated sensitized mice. Conversely, concentrations of IL-10- were significantly increased (*p<0.05).
MSC/OVA) and compared to AHR in OVA sensitized mice injected with PBS (OVA
PBS/OVA) and to AHR in sham sensitized mice (PBS/OVA). In some group of mice, MSC or PBS was injected following allergen challenge in sensitized mice (OVA/OVA
MSC and OVA/OVA
PBS respectively). Injection of MSC prior to or following allergen challenge significantly reduced AHR in OVA sensitized and challenged mice (*p<0.05 versus OVA/OVA+PBS and +p<0.05 versus PBS/OVA). In (B), effects of MSC injection on numbers of eosinophils in BAL fluids were assessed. Mice injected with MSC had significantly fewer eosinophils in their airways than PBS-injected mice (*p<0.05). In (C), representative photomicrographs of cellular responses in the lung tissue sections of sham-sensitized but OVA challenged mice (a and f), OVA sensitized and challenged mice received vehicle (b and g) or MSC injection (c and h) prior to OVA challenge, and vehicle (d and i) or MSC injection (e and j) following the last OVA challenge were assessed using routine histology. Panels a, b, c, d and e: H&E staining; panels f, g, h, i, and j: PAS staining. In (D), PAS+ areas were quantified and analyzed. PAS+ areas were significantly decreased in mice that received MSC (n=5 per group) (*p<0.05). In (E), effects of MSC injection on BAL concentrations of IL-4, IL-5, and IL-13 were assessed. Concentrations of IL-4, IL-5, and IL-13 were significantly decreased in MSC-treated mice compared to untreated sensitized mice. Conversely, concentrations of IL-10- were significantly increased (*p<0.05).
Impact of MSC administration on recruitment of monocytes to the lungs
Monocytes and/or macrophages play a critical role in MSC-induced anti-inflammatory activities [19]. CCR2 is mainly expressed on monocytes and has been recognized as a key molecule in the recruitment of monocytes to inflammatory sites as well as differentiation into macrophages [26]. We determined in this experimental model whether MSC could recruit monocytes to the lungs through activation of CCR2. To address this question, experiments were performed using labeled MSC to assess cell localization in the lungs following i.v. injection and to determine the impact of MSC injection on monocyte recruitment to the lungs using the CCR2-GFP reporter mice.
Labeled MSC were widely demonstrated throughout the lung parenchyma following i.v. administration (Supplemental Figure 1A) and remained abundant in lung tissue for at least 72 hours after injection; cells were still detected in the lungs up to 7 days after injection (data not shown). In addition, we observed that CCR2+ monocytes were rapidly (within 8 hours of MSC injection) recruited to the lungs following MSC injection and remained in the lungs for at least 7 days after MSC injection (Supplemental Figure 1B). Notably, extensive clustering of CCR2+ monocytes around injected MSC was observed 24 hours after MSC administration in the lung tissue (Figure 2). The association of CCR2+ monocytes with injected MSC persisted through later time points as well (72 hours, 7 days), although the clustering was not as tight as at 24 hours. Thus, these studies revealed a rapid and persistent influx of monocytes into the lungs in response to injection of MSC.
Figure 2. Pulmonary Monocyte Response to MSC Administration.

CCR2-GFP reporter mice (n=3 per group) were administered 2x105 MSC labeled with Cell Tracker Orange by the i.v. route, and pulmonary CCR2+ monocyte responses were assessed at 0 hours (panel a), 8 hours (panel b), 24 hours (panel c), and 72 hours (panel d) of MSC injection using confocal microscopy. CCR2-GFP+ monocytes were found to cluster around injected Cell Tracker Orange positive-MSC, particularly at early time points after MSC injection.
Flow cytometry was also performed to assess the effects of MSC injection on monocyte and macrophage populations in the lungs. We observed a significant increase in CD11b+F4/80+ monocytes and/or macrophages in lung tissues at 7 hours after MSC treatment (Figure 3A). In addition, numbers of CCR2+ monocytes, which may include macrophages, were increased following MSC treatment. Further, IL-10-producing monocytes and/or macrophages were significantly increased in lung tissue following MSC injection compared with vehicle injection. Immunohistochemical analyses of MSC-injected lung tissues also revealed an increase in numbers of monocytes and/or macrophages stained with anti-CD68 (a marker available for these cells in immunohistochemistry) in MSC-injected animals. Many of the cells were oriented around small airways (Figure 3B). There was also a significant increase in the numbers of CD4+Foxp3+-Tregs in the lungs of MSC injected mice (Supplemental Figure 2), consistent with observations from previous reports [9]. In contrast, MSC injection did not produce a significant change in the numbers of CD4+ or CD8+ T cells, neutrophils (CD11b+Ly6G+), B cells, or NK cells in the lungs at any time points examined (data not shown).
Figure 3. Monocyte and/or Macrophage Responses Following MSC Administration in the Lungs of Allergen Sensitized and Challenged Mice.

C57Bl/6 mice were sensitized and challenged with OVA, then injected i.v. with 2x105 MSC. (A) Seven hours after MSC injection, lung cells from MSC-injected (MSC) or PBS-injected (vehicle) mice were isolated, then immunostained for identification of CD11b+F4/80+ (per total lung mononuclear cells), CCR2+CD11b+F4/80+ (per total CD11b+F4/80+ cells), or IL-10+CD11b+F4/80+ monocytes and macrophages (per total CD11b+F4/80+ cells) by flow cytometry (n=6 per group) (*p<0.05). (B) Gating strategy to detect IL-10-positive monocyte/macrophage in the lung tissue and a representative flow cytometry data. IL-10- levels in CD11b+F4/80 cells were compared following with or without LPS stimulation as describe in Materials and Methods. In (C), lung tissues from PBS-injected (left panel) and MSC-injected (right panel) mice were immunostained for identification of CD68+ monocytes and macrophages. Lungs from MSC-injected mice had prominent accumulations of CD68+ cells (red stain) around airways, compared to PBS-injected mice. Similar results were observed in 2 additional control and treated mice, processed similarly.
Monocyte depletion eliminates MSC-induced suppression of airway inflammation
To determine whether monocytes were recruited to the airways in response to MSC administration and mediated the suppressive effects of MSC on airway inflammation, liposome clodronate was systemically administered to deplete circulating monocytes in allergen sensitized and challenged WT mice. As shown in Figure 4, treatment with liposome clodronate abrogated the anti-inflammatory activities of MSC, preventing MSC-induced reductions in AHR and suppression of airway eosinophilia and goblet cell metaplasia along the airways. Liposome clodronate treatment did not affect the viability of MSC (Supplemental Figure 3).
Figure 4. Effects of Monocyte Depletion and Migration Blockade on Allergen-Induced AHR and Airway Inflammation.

OVA sensitized and challenged mice (n=8 per group) received a single i.v. injection of MSC. Monocytes were concurrently depleted in one group of mice by i.v. administration of liposomal clodronate (OVA/OVA+MSC+Lip Clod) or vehicle alone (OVA/OVA+MSC+Vehicle). Liposomal clodronate was injected by i.p. 24 hours prior to and following MSC injection. The effects of liposomal clodronate administration on MSC-induced suppression of (A) AHR, (B) eosinophil numbers in BAL fluid, and (C) goblet cell metaplasia along the airways were assessed (*p<0.05).
Role of the CCL2-CCR2 axis in MSC-induced anti-inflammatory effects
To further explore the mechanisms of monocytes recruited in MSC-mediated suppression of inflammation, the CCR2 antagonist (RS102895) was administered to mice injected with MSC. In allergen sensitized and challenged WT mice treated with RS102895, the effects of MSC administration on AHR and airway inflammation were almost completely eliminated (Figures 5A, 5B, and 5C).
Figure 5. The Role of CCL2-CCR2 Axis for MSC-Induced Suppression of Allergen-Induced AHR and Airway Inflammation.

OVA sensitized and challenged mice (n=8 per group) received MSC treated with CCR2 antagonist RS102895 (OVA/OVA+MSC+RS102895) or vehicle alone (OVA/OVA+MSC+Vehicle). (A) AHR and (B) eosinophil numbers in BAL fluid were assessed (*p<0.05). To identify the source of CCL2, MSC were derived from CCL2−/− mice (CCL2−/− MSC) and the effects of administration into WT mice (OVA/OVA+CCL2−/− MSC) on (C) AHR and (D) eosinophil numbers in BAL fluid were compared to recipients of WT MSC (OVA/OVA+WT MSC) (n=8 per group) (*p<0.05).
As CCL2 is a potent ligand for CCR2, studies were performed to determine whether CCL2 production by injected MSC was necessary to suppress AHR and airway inflammation. Allergen sensitized WT recipient mice were injected with CCL2−/− or WT MSC prior to allergen challenge, and the effects on AHR and airway inflammation were assessed. Injection of CCL2−/− MSC had little effect on suppressing AHR, airway eosinophilia, or goblet cell metaplasia compared to the effects of injecting WT MSC (Figures 5D, 5E, and 5F). In addition, the numbers of monocytes and/or macrophages in the lungs decreased in mice treated with CCL2−/− MSC (data not shown).
IFN-γ is required for MSC production of CCL2 and in vivo MSC activation
IFN-γ has been reported previously to activate MSC production of several key immune regulatory enzymes and cytokines [27, 28]. Indeed, we found that when MSC were treated in vitro with IFN-γ, they produced large amounts of CCL2 (Figure 6A). We also demonstrated that IFN-γ levels were increased in the airways following allergen sensitization and challenge compared with challenge alone (Figure 6B). These results suggested that IFN-γ played an important role in the regulation of MSC responses with respect to airway inflammation in vivo. To address this question, MSC generated from IFN-γR−/− mice were injected into WT mice and their effects on airway responsiveness and inflammation were compared to those of injected WT MSC. While WT MSC significantly suppressed AHR and airway inflammation, IFN-γR−/− MSC exerted relatively little effect on AHR, airway eosinophilia, or goblet cell metaplasia in allergen sensitized and challenged WT recipient mice (Figures 6C, 6D, and 6E).
Figure 6. The Role for IFN-γ in MSC-Induced Suppression of Allergen-Induced AHR and Airway Inflammation.

In (A), cultured MSC were incubated with medium alone, or medium with 10 ng/ml recombinant mouse IFN-γ, and production levels of CCL2 in culture supernatants were determined by specific ELISA (3 replicates with 2 repeats) (*p<0.05). In (B), BAL levels of IFN-γ from mice that received sham-sensitization and OVA challenge (PBS/OVA) or OVA sensitization and challenge (OVA/OVA) were measured by high-sensitivity ELISA kit (n=10) (*p<0.05). Allergen sensitized WT mice received WT MSC (OVA/OVA+WT MSC) or MSC derived from mice lacking a functional IFN-γ receptor (OVA/OVA+IFN-γR−/− MSC) following sensitization but prior to challenge with OVA. The changes of (C) AHR and (D) eosinophil numbers in BAL fluid were assessed following MSC treatments (n=8 per group) (*p<0.05). In (E), goblet cell metaplasia along the airways were assessed in WT mice that received IFN-γ receptor deficient-MSC (OVA/OVA+IFN-γR−/− MSC) or WT MSC (OVA/OVA+WT MSC) following sensitization but prior to challenge with OVA (n=5 per group) (*p<0.05). In mice lacking a functional IFN-γ gene (IFN-γ−/− mice), the effects of treatment with WT MSC (IFNγ−/−OVA/OVA+MSC) or vehicle (IFNγ−/− OVA/OVA+Vehicle) on (F) AHR was analyzed following OVA sensitization and challenge (n=8 per group).
The suppressive activities of MSC in the development of AHR and airway inflammation were also assessed in recipient mice unable to produce IFN-γ. WT MSC were administered to IFN-γ−/− recipient mice and the suppressive activities of MSC on AHR and airway inflammation were compared with vehicle recipients. As shown in Figure 6F, WT MSC exerted little protective effect on AHR, as well as on airway eosinophilia or goblet cell metaplasia (data not shown) when administered to sensitized and challenged IFN-γ−/− recipient mice.
Both IFN-γ and CCL2 are required for MSC-dependent recruitment of IL-10- producing monocytes to the lungs
To determine the mechanisms underlying the impaired suppressive activities of administered IFN-γR−/− MSC and CCL2−/− MSC, the numbers of IL-10- producing monocytes and macrophages in the lungs of allergen sensitized and challenged mice following WT, IFN-γR−/− or CCL2−/− MSC injection were monitored by flow cytometry. As shown in Figure 7, IFN-γR−/− or CCL2−/− MSC failed to recruit IL-10- producing monocytes and macrophages into the lung.
Figure 7. Monocyte and/or Macrophage Responses Following WT, IFN-γR−/− or CCL2−/− MSC Administration in the Lungs of Allergen Sensitized and Challenged Mice.
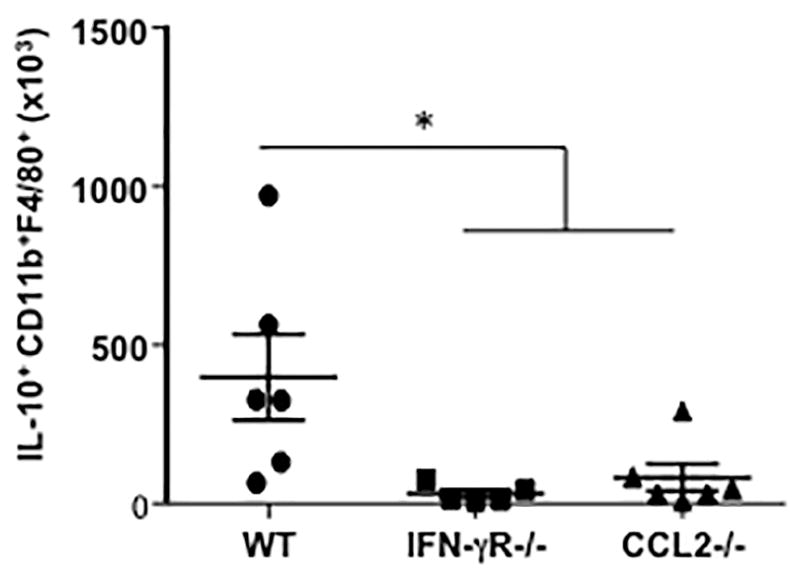
C57Bl/6 mice were sensitized and challenged with OVA, then injected i.v. with 2x105 of WT, IFN-γR−/− or CCL2−/− MSC. Seven hours after MSC injection, lung cells from MSC-injected (MSC) mice were isolated, then immunostained for identification of IL-10+CD11b+F4/80+ monocytes and macrophages (per lung) by flow cytometry (n=6 per group) (*p<0.05).
DISCUSSION
Our studies demonstrated that MSC administration following allergen sensitization but prior to allergen challenge inhibited the full complement of lung allergic responses including development of AHR, increases in airway eosinophils and Th2 type cytokine levels in BAL fluid, inflammatory cell infiltration, and goblet cell metaplasia in the airways. When MSC were administered after allergen challenge in sensitized mice, development of AHR and goblet cell metaplasia were inhibited. IL-10- levels in BAL fluid were increased following MSC treatment prior to allergen challenge in sensitized mice. In the lungs of OVA sensitized and challenged mice, Tregs as well as monocytes and/or macrophages were increased following MSC treatment. Among these cells, numbers of IL-10-producing monocytes and/or macrophages were increased. Depletion of monocytes with systemic administration of liposomal clodronate or blockade of monocyte migration with a CCR2 antagonist abrogated the suppressive effects of the MSC. These findings supported the notion that monocytes played a critical role in the effector arm of MSC suppressing lung allergic responses. The results from studies using CCL2-deficient MSC revealed that the CCL2-CCR2 axis played a crucial role in the suppressive activity of monocytes induced by administration of MSC. IFN-γ was shown to increase CCL2 release from MSC in vitro. In addition, in experiments with IFN-γ receptor-deficient MSC administered to WT mice combined with administration of WT MSC to IFN-γ deficient mice, IFN-γ was shown to be an essential initiator of the suppressive effects induced by MSC in allergen-induced airway inflammation in vivo.
Previous studies of MSC administration in animal models of asthma proposed several immune mechanisms for suppression of airway inflammation. Studies with MSC in experimental models of asthma identified suppressive effects of MSC treatment including increased levels of TGF-β and Th1 type cytokines, transfer of mitochondria to airway epithelial cells, increased numbers of Tregs, and decreased secretion of alarmins from epithelial cells [16, 19–31]. In this study, increases in numbers of Tregs in lung tissue were also demonstrated following MSC treatment. The potent roles of macrophages in MSC-induced anti-inflammatory activities have also been shown by MSC-induced reprogramming of monocytes and/or macrophages to produce IL-10- in a mouse model of sepsis [19]. Mathias et al. reported that alveolar macrophages play a role in human MSC-induced inhibition of the development of asthma in mice [20]. Treatment with human bone marrow-derived MSC was shown to inhibit development of experimental asthma in mice through induction of M2 type macrophages and TGF-β secretion [21]. In addition, MSC administration reduced allergen-induced airway inflammation and AHR by induction of TGF-β or IL-10-producing macrophages following phagocytosis of administered MSC [22]. A number of in vitro studies similarly demonstrated that MSC were capable of conditioning macrophages to produce IL-10- [20, 32]. In the present study, we also demonstrated that MSC treatment increased numbers of monocytes and/or macrophages in the lungs of allergen sensitized and challenged mice and that the cells were potent IL-10-producers. Furthermore, the recruited monocytes expressed CCR2, and the suppressive effects of MSC on allergen-induced airway inflammation and AHR were abrogated by treatment with a CCR2 antagonist or by administration of CCL2-deficient MSC. These results suggested that the recruitment of monocytes through activation of the CCL2-CCR2 axis in response to MSC administration was another critical mechanism underlying the immunomodulatory properties of MSC. Further support was gained in the studies with liposome clodronate, which also resulted in impaired MSC suppressive effects as this treatment likely depleted circulating monocytes rather than resident alveolar macrophages [33].
The results of these investigations do not rule out an important role for Tregs in suppressing AHR and airway inflammation. As monocyte depletion or inhibition of CCR2-CCL2 signaling significantly abolished the suppressive effects of MSC treatment, Treg expansion could be a secondary event following expansion of IL-10-producing monocytes and/or macrophages. Recent studies implicated the distinct roles of subsets of macrophages such as M1 or M2 in asthma [23, 34], and some studies reported that MSC treatment in allergen-induced airway inflammation resulted in induction of M2 macrophages [21, 22]. Based on current notions [35], it appears that IL-10-producing monocytes and M2 macrophages are important effector cells in MSC-induced suppression of allergen-induced airway inflammation.
We also demonstrated an essential role for IFN-γ in the MSC-mediated suppression of allergen-induced airway inflammation and AHR. Although asthma is thought to be a Th2-cytokine associated disease, up-regulation of IFN-γ has been reported in experimental asthma as well as human asthma (36–39). IFN-γ has been shown to activate MSC to secrete hepatocyte growth factor, TGF-β or indoleamine 2, 3, dioxygenase to inhibit T cell proliferation [27, 28, 40]. In this study, in vitro treatment with IFN-γ induced MSC release of CCL2, which was a critical chemoattractant for monocytes to the lung. Transfer of IFN-γ receptor-deficient MSC into WT mice or administration of WT MSC into IFN-γ-deficient mice, failed to demonstrate the suppressive effects on lung allergic responses. As described previously [41], we confirmed that the levels of IFN-γ were increased in the airways of allergen sensitized and challenged mice. Taken together, these findings implicate an important role for IFN-γ as a potent initiator of the immune suppressive effects of MSC in allergen-induced airway inflammation and AHR.
Based on our findings, a new model to explain MSC-mediated suppression of airway inflammation and AHR is proposed (Figure 8), in which MSC activated by IFN-γ secrete CCL2 and recruit monocytes to the lungs to amplify the overall immunomodulatory potential of MSC. MCS administration not only prior to allergen challenge but also after completion of allergen challenge demonstrated significant suppressive effects on AHR and airway inflammation. These findings may be relevant to the use of MSC in the treatment of asthma. For example, it may be possible to augment the effects of MSC treatment by pre-activating the MSC with IFN-γ prior to administration as in clinical trials in allotransplantation or Crohn’s disease [42, 43]. Alternatively, it may also be possible to engineer MSC to over-express chemokines such as CCL2 or CCL5 to augment recruitment of immune suppressive monocytes to the lungs. Our data also provide in vivo evidence that injection of MSC can exert a significant dampening effect on airway inflammation even after airway inflammation is established, further supporting the potential for MSC therapy in lung inflammatory diseases.
Figure 8. Proposed Model for IFN-γ Activation, CCL2 Production, and Monocyte Recruitment in MSC-Mediated Suppression of AHR and Airway Inflammation.

IFN-γ-activated MSC secrete CCL2 and recruit CCR2+ monocytes and macrophages into the inflammatory site. Recruited monocytes and macrophages are triggered to produce IL-10- and consequently resolve inflammation.
CONCLUSION
These studies indicate that MSC activated in vivo by IFN-γ suppress airway inflammation via CCL2 production and recruitment of anti-inflammatory monocytes and macrophages produces IL-10- in the lungs.
Supplementary Material
Acknowledgments
The authors wish to acknowledge the assistance of Dr. Scott Hafeman (CSU) for preparation of liposomal clodronate.
Grant Support: The studies at CSU were supported by PHS grant K08AI071724 to TW and by funding from the Shipley Foundation. Studies at National Jewish Health were supported by PHS grants HL-36577 and AI-36577 (EWG).
References
- 1.Lundbäck B, Backman H, Lötvall J, Rönmark E. Is asthma prevalence still increasing? Expert Rev Respir Med. 2016;10:39–51. doi: 10.1586/17476348.2016.1114417. [DOI] [PubMed] [Google Scholar]
- 2.Barnes PJ. New therapies for asthma: is there any progress? Trends Pharmacol Sci. 2010;31:335–343. doi: 10.1016/j.tips.2010.04.009. [DOI] [PubMed] [Google Scholar]
- 3.Wang Y, Chen X, Cao W, Shi Y. Plasticity of mesenchymal stem cells in immunomodulation: pathological and therapeutic implications. Nat Immunol. 2014;15:1009–1016. doi: 10.1038/ni.3002. [DOI] [PubMed] [Google Scholar]
- 4.Frenette PS, Pinho S, Lucas D, Scheiermann C. Mesenchymal stem cell: keystone of the hematopoietic stem cell niche and a stepping-stone for regenerative medicine. Annu Rev Immunol. 2013;31:285–316. doi: 10.1146/annurev-immunol-032712-095919. [DOI] [PubMed] [Google Scholar]
- 5.Cao W, Cao K, Cao J, Wang Y, Shi Y. Mesenchymal stem cells and adaptive immune responses. Immunol Lett. 2015;168:147–153. doi: 10.1016/j.imlet.2015.06.003. [DOI] [PubMed] [Google Scholar]
- 6.Eggenhofer E, Luk F, Dahlke MH, Hoogduijn MJ. The life and fate of mesenchymal stem cells. Front Immunol. 2014;5:148. doi: 10.3389/fimmu.2014.00148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Srour N, Thébaud B. Stem cells in animal asthma models: a systematic review. Cytotherapy. 2014;16:1629–1642. doi: 10.1016/j.jcyt.2014.08.008. [DOI] [PubMed] [Google Scholar]
- 8.Weiss DJ. Concise review: current status of stem cells and regenerative medicine in lung biology and diseases. Stem Cells. 2014;32:16–25. doi: 10.1002/stem.1506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Munir H, McGettrick HM. Mesenchymal stem cell therapy for autoimmune disease: risks and rewards. Stem Cells Dev. 2015;24:2091–2100. doi: 10.1089/scd.2015.0008. [DOI] [PubMed] [Google Scholar]
- 10.Duran NE, Hommes DW. Stem cell-based therapies in inflammatory bowel disease: promises and pitfalls. Therap Adv Gastroenterol. 2016;9:533–547. doi: 10.1177/1756283X16642190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Rani S, Ryan AE, Griffin MD, Ritter T. Mesenchymal stem cell-derived extracellular vesicles: toward cell-free therapeutic applications. Mol Ther. 2015;23:812–823. doi: 10.1038/mt.2015.44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lin HY, Xu L, Xie SS, Yu F, Hu HY, Song XL, Wang CH. Mesenchymal stem cells suppress lung inflammation and airway remodeling in chronic asthma rat model via PI3K/Akt signaling pathway. Int J Clin Exp Pathol. 2015;8:8958–8967. [PMC free article] [PubMed] [Google Scholar]
- 13.Cruz FF, Borg ZD, Goodwin M, Sokocevic D, Wagner DE, Coffey A, Antunes M, Robinson KL, Mitsialis SA, Kourembanas S, Thane K, Hoffman AM, McKenna DH, Rocco PR, Weiss DJ. Systemic administration of human bone marrow-derived mesenchymal stromal cell extracellular vesicles ameliorates aspergillus hyphal extract-induced allergic airway inflammation in immunocompetent mice. Stem Cells Transl Med. 2015;4:1302–1316. doi: 10.5966/sctm.2014-0280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zeng SL, Wang LH, Li P, Wang W, Yang J. Mesenchymal stem cells abrogate experimental asthma by altering dendritic cell function. Mol Med Rep. 2015;12:2511–2520. doi: 10.3892/mmr.2015.3706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cahill EF, Tobin LM, Carty F, Mahon BP, English K. Jagged-1 is required for the expansion of CD4+ CD25+ FoxP3+ regulatory T cells and tolerogenic dendritic cells by murine mesenchymal stromal cells. Stem Cell Res Ther. 2015;6:19. doi: 10.1186/s13287-015-0021-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Duong KM, Arikkatt J, Ullah MA, Lynch JP, Zhang V, Atkinson K, Sly PD, Phipps S. Immunomodulation of airway epithelium cell activation by mesenchymal stromal cells ameliorates house dust mite-induced airway inflammation in mice. Am J Respir Cell Mol Biol. 2015;53:615–624. doi: 10.1165/rcmb.2014-0431OC. [DOI] [PubMed] [Google Scholar]
- 17.Trzil JE, Masseau I, Webb TL, Chang CH, Dodam JR, Cohn LA, Liu H, Quimby JM, Dow SW, Reinero CR. Long-term evaluation of mesenchymal stem cell therapy in a feline model of chronic allergic asthma. Clin Exp Allergy. 2014;44:1546–1557. doi: 10.1111/cea.12411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Trzil JE, Masseau I, Webb TL, Chang CH, Dodam JR, Liu H, Quimby JM, Dow SW, Reinero CR. Intravenous adipose-derived mesenchymal stem cell therapy for the treatment of feline asthma: a pilot study. J Feline Med Surg. 2016;18:981–990. doi: 10.1177/1098612X15604351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Németh K, Leelahavanichkul A, Yuen PS, Mayer B, Parmelee A, Doi K, Robey PG, Leelahavanichkul K, Koller BH, Brown JM, Hu X, Jelinek I, Star RA, Mezey E. Bone marrow stromal cells attenuate sepsis via prostaglandin E(2)-dependent reprogramming of host macrophages to increase their interleukin-10 production. Nat Med. 2009;15:42–49. doi: 10.1038/nm.1905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mathias LJ, Khong SM, Spyroglou L, Payne NL, Siatskas C, Thorburn AN, Boyd RL, Heng TS. Alveolar macrophages are critical for the inhibition of allergic asthma by mesenchymal stromal cells. J Immunol. 2013;191:5914–5924. doi: 10.4049/jimmunol.1300667. [DOI] [PubMed] [Google Scholar]
- 21.Song X, Xie S, Lu K, Wang C. Mesenchymal stem cells alleviate experimental asthma by inducing polarization of alveolar macrophages. Inflammation. 2015;38:485–492. doi: 10.1007/s10753-014-9954-6. [DOI] [PubMed] [Google Scholar]
- 22.Braza F, Dirou S, Forest V, Sauzeau V, Hassoun D, Chesné J, Cheminant-Muller MA, Sagan C, Magnan A, Lemarchand P. Mesenchymal stem cells induce suppressive macrophages through phagocytosis in a mouse model of asthma. Stem Cells. 2016;34:1836–1845. doi: 10.1002/stem.2344. [DOI] [PubMed] [Google Scholar]
- 23.Jiang Z, Zhu L. Update on the role of alternatively activated macrophages in asthma. J Asthma Allergy. 2016;9:101–107. doi: 10.2147/JAA.S104508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Takeda K, Dow SW, Miyahara N, Kodama T, Koya T, Taube C, Joetham A, Park JW, Dakhama A, Kedl RM, Gelfand EW. Vaccine-induced CD8+ T cell-dependent suppression of airway hyperresponsiveness and inflammation. J Immunol. 2009;183:181–190. doi: 10.4049/jimmunol.0803967. [DOI] [PubMed] [Google Scholar]
- 25.Webb TL, Quimby JM, Dow SW. In vitro comparison of feline bone marrow-derived and adipose tissue-derived mesenchymal stem cells. J Feline Med Surg. 2012;14:165–168. doi: 10.1177/1098612X11429224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Varol C, Mildner A, Jung S. Macrophages: development and tissue specialization. Annu Rev Immunol. 2015;33:643–675. doi: 10.1146/annurev-immunol-032414-112220. [DOI] [PubMed] [Google Scholar]
- 27.DelaRosa O, Lombardo E, Beraza A, Mancheño-Corvo P, Ramirez C, Menta R, Rico L, Camarillo E, García L, Abad JL, Trigueros C, Delgado M, Büscher D. Requirement of IFN-γ gamma-mediated indoleamine 2, 3-dioxygenase expression in the modulation of lymphocyte proliferation by human adipose-derived stem cells. Tissue Eng Part A. 2009;15:2795–2806. doi: 10.1089/ten.TEA.2008.0630. [DOI] [PubMed] [Google Scholar]
- 28.Liang C, Jiang E, Yao J, Wang M, Chen S, Zhou Z, Zhai W, Ma Q, Feng S, Han M. Interferon-γ mediates the immunosuppression of bone marrow mesenchymal stem cells on T-lymphocytes in vitro. Hematology. 2017;5:1–6. doi: 10.1080/10245332.2017.1333245. [DOI] [PubMed] [Google Scholar]
- 29.Ogulur I, Gurhan G, Aksoy A, Duruksu G, Inci C, Filinte D, Kombak FE, Karaoz E, Akkoc T. Suppressive effect of compact bone-derived mesenchymal stem cells on chronic airway remodeling in murine model of asthma. Int Immunopharmacol. 2014;20:101–109. doi: 10.1016/j.intimp.2014.02.028. [DOI] [PubMed] [Google Scholar]
- 30.Mariñas-Pardo L, Mirones I, Amor-Carro O, Fraga-Iriso R, Lema-Costa B, Cubillo I, Rodríguez Milla MÁ, García-Castro J, Ramos-Barbón D. Mesenchymal stem cells regulate airway contractile tissue remodeling in murine experimental asthma. Allergy. 2014;69:730–740. doi: 10.1111/all.12392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ahmad T, Mukherjee S, Pattnaik B, Kumar M, Singh S, Kumar M, Rehman R, Tiwari BK, Jha KA, Barhanpurkar AP, Wani MR, Roy SS, Mabalirajan U, Ghosh B, Agrawal A. Miro1 regulates intercellular mitochondrial transport and enhances mesenchymal stem cell rescue efficacy. EMBO J. 2014;33:994–1010. doi: 10.1002/embj.201386030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kim J, Hematti P. Mesenchymal stem cell-educated macrophages: a novel type of alternatively activated macrophages. Exp Hematol. 2009;37:1445–1453. doi: 10.1016/j.exphem.2009.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Zasłona Z, Przybranowski S, Wilke C, van Rooijen N, Teitz-Tennenbaum S, Osterholzer JJ, Wilkinson JE, Moore BB, Peters-Golden M. Resident alveolar macrophages suppress, whereas recruited monocytes promote, allergic lung inflammation in murine models of asthma. J Immunol. 2014;193:4245–4253. doi: 10.4049/jimmunol.1400580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Rőszer T. Understanding the mysterious M2 macrophage through activation markers and effector mechanisms. Mediators Inflamm. 2015;2015:816460. doi: 10.1155/2015/816460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Murray PJ, Allen JE, Biswas SK, Fisher EA, Gilroy DW, Goerdt S, Gordon S, Hamilton JA, Ivashkiv LB, Lawrence T, Locati M, Mantovani A, Martinez FO, Mege JL, Mosser DM, Natoli G, Saeij JP, Schultze JL, Shirey KA, Sica A, Suttles J, Udalova I, van Ginderachter JA, Vogel SN, Wynn TA. Macrophage activation and polarization: nomenclature and experimental guidelines. Immunity. 2014;41:14–20. doi: 10.1016/j.immuni.2014.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Magnan AO, Mély LG, Camilla CA, Badier MM, Montero-Julian FA, Guillot CM, Casano BB, Prato SJ, Fert V, Bongrand P, Vervloet D. Assessment of the Th1/Th2 paradigm in whole blood in atopy and asthma. Increased IFN-gamma-producing CD8(+) T cells in asthma. Am J Respir Crit Care Med. 2000;161:1790–1796. doi: 10.1164/ajrccm.161.6.9906130. [DOI] [PubMed] [Google Scholar]
- 37.Yoshida M, Watson RM, Rerecich T, O’Byrne PM. Different profiles of T-cell IFN-gamma and IL-12 in allergen-induced early and dual responders with asthma. J Allergy Clin Immunol. 2005;115:1004–1009. doi: 10.1016/j.jaci.2005.02.003. [DOI] [PubMed] [Google Scholar]
- 38.Kanda A, Driss V, Hornez N, Abdallah M, Roumier T, Abboud G, Legrand F, Staumont-Sallé D, Quéant S, Bertout SFleury J, Rémy P, Papin JP, Julia V, Capron M, Dombrowicz D. Eosinophil-derived IFN-gamma induces airway hyperresponsiveness and lung inflammation in the absence of lymphocytes. J Allergy Clin Immunol. 2009;124:573–582. doi: 10.1016/j.jaci.2009.04.031. [DOI] [PubMed] [Google Scholar]
- 39.Yang M, Kumar RK, Foster PS. Interferon-gamma and pulmonary macrophages contribute to the mechanisms underlying prolonged airway hyperresponsiveness. Clin Exp Allergy. 2010;40:163–173. doi: 10.1111/j.1365-2222.2009.03393.x. [DOI] [PubMed] [Google Scholar]
- 40.Ryan JM, Barry F, Murphy JM, Mahon BP. Interferon-gamma does not break, but promotes the immunosuppressive capacity of adult human mesenchymal stem cells. Clin Exp Immunol. 2007;149:353–363. doi: 10.1111/j.1365-2249.2007.03422.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kumar RK, Yang M, Herbert C, Foster PS. Interferon-γ, pulmonary macrophages and airway responsiveness in asthma. Inflamm Allergy Drug Targets. 2012;11:292–297. doi: 10.2174/187152812800958951. [DOI] [PubMed] [Google Scholar]
- 42.Sivanathan KN, Gronthos S, Rojas-Canales D, Thierry B, Coates PT. Interferon-gamma modification of mesenchymal stem cells: implications of autologous and allogeneic mesenchymal stem cell therapy in allotransplantation. Stem Cell Rev. 2014;10:351–375. doi: 10.1007/s12015-014-9495-2. [DOI] [PubMed] [Google Scholar]
- 43.Taddio A, Tommasini A, Valencic E, Biagi E, Decorti G, De Iudicibus S, Cuzzoni E, Gaipa G, Badolato R, Prandini A, Biondi A, Ventura A. Failure of interferon-γ pre-treated mesenchymal stem cell treatment in a patient with Crohn’s disease. World J Gastroenterol. 2015;21:4379–4384. doi: 10.3748/wjg.v21.i14.4379. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.