

Abstract
Current models and theories of semantic knowledge primarily capture taxonomic relationships (DOG and WOLF), and largely do not address the role of thematic relationships in semantic knowledge (DOG and LEASH). Recent evidence suggests that processing or representation of thematic relationships may be distinct from taxonomic relationships. If taxonomic and thematic relations are distinct, then there should be a cost associated with switching between them even when the task remains constant. This hypothesis was tested using two different semantic relatedness judgment tasks: Experiment 1 used a triads task and Experiment 2 used an oddball task. In both experiments, participants were faster to respond when the same relationship appeared on consecutive trials than when the relationship types were different, even though the task remained the same and the specific relations were different on each trial. These results are consistent with the theory that taxonomic and thematic relations rely on distinct processes or representations.
Keywords: Semantic Memory, Taxonomic, Thematic
Semantic knowledge is a crucial aspect of human cognition. It allows us to understand language, interact with objects, remember the past, and imagine future events (Buxbaum & Saffran, 2002; Irish, Addis, Hodges, & Piguet, 2012; McRae & Jones, 2013). The representation, organization, cognitive algorithms, and neural implementation associated with this knowledge therefore plays an important role in various cognitive processes. Dating back at least to the 19th century, philosophers noted that there were different kinds of relationships within semantic knowledge. One particularly important distinction is between relationships based on similarity and relationships based on contiguity (Bain, 1864). In contemporary cognitive psychology and cognitive neuroscience, concepts are considered similar if they share features (e.g., DOG and WOLF; fur, tail, four legs etc.) and this relationship tends to place concepts into taxonomic structures based on category membership (e.g., animals). On the other hand, contiguity is based on co-occurrence in time and place (e.g., DOG and LEASH; walking a dog), which are typically called “thematic” relations. Although research has progressed quickly in regards to the representation and processing of concepts that share similar features, the field has not done as good a job of accounting for the integration and processing of relations based on contiguity and/or thematic semantics.
The taxonomic-thematic distinction is important because these relationships are dissociable and intrinsic to long-term semantic knowledge. Other kinds of semantic relations can be formed on-line in response to task instructions (e.g., ad-hoc categories such as “things to save from a burning house” or single-feature categories like “things that are orange”), but these are not part of long-term semantic knowledge and typically engage additional processes such as cognitive control (Lupyan, 2009; Lupyan & Mirman, 2013). Finally, associative relationships (e.g. those formed in free word association tasks) are defined by the free association task rather than a type of semantic relation, and are made up of various types of relationships (e.g., feature based, event based, compound words, etc.). For further review of the distinction between taxonomic and thematic semantics from other categories see Mirman, Landrigan, & Britt, 2017.
Contemporary theories of semantic memory agree that semantic knowledge relies on a distributed system that integrates modality-specific representations at various convergence zones and/or levels (Allport, 1985; Binder, Desai, Graves, & Conant, 2009; Fernandino et al., 2015; Gainotti, 2011; Meyer & Damasio, 2009). Although a variety of phenomena can be captured without including a “hub” or convergence zones (Cree, McRae, & McNorgan, 1999; Mirman & Magnuson, 2008; O’Connor, Cree, & McRae, 2009; Rabovsky & McRae, 2014), most models include either one central hub (e.g., (Lambon Ralph, Jefferies, Patterson, & Rogers, 2017; Rogers, Lambon Ralph, Garrard, et al., 2004) or a series of convergence zones (e.g., McNorgan, Reid, & McRae, 2011; Meyer & Damasio, 2009). Studies of the neural basis of semantic cognition have identified at least one central hub in the anterior temporal lobe (ATL) (Lambon Ralph et al., 2017; Patterson, Nestor, & Rogers, 2007). Computational implementations of this hub-and-spoke model have provided very precise accounts of a broad range of findings from individuals with semantic deficits as well as from neurologically intact participants (Barsalou, Simmons, Barbey, & Wilson, 2003; Chen, Lambon Ralph, & Rogers, 2017; Cree & McRae, 2003; Lambon Ralph et al., 2017; Patterson et al., 2007; Rogers, Lambon Ralph, Garrard, et al., 2004; Rogers, Lambon Ralph, Hodges, & Patterson, 2004; Rogers & Patterson, 2007). However, these models capture phenomena primarily related to taxonomic relationships, but ignore the role of thematic relationships. For example, individuals with semantic dementia (also called the semantic variant of primary progressive aphasia) produce a very specific pattern of exclusively taxonomic semantic errors in tasks such as picture naming, word comprehension, and object decision (e.g., Hurley, Paller, Rogalski, & Mesulam, 2012; Patterson et al., 2007). These error patterns are associated with ATL degeneration and are captured very well by hub-and-spoke computational models.
In contrast, individuals with post-stroke aphasia produce both taxonomic (e.g., HORSE for COW) and thematic (e.g., LEASH for DOG) errors in picture naming (Schwartz et al., 2011). In a recent systematic review, we synthesized results from over 100 studies that examined the distinction between taxonomic and thematic relations (Mirman et al., 2017). Key lines of evidence have shown that taxonomic and thematic relations make independent contributions to semantic relatedness (e.g., Estes, Gibbert, Guest, & Mazursky, 2012; Wisniewski & Bassok, 1999), that they have different time courses of activation (e.g., Kalénine, Mirman, Middleton, & Buxbaum, 2012), and that individuals differ in their reliance on taxonomic vs. thematic relations (e.g., Dunham & Dunham, 1995; Mirman & Graziano, 2012), including a neuropsychological double dissociation showing that taxonomic and thematic knowledge can be impaired separately (e.g., Au, Chan, & Chiu, 2003; Merck, Jonin, Laisney, Vichard, & Belliard, 2014; Semenza, Denes, Lucchese, & Bisiacchi, 1980; Semenza, Bisiacchi, & Romani, 1992).
Jackson and colleagues (2015) found that taxonomic and thematic processing activated overlapping brain regions1 and proposed that a single hub-and-spoke model could account for both kinds of relations by treating thematic relations as a kind of feature. Using their example: the thematic relation between croissants and coffee could be captured by allowing the concept CROISSANT to have a <coffee> feature (or perhaps <eaten with coffee>). However, on this approach, the <coffee> feature would be completely independent of the COFFEE concept because the hub-and-spoke model makes a strong distinction between concepts, which are representations based on convergent features in the hub, and features, which are represented in the spokes. Therefore, the <coffee> feature that is part of the CROISSANT concept would not be the same as the COFFEE concept, which would have its own set of features. Further, if thematic relations are features, then the CROISSANT-<coffee> relationship has nothing in common with other thematic relations such as BOAT-<river>, DOG-<leash>. As described below, the present experiments were designed to test this hypothesis – that thematic relations have something in common that is substantively distinct from taxonomic relations.
One possible explanation for the observed differences between taxonomic and thematic relations is that thematic relationships are also based on features, but either the features are different from those that typically form taxonomic relationships, or the features are integrated differently and therefore require a separate processing mechanism (Mirman et al., 2017). This is related to the account proposed by Jackson et al. (2015), but differs in that thematic relations are not themselves features, but, like taxonomic relations, are based on convergent, distributed representations over multiple features. The present experiments sought to test this hypothesis by adapting a method that has been previously used to study the role of modality-specific perceptual features in semantic processing (e.g., McNorgan, Reid, & McRae, 2011; Pecher, Zeelenberg, & Barsalou, 2003). In those studies, participants performed a feature verification task (e.g., “BLENDER can be LOUD”). The key finding was that reaction times were about 20–40ms slower when subsequent trials queried a feature from a different modality (e.g., “CRANBERRIES can be TART”) than the same modality (e.g., “LEAVES can be RUSTLING”). This switch cost was interpreted to mean that distinct, modality-specific feature representations were engaged during this task even though the task made no explicit reference to mental imagery or feature modality.
The present experiments used a similar approach, but with tasks that tapped multimodal taxonomic or thematic relations. Following Pecher et al. (2003), participants performed a single semantic relatedness judgment task throughout each experiment with no explicit instructions about the nature of the taxonomic or thematic relationships. The tasks ensured that the participants would engage their semantic system(s) and that they would identify particular semantic relations without explicit instruction to focus on taxonomic or thematic relatedness. This prevented any potential switch costs from being attributed to higher-level explicit task maintenance, updating, or conflict processes. The critical relation on each trial was either taxonomic or thematic and adjacent trials could have the same kind of relation (e.g. taxonomic to taxonomic or thematic to thematic) or different relations (e.g. taxonomic to thematic or thematic to taxonomic). The taxonomic and thematic relationships used in the present experiments were not reducible to single modalities or feature types. For example, some taxonomic relationships were purely feature based (e.g., SPAGHETTI and WORMS), while others were category based (e.g., BOAT and TRAIN). Similarly, the thematic relationships varied as well (e.g., SAW and TREE, MOON and ASTRONAUT). Thus, if a switch cost is observed between taxonomic and thematic trials, it would have to be above and beyond any cost associated with switching between the feature types within a single relationship type, suggesting a commonality among the taxonomic relationships that is distinct from thematic relationships. In sum, if taxonomic and thematic relationships rely on different features or if those features are integrated in different ways, then it should be possible to observe similar switch costs when the subsequent trials differ as compared to when they are the same.
Experiment 1: Triads
Materials and Methods
Participants
Thirty-two participants were recruited from Drexel University Psychology courses (24 females). The mean age of the participants was 19.9 years old (SD = 1.8). All participants were native English speakers or multilingual speakers who had learned English by the age of 5 years old. All participants provided informed consent in accordance with procedures approved by the Drexel University Institutional Review Board. Participants received course credit for their participation. No participants were excluded from participation or analysis.
Materials
Candidate word pairs were drawn from a prior norming study (Landrigan & Mirman, 2016). In short, each pair (a reference word and its potential taxonomic and thematic targets) were normed on average by 20 participants in separate surveys using Amazon Mechanical Turk for both their taxonomic relatedness (sharing similar features and/or belonging to the same category; participants rated the word pairs on a scale of 1 Not Similar at All to 7 Very Similar) and their thematic relatedness (co-occurrence in common events; participants rated the word pairs on a scale of 1 Not Related at All to 7 Very Related). Pairs and/or triads were then considered for inclusion if they showed a clear dichotomy between their taxonomic and thematic relationships. More specifically, the reference and its two targets had to exhibit unique relationships such that the reference and its taxonomic pair word had high taxonomic similarity and low thematic relatedness, whereas the reference and its thematic pair word had high thematic relatedness and low taxonomic similarity.2 For the full stimulus list see Appendix A and for a full description of the norming procedure see Landrigan and Mirman (2016). Taxonomically and thematically related words were shuffled to become the unrelated items for other triads and normed using the same method as the taxonomic and thematic pairs to control for unforeseen relationships. This shuffling was done to control for other properties of the words that may affect the task, thus all taxonomic items appeared as both a taxonomically-related item for its target word and as the unrelated item for a different target, and the same was done for thematic items. All conditions were also matched on word length in letters and phonemes (obtained from the Speech & Hearing Lab Neighborhood Database at Washington University in St. Louis), word frequency (Brysbaert & New, 2009), orthographic neighborhood sizes (Marian, Bartolotti, Chabal, & Shook, 2012), and imageability obtained from the MRC Psycholinguistic Database (Wilson, 1988) see Table 1.
Table 1.
Mean Semantic and Lexical Properties of all Taxonomic and Thematic Words
| Taxonomic | Thematic | Comparisons | |
|---|---|---|---|
| Similarity Rating | 4.79 (1.01) | 2.89 (0.87) | t(94) = 9.72, p < .001 |
| Relatedness Rating | 3.23 (0.94) | 5.79 (0.87) | t(94) = −13.69, p < .001 |
| Difference Score | 1.55 (0.55) | 2.90 (0.78) | t(94) = −9.72, p < .001 |
| Number of Letters | 5.25 (1.98) | 5.33 (1.65) | t(94) = −0.22, p = 0.83 |
| Number of Phonemes | 4.38 (1.73) | 4.52 (1.55) | t(94) = −0.43, p = 0.67 |
| Word Frequency (log) | 1.23 (0.6) | 1.39 (0.51) | t(94) = −1.40, p = 0.16 |
| Orthographic Neighborhood | 10.17 (10.17) | 7.21 (7.65) | t(94) = 1.59, p = 0.11 |
| Imageability* | 578.47 (42.14) | 590.48 (40.62) | t(67) = −1.20, p = 0.23 |
Note. SE in parentheses.
Imageability ratings were only available for 69 of the 96 words (72%).
Four experimental trial lists were created consisting of two blocks each. Lists were counterbalanced so that each reference item appeared in both taxonomic and thematic trials and in “same” (taxonomic-to-taxonomic or thematic-to-thematic) and “switch” trials (taxonomic-to-thematic defined as a thematic switch trial or thematic-to-taxonomic defined as a taxonomic switch trial). Each target word appeared once in a block. Within a single list, each reference word was presented in a taxonomic trial in one block and in a thematic trial in the other block; the same/switch counterbalancing was done across lists (and, thus, across participants). In sum, each list contained 96 trials in a 2 (trial type: taxonomic vs. thematic) by 2 (task type: same vs. switch) design with 24 trials in each cell of the design. Participants were randomly assigned to 1 of the 4 total lists resulting in 8 participants completing each list.
Procedure
The experiment was run using PsychoPy software. Instructions were to select the word that was most related to the word at the top of the screen (i.e. the reference word) by pressing Z for the word on the bottom left and M for the word on the bottom right. The response options were randomly assigned to sides on each trial. Reaction time recordings began at the presentation of the words until the participant’s response. Each trial was followed by an inter-trial interval (ITI) of 500 ms before the start of the next trial. After completing 5 practice trials with feedback, participants were told the experiment would begin and no more feedback would be given. The first 5 trials after the practice portion were filler trials (not analyzed) to allow for further practice and optimal performance on all analyzed trials. Midway through the experimental list of trials (between the two blocks) there was a break provided. The first trial after the midway break was excluded as it was neither a same nor a switch trial. The trial sequence is illustrated and described in Figure 1.
Figure 1.
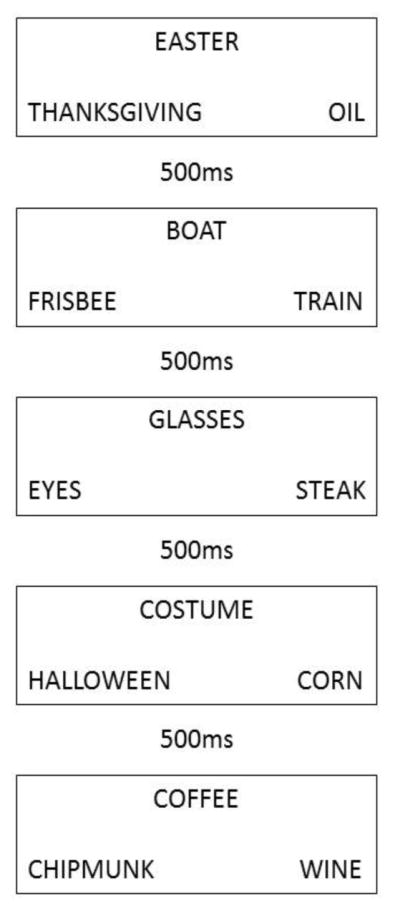
Example timeline of trials. Each box represents a single trial with a 500ms ITI between trials. The first (EASTER-THANKSGIVING) and second (BOAT-TRAIN) trials are taxonomic trials and thus the second trial is considered a taxonomic same trial. The third trial (GLASSES-EYES) is a thematic trial and therefore this is considered a thematic switch trial. The fourth trial (COSTUME-HALLOWEEN) in the sequence is also a thematic trial and is therefore a thematic same trial. The final trial (COFFEE-WINE) is once again a taxonomic trial and thus is a taxonomic switch trial.
Results
Data were analyzed using multilevel regression implemented in R version 3.1.2 (R Core Team, 2014). Reaction times of incorrect trials and trials where the reaction time was less than 250 ms were excluded. The subsequent trial was also excluded because, if the participant responded incorrectly or before processing the stimuli, then it was assumed they did not understand the probed relationship and therefore the subsequent trial would be neither a same nor a switch trial. Exclusion criteria resulted in a total of 218 out of the original 3072 experimental trials (7%) being dropped from further analysis. Trials with the target item WAGON were excluded due to poor overall accuracy (69%) across participants (N = 44 trials). All participants had reasonably high overall accuracy (range = 87% – 100%, M = 96%), so none were excluded from analysis. Error counts by condition are shown in Table 2. After exclusions, the total number of remaining observations was 2810. Using the lme4 package version 1.1–7, a linear mixed effects model was employed to analyze the raw reaction time data (Baayen, Davidson, & Bates, 2008). The model included fixed effects of trial type (taxonomic or thematic) and task type (same or switch) along with random effects of trial type by reference word and trial type and task type by participant (see Table 2 note for model syntax). All p-values were estimated using the normal distribution. Table 2 shows model-estimated condition means and SE.
Table 2.
Experiment 1 Results
| Reaction time (ms): Mean (SE) | Error rate | ||||
|---|---|---|---|---|---|
|
| |||||
| Same Trials | Switch Trials | Switch Cost | Same Trials | Switch Trials | |
| Taxonomic | 2044 (122) | 2132 (129) | 88 | .04 | .03 |
| Thematic | 1741 (74) | 1801 (81) | 60 | .03 | .03 |
Note. Model: ReactionTime ~ TrialType + TaskType + (TaskType : TrialType) + (1 + TrialType | RefWord) + (1 + TrialType + TaskType | Participant)
Participants were faster to respond to thematic trials (M = 1771ms, SE = 74ms) than taxonomic trials (M = 2088ms, SE = 123ms), Estimate = 158.55, SE = 51.29, p < 0.01. Critically, participants were slower in switch trials (M = 1965ms, SE = 93ms) than in same trials (M = 1891ms, SE = 85ms), Estimate = −37.19, SE = 17.48, p = 0.03. The interaction between trial type and task type was not statistically significant, Estimate = −6.96, SE = 16.81, p = 0.68.
Discussion
These results show a clear switch cost: responses were about 74ms faster when the kind of relatedness (taxonomic or thematic) was the same as the preceding trial, than when it was the other kind of relatedness. This pattern is consistent with the proposal outlined in the Introduction: taxonomic and thematic relationships may be represented or processed in different ways leading to the observed switch cost. Critically, this switch cost was observed in the absence of any explicit instructions about taxonomic and thematic relationships and across different kinds of taxonomic or thematic relationships.
Experiment 2: Oddball
To strengthen the evidence from Experiment 1, the switch cost hypothesis was further examined using a more complex semantic judgment task. Prior research suggests that similarity and dissimilarity judgments are not simply the inverse of each other (Golonka & Estes, 2009; Simmons & Estes, 2008). To test dissimilarity, studies have used an odd-one-out or oddball task in which participants need to process both relatedness and difference in order to identify the oddball, either based on general semantic relatedness or based on specific features (Davidoff & Roberson, 2004; Lupyan, 2009). Therefore, following this vein, we tested whether the observed switch cost from Experiment 1 would also emerge when participants made semantic difference or oddball judgments. In addition, the oddball task provided an opportunity to test the switch cost when the two relation types are (hypothetically) in competition with one another. That is, when the critical relation type from one trial needs to be suppressed on the subsequent trial, and when a previously-suppressed relation type becomes the critical relation on the next trial.
Experiment 2 used a similar same/switch design as Experiment 1, however participants had to identify which one of four items was least related to the others. Each trial contained a triplet of items that shared one kind of relationship and an “oddball” item that was related to one of the other items in the opposing relationship. Thus, in order to correctly identify the dominant triplet, participants needed to suppress the opposing relationship. Therefore, when switching between taxonomic and thematic dominance, participants would have to re-activate the previously suppressed relationship in order to correctly identify the new dominant relationship and subsequently identify the oddball. Although this differs from prior oddball experiments (e.g. Lupyan, 2009) using four items allowed us to present participants with both relationships on each trial so that they could compete with one another. This would therefore force participants to engage their semantic systems at an even deeper level than in the triads task as participants need to consider all possible relationships on each trial and suppress and reactivate relationships in order to identify the correct oddballs.
Materials and Methods
Participants
Thirty-two participants were recruited from Drexel University Psychology courses (22 females). Recruitment for the two experiments was independent, but two of the overall total of 62 participants completed both experiments. Both of them completed Experiment 2 at least 6 months after completing Experiment 1. The mean age of the participants was 20.8 years old (SD = 1.9). All participants were native English speakers or multilingual speakers who had learned English by the age of 6 years old. All participants provided informed consent in accordance with procedures approved by the Drexel University Institutional Review Board. Participants received course credit for their participation.
Materials
Each trial in this experiment consisted of the simultaneous presentation of four words. Three words were related either taxonomically or thematically and the fourth word was related to a reference word in the opposite relationship. Word quartets were constructed by taking the triads from Experiment 1 and adding an extra thematic or taxonomic item. Each quartet was then divided into the four possible three-word subsets and these triplets were normed using Qualtrics and Mechanical Turk. Participants were asked to rate on a scale of 1 to 7 how well the triplets grouped together based on co-occurrence in time or place or based on similarity (for an example see Table 3).
Table 3.
Example Trial Quartet: EGG, RABBIT, EASTER, THANKSGIVING.
| Triplet | Oddball | Group Rating |
|---|---|---|
| EASTER, RABBIT, EGG | THANKSGIVING | 6.15 |
| EASTER, RABBIT, THANKSGIVING | EGG | 2.37 |
| EASTER, EGG, THANKSGIVING | RABBIT | 2.35 |
| RABBIT, EGG, THANSGIVING | EASTER | 2.00 |
Note. The rows show each possible combination of triplet, oddball, and the mean group rating of the triplet. In this example, THANKSGIVING is the correct oddball and all triplets that included it received lower group ratings than the correct triplet that excluded it.
Trial quartets were only included in Experiment 2 if the correct triplet had a mean normed rating of at least 0.5 points higher than the other possible triplets for that quartet. That is, any triplet involving the “oddball” had to be at least 0.5 points less well-formed than the correct triplet that omitted the oddball. This relatively low threshold was used in order to maximize the number of possible trials: excluding error trials was selected over trying to guess which items were likely to elicit many errors. The oddball words and non-oddball words were also matched on word length in letters and phonemes (obtained from the Speech & Hearing Lab Neighborhood Database at Washington University in St. Louis), word frequency (Brysbaert & New, 2009), and orthographic neighborhood sizes (Marian et al., 2012). The norming criteria resulted in a final list of 64 quartets (i.e. 32 taxonomic quartets and 32 thematic quartets, see Appendix B for the full set of stimuli).
Experimental trial lists were created in the same manner as Experiment 1. Note that both kinds of relations were present on each trial: as illustrated in Figure 2, each trial contained a dominant relation that bound three items (in the example, the thematic relation that binds EASTER, RABBIT, and EGG) and a subordinate relation that bound the oddball with one of the items (in the example, the taxonomic relation between EASTER and THANKSGIVING). In sum, there were 32 trials per block, totaling 64 critical trials per participant in a 2 (trial type: taxonomic vs thematic) by 2 (task type: same vs switch) design with 16 trials in each cell. Participants were randomly assigned to 1 of the 4 lists resulting in 8 participants completing each list.
Figure 2.

Example trial presentation with a taxonomic oddball and dominant thematic grouping. Lines indicate relationships and are included in this schematic but not in the actual trials.
Procedure
The experiment was run using PsychoPy software. Participants were instructed to select the word that does not fit with the others by clicking on the word with the mouse. The words (including the oddball) were randomly assigned to one of four positions/quadrants on each trial. After each trial, the participant was presented with a cross in the middle of the screen, which they needed to click on in order to initiate the next trial (following an ITI of 500 ms). This was done so the mouse would be in the center of the screen at the start of each oddball trial. A mouse response was used in place of a key response due to the increased number of response options participants (i.e. 2 response options in Experiment 1, but 4 response options in Experiment 2). Thus, in order to eliminate errors from simply hitting the wrong key, the mouse response was used. Figure 3 shows an example trial sequence. Practice trials and breaks were carried out in the same manner as in Experiment 1.
Figure 3.
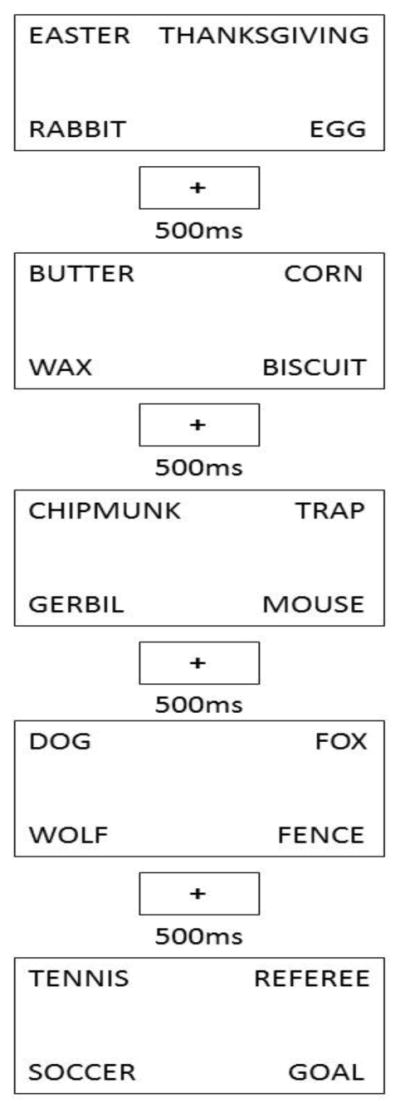
Example timeline of Experiment 2 trials. Each box represents a single trial with between trial cross presentations and 500ms ITI. The first trial (EASTER-RABBIT-EGG) is a thematic trial and the second trial (BUTTER-CORN-BISCUIT) is a thematic trial as well. Thus, the second trial is a thematic same trial. The third trial (CHIPMUNK-GERBIL-MOUSE) is a taxonomic trial and is therefore a taxonomic switch trial. The fourth trial (DOG-WOLF-FOX) is a taxonomic trial as well and is considered a taxonomic same trial. The last trial in this sequence (SOCCER-GOAL-REFEREE) is a thematic switch trial.
Results
Accuracy in Experiment 2 was much lower (M = 75%) than in Experiment 1, most likely due to the more open-ended nature of the task. Each trial contained a dominant relation that bound three items, but also a subordinate relation that bound two of the items. Binding a third item into the subordinate relation was all that was required for a participant to make an incorrect response, and given that a lower inclusion threshold was used in order to maximize the number of trials, some of the trials may have fallen victim to alternative interpretations.
Trial exclusion criteria for Experiment 2 were the same as Experiment 1. In total, 845 trials were excluded from analysis out of the original 2048 experimental trials (41%). In order to ensure a minimum level of data in each condition, 3 participants (38 total trials) were excluded from analyses because they did not have at least 3 analyzable trials in each of the 4 conditions. The final analyzed data set consisted of 1165 observations from 29 participants (M = 40 trials per participant, range = 14–58). Data were analyzed using the same model specifications as Experiment 1 (see Table 2).
Overall, participants were faster to respond to taxonomic trials (M = 5421ms, SE = 430ms) than thematic trials (M = 6526ms, SE = 475ms), Estimate = −554.93, SE = 221.09, p = 0.01. As in Experiment 1, participants were slower in switch trials (M = 6227ms, SE = 471ms) than in same trials (M = 5688ms, SE = 358ms), Estimate = −274.51, SE = 135.99, p = 0.04. The interaction between trial type and task type was not statistically significant, Estimate = 131.35, SE = 99.86, p = 0.19 (Table 4). Table 4 also shows that the errors followed the same pattern as the reaction times: more errors on switch trials than same trials. However, a logistic regression analysis with the same parameters as the reaction time model found no main effect of Task Type (Estimate = 0.12, SE = 0.08, p = 0.10), Trial Type (Estimate = 0.24, SE = 0.18, p = 0.19), nor their interaction (Estimate = −0.02, SE = 0.07, p = 0.82).
Table 4.
Experiment 2 Results
| Reaction time (ms): Mean (SE) | Error rate | ||||
|---|---|---|---|---|---|
|
| |||||
| Same Trials | Switch Trials | Switch Cost | Same Trials | Switch Trials | |
| Taxonomic | 5281 (401) | 5567 (516) | 286 | .18 | .21 |
| Thematic | 6128 (464) | 6940 (545) | 812 | .21 | .29 |
Note. Model: ReactionTime ~ TrialType + TaskType + (TaskType : TrialType) + (1 + TrialType | RefWord) + (1 + TrialType + TaskType | Participant)
Discussion
These results converge with the results of Experiment 1: when making an oddball judgment that one item does not fit with the dominant relation (taxonomic or thematic), if the preceding trial relied on the same kind of relation, responses were about 549ms faster than if that preceding trial relied on a different kind of relatedness. This switch cost is again consistent with the proposal that taxonomic and thematic relationships may be processed differently, such that when participants made judgements of the same type on consecutive trials while suppressing the other, their response times were quicker than when they switched between relationships.
General Discussion
The present study set out to test whether taxonomic and thematic relations rely on distinct representations or processing mechanisms. In both experiments, the task instructions were constant for all trials: triads relatedness judgment in Experiment 1, oddball judgment in Experiment 2. However, the materials for each trial were designed to make the participant engage in either taxonomic or thematic processing. The critical finding was that, across both experiments, participants were slower to respond to trials when the dominant relationship in consecutive trials switched (i.e. taxonomic to thematic, or thematic to taxonomic) compared to trials where the dominant relation type remained the same (Figure 4). These results suggest that taxonomic and thematic semantic processing are supported by different representations or processing mechanisms.
Figure 4.

Results of experiment 1 (left panel) and experiment 2 (right panel)
Within relation type, the individual pairs shared different features and relationship types, so this pattern is not reducible to a modality-specific effect. Any modality-specific effects (such as those demonstrated by Pecher et al., 2003; see also McNorgan et al., 2011) would cause virtually all trials to be “switch” trials, thus washing out the broader taxonomic-thematic switch cost. In other words, the variability in the relationship subtypes amongst thematic relations (e.g., agent-patient, location, etc.) and taxonomic relations (e.g., feature based, category based, etc.), prevents the observed switch cost effect from being attributed to those lower levels of relation types. At those lower levels, all of the trials would have been switch trials so there would not have been any additional effect of switching between taxonomic and thematic trials. Similarly, framing thematic relations as one kind of feature in a hub-and-spoke system (e.g., Jackson et al., 2015) would not account for the present switch costs because those thematic features (CROISSANT-<coffee>, BOAT-<river>, DOG-<leash>, etc.) would all be distinct and the cost of switching among them would be just as strong as the cost of switching to a taxonomic relation. Rather, these results indicate that thematic relations, as a group, share some commonality that is distinct from the analogous commonality shared by taxonomic relations. This could be accomplished by gathering all of thematic features into a single spoke and by connecting them to the other spokes so that, for example, the <coffee> feature could be connected to all of the features of the COFFEE concept. But doing so would create a thematic hub – a distinct representational basis for thematic semantics.
There was also a main effect of trial type (i.e., taxonomic versus thematic) in both experiments. The direction of this effect differed between experiments, most likely due to differences in the task demands. In the oddball experiment, the taxonomic trials may have been easier than thematic trials because the increased number of exemplars made it simpler to identify a common set of features between items or a common category to which the items belonged to (i.e., increased exemplars of a category or sharing features narrowed the search space). In contrast, themes are more flexible in nature, so all items and possible relationships would need to be considered on thematic trials in order to identify the true dominant relationship. A related effect has been observed in developmental studies in which increased numbers of taxonomic exemplars made learning and extending a superordinate label easier for children (Liu, Golinkoff, & Sak, 2001). In the triads task because there was a defined reference item and participants were only choosing between two response possibilities, a thematic relation may have been easier to detect than a taxonomic relation (a recent free sorting study found that adults exhibited more thematic sorting than taxonomic sorting: Lawson, Chang, & Wills, 2017). These effects are orthogonal to the observed switch costs that were the main finding of these experiments. The switch cost effects are explicitly consistent with distinct taxonomic and thematic processing mechanisms and the differential response to task demands only further suggests that these relationships are processed independently of each other.
We argue that distinct taxonomic and thematic representations or processing mechanisms are required to account for the present switch costs. If approached from the opposite view, it is not clear how a single processing mechanism would account for the present results. For example, a standard spreading activation model (e.g. Collins & Loftus, 1975; McNamara, 1992) does not distinguish between taxonomic and thematic relationships. In such a model, on a given trial, activation would spread to both taxonomically and thematically related concepts. The next trial would involve new, unrelated concepts and activation would again spread to both taxonomically and thematically related concepts. How long it takes to make a relatedness judgment would depend on strength of connections, but not the type of connection. A basic spreading activation model would need to be extended so that taxonomic and thematic connections differ in some way and use of one connection type facilitates subsequent use of the same type of connections or inhibits use of connections of the other type. This kind of “tagged connections” approach would predict a switch cost, but such a model would be simply one version of distinct taxonomic and thematic semantic systems – in this case, distinct systems of connections. In a hub-and-spoke model, as described above, the thematic features would need to be gathered into a single spoke and connected to the other spokes, which would create a thematic hub that is distinct from the taxonomic hub, in order to account for the switch cost.
Another alternative perspective is to consider relations as dynamic emergent properties of concepts and context. Indeed, context can influence which concept features and relations are activated and how strongly they are activated (e.g., Kalénine, Mirman, Middleton, & Buxbaum, 2012; Lee, Middleton, Mirman, Kalénine, & Buxbaum, 2013; Yee & Thompson-Schill, 2016). Although the overall task was constant within each of the present experiments, one could construe each trial context as dynamically emphasizing either taxonomic or thematic relations. However, the actual items and specific relations were different from trial to trial, so this context-specific emphasis would have to operate at the broad level of taxonomic vs. thematic relations, independent of the specific items and detailed relations. That is to say, it would also be a way of instantiating the distinct taxonomic and thematic systems.
It is important to acknowledge that semantic cognition does not operate independently of other cognitive systems. In particular, semantic cognition is strongly dependent on cognitive control processes (sometimes called “semantic control”) that allow manipulation and selection of semantic knowledge to suit task demands (Lambon Ralph et al., 2017; see also Jefferies, 2013) We are not aware of any evidence that either taxonomic or thematic semantic processing requires more semantic control. In fact, the well-documented neuropsychological double dissociation of taxonomic and thematic semantics argues against differences in the degree of semantic control required for taxonomic and thematic processing (for a review see Mirman et al., 2017). The reversal of the main effect of task type across our two experiments also argues against any differences in the amount of semantic control required. If differences existed then the reversal should not have appeared because the relationship requiring more semantic control would have elicited longer reaction times in both experiments. However, it is possible that taxonomic and thematic semantics require different semantic control. A switch cost could then arise as a result of switching between taxonomic control and thematic control processes operating over a single store of semantic knowledge. This interpretation shifts the dissociation from distinct taxonomic and thematic representations to distinct control mechanisms responsible for taxonomic and thematic processing. Further research is necessary to establish whether the dissociation of taxonomic and thematic semantics is in the representation of semantic knowledge or in the processing mechanisms that operate on that knowledge (for a review of the distinction between semantic knowledge and semantic access see Mirman & Britt, 2014).
These accounts are not the only possibilities, but they offer demonstrations of why the switch costs reported here require distinct taxonomic and thematic semantic representations or processing mechanisms. The present results do not require that taxonomic and thematic processes are modular – they could be distinct but highly interactive. These results also do not speak to specific neural implementations since the experiments were purely behavioral. From a levels-of-analysis perspective, these results suggest a computational or functional distinction between taxonomic and thematic processing, but do not speak to the algorithms or implementations that support this distinction. The degree of separation and interaction between taxonomic and thematic semantic systems, their neural basis, and whether the switch costs observed here are due to facilitation or inhibition are important topics for future research.
Acknowledgments
This study was supported by National Institutes of Health grant R01DC010805 to D.M. and by Drexel University. We thank Allison Britt, Amanda Kraft and Leah Friedman for help with data collection.
Appendix A: Stimuli used in experiment 1 (triads)
| Reference | Taxonomic | Sim | Rel | Sim - Rel | Thematic | Sim | Rel | Rel - Sim |
|---|---|---|---|---|---|---|---|---|
| easter | thanksgiving | 5.70 | 2.61 | 3.10 | rabbit | 3.31 | 5.98 | 2.67 |
| boat | train | 5.11 | 2.36 | 2.74 | river | 2.90 | 6.04 | 3.14 |
| glasses | goggles | 6.11 | 4.78 | 1.33 | eyes | 2.64 | 6.48 | 3.84 |
| honey | sap | 4.96 | 3.33 | 1.64 | bee | 3.84 | 6.67 | 2.83 |
| spoon | shovel | 4.57 | 2.56 | 2.01 | tea | 1.83 | 5.46 | 3.63 |
| backpack | dufflebag | 6.61 | 4.48 | 2.14 | student | 2.25 | 6.21 | 3.96 |
| ink | paint | 5.86 | 3.72 | 2.14 | printer | 3.64 | 6.29 | 2.65 |
| costume | uniform | 5.90 | 3.48 | 2.42 | halloween | 3.50 | 6.46 | 2.96 |
| peanut | bean | 4.75 | 3.05 | 1.71 | elephant | 1.65 | 5.09 | 3.44 |
| waitress | clerk | 4.73 | 2.62 | 2.11 | dinner | 2.82 | 5.57 | 2.75 |
| saw | knife | 5.43 | 3.23 | 2.20 | tree | 1.68 | 4.98 | 3.30 |
| spaghetti | worms | 2.61 | 1.27 | 1.35 | tomato | 3.57 | 5.88 | 2.31 |
| dorm | apartment | 5.12 | 4.26 | 0.86 | college | 3.59 | 6.21 | 2.62 |
| vault | box | 5.29 | 3.37 | 1.92 | bank | 4.64 | 6.16 | 1.52 |
| salt | sugar | 5.55 | 3.71 | 1.84 | pretzel | 3.32 | 5.74 | 2.42 |
| disk | frisbee | 4.86 | 4.31 | 0.56 | computer | 3.69 | 6.20 | 2.50 |
| pencil | stick | 4.25 | 2.92 | 1.34 | notebook | 3.50 | 6.82 | 3.32 |
| fork | rake | 3.83 | 2.22 | 1.61 | steak | 2.43 | 5.48 | 3.05 |
| dough | clay | 4.39 | 2.58 | 1.81 | pizza | 3.25 | 6.19 | 2.94 |
| syrup | oil | 4.06 | 2.13 | 1.93 | waffle | 3.38 | 6.43 | 3.05 |
| football | basketball | 5.87 | 3.59 | 2.28 | kick | 2.61 | 5.86 | 3.25 |
| moon | ball | 3.17 | 1.73 | 1.44 | astronaut | 3.44 | 6.43 | 3.00 |
| outlet | keyhole | 3.18 | 2.06 | 1.13 | plug | 4.61 | 6.44 | 1.83 |
| panda | grizzly | 5.28 | 3.20 | 2.09 | bamboo | 2.24 | 4.95 | 2.71 |
| dog | fox | 5.04 | 3.37 | 1.67 | fence | 1.47 | 4.60 | 3.14 |
| leash | rope | 5.09 | 3.73 | 1.36 | collar | 4.18 | 6.55 | 2.37 |
| dragon | snake | 3.79 | 2.65 | 1.14 | fire | 2.14 | 5.79 | 3.65 |
| butter | wax | 2.97 | 2.23 | 0.74 | corn | 2.58 | 5.11 | 2.54 |
| squirrel | rat | 5.23 | 3.25 | 1.98 | nut | 2.25 | 6.32 | 4.07 |
| ring | wheel | 3.69 | 2.29 | 1.40 | diamond | 4.01 | 6.14 | 2.13 |
| crown | hat | 5.29 | 4.15 | 1.14 | queen | 3.08 | 5.87 | 2.79 |
| church | store | 2.61 | 1.48 | 1.13 | bell | 2.00 | 5.78 | 3.78 |
| brick | boulder | 3.71 | 2.86 | 0.85 | wall | 3.85 | 5.79 | 1.95 |
| duck | swan | 5.99 | 4.88 | 1.11 | tub | 1.53 | 3.23 | 1.70 |
| wagon | car | 4.54 | 3.48 | 1.06 | fruit | 1.07 | 2.53 | 1.46 |
| ocean | lake | 5.59 | 4.12 | 1.48 | shark | 2.44 | 6.17 | 3.73 |
| gun | sword | 4.79 | 3.34 | 1.45 | cop | 2.25 | 6.36 | 4.11 |
| sign | label | 5.70 | 4.35 | 1.35 | street | 2.75 | 5.66 | 2.91 |
| needle | pin | 5.99 | 5.17 | 0.82 | doctor | 1.81 | 5.28 | 3.48 |
| stewardess | nurse | 4.29 | 2.16 | 2.13 | airplane | 2.36 | 6.48 | 4.13 |
| mouse | chipmunk | 5.08 | 3.16 | 1.92 | hole | 1.63 | 5.18 | 3.55 |
| breakfast | lunch | 5.39 | 4.07 | 1.32 | bacon | 4.53 | 6.28 | 1.75 |
| turnip | beet | 5.66 | 4.37 | 1.30 | salad | 3.40 | 4.06 | 0.67 |
| oyster | scallop | 5.59 | 4.65 | 0.94 | pearl | 3.75 | 6.50 | 2.75 |
| editor | host | 2.32 | 1.59 | 0.74 | article | 3.04 | 6.69 | 3.65 |
| anchor | weight | 5.64 | 3.90 | 1.74 | navy | 2.50 | 4.61 | 2.11 |
| soccer | tennis | 4.32 | 3.55 | 0.77 | goal | 2.79 | 6.65 | 3.86 |
| coffee | wine | 4.27 | 2.77 | 1.50 | mug | 3.06 | 6.54 | 3.49 |
Appendix B: Stimuli used in experiment 2 (oddball)
| Taxonomic Trials
| |||
|---|---|---|---|
| Reference | Related Item 1 | Related Item 2 | Oddball |
| anchor | weight | dumbbell | ship |
| backpack | suitcase | purse | student |
| boat | train | bus | wave |
| breakfast | lunch | snack | egg |
| brick | boulder | cement | roof |
| butter | wax | lard | corn |
| church | court | store | cross |
| coffee | wine | soda | donut |
| costume | uniform | disguise | play |
| crown | hat | yarmulke | castle |
| disk | frisbee | saucer | computer |
| dog | fox | wolf | fence |
| dorm | apartment | barrack | freshman |
| dough | clay | mortar | pizza |
| dragon | snake | iguana | fire |
| easter | thanksgiving | christmas | rabbit |
| football | basketball | hockey | tackle |
| fork | rake | hoe | steak |
| heart | stomach | kidney | valentine |
| honey | jelly | ketchup | bee |
| mouse | chipmunk | gerbil | trap |
| needle | thorn | spike | thread |
| outlet | keyhole | slot | electric |
| panda | grizzly | lion | bamboo |
| peanut | bean | walnut | elephant |
| salt | sugar | sand | fries |
| saw | knife | axe | tree |
| sign | label | billboard | directions |
| soccer | tennis | lacrosse | goal |
| squirrel | rat | hamster | nut |
| stewardess | nurse | technician | airplane |
| vault | box | container | money |
| anchor | navy | ship | dumbbell |
| backpack | books | student | purse |
| boat | river | wave | bus |
| breakfast | bacon | egg | snack |
| brick | roof | wall | boulder |
| butter | corn | biscuit | wax |
| church | bell | cross | court |
| coffee | donut | cream | soda |
| costume | halloween | play | uniform |
| crown | queen | castle | yarmulke |
| disk | music | computer | frisbee |
| dog | fence | house | wolf |
| dorm | college | freshman | barrack |
| dough | pizza | noodle | clay |
| dragon | fire | knight | iguana |
| easter | rabbit | egg | thanksgiving |
| football | kick | tackle | basketball |
| fork | fondue | steak | rake |
| heart | valentine | love | stomach |
| honey | bee | bear | jelly |
| mouse | cheese | trap | gerbil |
| needle | thread | seamstress | spike |
| outlet | electric | plug | keyhole |
| panda | bamboo | forest | lion |
| peanut | elephant | zoo | bean |
| salt | fries | pretzel | sand |
| saw | tree | carpenter | knife |
| sign | street | directions | label |
| soccer | goal | referee | tennis |
| squirrel | nut | nest | rat |
| stewardess | airplane | beverage | nurse |
| vault | bank | money | container |
Footnotes
The neural basis of semantic cognition is not the focus of this study, but we note that although Jackson et al. found no dissociation between taxonomic and thematic neural systems, several other studies have found that they do dissociate, so the existence and nature of a possible neural dissociation remains an open question (for systematic review and discussion see Mirman et al., 2017).
This dichotomous approach resulted in a bimodal distribution of relationship strengths, which was optimal for the factorial design but precluded a continuous analysis of the association between relationship strength of the word pairs and the observed switch cost. For bimodally-distributed data, continuous analyses tend to reproduce categorical analysis results even if the within-group patterns are in the opposite direction (Simpson’s paradox), making the continuous analysis redundant and possibly misleading.
References
- Allport DA. Distributed memory, modular subsystems and dysphasia. Current Perspectives in Dysphasia. 1985:32–60. [Google Scholar]
- Au A, Chan AS, Chiu H. Conceptual organization in Alzheimer’s dementia. Journal of Clinical and Experimental Neuropsychology. 2003;25(6):737–750. doi: 10.1076/jcen.25.6.737.16468. http://doi.org/10.1076/jcen.25.6.737.16468. [DOI] [PubMed] [Google Scholar]
- Baayen RH, Davidson DJ, Bates DM. Mixed-effects modeling with crossed random effects for subjects and items. Journal of Memory and Language. 2008;59(4):390–412. http://doi.org/10.1016/j.jml.2007.12.005. [Google Scholar]
- Bain A. In: The senses and the intellect. Longman Roberts, Green, editors. London: 1864. [Google Scholar]
- Barsalou LW, Simmons WK, Barbey AK, Wilson CD. Grounding conceptual knowledge in modality-specific systems. Trends in Cognitive Sciences. 2003;7(2):84–91. doi: 10.1016/s1364-6613(02)00029-3. [DOI] [PubMed] [Google Scholar]
- Binder JR, Desai RH, Graves WW, Conant LL. Where is the semantic system? A critical review and meta-analysis of 120 functional neuroimaging studies. Cerebral Cortex. 2009;19(December):2767–2796. doi: 10.1093/cercor/bhp055. http://doi.org/10.1093/cercor/bhp055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brysbaert M, New B. Moving beyond Kucera and Francis: a critical evaluation of current word frequency norms and the introduction of a new and improved word frequency measure for American English. Behavior Research Methods. 2009;41(4):977–990. doi: 10.3758/BRM.41.4.977. [DOI] [PubMed] [Google Scholar]
- Buxbaum LJ, Saffran EM. Knowledge of object manipulation and object function: Dissociations in apraxic and nonapraxic subjects. Brain and Language. 2002;82(2):179–199. doi: 10.1016/s0093-934x(02)00014-7. http://doi.org/10.1016/S0093-934X(02)00014-7. [DOI] [PubMed] [Google Scholar]
- Chen L, Lambon Ralph MA, Rogers TT. A unified model of human semantic knowledge and its disorders. Nature Human Behaviour. 2017;1(March):39. doi: 10.1038/s41562-016-0039. http://doi.org/10.1038/s41562-016-0039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cree GS, McRae K. Analyzing the factors underlying the structure and computation of the meaning of chipmunk, cherry, chisel, cheese, and cello (and many other such concrete nouns) Journal of Experimental Psychology. 2003;132(2):163–201. doi: 10.1037/0096-3445.132.2.163. http://doi.org/10.1037/0096-3445.132.2.163. [DOI] [PubMed] [Google Scholar]
- Cree GS, McRae K, McNorgan C. An attractor model of lexical conceptual processing: simulating semantic priming. Cognitive Science. 1999;23(3):371–414. http://doi.org/10.1016/S0364-0213(99)00005-1. [Google Scholar]
- Davidoff JB, Roberson D. Preserved thematic and impaired taxonomic categorisation: a case study. 2004:1–65. http://doi.org/10.1080/01690960344000125.
- Dunham PJ, Dunham F. Developmental antecedents of taxonomic and thematic strategies at 3 years of age. Developmental Psychology. 1995;31(3):483–493. [Google Scholar]
- Estes Z, Gibbert M, Guest D, Mazursky D. A dual-process model of brand extension: Taxonomic feature-based and thematic relation-based similarity independently drive brand extension evaluation. Journal of Consumer Psychology. 2012;22(1):86–101. http://doi.org/10.1016/j.jcps.2011.11.002. [Google Scholar]
- Fernandino L, Humphries CJ, Seidenberg MS, Gross WL, Conant LL, Binder JR. Predicting brain activation patterns associated with individual lexical concepts based on five sensory-motor attributes. Neuropsychologia. 2015;76:17–26. doi: 10.1016/j.neuropsychologia.2015.04.009. http://doi.org/10.1016/j.neuropsychologia.2015.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gainotti G. The organization and dissolution of semantic-conceptual knowledge: Is the “amodal hub” the only plausible model? Brain and Cognition. 2011;75(3):299–309. doi: 10.1016/j.bandc.2010.12.001. http://doi.org/10.1016/j.bandc.2010.12.001. [DOI] [PubMed] [Google Scholar]
- Golonka S, Estes Z. Thematic relations affect similarity via commonalities. Journal of Experimental Psychology. Learning, Memory, and Cognition. 2009;35(6):1454–1464. doi: 10.1037/a0017397. http://doi.org/10.1037/a0017397. [DOI] [PubMed] [Google Scholar]
- Hurley RS, Paller KA, Rogalski EJ, Mesulam MM. Neural Mechanisms of Object Naming and Word Comprehension in Primary Progressive Aphasia. Journal of Neuroscience. 2012;32(14):4848–4855. doi: 10.1523/JNEUROSCI.5984-11.2012. http://doi.org/10.1523/JNEUROSCI.5984-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Irish M, Addis DR, Hodges JR, Piguet O. Considering the role of semantic memory in episodic future thinking : evidence from semantic dementia. Brain. 2012:2178–2191. doi: 10.1093/brain/aws119. http://doi.org/10.1093/brain/aws119. [DOI] [PubMed]
- Jackson RL, Hoffman P, Pobric G, Ralph MaL. The Neural Correlates of Semantic Association versus Conceptual Similarity. Cerbral Cortex. 2015;20(2010):8524. doi: 10.1093/cercor/bhv003. http://doi.org/10.1093/cercor/bhv003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jefferies E. The neural basis of semantic cognition : Converging evidence from neuropsychology, neuroimaging and TMS. Cortex. 2013;49(3):611–625. doi: 10.1016/j.cortex.2012.10.008. http://doi.org/10.1016/j.cortex.2012.10.008. [DOI] [PubMed] [Google Scholar]
- Kalénine S, Mirman D, Middleton EL, Buxbaum LJ. Temporal dynamics of activation of thematic and functional knowledge during conceptual processing of manipulable artifacts. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2012;38(5):1274–1295. doi: 10.1037/a0027626. http://doi.org/10.1037/a0027626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lambon Ralph MA, Jefferies E, Patterson K, Rogers TT. The neural and computational bases of semantic cognition. Nature Reviews Neuroscience. 2017:1–14. doi: 10.1038/nrn.2016.150. http://doi.org/10.1038/nrn.2016.150. [DOI] [PubMed]
- Landrigan J-F, Mirman D. Data from: “Taxonomic and Thematic Relatedness Ratings for 659 Word Pairs”. 2016 http://doi.org/http://dx.doi.org/10.7910/DVN/FKTQ4C.
- Lawson R, Chang F, Wills AJ. Free classification of large sets of everyday objects is more thematic than taxonomic. Acta Psychologica. 2017;172:26–40. doi: 10.1016/j.actpsy.2016.11.001. http://doi.org/10.1016/j.actpsy.2016.11.001. [DOI] [PubMed] [Google Scholar]
- Lee C, Middleton EL, Mirman D, Kalénine S, Buxbaum LJ. Incidental and context-responsive activation of structure- and function-based action features during object identification. Journal of Experimental Psychology Human Perceptual Performance. 2013;39(1):257–270. doi: 10.1037/a0027533. http://doi.org/10.1037/a0027533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu J, Golinkoff RM, Sak K. One cow does not an animal make: young children can extend novel words at the superordinate level. Child Development. 2001;72(6):1674–94. doi: 10.1111/1467-8624.00372. http://doi.org/10.1111/1467-8624.00372. [DOI] [PubMed] [Google Scholar]
- Lupyan G. Extracommunicative functions of language: verbal interference causes selective categorization impairments. Psychonomic Bulletin & Review. 2009;16(4):711–718. doi: 10.3758/PBR.16.4.711. http://doi.org/10.3758/PBR.16.5.986. [DOI] [PubMed] [Google Scholar]
- Lupyan G, Mirman D. Linking language and categorization: Evidence from aphasia. Cortex. 2013;49(5):1187–1194. doi: 10.1016/j.cortex.2012.06.006. http://doi.org/10.1016/j.cortex.2012.06.006. [DOI] [PubMed] [Google Scholar]
- Marian V, Bartolotti J, Chabal S, Shook A. CLEARPOND: cross-linguistic easy-access resource for phonological and orthographic neighborhood densities. PloS One. 2012;7(8) doi: 10.1371/journal.pone.0043230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McNorgan C, Reid J, McRae K. Integrating conceptual knowledge within and across representational modalities. Cognition. 2011;118:211–233. doi: 10.1016/j.cognition.2010.10.017. http://doi.org/10.1016/j.cognition.2010.10.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McRae K, Jones M. Semantic Memory. The Oxford Handbook of Cognitive Psychology. 2013 Retrieved from http://books.google.co.il/books?id=ktFMAgAAQBAJ.
- Merck C, Jonin PY, Laisney M, Vichard H, Belliard S. When the zebra loses its stripes but is still in the savannah: Results from a semantic priming paradigm in semantic dementia. Neuropsychologia. 2014;53(1):221–232. doi: 10.1016/j.neuropsychologia.2013.11.024. http://doi.org/10.1016/j.neuropsychologia.2013.11.024. [DOI] [PubMed] [Google Scholar]
- Meyer K, Damasio A. Convergence and divergence in a neural architecture for recognition and memory. Trends in Neurosciences. 2009;32(7):376–382. doi: 10.1016/j.tins.2009.04.002. http://doi.org/10.1016/j.tins.2009.04.002. [DOI] [PubMed] [Google Scholar]
- Mirman D, Britt AE. What we talk about when we talk about access deficits. Philosophical Transactions of the Royal Society B: Biological Sciences. 2014;369(1634):1–14. doi: 10.1098/rstb.2012.0388. http://doi.org/10.1098/rstb.2012.0388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mirman D, Graziano KM. Individual differences in the strength of taxonomic versus thematic relations. Journal of Experimental Psychology: General. 2012;141(4):601–609. doi: 10.1037/a0026451. http://doi.org/10.1037/a0026451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mirman D, Landrigan JF, Britt AE. Taxonomic and Thematic Semantic Systems. Psychological Bulletin. 2017;143(5):499–520. doi: 10.1037/bul0000092. http://doi.org/10.1037/bul0000092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mirman D, Magnuson JS. Attractor dynamics and semantic neighborhood density: Processing is slowed by near neighbors and speeded by distant neighbors. Journal of Experimental Psychology. Learning, Memory, and Cognition. 2008;34(1):65–79. doi: 10.1037/0278-7393.34.1.65. http://doi.org/10.1007/s11103-011-9767-z.Plastid. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Connor CM, Cree GS, McRae K. Conceptual hierarchies in a flat attractor network: Dynamics of learning and computations. Cognitive Science. 2009;33(4):665–708. doi: 10.1111/j.1551-6709.2009.01024.x. http://doi.org/10.1111/j.1551-6709.2009.01024.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patterson K, Nestor PJ, Rogers TT. Where do you know what you know? The representation of semantic knowledge in the human brain. Nature Reviews. Neuroscience. 2007;8(december):976–987. doi: 10.1038/nrn2277. http://doi.org/10.1038/nrn2277. [DOI] [PubMed] [Google Scholar]
- Pecher D, Zeelenberg R, Barsalou LW. Verifying different madality properties for concepts produces swtiching costs. Psychological Science. 2003;14(2):119–124. doi: 10.1111/1467-9280.t01-1-01429. [DOI] [PubMed] [Google Scholar]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing; Vienna, Austria: 2014. URL: http://www.R-project.org/ [Google Scholar]
- Rabovsky M, McRae K. Simulating the N400 ERP component as semantic network error: Insights from a feature-based connectionist attractor model of word meaning. Cognition. 2014;132(1):68–89. doi: 10.1016/j.cognition.2014.03.010. http://doi.org/10.1016/j.cognition.2014.03.010. [DOI] [PubMed] [Google Scholar]
- Rogers TT, Lambon Ralph Ma, Hodges JR, Patterson K. Natrual Selection: The Impact of Semantic Impairment on Lexical and Object Decision. Cognitive Neuropsychology. 2004;21:331–352. doi: 10.1080/02643290342000366. http://doi.org/10.1016/j.limno.2013.04.005. [DOI] [PubMed] [Google Scholar]
- Rogers TT, Lambon Ralph Ma, Garrard P, Bozeat S, McClelland JL, Hodges JR, Patterson K. Structure and deterioration of semantic memory: a neuropsychological and computational investigation. Psychological Review. 2004;111(1):205–235. doi: 10.1037/0033-295X.111.1.205. http://doi.org/10.1037/0033-295X.111.1.205. [DOI] [PubMed] [Google Scholar]
- Rogers TT, Patterson K. Object categorization: reversals and explanations of the basic-level advantage. Journal of Experimental Psychology: General. 2007;136(3):451–469. doi: 10.1037/0096-3445.136.3.451. http://doi.org/http://doi.org/10.1037/0096-3445.136.3.451. [DOI] [PubMed] [Google Scholar]
- Schwartz MF, Kimberg DY, Walker GM, Brecher A, Faseyitan OK, Dell GS, … Coslett HB. Neuroanatomical dissociation for taxonomic and thematic knowledge in the human brain. Proceedings of the National Academy of Sciences of the United States of America. 2011;108:8520–8524. doi: 10.1073/pnas.1014935108. http://doi.org/10.1073/pnas.1014935108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Semenza C, Bisiacchi PS, Romani L. Naming disorders and semantic representations. Journal of Psycholinguistic Research. 1992;21(5):349–364. doi: 10.1007/BF01067920. http://doi.org/10.1007/BF01067920. [DOI] [PubMed] [Google Scholar]
- Semenza C, Denes G, Lucchese D, Bisiacchi P. Selective deficit of conceptual structures in aphasia: class versus thematic relations. Brain and Language. 1980;10(2):243–248. doi: 10.1016/0093-934x(80)90054-1. http://doi.org/10.1016/0093-934X(80)90054-1. [DOI] [PubMed] [Google Scholar]
- Simmons S, Estes Z. Individual differences in the perception of similarity and difference. Cognition. 2008;108(3):781–795. doi: 10.1016/j.cognition.2008.07.003. http://doi.org/10.1016/j.cognition.2008.07.003. [DOI] [PubMed] [Google Scholar]
- Wilson MD. The MRC Psycholinguistic Database: Machine Readable Dictionary, Version 2. Behavior ResearchMethods, Instruments, & Computers. 1988;20:6–11. [Google Scholar]
- Wisniewski EJ, Bassok M. What makes a man similar to a tie? Stimulus compatibility with comparison and integration. Cognitive Psychology. 1999;39:208–238. doi: 10.1006/cogp.1999.0723. http://doi.org/10.1006/cogp.1999.0723. [DOI] [PubMed] [Google Scholar]
- Yee E, Thompson-Schill SL. Putting Concepts into Context. Psychonomic Bulletin & Review. 2016 Jun;:1015–1027. doi: 10.3758/s13423-015-0948-7. http://doi.org/10.3758/s13423-015-0948-7. [DOI] [PMC free article] [PubMed]