

ABSTRACT
Dispersal is a process of central importance for the ecological and evolutionary dynamics of populations and communities, because of its diverse consequences for gene flow and demography. It is subject to evolutionary change, which begs the question, what is the genetic basis of this potentially complex trait? To address this question, we (i) review the empirical literature on the genetic basis of dispersal, (ii) explore how theoretical investigations of the evolution of dispersal have represented the genetics of dispersal, and (iii) discuss how the genetic basis of dispersal influences theoretical predictions of the evolution of dispersal and potential consequences.
Dispersal has a detectable genetic basis in many organisms, from bacteria to plants and animals. Generally, there is evidence for significant genetic variation for dispersal or dispersal‐related phenotypes or evidence for the micro‐evolution of dispersal in natural populations. Dispersal is typically the outcome of several interacting traits, and this complexity is reflected in its genetic architecture: while some genes of moderate to large effect can influence certain aspects of dispersal, dispersal traits are typically polygenic. Correlations among dispersal traits as well as between dispersal traits and other traits under selection are common, and the genetic basis of dispersal can be highly environment‐dependent.
By contrast, models have historically considered a highly simplified genetic architecture of dispersal. It is only recently that models have started to consider multiple loci influencing dispersal, as well as non‐additive effects such as dominance and epistasis, showing that the genetic basis of dispersal can influence evolutionary rates and outcomes, especially under non‐equilibrium conditions. For example, the number of loci controlling dispersal can influence projected rates of dispersal evolution during range shifts and corresponding demographic impacts. Incorporating more realism in the genetic architecture of dispersal is thus necessary to enable models to move beyond the purely theoretical towards making more useful predictions of evolutionary and ecological dynamics under current and future environmental conditions. To inform these advances, empirical studies need to answer outstanding questions concerning whether specific genes underlie dispersal variation, the genetic architecture of context‐dependent dispersal phenotypes and behaviours, and correlations among dispersal and other traits.
Keywords: dispersal kernel, eco‐evolutionary models, gene flow, genetic architecture, genotype–environment interactions, heritability, life‐history traits, migration, mobility, movement
I. INTRODUCTION
Dispersal is any movement of individuals or propagules that has potential consequences for gene flow across space (Ronce, 2007; Appendix I). Dispersal has a central role in life history and its evolution is fundamental in determining the consequences of land use change, habitat degradation, and climate change for species persistence, or a species' invasive potential (Clobert et al., 2012; Travis et al., 2013a). Whether or not, and how far, an organism moves between the place of its birth and reproduction can be the target of intense selection (Ronce, 2007). However, any evolutionary change in dispersal ability will occur only if the underlying traits (see Appendix I) have a heritable component (Falconer & Mackay, 1996; Lynch & Walsh, 1998; Danchin et al., 2011). The speed of the evolutionary change will depend on the underlying genetic architecture (Orr, 2005; Richards, Bossdorf & Pigliucci, 2010; MacKay et al., 2012), the dispersal trait's genetic variance, and its covariance with other traits that may be under selection (Appendix I).
Dispersal is a complex and multidimensional phenotype, which makes describing its genetic basis highly challenging (see online Table S1). Dispersal comprises departure (initiation to leave the natal habitat), transfer (movement), and settlement (establishment in the novel breeding habitat), and often involves multiple morphological, physiological and behavioural traits (Clobert et al., 2009). Further, departure and transfer can be actively undertaken by the organism or vectored by other abiotic or biotic agents, and in most cases even vectored dispersal is influenced by morphological, physiological or behavioural adaptations (Bonte et al., 2012), potentially leading to ‘dispersal syndromes’ (Appendix I) (Clobert et al., 2009). In actively dispersing organisms, an individual's ability or capacity to move is often related to its morphology, size or endurance, although high movement abilities only lead to dispersal when individuals have a propensity to disperse. Dispersal may be the result of a ‘dedicated behaviour’, the main function of which is to disperse; or it may occur as a by‐product of other movements, such as foraging or mate searching (Van Dyck & Baguette, 2005; Burgess et al., 2015; Bonte & Dahirel, 2017). While some species show a recognizable locomotory or behavioural ‘dispersal morph’, which allows a more accurate quantification of dispersal [see Zera & Denno, 1997 for a review], in most species dispersal traits show continuous phenotypes, and hence are more difficult to quantify. Passive dispersal has the added complexity of dispersers being vectored by agents such as animals, wind, water or gravity, with the eventual dispersal process resulting from the interaction between the vector and individual traits (Cousens, Dytham & Law, 2008). Seed dispersal by wind, for example, depends on the overall size of the plant, including height of seed release, and on the seed size and morphology (e.g. presence of plumes or wings; Soons et al., 2004; Thomson et al., 2010). Seed dispersal by animals is affected by traits such as plant height and the seed's adhesive structures (Albert et al., 2015), while fruit placement, size, colour and nutritional content affect their attractiveness to potential frugivorous dispersers (Russo, 2003; Lomáscolo et al., 2010). Passive dispersal and phoretic (transport of one species by another) behaviour is also found in animal taxa (e.g. Houck & O'Connor, 1991). Whether active or passive, dispersal is often correlated with other key life‐history characteristics, such as morphological characteristics or reproductive traits (Stevens et al., 2012, 2014; Bonte & Dahirel, 2017), resulting in the potential for trait co‐evolution if the correlations are genetic. Additionally, environmental conditions can strongly impact dispersal. For example, dispersal propensity may be affected by population density (Bowler & Benton, 2005; Matthysen, 2005), and wind force affects seed dispersal distances (Greene, 2005). Understanding whether different genotypes are differently influenced by environmental conditions – i.e. genotype–environment (G × E) interactions (Appendix I) – is also important (e.g. Donohue, Polisetty & Wender, 2005; Sinervo et al., 2006; Ducatez et al., 2012; Pennekamp et al., 2014; Cote et al., 2017a).
How do evolutionary models of dispersal deal with this complexity? There are two main categories of dispersal models: theoretical ecological and/or evolutionary models that aim at a general understanding of the causes and consequences of dispersal, and taxon‐specific predictive models. While the vast majority of models of dispersal fall into the former category, the latter approach is mostly used for tactically describing movement in a specific ecological context (e.g. spread of invasive species, species range shift; see online Table S2 and do not usually consider evolution. In models that do consider evolution, the genetic basis of dispersal is usually kept as simple as possible: a typical assumption is a single locus affecting a single, broadly defined trait related to dispersal (e.g. dispersal propensity, shape of the dispersal kernel, environmentally cued rules for individual departure and settling; see online Table S2). Hence, a critical question is whether the accumulation of empirical information about genetic architectures for dispersal should result in new approaches to modelling dispersal evolution, and whether this increased complexity matters.
Our goal in this review is twofold. First, even though several reviews already exist on different aspects of the genetics of dispersal (e.g. Gatehouse, 1989; Wheat, 2012; Zera & Brisson, 2012; Dingle, 2014; see also reviews on the genetics of other movement types: Pulido, Berthold & van Noordwijk, 1996; van Noordwijk et al., 2006; Pulido, 2007; Dingle, 2014), we review current knowledge across different taxa. We focus on inferences of the genetic basis of dispersal using direct estimates of their heritability or population genetic variance, comparisons of dispersal traits among populations, and genetic correlations/covariances between dispersal traits (Appendix I) and other traits. Our perspective is on the genetic architecture of dispersal components, which has not been reviewed previously, and we review the few cases in which loci influencing dispersal have been identified. Second, we review current theory on dispersal evolution. Specifically, we review how genetics have been accounted for in existing models, which to date have almost exclusively been developed to provide qualitative, theoretical predictions on dispersal evolution, and we discuss how predicted evolutionary outcomes may depend on assumptions about dispersal genetic architecture. Finally, we present results from a new model that we developed to illustrate how the evolution of dispersal probability in a fragmented landscape depends upon the underlying genetic architecture.
II. EMPIRICAL EVIDENCE FOR THE GENETIC BASIS OF DISPERSAL
We carried out an extensive literature search using the Web of Science (WOS) database (see online Appendix S1: Methods). We targeted not only dispersal traits sensu stricto (such as distance covered or propensity to leave a patch), but also dispersal‐related traits such as locomotory capacity or wing morphology that may serve as proxies for dispersal. We selected papers from the search results on the criteria that the study dealt strictly with (i) assessing the existence of the genetic basis/variation of dispersal traits, or (ii) estimating the heritability values of dispersal and related traits, or (iii) determining their genetic architecture. We moreover reviewed studies on evolutionary responses of dispersal traits to environmental changes or on population differentiation in dispersal. We also collected information on whether dispersal traits are correlated with each other or with other traits, whether due to chromosomal linkage or pleiotropy (Appendix I), with the aim to determine whether dispersal can be a direct target of natural selection, whether it can evolve via selection on correlated traits, or both.
(1). Heritability of dispersal and dispersal‐related traits
The most straightforward approach to investigating the genetic basis of phenotypes is to measure their heritability (h 2; Appendix I). In animals, heritability of dispersal‐related traits has been estimated in diverse taxa, from aquatic invertebrates to mammals (see online Table S1). However, as pointed out in previous reviews of dispersal (e.g. Zera & Brisson, 2012; Dingle, 2014), the literature on heritability estimates remains somewhat unbalanced, with most studies focusing on birds and insects. For mammals, fish, amphibians and reptiles, few studies are available, and information on invertebrates other than insects was almost absent. Heritability estimates of dispersal‐related traits in the two best‐represented animal taxa (birds and insects) range from 0 to 1, with average values of 0.35 for both. Most studies – with the exception of those on birds – did not quantify dispersal distance or propensity per se but used proxies related to movement ability during the transfer stage of dispersal, such as flight capacity in insects, and speed or locomotion performance in fish and reptiles (e.g. Garland, 1990; Robertson et al., 2002; Zera & Brisson, 2012; Drangsholt, Damsgard & Olesen, 2014; Mattila & Hanski, 2014; see online Table S1). Morphological traits are also often used as proxies of dispersal. Examples of such traits in insects include thorax ratio, wing shape, and wing muscle strength (e.g. Van Dyck, Matthysen & Wiklund, 1998; Roff & Bradford, 1998; Keena, Grinberg & Wallner, 2007; Zera & Brisson, 2012; see online Table S1).
Very few animal studies have estimated the heritability of traits related to stages of dispersal other than transfer. A notable exception is pre‐dispersal behaviour in arachnid species (Bonte, Bossuyt & Lens, 2007; Nachappa et al., 2010; see online Table S1). Most other estimates of heritability that are related to departure or settlement stages are from bird species, with h 2 reported for the propensity to leave the natal area, distance moved to the breeding site, and/or habitat choice in the settlement phase. We found 11 examples in bird species with estimates of h 2 significantly greater than zero in at least one of these traits, and five in which h 2 was reported to be non‐significant (see online Table S1). Examples of very high estimates are h 2 = 0.95 for propensity to leave the natal population in western bluebirds, Sialia mexicana, based on an Animal Model analysis (Duckworth & Kruuk, 2009), and h 2 = 0.86 for natal dispersal estimated from mother–daughter regression in the wandering albatross, Diomedea exulans (Charmantier et al., 2011). Other studies report modest heritabilities, such as h 2 = 0.15 for dispersal distance in great tits, Parus major (Korsten et al., 2013). Few studies have published heritability estimates for traits related to the settlement phase of dispersal (see online Table S1). Most that do pertain to conspecific attraction (Møller, 2002; Serrano & Tella, 2007; Roche, Brown & Brown, 2011), and one pertains to heterospecific attraction (Wheelwright, Lawler & Weinstein, 1997).
In plants, significant among‐family variance and heritability have been documented in a number of dispersal‐related traits, mostly in annual herbs and crop species (see online Table S1). Some species of the Brassicaceae family, such as the rapeseed (Brassica napus) show high levels of heritability in pod shatter resistance that varied from h 2 = 0.85 to 0.92 (Liu et al., 2016). Pod shatter resistance is a trait under strong selection in cereal crops, such as rice (Oryza sativa) (Konishi et al., 2006) and soybean (Glycine max (L.) Merr.) (Dong et al., 2014) because it prevents seed dispersal and, therefore, the loss of grains in terms of yield. High heritability values were also recorded for a diverse array of traits in the spikelets of a grass species Brachypodium hybridum (see online Table S1). On average these traits showed h 2 = 0.54, with values ranging from 0.86 (average number of seeds per spikelet) to 0 (average length of inflorescence) (Neji et al., 2015). The heritability of dispersal‐related traits in wild species has been scarcely documented, but studies show that broad‐sense heritability of achene and capitulum traits range from h 2 = 0.4 to 0.8 in Heterosperma pinnatum (Venable & Burquez, 1989) and that the narrow‐sense heritability of the proportion of achenes that disperse in Crepis sancta is greater than h 2 = 0.2 (Imbert, 2001). The narrow‐sense heritability of dispersal‐related traits measured in the panicle of the canary grass (Phalaris aquatic), an invasive species in native grassland across California, show values above h 2 = 0.5 (Kelman & Culvenor, 2003). Another example of a passively dispersed species shows that heritability is environment‐dependent. For example, in Arabidopsis thaliana, Donohue et al. (2005) quantified broad‐sense heritability estimates of fruit and plant architectural traits associated with dispersal, based on genetic differences among ecotypes. The values ranged from h 2 = 0 to 0.7, depending on the trait and the environment in which the trait was measured. The heritability of aspects of seed dispersal patterns per se under controlled conditions ranged from h 2 = 0.01 to 0.4 (Donohue et al., 2005), and were strongly dependent on how dispersal‐related traits of maternal plants responded to density (Wender, Polisetty & Donohue, 2005). Furthermore, some studies that investigate seed dispersal by tropical vertebrates have shown moderate values of broad‐sense heritability in natural populations. Wheelwright (1993) reported h 2 = 0.6 for seed diameter in the sweetwood tree (Ocotea tenera), and Galetti et al. (2013) quantified h 2 = 0.35 for seed size in Euterpe edulis, a keystone palm species.
What have we learned from these studies? While there are many examples of heritability estimates for putative proxies of dispersal and movement ability in general, heritability estimates for specific traits involved in the entire process of dispersal, such as propensity to leave the natal patch or distance travelled between sites, are very rare and would be important to target for future studies. Birds are the most broadly studied taxon in this regard, as the heritability of dispersal has been studied in multiple species in free‐living conditions. These studies are based on marked individuals of known relatedness, in one case combined with in situ cross‐fostering (Roche et al., 2011). Until recently most studies were based on parent–offspring or sibling similarities, but several analyses have now been published based on Animal Models (Appendix I) that make use of complete pedigree information (Duckworth & Kruuk, 2009; Korsten et al., 2010; Charmantier et al., 2011; Doligez et al., 2012). Parent–offspring and sib–sib analyses typically yield higher heritability estimates than Animal Models (McCleery et al., 2004; Charmantier et al., 2011; Doligez et al., 2012), due to maternal and common environment effects (van Noordwijk et al., 2006; Zera & Brisson, 2012). Further, heritability estimates may also be inflated by biases in detectability due to limited study areas (van Noordwijk, 1984; Doligez & Pärt, 2008) and to more numerous social interactions among siblings synchronizing their movements (Matthysen, Van de Casteele & Adriaensen, 2005; Cote et al., 2017b). As a whole, heritability studies suggest that variation in dispersal can arise as a result of additive genetic variation, but also that in many cases much of the phenotypic variation is explained by environmental variation, including transgenerational effects (Appendix I). Recent work, for instance, has demonstrated strong epigenetic signals in dispersal propensity (van Petegem et al., 2015). Furthermore, similarity in dispersal phenotypes among related individuals may be caused by (maternally transmitted) symbionts or parasites (Goodacre et al., 2009; Debeffe et al., 2014).
(2). Genetic architecture of dispersal
A fundamental question regarding the evolution of any trait is whether the trait's heritable variation arises from a single gene, a few genes (each of large effect, oligogenic), or many genes (each of very small effect, polygenic; Appendix I) (Mackay, Stone & Ayroles, 2009; Rockman, 2012; Zera & Brisson, 2012). While research has revealed some loci of major effect affecting a range of quantitative traits (reviewed in Slate, 2005; Schielzeth & Husby, 2014), more recent work using linkage analysis or association analysis [genome‐wide association study (GWAS); Appendix I] indicates that many quantitative traits are polygenic (MacKay et al., 2012; Husby et al., 2015; Santure et al., 2015; Kardos et al., 2016). The form of the genetic architecture has significant impacts on evolutionary dynamics, and this will be discussed in more detail in Section III. Below we review studies that have attempted to characterize loci associated with dispersal and the magnitude of their effects, as well as some studies that have identified genes associated with dispersal.
Pea aphid, Acyrthosiphon pisum, offers the most obvious example of pronounced genetic determination of dispersal, as male wing polymorphism is determined by a single locus on the X‐chromosome (Caillaud et al., 2002). Beyond this example, we lack a robust mechanistic insight into loci with large dispersal effects for most organisms. In Drosophila melanogaster studies of variation affecting locomotion behaviour provide us with a list of potential candidate genes for dispersal (Jordan et al., 2007, 2012) (Appendix I), and these genes may also regulate dispersal in other organisms [see also reviews by Zera & Brisson, 2012 and by Wheat, 2012]. One of the first genes identified to influence both larval and adult locomotion behaviour was the foraging gene (Pereira & Sokolowski, 1993), where larvae with the dominant ‘rover’ allele (for R) move further than individuals homozygous for the ‘sitter’ allele (for S) in the presence of food (Edelsparre et al., 2014). Importantly, adult dispersal distance in the field is also higher for individuals with the for R allele (Edelsparre et al., 2014). In both cases, detailed functional genomic validation has demonstrated that these effects arise from variation in expression of the gene itself. A transcriptome study of long‐wing and short‐wing artificially selected lines of the cricket Gryllus firmus identified a number of large‐magnitude differences in expression of genes that function in flight and reproduction (Nanoth Vellichirammal et al., 2014; reviewed in Zera, 2016). These data on genetic covariances in expression of genes related to flight and reproduction are also relevant to the discussion of genetic correlations (see Section II.5). By developing a state‐of‐the‐art phenotyping platform, a further recent study of the cotton bollworm Helicoverpa armigera showed that a suite of expressed candidate genes linked to physiological adaptations are required for long‐distance flight, allowing this insect to respond to deteriorating conditions (Jones et al., 2015).
Indications of oligogenic architectures related to dispersal have also been uncovered in other organisms. Allelic variation in the gene responsible for the enzyme phosphoglucose isomerase (Pgi), which is associated with dispersal rate in the Glanville fritillary butterfly (Melitaea cinxia), is probably the most thoroughly studied case (reviewed by Niitepõld & Saastamoinen, 2017). Pgi is a metabolic enzyme responsible for cellular energetics (e.g. Mattila & Hanski, 2014). The butterfly forms a metapopulation in the Åland Islands, Finland, and more‐dispersive individuals are known to establish new populations (Hanski et al., 2004). This is especially true in habitat patches that are more isolated from established populations. The frequency of a specific allele of the Pgi gene is highest in these newly established, isolated populations (Haag et al., 2005). In addition, individuals with this allele have a higher flight metabolic rate as well as higher dispersal propensity in the field (Haag et al., 2005; Niitepõld et al., 2009, 2011). Allelic variations are under balancing selection according to their associated higher individual fitness under different environmental conditions (Saastamoinen, 2008; Klemme & Hanski, 2009; Orsini et al., 2009; Saastamoinen, Ikonen & Hanski, 2009; Wheat et al., 2010). These changes in Pgi allele frequencies translate into phenotypic variation in dispersal, and hence influence demographic rates in ways that impact population dynamics (Saccheri & Hanski, 2006; Hanski, 2011). Recently, alleles of a second gene, succinate dehydrogenase D (sdhd), have been found to show epistatic interaction with Pgi (Wheat et al., 2011). sdhd affects flight endurance through oxygen delivery (Marden et al., 2003) and as for Pgi, sdhd alleles also differ in frequency between new and established populations within the metapopulation (Wheat et al., 2011). It is notable, however, that detailed functional genomic validation of Pgi has not been demonstrated, and hence the causal role of Pgi as a driver of the above‐mentioned processes has not been proven [see Niitepõld & Saastamoinen, 2017 and Wheat & Hill, 2014 for more details].
All studies examining the genetic basis of dispersal in natural populations in animals have so far either used a candidate gene approach (e.g. the Pgi studies described above) or a limited number of markers (all other studies than the Pgi studies described above). The low resolution of marker densities, often in combination with a small number of genotyped individuals, means that the power to detect genetic variants associated with dispersal is modest at best. One way to circumvent such issues is to study dispersal in model systems, for instance using the Drosophila Genetic Reference Panel (Mackay et al., 2012), which consists of more than 200 fully sequenced inbred lines derived from North Carolina (Mackay et al., 2012), and thus offers genomic tools for understanding the genetic basis of complex traits with high precision and statistical power. Jordan et al. (2012) investigated the genomic regions affecting locomotion variation across 167 lines of D. melanogaster. Using nearly 2.5 million single nucleotide polymorphisms (SNPs) to explain inter‐line variation in locomotion, they found 220 SNPs (Appendix I) in 192 genes associated with this phenotype. Of these, 12 SNPs explained about 60% of the variation observed in both sexes. Importantly, they found a negative relationship between the effect size of individual SNPs and their frequency in the sample as a whole, indicating that rare alleles had the largest effects. Similar results appear to be emerging from a recent study in the M. cinxia butterfly's metapopulation (Fountain et al., 2016). In this study, individual butterflies were genotyped for 222 SNPs across the genome, including SNPs from candidate genes and neutral regions. Fifteen SNP loci, mostly associated with flight, changed in frequency in a non‐random way in response to habitat fragmentation (Fountain et al., 2016). These loci were identified in comparisons of new versus old populations and therefore are likely enriched in rare alleles with respect to the entire metapopulation.
In plants, the best‐studied examples of genes involved in dispersal come from domesticated species and model genetic organisms. In domesticated species, research has focused on dispersal features such as seed shattering in cereals (Konishi et al., 2006), and fruit/pod shattering (dehiscence) of multi‐seeded fruits such as pods in legumes and siliques in the mustard family, Brassicaceae (Raman et al., 2014) or on the proportion of central achenes produced per capitulum of some species of the Asteraceae family (Imbert, 2001). Depending on the species and population studied, genetic analysis of pod shattering has documented between one and 13 quantitative trait loci (QTLs) or causative loci associated with this trait (Konishi et al., 2006; Raman et al., 2014; reviewed by Li & Olsen, 2016).
In contrast to the above examples, where the small number of loci associated with dispersal variation suggests an oligogenic genetic architecture, numerous studies (outside the field of dispersal) indicate that quantitative traits generally arise via a polygenic architecture (Mackay et al., 2012; Husby et al., 2015; Santure et al., 2015). While the relatively few examples of genes with large effect size could arise from underpowered QTL or GWA studies, it could also indicate the absence of such loci. Indeed, it has been argued that publishing bias is responsible for inflating the frequency of known oligogenic examples relative to their true general relevance for evolution in the wild (Rockman, 2012). The multi‐factorial nature of dispersal is frequently manifested as quantitative phenotypic variation in dispersal and in traits that contribute to dispersal. On the whole, there is good evidence for genetic variation in dispersal in natural populations, and evidence from other complex life‐history traits suggests that this variation is seldom caused by single genes of large effect. Instead, multiple loci probably combine to affect dispersal. Further studies, using genome‐wide approaches, are required to confirm this hypothesis.
Why is knowing the number of genes contributing to dispersal variation relevant? One reason is that it may influence patterns of convergent evolution. With a small number of genes involved in trait expression, the same genes may evolve in multiple taxa to create similar phenotypes. For example, orthologues of a number of candidate genes identified in D. melanogaster also appear to regulate dispersal in other insects (Wheat, 2012). In plants, orthologues of a major gene that controls pod shattering INDEHISCENT (IND), are expressed in diverse species in the mustard family (Brassicaceae); in Lepidium campestre, which has dehiscent fruits, expression patterns of the dehiscence‐pathway genes were similar to that in Arabidopsis thaliana (Lenser & Theißen, 2013). Homologues of IND have been found even outside of the Brassicaceae (Gioia et al., 2013). Another reason is that deciphering the genetic architecture of dispersal may provide information on the speed at which this phenotype may evolve in response to environmental changes (Hoffmann & Willi, 2008). For example, the existence of complex epistatic interactions (Appendix I) between genes involved in quantitative traits can decrease the amount of genetic variation at these genes compared to an additive architecture (Hansen, 2006). This means that a complex epistatic architecture of dispersal may limit its evolutionary potential in the context of environmental changes because of a limited pool of available genetic variants on which selection can act.
(3). Artificial selection and experimental evolution of dispersal
Besides the approaches detailed above that characterize genetic variation for dispersal, indirect evidence for the genetic basis of dispersal can be provided through studies that elicit evolutionary responses directly by imposing selection on dispersal phenotypes. Experimental evolution and artificial selection can be used to estimate heritability (e.g. Jordan et al., 2007; Kawecki et al., 2012), but they can also directly demonstrate the evolutionary potential of a trait. Experimental evolution allows evolutionary changes of dispersal in experimental populations subjected to specific conditions while artificial selection exerts selection on dispersal traits alone by breeding partners chosen for their dispersal phenotype (Zera & Brisson, 2012). This type of approach is hence very powerful but its application is generally limited to species that can easily be reared in the laboratory.
Part of the current knowledge of the evolutionary potential of dispersal comes from artificial selection on domesticated species, particularly plants, and on model micro‐organisms. Decreased seed dispersal has been subject to strong artificial selection in diverse domesticated plant species (Li & Gill, 2006; Weeden, 2007; Meyer & Purugganan, 2013; Mach, 2015), and reversion to dispersing forms has been documented in naturalized crop species (De Wet & Harlan, 1975). Based on genetic analyses of domestication traits, Fuller & Allaby (2009) inferred that domestication first imposed selection on seed size and germination even before agriculture was widely practiced, and as crops became cultivated, selection for lack of dispersal followed. Similarly in bacteria, experimental evolution and selection experiments show that bacterial motility, as well as sporulation, are evolvable traits (Girgis et al., 2007; Brown et al., 2011; Koskella et al., 2011; Taylor & Buckling, 2011). Mobility was also successfully selected for in the nematode Caenorhabditis elegans (Friedenberg, 2003), in a protist (Fronhofer & Altermatt, 2015), and in different mite species (Gu & Danthanarayana, 1992; Knülle, 1995; Nachappa et al., 2010; Bal, Michel & Grewal, 2014).
Results of selection experiments on dispersal traits are however not always straightforward, as exemplified by the case of the spider mite Tetranychus urticae. In this species, aerial dispersal was initially found to be highly heritable in a narrow sense (Li & Margolies, 1993, 1994) but recently, this trait has been discovered to be driven by epigenetic control (van Petegem et al., 2016). Yano & Takafuji (2002) reported a response to selection, while Tien, Sabelis & Egas (2011) did not detect any response for this trait. A third trait, dispersal distance, was not amenable to selection due to substantial intergenerational plasticity (Bitume et al., 2011, 2015). Experimental landscapes that differed in the spatial correlation of patch extinctions were found to cause evolution of dispersal in the spider mite, with increased spatial correlation selecting for less emigration (departure) but more long‐distance dispersal (Fronhofer et al., 2014). Further work using experimental metapopulations showed that increased spatial variation in habitat availability reduced dispersal rates in terms of departure from patches (De Roissart, Wang & Bonte, 2015; De Roissart et al., 2016). Although the various traits involved in dispersal contrasted in their response to selection, experimental evolution in the spider mite has resulted in some responses to selection, demonstrating both a genetic basis of dispersal and existing variation for dispersal within sampled populations.
(4). Phenotypic variation in dispersal among populations
If dispersal has a genetic basis and can evolve, divergent selection on dispersal should produce predictable differences in dispersal phenotypes among different types of populations. While Section II.3 documented the evolvability of dispersal, here we address an extension to that concept, focusing on the evolution of dispersal differences among populations, and its role in linking ecological and evolutionary dynamics. It is notable, however, that differences between populations in any trait, even if in accordance with predictions, are not in itself evidence for genetic variation, and should always be accompanied by common‐garden breeding trials.
Some of the best‐known examples indicating micro‐evolution of dispersal come from range‐expanding species (e.g. Thomas et al., 2001; Lindström et al., 2013; Cheptou et al., 2017; Cote et al., 2017b) and from metapopulations showing contrasting levels of connectivity. Individuals from range margins might be predicted to evolve a higher dispersal propensity than core populations, and newly founded populations are likely to comprise more dispersive individuals than old populations in a metapopulation. In fact, both the evolution of increased or decreased dispersal can be predicted for species expanding over fragmented landscapes or experiencing a sudden fragmentation event (Leimar & Norberg, 1997 ; Heino & Hanski, 2001 ; Gyllenberg, Parvinen & Dieckmann, 2002; Travis, Smith & Ranwala, 2010b; Poethke, Gros & Hovestadt, 2011b ; Cote et al., 2017a). On the one hand, as habitat fragmentation imposes diverse costs during transfer (Bonte et al., 2012; Baguette & Van Dyck, 2007), we can simply expect a reduction in dispersal propensity (reviewed in Cote et al., 2017b). In the bog fritillary butterfly Boloria (Proclossiana) eunomia, dispersal propensity was dramatically decreased by landscape fragmentation, which induced higher dispersal mortality (Schtickzelle, Mennechez & Baguette, 2006; Schtickzelle et al., 2007). A reduced propensity to initiate wind‐assisted dispersal has been reported in spiders in fragmented landscapes (Bonte et al., 2006, 2007). The proportion of dispersing versus non‐dispersing achenes has been shown to differ among populations in desert annual herbs (Venable & Burquez, 1989), as well as among other species populations that differ in the degree of habitat fragmentation (Imbert, 2001; Riba et al., 2009). Isolated populations of the annual herb Crepis sancta along pavements in the city of Montpellier evolved less‐dispersive seeds within approximately 10 generations (Cheptou et al., 2008). Similarly, Mycelis muralis populations collected from fragmented landscapes had propagules with a lower terminal velocity (a proxy of dispersal ability), than those in non‐fragmented landscapes (Riba et al., 2009). Habitat fragmentation, on the other hand, may select for adaptations to improve transfer and settlement through, for example, movement and navigation abilities and, in highly fragmented landscapes, for higher emigration rates (Olivieri & Gouyon, 1997; Baguette & Van Dyck, 2007). This occurs because the increased habitat loss and local extinction probability increase the dispersal costs, but also raise the dispersal benefits gained through recolonizations (e.G. Olivieri & Gouyon, 1997; Travis & Dytham, 1999; see also the Glanville fritillary butterfly example in Section III.2d ). In some insects, more efficient transfer has been suggested to evolve in fragmented landscapes, for example by increased investments in thorax and flight muscle mass and by improved navigation skills (Thomas, Hill & Lewis, 1998; Merckx & van Dyck, 2007; Gibbs & van Dyck, 2010; Turlure, Schtickzelle & Baguette, 2010; Gomez & Van Dyck, 2012). Improved settlement capacities through increased perceptual ability are found in speckled wood butterflies Pararge aegeria in fragmented relative to more continuous habitat (Öckinger & Van Dyck, 2012). Whether dispersal will increase or decrease in response to habitat fragmentation will depend on factors such as the proportion of empty habitats to colonize, the degree of environmental heterogeneity and, importantly, the amount of genetic variation in dispersal traits (Cheptou et al., 2017; Cote et al., 2017b).
Comparisons of individuals from expanding range fronts with those from core populations also provide interesting insights about eco‐evolutionary dynamics (Appendix I) involved in dispersal evolution. For example, in northern Australia, research on the invasive cane toad Rhinella marina has documented behavioural and morphological changes associated with increased dispersal rate in populations at the expanding range front (Phillips et al., 2006, 2008). Common‐garden breeding trials have suggested a genetic basis to dispersal rate, as toads with parents from the range front have higher dispersal rates than toads with parents from the core populations (Phillips, Brown & Shine, 2010). A number of candidate genes linked to dispersal in cane toads have been identified recently (Rollins, Richardson & Shine, 2015). Similar results supporting an increased dispersal rate at the expansion front have been observed for other taxa including butterflies (Hill, Thomas & Blakeley, 1999; Hill et al., 2001; Thomas et al., 2001), damselflies (Therry, Bonte & Stoks, 2015), crickets (Thomas et al., 2001; Simmons & Thomas, 2004), aphids (Lombaert et al., 2014), pine trees (Cwynar & MacDonald, 1987), fishes (Myles‐Gonzalez et al., 2015) and some invasive plants (Monty & Mahy, 2010; Huang et al., 2015). These findings from field populations are closely paralleled by results from laboratory studies on range expansions in protists (Fronhofer & Altermatt, 2015; Fronhofer, Nitsche & Altermatt, 2017) and plants (Williams, Kendall & Levine, 2016). Increased seed dispersal at range edges compared to central locations has been observed in Senecio inaequidens (Monty & Mahy, 2010), but not in Senecio madagascariensis (Bartle, Moles & Bonsor, 2013), indicating that dispersal ability may evolve rapidly in response to natural selection, but that some species or populations may be limited in their ability to do so.
In summary, between‐population differentiation in dispersal may reflect a history of divergent selection on dispersal, imposed for example by habitat fragmentation or range expansions. Divergence among natural populations therefore provides evidence not only for a genetic basis of dispersal but also for its, sometimes rapid, evolutionary responses. However, the genetic architecture of dispersal traits has mostly been ignored from empiricists working on the eco‐evolutionary dynamics of dispersal. This is despite the fact that the speed of evolutionary changes and the likelihood that eco‐evolutionary dynamics occur likely involve a combination of standing genetic variation, new mutations and genetic covariances among traits (Reznick & Ghalambor, 2001; Etterson, 2004; Becks et al., 2010; Hendry, 2013).
(5). Genetic covariances among dispersal traits and between dispersal and other traits
As discussed above, dispersal is a multidimensional process made of different components (propensity to disperse, distance and direction of movements, and settlement choice), determined by various physiological, morphological and behavioural traits and induced by multiple ecological factors (reviewed in Zera & Brisson, 2012). Important questions are (i) how to characterize the complexity and the genetic drivers of dispersal, (ii) whether the multiple traits determining dispersal are genetically correlated, (iii) whether they are genetically correlated to other fitness‐related traits, and (iv) whether these correlations generally result from selection or genetically based traits or from plasticity (Clobert et al., 2009; Stevens et al., 2014; Cote et al., 2017a). The answer to these questions has important repercussions for the evolution of dispersal, as genetic covariances may either facilitate or constrain evolutionary changes (Futuyma, 2010; Kirkpatrick, 2010; Duputié et al., 2012). It is therefore surprising that although phenotypic correlations between dispersal and other phenotypic traits have been studied extensively in many organisms (reviewed in Cote et al., 2017b), studies on genetic correlations or covariances are relatively scarce.
The interrelationships between events at the different dispersal stages (Clobert et al., 2009) involve correlations between dispersal components. Some of these relationships are trivial, such as the emigration status intrinsically predicting the distance moved (i.e. non‐dispersing individuals moving null to short distances), and others are subtler, such as the correlation between the distance moved and the choice of settlement. Few theoretical studies incorporate these types of correlations: for example, the movement ability and the propensity to disperse may co‐vary, particularly in fragmented landscapes where the costs of transfer between habitats are high (Travis et al., 2012). Empirical studies, however, rarely measure more than one dispersal component or dispersal stage, and when they do, they rarely estimate correlations between them. Some studies focused on the transfer stage have, however, found that dispersal distance is often correlated with the direction or the linearity of movements and settlement behaviours (Selonen & Hanski, 2004; Merckx & van Dyck, 2007; Delgado & Penteriani, 2008; Delgado et al., 2010). Individuals dispersing over longer distances display straighter movements directed towards other habitats, while short‐distance dispersers wander around. These relationships at the phenotypic level can result from genetic correlations between dispersal components or from plasticity, a common environmental effect on all components or a third, unknown trait covarying with both components. To our knowledge, these alternative sources of covariation have not been tested so far.
Along with covariation among traits directly relevant to the dispersal process, dispersal components also covary with various other traits that improve the likelihood and success of dispersal (Clobert et al., 2009; Ronce & Clobert, 2012; Cote et al., 2017a). These correlations, defining dispersal syndromes (Appendix I), have been well studied at the species level (e.g. Zera & Brisson, 2012; Stevens et al., 2013, 2014; Cote et al., 2017a , and references therein), whereas information about the phenotypic structure, and occasionally about the genetic structure, at the individual level is only starting to accumulate. The most commonly observed correlation is that between dispersal and body size, body mass or derivatives (e.g. leg, wing or thorax size) in many animals (Bradford & Taylor, 1997; Zera & Denno, 1997; Hill et al., 1999; Lowe, Likens & Cosentino, 2006; Phillips et al., 2006; Cote & Clobert, 2010; Saastamoinen, Brakefield & Ovaskainen, 2012; Zera & Brisson, 2012; Trochet et al., 2013). Such relationships may arise because morphology directly affects the movement efficiency or the competitive ability of individuals, and therefore their dispersal success. Both dispersal traits and correlated morphological traits can be heritable (Thomas et al., 1998; Saastamoinen et al., 2012; Legrand et al., 2016), and it is therefore not surprising to see that their correlations also can have a genetic basis (e.g. Roff & Bradford, 1998; Lobón et al., 2011; Saastamoinen et al., 2012). However, when tested, these correlations sometimes have proven to be environmentally driven as well (Saastamoinen et al., 2012; Legrand et al., 2016).
Investment in morphology and the physiological machinery that supports dispersal may co‐vary with a number of other traits and processes. Arguably the most intensively studied species with regard to the physiological and genetic underpinnings of dispersal is the wing‐dimorphic cricket Gryllus firmus, which shows systematic differences between the long‐winged and short‐winged morphs (reviewed in Zera & Harshman, 2001; Zera & Brisson, 2012). Selection studies demonstrate that wing length is associated with differences in juvenile hormone and ecdysteroid levels (Zera, 2006). The morphs also differ in genetically controlled lipid synthesis and triglyceride accumulation (Zera & Larsen, 2001; Zera & Zhao, 2003), investment in the flight apparatus, mainly flight muscle mass (Zera & Denno, 1997; Zera & Harshman, 2001), and respiration rate (Crnokrak & Roff, 2002). In addition to morphology, dispersal syndromes also involve traits related to reproductive performance and hence demographic parameters (Zera & Brisson, 2012; Bestion, Clobert & Cote, 2015; Závorka et al., 2015). Here too, some of the best examples of genetic covariances between dispersal and reproductive traits are those from wing‐dimorphic crickets Gryllus firmus and G. veletis (Crnokrak & Roff, 1998; Roff, 1995; Begin & Roff, 2002; Zera & Brisson, 2012), in which a negative genetic correlation between dispersal and traits, such as embryo viability (Evans, García‐González & Marshall, 2007), fecundity (Roff & Bradford, 1998; Johnson, Johnson & Bradley Shaffer, 2010; Lobón et al., 2011) and ovary development (Zera & Brink, 2000; Roff & Gélinas, 2003) is evident. In a similar manner, in plants, genes that control plant architecture are expected to influence not only seed dispersal, but also floral presentation, ability to compete for light, or even size (Kirchoff & Claßen‐Bockhoff, 2013). Genes associated with plant height or size are likely to be associated with overall growth rates, competitive ability, and fitness itself (Donohue, 1998, 1999; Wender et al., 2005), and genes that influence the timing of dispersal are also expected to influence the timing of flowering. Finally, few studies have tested genetic correlations between dispersal and behavioural traits or between dispersal and physiological traits. Pedigree analyses showed that exploratory behaviour is genetically correlated with movement propensity in great tits Parus major and that departure and settlement are genetically correlated with aggressiveness in western bluebirds Sialia mexicana (Duckworth & Kruuk, 2009; Korsten et al., 2013). In marmots Marmota flaviventris, there is a negative genetic correlation between dispersal and time allocated to vigilance (Blumstein et al., 2010).
Dispersal traits can therefore be genetically correlated with other traits under selection, and it is unclear whether such correlations result from joint selection on multiple traits, or from pleiotropy. Some trait combinations lead to higher dispersal success than others and are therefore likely to be under correlational selection. For example, in western bluebirds, higher aggressiveness in dispersers increases their colonization success at the range‐expanding front (Duckworth & Badyaev, 2007). Given that selective pressures vary along environmental gradients, when scaled up towards among‐population variation, divergent environmental gradients may eventually select for specific dispersal‐related trait combinations and/or specific dispersal syndromes. Consequently, different syndromes among populations could emerge and be maintained (Clobert et al., 2009; Cote et al., 2010, 2017a ; Ronce & Clobert, 2012). Conditional dispersal syndromes have now been reported in several studies (Byers, 2000; Gilliam & Fraser, 2001; Cote & Clobert, 2007; Bonte et al., 2008; Bolnick et al., 2009; Cote et al., 2013; Bestion et al., 2014, 2015; Pennekamp et al., 2014; Myles‐Gonzalez et al., 2015; Jacob et al., 2016). However, although the strength of genetic and environmental components of dispersal syndromes and their interactions will undoubtedly influence the eco‐evolutionary outcomes of dispersal (Legrand et al., 2017), they remain almost unexplored.
(6). Genotype–environment interactions
Active dispersers are known to adjust their dispersal decisions according to the environmental conditions they encounter at each of the three dispersal stages (i.e. context‐dependent dispersal or conditional dispersal; Clobert et al., 2009). Similarly, passive dispersal can be impacted by the environment indirectly – such as temperature effects on plant height and thus dispersal distance (Zhang, Jongejans & Shea, 2011) – or directly, such as changes in ocean currents transporting plankton (McManus & Woodson, 2012). At the molecular level, the associations between certain candidate genes and dispersal have also been shown to depend on environmental conditions. Dispersal phenotypes therefore result from the combination of genetic (G) and environmental (E) effects, and genotype–environment (G × E) interactions. For instance, the associations between the Pgi gene and flight metabolic rate or other traits (see Section II.2) depend on thermal conditions in several butterfly species (Karl & Fischer, 2008; Niitepõld, 2010; Mattila, 2015) and food availability in the blue‐tailed damselfly Ischnura elegans (Block & Stoks, 2012). Similarly, when D. melanogaster larvae are reared under low‐nutrient conditions, adults with for S alleles exhibit more exploratory behaviour and a lower reproductive output than for R individuals, the opposite association than when reared under high‐nutrient conditions (Burns et al., 2012). Studies of the migratory African armyworm, Spodoptera exempta, are good examples of the existence of G × E interaction (Gatehouse, 1989, 1997). Selection experiments of this species show that only individuals raised in the laboratory under high density are selected to fly long distances. Otherwise, individuals showed a short flight duration. The existence of significant G × E interactions would indicate that genetic variation underlies such plasticity, and the potential for selection to act on conditional dispersal (e.g. Venable, 1985; Gatehouse, 1989, 1997; Imbert & Ronce, 2001; Riba et al., 2009, reviewed in Zera & Brisson, 2012; Arendt, 2015).
While evidence of conditional dispersal is substantial, quantitative studies on the genetic component of such plasticity are still relatively rare. Pennekamp et al. (2014) quantified the contribution of G, E, and G × E to this variable for 44 different genotypes from three population densities in the ciliate Tetrahymena thermophila. They found that 43% of the variance in dispersal propensity was explained by G, 13% by E and 2% by G × E, with reaction norms (Appendix I) to density varying among genotypes from negative to positive density dependencies (see also Fronhofer, Kropf & Altermatt, 2015a). A recent study showed that this significant G × E interaction could be explained by other traits like cooperation and result in the variation of dispersal syndromes along a density gradient (Jacob et al., 2016). In the spider Erigone atra, temperatures during development affected both long‐ and short dispersal strategies (Bonte et al., 2008). Using parent–offspring regressions, Bonte et al. (2003) found that 16 and 13% of the phenotypic variation in pre‐dispersal behaviour in response to multiple stressors could be attributed to genotypic and genotype × environment interactions, respectively. Both studies thus point to the existence of substantial genetic variation in dispersal reaction norms for long‐distance dispersal strategies, but not for more routine (short‐distance) behaviours. In another study using saltmarsh‐inhabiting wolf spiders Pardosa purbeckensis, population‐specific reaction norms relative to wind conditions were found to be adaptive relative to putative costs of transfer. Genetic variation in the reaction norms declined significantly with habitat and thus population size (Bonte et al., 2007). Similarly, in the plant Arabidopsis thaliana, significant heritability was detected for seed dispersal patterns, but the magnitude of heritability depended on the density at which the plants were grown (Donohue et al., 2005). Since density is, in part, determined by seed dispersal patterns, this suggests that the evolutionary potential (expression of genetic variation) of dispersal could depend on the level of dispersal itself. Hence, evolutionary or plastic changes in dispersal, which determine post‐dispersal density, could either mask or release further genetic variation for dispersal, depending on specific patterns of genotype–environment interaction (Donohue, 2005).
Importantly, the environmental conditions that influence dispersal can be experienced by the individual itself either during its development or during the adult stage, but also by its parents (so‐called parental or transgenerational effects; e.g. Sinervo et al., 2006; Tschirren, Fitze & Richner, 2007; Ducatez et al., 2012; Bestion et al., 2014; van Petegem et al., 2015). When plasticity occurs through maternal effects, it might be difficult to tease apart environmental from genotype × environment effects. This is because maternal genes and broad‐sense epigenetic factors (including hormonal conditions or endosymbionts) are confounded, and maternal environmental conditions can select simultaneously for different genotypes through selective embryo mortality (Duong et al., 2011). Maternal effects can be distinguished from genetic effects through comparisons between father–offspring and mother–offspring regression (Charmantier et al., 2011), Animal Models using full pedigrees (Charmantier et al., 2011), or manipulations of endosymbiotic communities or of the environmental conditions of mothers during gestation (Sinervo et al., 2006; Tschirren et al., 2007; Goodacre et al., 2009; Bestion et al., 2014).
III. REPRESENTATION OF DISPERSAL GENETICS IN MODELS
To date, models that explore the evolution of dispersal have almost exclusively been developed to provide general, and typically qualitative, theoretical insights. This is largely because without information on dispersal genetics it is extremely challenging to develop models that can provide more quantitative insights into, for example, the number of years we might expect it to take for emigration rate of a particular mammal species to evolve to a new equilibrium following fragmentation of its habitat. As we have seen above, it is only in very recent years that we have begun to gain empirical understanding of dispersal genetics. This will now allow us to develop models that can make the more quantitative predictions that are required if we are to integrate dispersal evolution into models developed for forecasting how ecological systems will respond to environmental changes (Urban et al., 2016), and for informing management. In this section, we start by reviewing the questions asked, and approaches typically used, in theoretical models for dispersal evolution. We subsequently highlight how new theory can usefully be developed by considering greater genetic complexity. New genetically explicit models can, together with the emerging empirical information, provide the foundations required to build a new generation of models that can yield (quantitative) eco‐evolutionary predictions and that can be used to develop more‐effective management practices.
When developing general theory about the evolution of any aspect of life history, biological assumptions need to be justified. While it is understood that dispersal phenotypes are the product of multiple traits, our review below illustrates that most models have not incorporated the underlying complex mechanisms. Rather, models generally represent compound phenotypes, such as the rate of leaving the natal patch (a proportion), or parameters of a simple dispersal kernel (i.e. the distribution of distances over which individuals are displaced from their natal site). Despite the potentially complex genetic architecture and apparently polygenic genetic basis of dispersal, models have remained genetically very simple, with most of them assuming a single locus affecting a single trait. Such an oversimplification of the genetic basis of a phenotype has been dubbed the ‘phenotypic gambit’ (Grafen, 1984), because the loss of genetic realism trades off favourably with the mathematical/computational tractability and generality of the models.
(1). What dispersal phenotypes are modelled?
Theoretical models have typically focused on the evolution of one of the three phases of dispersal (departure, transfer or settlement) and have fixed the other two (see online Table S2). The vast majority of models have focused on the departure phase by modelling the evolution of rate/probability of emigration from a natal patch (see online Table S2). Early models, considering context‐ (e.g. density‐) independent rates of emigration, were fundamental in elucidating what we now understand to be the major drivers of selection for emigration, such as kin‐competition (e.g. Hamilton & May, 1977; Motro, 1982a , b ; Frank, 1986; Taylor, 1988; Gandon & Michalakis, 1999), environmental variability (e.g. Comins, Hamilton & May, 1980; Hastings, 1983; Levin, Cohen & Hastings, 1984) and inbreeding depression (e.g. Motro, 1991; Gandon, 1999; Perrin & Mazalov, 1999). There is now an increasing number of models that explore the evolution of more‐complex emigration rules that may depend, for example, on environmental conditions, local density of conspecifics (McPeek & Holt, 1992; Travis, Murrell & Dytham, 1999; Poethke & Hovestadt, 2002; Kun & Scheuring, 2006), and local density of prey/parasites/predators (Travis et al., 2013a; Iritani & Iwasa, 2014). Further, some recent work has begun to consider how the quality of information available to actively dispersing individuals influences the evolution of emigration and the evolution of investment in the acquisition of information (Armsworth, 2009; Enfjäll & Leimar, 2009; Bocedi, Heinonen & Travis, 2012; Delgado et al., 2014).
An interesting set of studies explores the evolution of dispersal kernels (see online Table S2). These studies focus primarily on the distances over which individuals are selected to travel, so they consider mainly the transfer phase of dispersal. However, it should be noted that in some cases, the dispersal kernel that evolves also determines the emigration rate, since only those individuals that disperse far enough to leave their natal habitat are emigrants (e.g. Hovestadt, Messner & Poethke, 2001; Rousset & Gandon, 2002; North, Cornell & Ovaskainen, 2011; Fronhofer, Poethke & Dieckmann, 2015b). Notably, these studies on the evolution of dispersal kernels have not tackled context dependencies in the dispersal distances to the same degree as context‐dependent emigration has been treated. Recent theory has taken an alternative approach to modelling the transfer phase by incorporating evolution into more‐mechanistic descriptions of movement. Examples include the evolution of plant height (and thus seed release) with a mechanistic model determining how far individual seeds are dispersed (Travis et al., 2010b), the evolution of the correlation of animal movement trajectories (Heinz & Strand, 2006; Bartoń et al., 2009, 2012), the evolution of boundary‐crossing behaviours (Martin & Fahrig, 2015), and the evolution of behavioural rules in plankton that influence the currents in which individuals are dispersed and thus how far they travel (Pringle et al., 2014). Theory on the settlement phase of dispersal is the scarcest. There are however a handful of examples looking at the evolution of settlement behaviours (typically probability of settling) based on habitat selection (Stamps, Krishnan & Reid, 2005), mate finding (Shaw & Kokko, 2014, 2015), prey (Travis et al., 2013a) and conspecific density (Poethke et al., 2011b).
(2). How is the genetics of dispersal modelled?
(a). The ‘phenotypic gambit’: modelling dispersal phenotypes with hints of genetic architecture
Early analytical models focused on the evolution of emigration rate. They typically proceeded by considering the fate of a single mutant allele, or phenotype, of a small effect in an otherwise monomorphic haploid population. These models predicted the circumstances under which an optimal dispersal rate is evolutionarily stable, and can be reached by small mutational steps. Such models usually considered asexual species with a single phenotype that had one of two possible states, migrant versus non‐migrant, or resident versus mutant, in game‐theoretical or adaptive dynamics approaches (e.g. Hamilton & May, 1977; Comins et al., 1980; Hastings, 1983; Levin et al., 1984; Frank, 1986; Taylor, 1988; see online Table S2). A few early population‐genetic models considered sexual species with either a haploid locus bearing discrete alleles (e.g. van Valen, 1971; Motro, 1982a) or a diploid bi‐allelic locus, that directly set the dispersal probability of the gene carriers and followed changes in allele frequencies (e.g. Roff, 1975; Motro, 1982b, 1983). Although more complex analytical models are being developed and are still being used fruitfully (e.g. Ravigné et al., 2006; Vitalis et al., 2013; Blanquart & Gandon, 2014; Massol & Debarré, 2015), such models still focus on discrete phenotypes and ignore their complex polygenic determination and any genetic mechanisms that might impact its evolution. While this approach might be appropriate to answer questions on what strategy eventually evolves and if it is evolutionarily stable, it does not allow consideration of transient dynamics that are likely to be most affected by the genetic architecture, and are fundamental in eco‐evolutionary dynamics and responses to environmental changes (see Sections III.2d and III.3; Dunlop, Heino & Dieckmann, 2009; North et al., 2011). Ultimately, the constraints imposed by the genetic architecture and the possibility for eco‐evolutionary dynamics (see Section III.2d ) may even determine the possibility of reaching alternative evolutionary outcomes (Crespi, 2004).
In the last three decades, the theory of dispersal evolution has benefited from the development of more flexible population‐based and individual‐based simulation models (see online Table S2). While these approaches have less generality, they can achieve greater realism of the modelled evolutionary scenarios. However, in terms of genetic realism, these models remain simplistic. For example, emigration probability is usually coded by a single haploid or diploid locus carrying continuous and additive allelic values bounded between 0 and 1 (e.g. Poethke & Hovestadt, 2002; Travis & Dytham, 2002; Guillaume & Perrin, 2006), or a set of discrete allelic values (e.g. McPeek & Holt, 1992; Travis & Dytham, 1999). The advantage of this simplification, however, is the possibility to match analytical model predictions with simulation results, and to ignore complications arising from multi‐locus genetics.
Some simple elements of single‐locus genetic architectures have been implemented in population‐genetic and individual‐based simulation models (see online Table S2). Regarding intra‐locus allelic interactions, in most of the diploid models the two alleles are co‐dominant in heterozygotes and the genotypic value at the locus is determined by either the arithmetic mean or the sum of the two allelic values. A few models with discrete alleles consider dominance interactions, generally modelling complete dominance of one allele over the other (Leimar & Norberg, 1997; Motro, 1982b; Roff, 1994). More recently, Fronhofer et al. (2011) modelled evolution of emigration probability considering evolution of dominance interactions between two continuous alleles at a single locus. The dominance of one allele over the other was determined by a ‘dominance modifier’ diploid locus determining the dominance value of each allele. Dominance interactions were therefore allowed to evolve from co‐dominance, to partial dominance, to complete dominance. Interestingly, this study showed that evolving dominance allows the maintenance of dispersal polymorphisms, which otherwise disappear when assuming fixed co‐dominance.
The mapping between genotype and phenotype is most often very simple, with complete equivalence being assumed, and hence no environmental variance. Thus, most models consider the heritability of the modelled phenotypes to be equal to 1. Very few models have taken a more explicit quantitative‐genetic approach (e.g. Perkins et al., 2013) where environmental variance contributes to the dispersal phenotype, hence decreasing the heritability of traits to less than 1. Phenotypic expression, typically of wing dimorphism, has also been modelled through the quantitative genetic threshold model (Roff, 1994). Here, a continuous and normally distributed characteristic, called liability (Falconer & Mackay, 1996), underlies a discrete phenotypic variation (e.g. dimorphism). Individuals with liability above a certain threshold express one morph (e.g. wings), while individuals with liability below the threshold express the alternative morph (e.g. no wings). While the liability might potentially be modelled in a genetically explicit manner (i.e. with individuals carrying multiple loci; e.g. Duthie, Bocedi & Reid, 2016), we are not aware of models that do so for dispersal traits. Models considering the evolution of sex‐biased dispersal assume sex‐limited phenotypic expression, where two loci carried by both sexes separately code for male and female dispersal, while each sex expresses only the relevant phenotype (e.g. Motro, 1991; Guillaume & Perrin, 2006, 2009; Gros, Hovestadt & Poethke, 2008; Gros, Poethke & Hovestadt, 2009; Meier, Starrfelt & Kokko, 2011; Shaw & Kokko, 2014, 2015; Henry, Coulon & Travis, 2016).
(b). Reaction norms and multiple‐traits models
Several studies have investigated the evolution of plastic phenotypic responses to biotic or abiotic conditions (reaction norms) where, for example, the probability of emigration depends on conspecific density or habitat quality. In this case, the modelled phenotype is a function describing the individual's response to a varying condition. Each of the function's parameters (one or more) is coded by one locus, and free recombination and no pleiotropy are assumed among loci coding for different parameters (e.g. Kun & Scheuring, 2006; Bonte & De La Peña, 2009; Enfjäll & Leimar, 2009; Travis et al., 2009; Shaw & Kokko, 2015).
A few studies have modelled the joint evolution of multiple dispersal traits, for example traits involved in more than one dispersal phase (e.g. Yukilevich, 2005; Poethke et al., 2011b; Travis et al., 2012), and some studies have considered joint evolution of dispersal and other traits (see online Table S2). These include local adaptation (Billiard & Lenormand, 2005; Kubisch et al., 2013; Blanquart & Gandon, 2014; Berdahl et al., 2015; Hargreaves, Bailey & Laird, 2015; ), rate of self‐fertilization (Cheptou & Massol, 2009; Sun & Cheptou, 2012), seed dormancy fraction (Vitalis et al., 2013), sex‐ratio (Leturque & Rousset, 2003), animal perceptual range (Delgado et al., 2014), and resource investment in reproduction and competitive ability (Burton, Phillips & Travis, 2010; Fronhofer & Altermatt, 2015; Fronhofer et al., 2017) or in acquisition of information (Bocedi et al., 2012). As for the single‐trait models, multiple‐trait models include analytical and quantitative genetic models, as well as population‐genetic and individual‐based models. Where a genetic basis is modelled, each trait is coded by one locus. Different loci are unlinked (i.e. freely recombining) in most models (e.g. Poethke, Dytham & Hovestadt, 2011a; Bocedi et al., 2012; Travis et al., 2012; Kubisch et al., 2013), but in others are completely linked assuming a biologically sensible trade‐off (Fronhofer et al., 2011), or recombine at a certain rate (Billiard & Lenormand, 2005; Yukilevich, 2005; Blanquart & Gandon, 2014). We are not aware of any model implementing pleiotropy, where the same gene can affect more than one trait.
(c). Dispersal evolution and the genetic architecture of inbreeding depression
More‐explicit genetic architecture has been considered in models investigating the role of inbreeding depression in the evolution of emigration probability (Morgan, 2002; Roze & Rousset, 2005, 2009; Guillaume & Perrin, 2006, 2009; Ravigné et al., 2006; Henry et al., 2016). While the dispersal trait is still coded by a single locus, different multilocus architectures are implemented for loci determining inbreeding depression, thus allowing it to evolve. In these models, the mutation load is determined by multiple diploid loci carrying deleterious mutations. At each locus, the fitness effect of the deleterious mutation is determined by the selection coefficient against the mutation and its dominance coefficient (Higgins & Lynch, 2001). Typically, models assume selection and dominance coefficients to be the same for all loci and also assume multiplicative effects on fitness across loci. These models have revealed important effects of the genetic architecture of the mutation load on inbreeding depression and, consequently, on the evolution of dispersal. For example, Guillaume & Perrin (2006) showed that in small populations, inbreeding depression, and consequent heterosis, can drive dispersal evolution as a means of inbreeding avoidance when deleterious mutations are strongly recessive, so that they do not go to fixation and are not purged from the population, thus maintaining heterosis. However, mildly deleterious and partially recessive mutations appearing at high rates may quickly accumulate in small populations, increasing the mutation load to the point of causing population extinctions and high demographic stochasticity, which also induces selection for higher dispersal. The amount of heterosis maintained in the population, and hence the strength of selection for increased dispersal, also depends on the number of loci that contribute to inbreeding depression, where a low number of loci can substantially decrease heterosis (Roze & Rousset, 2009).
(d). Eco‐evolutionary dynamics and the importance of genetic architecture
Dispersal is key in linking ecological and evolutionary dynamics as it connects populations both demographically and genetically (Hanski, 2012; Hendry, 2016). Variation in population densities and demography caused by dispersal feeds back on the evolutionary dynamics of the dispersal trait itself, or other life‐history or fitness‐related traits, by affecting changes in allele frequencies, the strength of density‐dependent mechanisms, the opportunity to either avoid conspecifics (e.g. Ronce, 2007; Kubisch et al., 2014) or to follow them in some other cases (e.g. Penteriani & Delgado, 2011; Coppock, Gardiner & Jones, 2016). The resulting eco‐evolutionary dynamics (also called eco‐evolutionary feedback when the effects between ecology and evolution are reciprocal; Post & Palkovacs, 2009; Hanski, 2012) are characterized by rapid evolution due to strong selection, and are likely to occur, or at least to be most detectable, during environmental change or in highly spatio‐temporally variable environments (Reznick & Ghalambor, 2001; Crespi, 2004; Hanski, 2011, 2012). Accordingly, rapid evolution of dispersal has been predicted and documented during range expansion or shifts and as a consequence of habitat fragmentation (see Section II.1; Hanski, 2012; Cheptou et al., 2017; Legrand et al., 2017).
Theoretical models predict interesting eco‐evolutionary dynamics involving dispersal evolution (see also Section II.1). For example, both ‘evolutionary rescue’ determined by evolution of increased dispersal, and ‘evolutionary suicide’ determined by loss of dispersal, can be predicted for species expanding over fragmented landscapes or experiencing a sudden fragmentation event (Leimar & Norberg, 1997; Heino & Hanski, 2001; Gyllenberg et al., 2002; Travis et al., 2010b; Poethke et al., 2011a). The outcome will depend on factors such as the stage of expansion, the manner and rapidity of fragmentation and, importantly, the amount of genetic variation in dispersal traits. Further, models of range shifting following a shift in environmental conditions predict ‘elastic’ range margins, where the range shifts and expands due to rapid increase in dispersal during the environmental change, but then retracts due to decreased dispersal following the cease of the change (Holt, 2003; Kubisch, Hovestadt & Poethke, 2010; Henry, Bocedi & Travis, 2013). The type of environmental gradient determines the amplitude of the fluctuation in range size.
The impact of the genetic architecture of dispersal traits on dispersal eco‐evolutionary dynamics has mostly been ignored. An exception is the model by Kubisch et al. (2014) which showed that ploidy affects dispersal evolution, and hence rate of range expansion, depending on the type of environmental gradient over which expansion occurs. While clonal species expanded faster in shallow gradients relative to sexually reproducing species, they expanded more slowly over steeper gradients. Whether or not genetic architecture might affect evolutionary equilibria, it surely affects the rate of evolution (see Section II.3) which is key for eco‐evolutionary dynamics to occur. For example, rapid evolutionary changes likely involve a combination of standing genetic variation and new mutations (Hendry, 2013), with the balance between the two determining not only the speed of evolutionary changes, but also the likelihood that eco‐evolutionary dynamics will occur (Becks et al., 2010). For example, North et al. (2011) showed that high mutation rates allowed maintenance of dispersal polymorphisms in fragmented landscapes with high patch turnover rate. In turn, the occurrence or not of eco‐evolutionary feedbacks might affect the amount of adaptive genetic variation that is maintained (Becks et al., 2010; Hanski, 2012). Hence, the underlying genetic architecture determining maintenance of genetic variation prior to changes, as well as mutation rate and magnitude, will likely affect dispersal eco‐evolutionary dynamics (R. C. Henry, G. Bocedi & J. M. J. Travis, in preparation). How much additive genetic variation is generally present within a population is still much debated and indeed this might be idiosyncratic to the population, its evolutionary history and spatial location (e.g. marginal versus core habitats; bottlenecks) (Kawecki, 2008; Hanski, 2012; Hendry, 2013). However, even in cases where populations seem to be genetically depauperate, rapid evolution (including of dispersal) has been observed (e.g. Rollins et al., 2015). The genetic mechanisms behind this are unknown although heritable epigenetic variation might be a potential one (Bonduriansky & Day, 2009; Rollins et al., 2015; Legrand et al., 2017). Further genetic variation can be maintained by non‐additive genetic effects (dominance and epistasis) and be converted to additive genetic variation following a bottleneck (Roff & Emerson, 2006). Finally, genetic covariances among traits might either favour or hinder rapid evolutionary responses (Reznick & Ghalambor, 2001; Etterson, 2004; Hendry, 2013).
While a few theoretical studies have begun to consider explicit genetic architecture in modelling the eco‐evolution of dispersal, models developed and parameterized for addressing questions related to particular species in particular environments remain even scarcer. A recent exception is Hanski et al. (2017), who used a metapopulation model to predict the distribution of the Glanville fritillary butterfly over 22 years across the heterogeneous landscape of the Åland islands SW Finland. Of the 125 population networks 33 were above the extinction threshold for long‐term persistence. Spatial configuration and habitat area predicted metapopulation size and persistence, but interestingly in the 33 networks above the extinction threshold, allelic variation in SNPs of the Pgi gene explained ∼30% of variation in metapopulation size. Based on previous knowledge (see Section II.2), this indicates that individuals with specific Pgi genotypes that are more dispersive increase colonization rate and decrease extinction rate of these population networks (Hanski et al., 2017). Consistently, three viable networks that were extinct for at least 5 years had lower frequency of the dispersive genotype than the 27 networks that persisted for the full 22 years (Hanski et al., 2017).
(3). Why is it important to include the genetic architecture of dispersal in models?
Dispersal traits are often quantitative traits showing continuous and heritable variation within populations (see Section II and see online Table S1) and can thus be assumed polygenic in nature. This assumption, however, potentially introduces many complications when modelling the evolution of a trait. Ignoring, or not, these complications may matter in some cases but not in others. For example, and as alluded to above, details of the distribution of allelic effects at loci affecting a trait will affect its evolutionary dynamics. Previous models, nevertheless, have ignored these genetic details and have been based on much simpler assumptions (reviewed in Section III.2a ), in part for practical reasons but also because modellers were mostly interested in finding optimal strategies, corresponding to equilibrium points located at maxima of fitness landscapes. Whether and how the optimal strategy is reached by evolution then depends mostly on the shape of the adaptive landscape around an equilibrium point and less on the genetic architecture of the trait [see Ronce, 2007 for a review of modelling techniques]. This is because we generally assume that, as long as genetic variation exists in the direction of selection on the adaptive landscape, a population will reach the equilibrium point.
Details of the genetic architecture of dispersal trait(s) may then matter most when considering the evolutionary dynamics of a (meta)population, especially in non‐equilibrium situations such as range expansions, or extinction–colonization dynamics. That is, the number of loci, their distribution of allelic effects and possible epistatic interactions, their mutation and recombination rates, and their number of pleiotropic effects will all affect the rate of trait evolution towards an optimal strategy even if they do not affect the existence or position of that strategy on an adaptive landscape. However, when dispersal evolution and eco‐evolutionary dynamics are co‐determined, for instance during range expansion, genetic details underlying trait variation may have quantitative and qualitative effects on model outcomes. Typically, of the features just enumerated, some will enhance and some will constrain the rate of evolution of a dispersal trait and are thus worth considering because they may feedback on the evolutionary process itself. We will, however, not provide an exhaustive review of all possible effects of complex polygenic architectures on trait evolution but rather focus on a few simple examples.
First, keeping in mind that rates of evolution depend on the amount of additive (heritable) genetic variation in a trait, any characteristic that increases the genetic and thus the phenotypic variance of a trait will be favourable under directional selection towards an optimum value. Increasing the number of loci, the mutation rate, or the mutational effect size (i.e. proportion of variance contributed by a locus) will speed up evolution by increasing trait variation, but will become an impediment when selection is stabilizing by generating a variance load (i.e. reduction of mean population fitness due to phenotypic variation around the optimum). Furthermore, when loci contribute unequally to trait variation, the shape of the phenotypic distribution may depart from normality, and become skewed, or have higher kurtosis. For example, Débarre, Yeaman & Guillaume (2015) showed that when traits are affected by a few loci of large effect (i.E. major loci) together with many loci of small effect (i.E. minor loci), they are better able to resist homogenization by gene flow and lead to better local adaptation under diversifying selection between divergent habitats than when only minor loci are considered. In Débarre et al. (2015), increasing the allelic effect of major loci increased the skewness of the phenotypic distributions in divergent populations connected by gene flow, which increases the response to selection towards phenotypic optima. The enhancing effect of major loci on rates of evolution has also been shown when populations face a sudden, large shift in their environment and when a trait is determined by a mix of major and minor loci (Gomulkiewicz et al., 2010), or during range expansion on coarse or stepwise environmental gradients (Gilbert & Whitlock, 2017). Traits determined by many minor loci can, however, confer faster adaptation to coarse gradients when the mutation rate is large (Gilbert & Whitlock, 2017). Finally, genetic architectures with many possible mutational paths will cause faster trait evolution than genetic architectures that constrain trait variation on a single path. That is, traits determined by many loci at which multiple allelic combinations may provide the same trait value (i.e. when there is genetic redundancy) will achieve higher rates of evolution compared to when trait values can only be reached by a single combination of alleles (Yeaman, 2015). All these aspects must thus be carefully considered when modelling dispersal as a polygenic trait.
Second, when selection acts on allelic variants at multiple loci, statistical linkage, and thus linkage disequilibrium (LD) may ensue because some allelic combinations may be favoured over others. Selection will then maintain LD among beneficial allelic combinations, which contributes to adaptation in heterogeneous environments despite recombination and sometimes strong gene flow (Le Corre & Kremer, 2003, 2012). This is especially true when adapting from standing genetic variation in the presence of gene flow and for genetic architectures with high genetic redundancy, which can maintain clusters of co‐adapted loci that can function as loci of major effect (Yeaman & Guillaume, 2009; Yeaman & Whitlock, 2011). The rate of recombination, in addition to the number and effect sizes of loci under selection, is thus an important parameter to consider when modelling polygenic traits.
Third, dominance and epistasis, both representing non‐additive allelic effects, are important in the evolution of quantitative traits because of their contributions to trait genetic variance (Falconer & Mackay, 1996; Lynch & Walsh, 1998). They are seldom taken into account, mostly because both are very complex features to model in multi‐locus systems, and are expected to be of most influence in small or inbred populations (e.g. for dominance: Wolak & Keller, 2014; see Section III.2a for single‐locus models with dominance). Whether epistasis contributes significantly to trait evolution is also still under intense debate (e.g. Hansen, 2013; Barton, 2017). However, it is safe to assume that epistasis can be ignored for polygenic traits affected by many loci of small effects at which selection is of the same intensity as drift, in which case epistasis will not contribute much to trait variation (Barton, 2017).
Finally, genetic non‐independence among traits, caused by physically linked or pleiotropic loci, has the potential either to constrain or enhance the co‐evolution of correlated traits (Lande, 1979; Walsh & Blows, 2009). For instance, traits that are under uniform selection across populations, such as dispersal traits or other life‐history traits, may be displaced from their optimum if they are genetically correlated to traits under directional selection (Guillaume, 2011). Such genetic non‐independence may have far‐reaching eco‐evolutionary consequences (see Section III.2d ). As reviewed in Section II.5, many dispersal traits are strongly correlated with each other and/or with other traits under selection. While the mechanisms underlying these correlations (e.g. pleiotropy, physical linkage, environmentally induced correlations) have been identified in only a few cases, many of these traits likely share a common genetic basis and are subject to correlated evolution. However, current models consider different traits as evolving independently. In some models, correlated evolution can arise because of imposed or emerging trade‐offs (e.g. dispersal versus reproductive abilities), likely generating statistical linkage, but genetic constraints have not yet been considered.
(4). Case study: habitat fragmentation and the genetic architecture of dispersal
To assess the importance of considering genetic architecture when modelling eco‐evolutionary dynamics of dispersal, we present results from a genetically and spatially explicit individual‐based model used to investigate how the genetic architecture of dispersal affects a population's response to habitat fragmentation. Specifically, we illustrate that a population's evolutionary response to habitat fragmentation is slowed when a large number of loci with frequent smaller mutations, as opposed to a small number of loci with fewer larger mutation, are controlling a dispersal trait.
(a). The model
We modelled a spatially explicit landscape of 400 patches, each with a carrying capacity of 30 individuals. Individuals are hermaphroditic and generations are non‐overlapping. Each individual in a patch produces a number of offspring drawn from a Poisson distribution with mean 3. These offspring are each fathered by an individual randomly drawn from the same patch. Selfing occurs with the same probability as mating with any other individual. Individuals have a diploid genome composed of L freely recombining loci (L = 1, 2, 5, 10, 20, 40 loci) with continuous alleles, coding for an individual's probability of leaving the natal patch. The individual's genotype is calculated as the sum of the 2L allelic values. Phenotypic values are directly determined by genotypes (heritability equals 1). The phenotypic value of an individual gives the probability that the individual will emigrate from its natal patch and therefore, while alleles can assume any continuous value, phenotypic values are constrained between 0 and 1. We assume no environmental variance. To ensure that the initial phenotypic variance is the same among simulations with varying numbers of loci (L), we scale the distribution of allelic and mutational effects by L. At initialization, individuals are assigned allelic values drawn from a normal distribution with a zero mean and standard deviation (σ a) that has been scaled according to the number of loci,
where σ is the initial standard deviation equal to 0.5. Offspring inherit their dispersal alleles from their parents with a probability of mutation μ = 0.001 per allele per generation. When a mutation occurs, it is sampled from the same distribution as above and added to the existing allele value.
Thus for small genomes, alleles have larger effects and few large‐effect mutations occur; for large genomes, alleles have small effects and many small‐effect mutations occur. If an offspring disperses, it moves to one of eight neighbouring cells with equal probability and a 0.05 chance of mortality. Additional mortality is incurred by dispersing out of suitable patches upon habitat fragmentation. After dispersal, the pool of individuals in each patch is reduced to carrying capacity. Density regulation is random with respect to an individual's genotype and results in patches filled to capacity when enough individuals are present. The model initially runs for 50000 generations with 100% of patches suitable to ensure equilibrium in emigration probability is reached. After this time a sudden habitat fragmentation event occurs and the proportion of suitable patches is reduced to 60%. Patches that remain suitable are chosen at random. The model then runs for a further 10000 generations.
(b). Findings
Prior to the fragmentation event the mean and variance of the phenotypic distributions of dispersal probability are similar between simulations with different numbers of loci (Fig. 1). Despite this, the speed of the response to fragmentation differs among simulations and this has associated effects on the population sizes. In populations in which a large number of loci control dispersal, emigration probability evolves less quickly to low values, and there is typically a smaller reduction in population size (Fig. 2). This is because when dispersal probabilities take longer to decline, more individuals disperse and thus recolonize empty suitable patches despite the higher chance of dispersal mortality due to the presence of unsuitable habitat.
Figure 1.

Distribution of phenotypes for dispersal probability in the population prior to the fragmentation event, generation 49999, for scenarios with one locus (A), 10 loci (B), and 40 loci (C).
Figure 2.
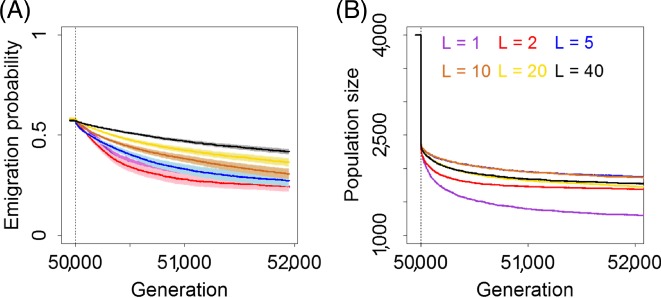
(A) Evolution of dispersal and (B) total population size after a fragmentation event, highlighted by the dashed line at generation 50000. Lines represent the mean and shading, when shown, represents the standard error of the mean of 30 replicates. L = number of loci.
These results highlight that the genetic architecture underlying dispersal, even simply the number of loci controlling dispersal traits, can have evolutionary and ecological consequences for populations. In this case, alternative monogenic and polygenic representations of the genetic architecture of dispersal led to significant differences in the rate of dispersal evolution, although the direction was the same. There is clearly much scope for future work on the effect of genetic architecture on the evolution of dispersal traits and their demographic consequences.
IV. CONCLUSIONS
(1) Our review of the empirical literature on the genetic basis of dispersal shows that the available evidence is largely skewed to a few groups of organisms, notably insects, birds and plants. Overall, these studies show that dispersal often has a genetic basis, that it has evolutionary potential in the form of genetic variation, and that dispersal may evolve in response to natural selection, sometimes even over relatively short timescales. Dispersal, which is a complex process involving departure, transfer and settling and is the outcome of several interacting traits, appears to be polygenic, even though genes of moderate to large effect may control certain aspects of dispersal. Correlations among the multiple traits determining dispersal, and among other life‐history traits, are still relatively poorly understood, even though they might contribute to the evolution of dispersal syndromes, and may constrain dispersal from evolving to its adaptive optimum. Further, dispersal can be highly plastic and, as a result, genetic variation for dispersal plasticity can lead to environment‐dependent genetic variation for dispersal. We still lack understanding of the relative importance of plasticity versus allelic variation in influencing dispersal phenotypes.
(2) Major unanswered empirical questions regarding the genetic basis of dispersal include the degree to which dispersal traits are correlated with each other and with other traits under selection. This is especially important for a complex process such as dispersal, which is likely to be determined by diverse physiological, morphological, metabolic, and behavioural traits, each of which may be exposed to selection via processes other than dispersal. Another arena for further empirical investigation is the environmental dependence of the genetic architecture of dispersal. This is important particularly if dispersal itself causes a change in the environment that the organism experiences. Finally, identification of specific genes underlying dispersal variation in natural populations would provide access to resolving important issues of dominance, epistasis, and allelic effects of dispersal genes.
(3) Importantly, the potential roles of epigenetic mechanisms underlying variation in dispersal are almost unknown although such mechanisms can influence both the context dependency of dispersal (i.e. the intragenerational level) and the evolution of dispersal (i.e. the transgenerational level). As environmentally sensitive molecular elements causing phenotypic changes, epigenetic factors have the potential to provoke rapid responses to environmental changes. Therefore, ignoring the epigenetic architecture of dispersal can bias predictions of both its evolution and the dynamics of the response to environmental changes. Either performed in nature or under controlled conditions, future studies on the molecular bases of dispersal would be particularly useful if they include parallel investigations on the genetic and epigenetic architectures in predefined environments. This would help to unravel the interactions between the epigenetic and genetic determinants of dispersal (and its correlated traits), and also help evaluate interactions with the environmental context.
(4) Models describing the genetic basis of dispersal have remained surprisingly simple. Hence, we do not know the degree to which existing theory on dispersal evolution is robust to the limiting assumptions made about the underlying genetics, especially when considering transient and eco‐evolutionary dynamics. Further theoretical development should look at how elements of the genetic architecture of dispersal traits, such as number of loci and allelic effect size, rate of recombination and linkage disequilibrium, and non‐additive allelic effects (dominance and epistasis), will affect maintenance of additive genetic variation in dispersal traits and their potential rate of evolution. Explicitly considering genetic correlations among dispersal traits and other traits under selection may be especially productive. Addressing these gaps, and understanding the potential for rapid dispersal evolution, is particularly important in the context of non‐equilibrium scenarios such as ongoing habitat fragmentation, range expansion and shifts. Especially in cases where eco‐evolutionary feedbacks are involved, genetic architecture might also affect the ultimate evolutionary outcome in terms of evolutionarily stable dispersal strategy. Theory that combines eco‐evolutionary dynamics with elements of genetic architecture is now required to understand under what circumstances this might occur.
(5) New empirical understanding on the genetic basis of dispersal, together with novel genetically explicit theory on dispersal evolution, will enable addressing the challenge of incorporating dispersal evolution into predictive models for forecasting species' eco‐evolutionary responses to environmental changes. This will enable moving towards more realistic quantitative predictions that might ultimately inform effective species' management under ongoing environmental change.
Supporting information
Appendix S1. Methods.
Table S1. Examples of heritability (h 2) estimates in the reviewed studies.
Table S2. Models of dispersal evolution and assumptions made on the genetic architecture of the evolving traits.
V. ACKNOWLEDGEMENTS
M. S. thanks the European Research Council (Independent Starting grant META‐STRESS; 637412) and Academy of Finland (Decision numbers 273098 & 265461) for funding. M. M. D. was supported by Ministerio de Economia y Competitividad “Ramón y Cajal program” grant contract no. RYC‐2014‐16263. D. B., E. A. F., J. M. J. T. and E. M. are funded by the FWO research community EVENET. D. B. is funded by FWO research grant INVADED G.018017.N. D. L., J. C., V. M. S. and M. B. are part of the Laboratoire d'Excellence (LABEX) entitled TULIP (ANR‐10‐LABX‐41). D. L. thanks Fyssen foundation for research funding. J. C. thanks an ANR‐12‐JSV7‐0004‐01. F. G. is supported by SNSF grant PP00P3_144846. E. N. is supported by an International postdoctoral fellowship from the Swedish Research Council (Vetenskapsrådet). E. A. F. thanks Eawag for funding. J. M. B. thanks CEH for funding under project NEC05264. C. G. thanks FCT (Fundação para a Ciencia e a Tecnologia) for funding (FCT‐ANR/BIA‐BIC/0010/2013 and IF Fellowship). M. B. and A. C. thank the ANR for funding (INDHET program, ANR‐12‐BSV7‐0023). V. M. S. thanks the ANR for funding (GEMS program ANR‐13‐JSV7‐0010‐01). K. N. thanks the Ella and Georg Ehrnrooth Foundation and the Emil Aaltonen Foundation for funding. E. M. is supported by UA‐TOPBOF and FWO‐project G030813N.
GLOSSARY
| additive genetic variation | The variance in breeding values among individuals in a population (one component of the total genetic variance; Nielsen & Slatkin, 2013) |
| animal model | A linear mixed‐effects model to partion phenotypic variance into additive genetic variance and other sources of variance using relatedness between individuals as a random effects (Wilson et al., 2010) |
| candidate gene approach | The study of genes that a priori are hypothesized to contribute to the gene's functional impact on a trait (Fitzpatrick et al., 2005) |
| dispersal | Any movement of individuals or propagules that has potential consequences for gene flow across space (Ronce, 2007) |
| dispersal syndromes | Patterns of covariation of morphological, behavioural or life‐history traits associated with dispersal across species, populations or individuals (Stevens et al., 2014) |
| eco‐evolutionary feedback/dynamics | Occur when ecological and evolutionary dynamics reciprocally influence each other on a contemporary timescale (Travis et al., 2014) |
| epigenetic inheritance | Inheritance of gene expression patterns without altering the underlying DNA sequencing. The most common mechanisms are DNA methylation and histone modification (Allis & Jenuwein, 2016) |
| epistatic/epistatis interaction | Interaction between different alleles in different loci (Fox & Wolf, 2006) |
| environmental variation | Differences among individuals in a population that are due to differences in the environments they have experienced (Freeman & Herron, 2004) |
| genetic covariance/correlation | A statistical measure of the degree to which two characters vary together at the genetic level; when standardized it is known as correlation (Roff, 2002) |
| genetic architecture | A description of the number of genes, effect size of genes, interactions among genes or, if segregating polymorphisms are regulatory or structural, that influence a phenotypic trait (Schielzeth & Husby, 2014) |
| genetic variance | Differences among individuals in a population that are due to differences in genotype (Freeman & Herron, 2004). Three major components: additive genetic variance, dominance variance and epistatic variance |
| genome‐wide association study (GWAS) | A study that is designed to find a non‐random association between marker loci spread throughout the genome and a phenotypic trait (Nielsen & Slatkin, 2013) |
| genotype–environment (G × E) interaction | Differences in the effect of the environment on the phenotype displayed by different genotypes (Freeman & Herron, 2004) |
| linkage disequilibrium | Non‐random association between alleles at different loci on a chromosome (Freeman & Herron, 2004) |
| narrow‐sense heritability (h 2 ) | Proportion of the total phenotypic variance explained by additive genetic variance (V A/V P; Freeman & Herron, 2004) |
| oligogenic | When few genes are responsible for the major heritable changes in the phenotype (Barton et al., 2007) |
| phenotypic variation | The variance in phenotypes among individuals in a population (Nielsen & Slatkin, 2013). Composed of genetic and environmental variance |
| polygenic | When a trait is influenced by many genes of individually small effect (Barton et al., 2007) |
| pleiotropy | When a single locus influences multiple phenotypes (Fitzpatrick et al., 2005) |
| quantitative trait locus (QTL) | A region in the genome that contains one or several genes that contribute to a quantitative trait (Fitzpatrick et al., 2005) |
| reaction norm | Depicts the manner in which a genotype responds to the environment; can be continuous or not (Roff, 2002) |
| single‐nucleotide polymorphism (SNP) | Alteration of a single nucleotide that occurs at a specific position in the genome, found in at least 1% of the population (Fox & Wolf, 2006) |
| trait | Morpho‐physio‐phenological features of an organism which impact fitness indirectly via their effects on growth, reproduction, survival and dispersal (Violle et al., 2007) |
| transgenerational effect/parental effects | Variation among individuals due to variation in non‐genetic influences exerted by their parents or grandparents (Nielsen & Slatkin, 2013) |
VI. REFERENCES
References marked with asterisk have been cited within the supporting information.
- Albert, A. , Aufret, A. G. , Cosyns, E. , Cousins, S. A. O. , D'hondt, B. , Eichberg, C. , Eycott, A. E. , Heinken, T. , Hoffmann, M. , Jaroszewicz, B. , Malo, J. E. , Marell, A. , Mouissie, M. , Pakeman, R. J. , Picard, M. , et al. (2015). Seed dispersal by ungulates as an ecological filter: a trait‐based meta‐analysis. Oikos 124, 1109–1120. [Google Scholar]
- Allis, C. D. & Jenuwein, T. (2016). The molecular hallmarks of epigenetic control. Nature Review Genetics 17, 487–500. [DOI] [PubMed] [Google Scholar]
- Arendt, J. D. (2015). Effects of dispersal plasticity on population divergence and speciation. Heredity 115, 306–311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Armsworth, P. R. (2009). Conditional dispersal, clines, and the evolution of dispersiveness. Theoretical Ecology 2, 105–117. [Google Scholar]
- * Artacho, P. , Figueroa, C. C. , Cortes, P. A. , Simon, J.‐C. & Nespolo, R. F. (2011). Short‐term consequences of reproductive mode variation on the genetic architecture of energy metabolism and life‐history traits in the pea aphid. Journal of Insect Physiology 57, 986–994. [DOI] [PubMed] [Google Scholar]
- Baguette, M. & Van Dyck, H. (2007). Landscape connectivity and animal behavior: functional grain as a key determinant for dispersal. Landscape Ecology 22, 1117–1129. [Google Scholar]
- Bal, H. K. , Michel, A. P. & Grewal, P. S. (2014). Genetic selection of the ambush foraging entomopathogenic nematode, Steinernema carpocapsae for enhanced dispersal and its associated trade‐offs. Evolutionary Ecology 28, 923–939. [Google Scholar]
- * Baliuckas, V. , Lagerström, T. , Norell, L. & Eriksson, G. (2005). Genetic variation among and within populations in Swedish species of Sorbus aucuparia L. and Prunus padus L. assessed in a nursery trial. Silvae Genetica 54, 1–8. [Google Scholar]
- Bartle, K. , Moles, A. T. & Bonsor, S. P. (2013). No evidence for rapid evolution of seed dispersal ability in range edge populations of the invasive species Senecio madagascariensis . Austral Ecology 38, 915–920. [Google Scholar]
- Bartoń, K. A. , Hovestadt, T. , Phillips, B. L. & Travis, J. M. J. (2012). Risky movement increases the rate of range expansion. Proceedings of the Royal Society of London B 279, 1194–1202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bartoń, K. A. , Phillips, B. L. , Morales, J. M. & Travis, J. M. J. (2009). The evolution of an “intelligent” dispersal strategy: biased, correlated random walks in patchy landscapes. Oikos 118, 309–319. [Google Scholar]
- Barton, N. H. (2017). How does epistasis influence the response to selection? Heredity 118, 96–109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barton, N. H. , Briggs, D. E. G. , Eisen, J. A. , Goldstein, D. B. & Patel, N. H. (2007). Evolution. Cold Spring Harbor Laboratory Press, Cold Spring Harbor. [Google Scholar]
- Becks, L. , Ellner, S. P. , Jones, L. E. & Hairston Nelson, G. J. G. (2010). Reduction of adaptive genetic diversity radically alters eco‐evolutionary community dynamics. Ecology Letters 13, 989–997. [DOI] [PubMed] [Google Scholar]
- Begin, M. & Roff, D. A. (2002). The common quantitative genetic basis of wing morphology and diapause occurrence in the cricket Gryllus veletis . Heredity 89, 473–479. [DOI] [PubMed] [Google Scholar]
- Berdahl, A. , Torney, C. J. , Schertzer, E. & Levin, S. A. (2015). On the evolutionary interplay between dispersal and local adaptation in heterogeneous environments. Evolution 69, 1390–1405. [DOI] [PubMed] [Google Scholar]
- * van Berkum, F. H. , Huey, R. B. , Tsuji, J. S. & Garland, T. Jr. (1989). Repeatability of individual differences in locomotor performance and body size during early ontogeny of the lizard Sceloporus occidentalis . Functional Ecology 3, 97–105. [Google Scholar]
- * Berwaerts, K. , Matthysen, E. & van Dyck, H. (2008). Take‐off flight performance in the butterfly Pararge aegeria relative to sex and morphology: a quantitative genetic assessment. Evolution 62, 2525–2533. [DOI] [PubMed] [Google Scholar]
- Bestion, E. , Clobert, J. & Cote, J. (2015). Dispersal response to climate change: scaling down to intra specific variation. Ecology Letters 18, 1226–1233. [Google Scholar]
- Bestion, E. , Teyssier, A. , Aubret, F. , Clobert, J. & Cote, J. (2014). Maternal exposure to predator scents: offspring phenotypic adjustment and dispersal. Proceedings of the Royal Society of London B 281, 20140701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Billiard, S. & Lenormand, T. (2005). Evolution of migration under kin selection and local adaptation. Evolution 59, 13–23. [PubMed] [Google Scholar]
- Bitume, E. , Bonte, D. , Ronce, O. , Olivieri, I. & Nieberding, C. M. (2015). Dispersal distance is influenced by parental and grand‐parental density. Proceedings of the Royal Society of London B 281, 20141061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bitume, E. V. , Bonte, D. , Magalhaes, S. , San Martin, G. , Van Dongen, S. , Bach, F. , Anderson, J. M. , Olivieri, I. & Nieberding, C. M. (2011). Heritability and artificial selection on ambulatory dispersal distance in Tetranychus urticae: effects of density and maternal effects. PLoS One 6, e26927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blanquart, F. & Gandon, S. (2014). On the evolution of migration in heterogeneous environments. Evolution 68, 1617–1628. [DOI] [PubMed] [Google Scholar]
- Block, M. & Stoks, R. (2012). Phosphoglucose isomerase genotype effects on life history depend on latitude and food stress. Functional Ecology 26, 1120–1126. [Google Scholar]
- Blumstein, D. T. , Lea, A. J. , Olson, L. E. & Martin, J. G. A. (2010). Heritability of anti‐predatory traits: vigilance and locomotor performance in marmots. Journal of Evolutionary Biology 23, 879–887. [DOI] [PubMed] [Google Scholar]
- Bocedi, G. , Heinonen, J. & Travis, J. M. J. (2012). Uncertainty and the role of information acquisition in the evolution of context‐dependent emigration. American Naturalist 179, 606–620. [DOI] [PubMed] [Google Scholar]
- * Boeye, J. , Travis, J. M. J. , Stoks, R. & Bonte, D. (2013). More rapid climate change promotes evolutionary rescue through selection for increased dispersal distance. Evolutionary Applications 6, 353–364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolnick, D. I. , Snowberg, L. K. , Patenia, C. , Stutz, W. E. , Ingram, T. & Lau, O. L. (2009). Phenotype‐dependent native habitat preference facilitates divergence between parapatric lake and stream stickleback. Evolution 63, 2004–2016. [DOI] [PubMed] [Google Scholar]
- Bonduriansky, R. & Day, T. (2009). Nongenetic inheritance and its evolutionary implications. Annual Reviews in Ecology, Evolution and Systematics 40, 103–125. [Google Scholar]
- Bonte, D. , Bossuyt, B. & Lens, L. (2007). Aerial dispersal plasticity under different wind velocities in a salt marsh wolf spider. Behavioral Ecology 18, 438–443. [Google Scholar]
- Bonte, D. & Dahirel, M. (2017). Dispersal: a central and independent trait in life history. Oikos 126, 472–479. [Google Scholar]
- Bonte, D. & De La Peña, E. (2009). Evolution of body condition‐dependent dispersal in metapopulations. Journal of Evolutionary Biology 22, 1242–1251. [DOI] [PubMed] [Google Scholar]
- * Bonte, D. , Hovestadt, T. & Poethke, H. J. (2009). Sex‐specific dispersal and evolutionary rescue in metapopulations infected by male killing endosymbionts. BMC Evolutionary Biology 9, 16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- * Bonte, D. , Hovestadt, T. & Poethke, H. J. (2010). Evolution of dispersal polymorphism and local adaptation of dispersal distance in spatially structured landscapes. Oikos 119, 560–566. [Google Scholar]
- Bonte, D. , Travis, J. M. J. , De Clercq, N. , Zwertvaegher, I. & Lens, L. (2008). Thermal conditions during juvenile development affect adult dispersal in a spider. Proceedings of the National Academy of Sciences of the United States of America 105, 17000–17005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonte, D. , Vanden Borre, J. , Lens, L. & Maelfait, J. P. (2006). Geographical variation in wolf spider dispersal behaviour is related to landscape structure. Animal Behaviour 72, 655–662. [Google Scholar]
- Bonte, D. , Vandenbroecke, N. , Lens, L. & Maelfait, J.‐P. (2003). Low propensity for aerial dispersal in specialist spiders from fragmented landscapes. Proceedings of the Royal Society of London B 270, 1601–1607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonte, D. , Van Dyck, H. , Bullock, J. M. , Coulon, A. , Delgado, M. , Gibbs, M. , Lehouck, V. , Matthysen, E. , Mustin, K. , Saastamoinen, M. , Schtickzelle, N. , Stevens, V. M. , Vandewoestijne, S. , Baguette, M. , Barton, K. , et al. (2012). Costs of dispersal. Biological Reviews 87, 290–312. [DOI] [PubMed] [Google Scholar]
- * Boonstra, R. & Hochachka, M. (1997). Maternal effects and additive genetic inheritance in the collared lemming Dicrostonyx groenlandicus . Evolutionary Ecology 11, 169–182. [Google Scholar]
- Bowler, D. E. & Benton, T. G. (2005). Causes and consequences of animal dispersal strategies: relating individual behaviour to spatial dynamics. Biological Reviews 80, 205–225. [DOI] [PubMed] [Google Scholar]
- Bradford, M. J. & Taylor, G. C. (1997). Individual variation in dispersal behaviour of newly emerged Chinook salmon (Oncorhynchus tshawytscha) from the upper Fraser River, British Columbia. Canadian Journal of Fisheries and Aquatic Sciences 54, 1585–1592. [Google Scholar]
- Brown, C. T. , Fishwick, L. K. , Chokshi, B. M. , Cuff, M. A. , Jackson, J. M. IV , Oglesby, T. , Rioux, A. T. , Rodriguez, E. , Stupp, G. S. , Trupp, A. H. , Woollcombe‐Clarke, J. S. , Wright, T. N. , Zaragoza, W. J. , Drew, J. C. , Triplett, E. W. & Nicholson, W. L. (2011). Whole‐genome sequencing and phenotypic analysis of Bacillus subtilis mutants following evolution under conditions of relaxed selection for sporulation. Applied and Environmental Microbiology 77, 6867–6877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- * Brown, G. P. , Phillips, B. L. , Dubey, S. & Shine, R. (2014). Invader immunology: invasion history alters immune system function in cane toads (Rhinella marina) in tropical Australia. Ecology Letters 18, 57–65. [DOI] [PubMed] [Google Scholar]
- * Brown, G. P. & Shine, R. (2014). Immune response varies with rate of dispersal in invasive cane toads (Rhinella marina). PloS One 6, e99734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burgess, S. C. , Baskett, M. L. , Grosberg, R. K. , Morgan, S. G. & Strathmann, R. R. (2015). When is dispersal for dispersal? Unifying marine and terrestrial perspectives. Biological Reviews 91, 867–882. [DOI] [PubMed] [Google Scholar]
- Burns, J. G. , Svetec, N. , Rowe, L. , Mery, F. , Dolan, M. J. , Boyce, W. T. & Sokolowski, M. B. (2012). Gene‐environment interplay in Drosophila melanogaster: chronic food deprivation in early life affects adult exploratory and fitness traits. Proceedings of the National Academy of Sciences of the United States of America 109, 17239–17244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burton, O. J. , Phillips, B. L. & Travis, J. M. J. (2010). Trade‐offs and the evolution of life‐histories during range expansion. Ecology Letters 13, 1210–1220. [DOI] [PubMed] [Google Scholar]
- Byers, J. E. (2000). Effects of body size and resource availability on dispersal in a native and a non‐native estuarine snail. Journal of Experimental Marine Biology and Ecology 248, 133–150. [DOI] [PubMed] [Google Scholar]
- * Cadet, C. , Ferrière, R. , Metz, J. A. J. & van Baalen, M. (2003). The evolution of dispersal under demographic stochasticity. American Naturalist 162, 427–441. [DOI] [PubMed] [Google Scholar]
- Caillaud, M. C. , Boutin, M. , Braendle, C. & Simon, J. C. (2002). A sex‐linked locus controls wing polymorphism in males of the pea aphid, Acyrthosiphon pisum (Harris). Heredity 89, 346–352. [DOI] [PubMed] [Google Scholar]
- * Chaine, A. S. , Legendre, S. & Clobert, J. (2013). The co‐evolution of multiply‐informed dispersal: information transfer across landscapes from neighbors and immigrants. PeerJ 1, e44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charmantier, A. , Buoro, M. , Gimenez, O. & Weimerskirch, H. (2011). Heritability of short‐scale natal dispersal in a large‐scale foraging bird, the wandering albatross. Journal of Evolutionary Biology 24, 1487–1496. [DOI] [PubMed] [Google Scholar]
- * Charmantier, A. , Keyser, A. J. & Promislow, D. E. L. (2007). First evidence for heritable variation in cooperative breeding behaviour. Proceedings of the Royal Society of London B 274, 1757–1761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheptou, P.‐O. , Carrue, O. , Rouifed, S. & Cantarel, A. (2008). Rapid evolution of seed dispersal in an urban environment in the weed Crepis sancta . Proceedings of the National Academy of Sciences of the United States of America 105, 3796–3799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheptou, P-O , Hargreaves, A. L , Bonte, D . & Jacquemyn, H . (2017). Adaptation to fragmentation: evolutionary dynamics driven by human influences. Phil. Trans. R. Soc. B 372: 20160037 http://dx.doi.org/10.1098/rstb.2016.0037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheptou, P.‐O. & Massol, F. (2009). Pollination fluctuations drive evolutionary syndromes linking dispersal and mating system. American Naturalist 174, 46–55. [DOI] [PubMed] [Google Scholar]
- Clobert, J. , Baguette, M. , Benton, T. & Bullock, J. (2012). Dispersal Ecology and Evolution. Oxford University Press, Oxford. [Google Scholar]
- Clobert, J. , Le Galliard, J.‐F. , Cote, J. , Meylan, S. & Massot, M. (2009). Informed dispersal, heterogeneity in animal dispersal syndromes and dynamics of spatially structures populations. Ecology Letters 12, 197–209. [DOI] [PubMed] [Google Scholar]
- * Cobben, M. M. P. , Verboom, J. , Opdam, P. F. M. & Hoekstra, R. F. (2015). Spatial sorting and range shifts: consequences for evolutionary potential and genetic signature of a dispersal trait. Journal of Theoretical Biology 373, 92–99. [DOI] [PubMed] [Google Scholar]
- Comins, H. , Hamilton, W. & May, R. (1980). Evolutionarily stable dispersal strategies. Journal of Theoretical Biology 82, 205–230. [DOI] [PubMed] [Google Scholar]
- Coppock, A. G. , Gardiner, N. M. & Jones, G. P. (2016). Sniffing out the competition? Juvenile coral reef damselfishes use chemical cues to distinguish the presence of conspecific and heterospecific aggregations. Behavioural Processes 125, 43–50. [DOI] [PubMed] [Google Scholar]
- Cote, J. , Bestion, E. , Jacob, S. , Travis, J. , Legrand, D. & Baguette, M. (2017a). Evolution of dispersal strategies and dispersal syndromes in fragmented landscapes. Ecography 40, 56–73. [Google Scholar]
- Cote, J. , Bocedi, G. , Debeffe, L. , Chudzińska, M. E. , Weigang, H. C. , Dytham, C. , Gonzalez, G. , Matthysen, E. , Travis, J. M. J. , Baguette, M. & Hewison, A. J. M. (2017b). Behavioural synchronization of large‐scale animal movements – disperse alone, but migrate together? Biological Reviews 92, 1275–1296. https://doi.org/10.1111/brv.12279. [DOI] [PubMed] [Google Scholar]
- Cote, J. & Clobert, J. (2007). Social personalities influence natal dispersal in a lizard. Proceedings of the Royal Society of London B 274, 383–390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cote, J. & Clobert, J. (2010). Risky dispersal: avoiding kin competition despite uncertainty. Ecology 91, 1485–1493. [DOI] [PubMed] [Google Scholar]
- Cote, J. , Clobert, J. , Brodin, T. , Fogarty, S. & Sih, A. (2010). Personality‐dependent dispersal: characterization, ontogeny and consequences for spatially structured populations. Philosophical Transactions of the Royal Society B: Biological Sciences 365, 4065–4076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cote, J. , Fogarty, S. , Tymen, B. , Sih, A. & Brodin, T. (2013). Personality‐dependent dispersal cancelled under predation risk. Proceedings of the Royal Society of London B 280, 20132349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- * Cotto, O. , Olivieri, I. & Ronce, O. (2013). Optimal life‐history schedule in a metapopulation with juvenile dispersal. Journal of Evolutionary Biology 26, 944–954. [DOI] [PubMed] [Google Scholar]
- Cousens, R. D. , Dytham, C. & Law, R. (2008). Dispersal in Plants: A Population Perspective. Oxford University Press, Oxford. [Google Scholar]
- * Crespi, B. & Taylor, P. (1990). Dispersal rates under variable patch density. American Naturalist 135, 48–62. [Google Scholar]
- Crespi, B. J. (2004). Vicious circles: positive feedback in major evolutionary and ecological transitions. Trends in Ecology and Evolution 19, 627–633. [DOI] [PubMed] [Google Scholar]
- Crnokrak, P. & Roff, D. A. (1998). The genetic basis of the trade‐off between calling and wing morph in males of the cricket, Gryllus firmus . Evolution 52, 1111–1118. [DOI] [PubMed] [Google Scholar]
- Crnokrak, P. & Roff, D. A. (2002). Trade‐offs to flight capability in Gryllus firmus: the influence of whole‐organism respiration rate on fitness. Journal of Evolutionary Biology 15, 388–398. [Google Scholar]
- Cwynar, L. C. & MacDonald, G. M. (1987). Geographical variation of Lodgepole pine in relation to population history. American Naturalist 129, 463–469. [Google Scholar]
- Danchin, É. , Charmantier, A. , Champagne, F. A. , Mesoudi, A. , Pujol, B. & Blanchet, S. (2011). Beyond DNA: integrating inclusive inheritance into an extended theory of evolution. Nature Reviews Genetics 12, 475–486. [DOI] [PubMed] [Google Scholar]
- Débarre, F. , Yeaman, S. & Guillaume, F. (2015). Evolution of quantitative traits under a migration‐selection balance: when does skew matter? American Naturalist 186, S37–S47. [DOI] [PubMed] [Google Scholar]
- Debeffe, L. , Morellet, N. , Verheyden‐Tixier, H. , Hoste, H. , Gaillard, J.‐M. , Cargnelutti, B. , Picot, D. , Sevila, J. & Hewison, A. J. M. (2014). Parasite abundance contributes to condition‐dependent dispersal in a wild population of large herbivore. Oikos 123, 1121–1125. [Google Scholar]
- Delgado, M. M. , Bartoń, K. A. , Bonte, D. & Travis, J. M. J. (2014). Prospecting and dispersal: their eco‐evolutionary dynamics and implications for population patterns. Proceedings of the Royal Society of London B 281, 20132851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delgado, M. M. & Penteriani, V. (2008). Behavioral states help translate dispersal movements into spatial distribution patterns of floaters. American Naturalist 172, 475–485. [DOI] [PubMed] [Google Scholar]
- Delgado, M. M. , Penteriani, V. , Revilla, E. & Nams, V. O. (2010). The effect of phenotypic traits and external cues on natal dispersal movements. Journal of Animal Ecology 79, 620–632. [DOI] [PubMed] [Google Scholar]
- De Roissart, A. , Wang, S. & Bonte, D. (2015). Spatial and spatiotemporal variation in metapopulation structure affects population dynamics in a passively dispersing arthropod. Journal of Animal Ecology 84, 1565–1574. [DOI] [PubMed] [Google Scholar]
- De Roissart, A. , Wybouw, N. , Renault, D. , Van Leeuwen, T. & Bonte, D. (2016). Life history evolution in response to changes in metapopulation structure in an arthropod herbivore. Functional Ecology 30, 1408–1417. [Google Scholar]
- De Wet, J. M. J. & Harlan, J. R. (1975). Weeds and domesticates: evolution in the man‐made habitat. Economic Botany 29, 99–107. [Google Scholar]
- Dingle, H. (2014). Migration: The Biology of Life on the Move, Second Edition. Oxford University Press, Oxford. [Google Scholar]
- Doligez, B. , Daniel, G. , Warin, P. , Pärt, T. , Gustafsson, L. & Réale, D. (2012). Estimation and comparison of heritability and parent–offspring resemblance in dispersal probability from capture–recapture data using different methods: the collared flycatcher as a case study. Journal of Ornithology 152, 539–554. [Google Scholar]
- Doligez, B. & Pärt, T. (2008). Estimating fitness consequences of dispersal: a road to 'know‐where'? Non‐random dispersal and the underestimation of dispersers' fitness. Journal of Animal Ecology 77, 1199–1211. [DOI] [PubMed] [Google Scholar]
- Dong, Y. , Yang, X. , Liu, J. , Wang, B.‐H. , Liu, B.‐L. & Wang, Y.‐Z. (2014). Pod shattering resistance associated with domestication is mediated by a NAC gene in soybean. Nature Communications 5, 3352. [DOI] [PubMed] [Google Scholar]
- Donohue, K. (1998). Maternal determinants of seed dispersal in Cakile edentula: fruit, plant and site traits. Ecology 79, 2771–2788. [Google Scholar]
- Donohue, K. (1999). Seed dispersal as a maternally‐influenced character: mechanistic basis of maternal effects and selection on maternal characters in an annual plant. American Naturalist 154, 674–689. [DOI] [PubMed] [Google Scholar]
- Donohue, K. (2005). Niche construction through phenological plasticity: life history dynamics and ecological consequences. New Phytologist 166, 83–92. [DOI] [PubMed] [Google Scholar]
- Donohue, K. , Polisetty, C. R. & Wender, N. J. (2005). Genetic basis and consequences of niche construction: plasticity‐induced genetic constraints on the evolution of seed dispersal in Arabidopsis thaliana . American Naturalist 165, 537–550. [DOI] [PubMed] [Google Scholar]
- Drangsholt, T. M. K. , Damsgard, B. & Olesen, I. (2014). Quantitative genetics of behavioral responsiveness in Atlantic cod (Gadus morhua L.). Aquacultures 420–421, 282–287. [Google Scholar]
- Ducatez, S. , Baguette, M. , Stevens, V. M. , Legrand, D. & Freville, H. (2012). Complex interactions between paternal and maternal effects: parental experience and age at reproduction affect fecundity and offspring performance in a butterfly. Evolution 66, 3558–3569. [DOI] [PubMed] [Google Scholar]
- Duckworth, R. A. & Badyaev, A. V. (2007). Coupling of dispersal and aggression facilitates the rapid range expansion of a passerine bird. Proceedings of the National Academy of Sciences of the United States of America 104, 15017–15022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duckworth, R. A. & Kruuk, L. E. B. (2009). Evolution of genetic integration between dispersal and colonization ability in a bird. Evolution 63, 968–977. [DOI] [PubMed] [Google Scholar]
- Dunlop, E. S. , Heino, M. & Dieckmann, U. (2009). Eco‐genetic modeling of contemporary life‐history evolution. Ecological Applications 19, 1815–1834. [DOI] [PubMed] [Google Scholar]
- Duong, T. Y. , Scribner, K. T. , Crossman, J. A. , Forsythe, P. S. & Baker, E. A. (2011). Environmental and maternal effects on embryonic and larval developmental time until dispersal of lake sturgeon (Acipenser fulvescens). Canadian Journal of Fisheries and Aquatic Sciences 68, 643–654. [Google Scholar]
- Duputié, A. , Massol, F. , Chuine, I. , Kirkpatrick, M. & Ronce, O. (2012). How do genetic correlations affect species range shifts in a changing environment? Ecology Letters 15, 251–259. [DOI] [PubMed] [Google Scholar]
- Duthie, A. B. , Bocedi, G. & Reid, J. M. (2016). When does female multiple mating evolve to adjust inbreeding? Effects of inbreeding depression, direct costs, mating constraints, and polyandry as a threshold trait. Evolution 70, 1927–1943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- * Dytham, C. (2009). Evolved dispersal strategies at range margins. Proceedings of the Royal Society of London B 276, 1407–1413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edelsparre, A. H. , Lim, J. H. , Anwari, M. & Fitzpatrick, M. J. (2014). Alleles underlying larval foraging behaviour influence adult dispersal in nature. Ecology Letters 17, 333–339. [DOI] [PubMed] [Google Scholar]
- Enfjäll, K. & Leimar, O. (2009). The evolution of dispersal – the importance of information about population density and habitat characteristics. Oikos 118, 291–299. [Google Scholar]
- Etterson, J. (2004). Evolutionary potential of Chamaecrista fasciculate in relation to climate change. II. Genetic architecture of three populations reciprocally planted along an environmental gradient in the great plains. Evolution 58, 1459–1471. [DOI] [PubMed] [Google Scholar]
- Evans, J. P. , García‐González, F. & Marshall, D. J. (2007). Sources of genetic and phenotypic variance in fertilization rates and larval traits in a sea urchin. Evolution 61, 2832–2838. [DOI] [PubMed] [Google Scholar]
- Falconer, D. S. & Mackay, T. F. C. (1996). Introduction to Quantitative Genetics, Fourth Edition. Longman Group Limited, Essex. [Google Scholar]
- Fitzpatrick, M. J. , Ben‐Shahar, Y. , Smid, H. M. , Vet, L. E. M. , Robinson, G. E. & Sokolowski, M. B. (2005). Candidate genes for behavioural ecology. Trends in Ecology and Evolution 20, 96–104. [DOI] [PubMed] [Google Scholar]
- Fountain, T. , Nieminen, M. , Siren, J. , Wong, S. C. & Hanski, I. (2016). Predictable allele frequency changes due to habitat fragmentation in the Glanville fritillary butterfly. Proceedings of the National Academy of Sciences of the United States of America 113, 2678–2683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox, C. W. & Wolf, J. B. (2006). Evolutionary Genetics: Concepts and Case Studies. Oxford University Press, Oxford. [Google Scholar]
- Frank, S. A. (1986). Dispersal polymorphisms in subdivided populations. Journal of Theoretical Biology 122, 303–309. [DOI] [PubMed] [Google Scholar]
- Freeman, S. & Herron, J. C. (2004). Evolutionary Analyses. Pearson Education Inc., Upper Saddle River, N.J. [Google Scholar]
- Friedenberg, N. A. (2003). Experimental evolution of dispersal in spatiotemporally variable microcosms. Ecology Letters 6, 953–959. [Google Scholar]
- Fronhofer, E. A. & Altermatt, F. (2015). Eco‐evolutionary feedbacks during experimental range expansions. Nature Communications 6, 6844. https://doi.org/10.1038/ncomms7844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fronhofer, E. A. , Kropf, T. & Altermatt, F. (2015a). Density‐dependent movement and the consequences of the Allee effect in the model organism Tetrahymena . Journal of Animal Ecology 84, 712–722. [DOI] [PubMed] [Google Scholar]
- Fronhofer, E. A. , Poethke, H. J. & Dieckmann, U. (2015b). Evolution of dispersal distance: maternal investment leads to bimodal dispersal kernels. Journal of Theoretical Biology 365, 270–279. [DOI] [PubMed] [Google Scholar]
- Fronhofer, E. A. , Kubisch, A. , Hovestadt, T. & Poethke, H. J. (2011). Assortative mating and the evolution of dispersal polymorphisms. Evolution 65, 2461–2469. [DOI] [PubMed] [Google Scholar]
- Fronhofer, E. A. , Nitsche, N. & Altermatt, F. (2017). Information use shapes range expansion dynamics into environmental gradients. Global Ecology and Biogeography 26, 400–411. [Google Scholar]
- Fronhofer, E. A. , Stelz, J. M. , Lutz, E. , Poethke, H. J. & Bonte, D. (2014). Spatially correlated extinctions select for less emigration but larger dispersal distances in the spider mite Tetranychus urticae . Evolution 68, 1838–1844. [DOI] [PubMed] [Google Scholar]
- Fuller, D. Q. & Allaby, R. (2009). Seed dispersal and crop domestication: shattering, germination, and seasonality in evolution under cultivation. Annual Plant Reviews 38, 238–295. [Google Scholar]
- Futuyma, D. J. (2010). Evolutionary constraint and ecological consequences. Evolution 64, 1865–1884. [DOI] [PubMed] [Google Scholar]
- Galetti, M. , Guevara, R. , Côrtes, M. C. , Fadini, R. , VonMatter, S. , Leite, A. B. , Labecca, F. , Ribeiro, T. , Carvalho, C. S. , Collevatti, R. G. , Pires, M. M. , Guimarães, P. R. Jr. , Brancalion, P. H. S. , Ribeiro, M. C. & Jordano, P. (2013). Functional extinction of birds drives rapid evolutionary changes in seed size. Science 340, 1086–1090. [DOI] [PubMed] [Google Scholar]
- Gandon, S. (1999). Kin competition, the cost of inbreeding and the evolution of dispersal. Journal of Theoretical Biology 200, 345–364. [DOI] [PubMed] [Google Scholar]
- Gandon, S. & Michalakis, Y. (1999). Evolutionarily stable dispersal rate in a metapopulation with extinctions and kin competition. Journal of Theoretical Biology 199, 275–290. [DOI] [PubMed] [Google Scholar]
- * Garenc, C. , Silversides, F. G. & Guderley, H. (1998). Burst swimming and its enzymatic correlates in the threespine stickleback (Gasterosteus aculeatus): full‐sib heritabilities. Canadian Journal of Zoology 76, 680–688. [Google Scholar]
- Garland, T. Jr. (1990). Locomotor performance and activity metabolism of Cnemidophorus tigris in relation to natural behavior In Biology of Cnemidophorus (ed. Wright J.), pp. 163–210. Los Angeles County Museum and University of Washington Press, Los Angeles. [Google Scholar]
- Gatehouse, A. G. (1989). Genes, environment and insect flight In Insect Flight (eds Goldworthy G. J. and Wheeler C. H.), pp. 115–138. CRC Press, Boca Raton. [Google Scholar]
- Gatehouse, A. G. (1997). Behavior and ecological genetics of wind‐borne migration by insects. Annual Review of Entomology 42, 475–502. [DOI] [PubMed] [Google Scholar]
- Gibbs, M. & van Dyck, H. (2010). Butterfly flight activity affects reproductive performance and longevity relative to landscape structure. Oecologia 163, 341–350. [DOI] [PubMed] [Google Scholar]
- Gilbert, K. J. & Whitlock, M. C. (2017). The genetics of adaptation to discrete heterogeneous environments: frequent mutation or large‐effect alleles can allow range expansion. Journal of Evolutionary Biology 30, 591–602. https://doi.org/10.1111/jeb.13029. [DOI] [PubMed] [Google Scholar]
- Gilliam, J. F. & Fraser, D. F. (2001). Movement in corridors: enhancement by predation threat, disturbance, and habitat structure. Ecology 82, 258–273. [Google Scholar]
- Gioia, T. , Logozzo, G. , Kami, J. , Spagnoletti Zeuli, P. & Gepts, P. (2013). Identification and characterization of a homologue to the Arabidopsis INDEHISCENT gene in common bean. Journal of Heredity 104, 273–286. [DOI] [PubMed] [Google Scholar]
- Girgis, H. S. , Liu, Y. , Ryu, W. S. & Tavazoie, S. (2007). A comprehensive genetic characterization of bacterial motility. PLoS Genetics 3, e154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomez, G. S. Y. & Van Dyck, H. (2012). Ecotypic differentiation between urban and rural populations of the grasshopper Chorthippus brunneus relative to climate and habitat fragmentation. Oecologia 169, 125–133. [DOI] [PubMed] [Google Scholar]
- Gomulkiewicz, R. , Holt, R. D. , Barfield, M. & Nuismer, S. L. (2010). Genetics, adaptation, and invasion in harsh environments. Evolutionary Applications 3, 97–108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goodacre, S. , Martin, O. , Bonte, D. , Hutchings, L. , Woolley, C. , Ibrahim, K. , Hewitt, G. M. & Thomas, C. F. G. (2009). Microbial modification of host long‐distance dispersal capacity. BMC Biology 7, 32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grafen, A. (1984). Natural selection, kin selection, and group dispersal In Behavioural Ecology: An Evolutionary Approach (eds Krebs J. R. and Davies N. B.), pp. 62–84. Blackwell, Oxford. [Google Scholar]
- Greene, D. F. (2005). The role of abscission in long‐distance seed dispersal by the wind. Ecology 86, 3105–3110. [Google Scholar]
- * Greenwood, P. J. , Harvey, P. H. & Perrins, C. M. (1979). Kin selection and territoriality in birds? A test. Animal Behaviour 27, 645–651. [Google Scholar]
- Gros, A. , Hovestadt, T. & Poethke, H. J. (2008). Evolution of sex‐biased dispersal: the role of sex‐specific dispersal costs, demographic stochasticity, and inbreeding. Ecological Modelling 219, 226–233. [Google Scholar]
- Gros, A. , Poethke, H. J. & Hovestadt, T. (2009). Sex‐specific spatio‐temporal variability in reproductive success promotes the evolution of sex‐biased dispersal. Theoretical Population Biology 76, 13–18. [DOI] [PubMed] [Google Scholar]
- Gu, H. & Danthanarayana, W. (1992). Quantitative genetic analysis of dispersal in Epiphyas postvittana. I. Genetic variation in flight capacity. Heredity 68, 53–60. [Google Scholar]
- Guillaume, F. (2011). Migration‐induced phenotypic divergence: the migration‐selection balance of correlated traits. Evolution 65, 1723–1738. [DOI] [PubMed] [Google Scholar]
- Guillaume, F. & Perrin, N. (2006). Joint evolution of dispersal and inbreeding load. Genetics 173, 497–509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guillaume, F. & Perrin, N. (2009). Inbreeding load, bet hedging, and the evolution of sex biased dispersal. American Naturalist 173, 536–541. [DOI] [PubMed] [Google Scholar]
- Gyllenberg, M. , Parvinen, K. & Dieckmann, U. (2002). Evolutionary suicide and evolution of dispersal in structured metapopulations. Journal of Mathematical Biology 45, 79–105. [DOI] [PubMed] [Google Scholar]
- Haag, C. R. , Riek, M. , Hottinger, J. W. , Pajunen, V. I. & Ebert, D. (2005). Genetic diversity and genetic differentiation in Daphnia metapopulations with subpopulations of known age. Genetics 170, 1809–1820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamilton, W. & May, R. M. (1977). Dispersal in stable habitats. Nature 269, 578–581. [Google Scholar]
- * Han, L. Z. , Gu, H. N. & Zhang, X. X. (2009). Genetic effects on flight capacity in the beet armyworm, Spodoptera exigua (Lep., Noctuidae). Journal of Applied Entomology 133, 262–271. [Google Scholar]
- Hansen, T. F. (2006). The evolution of genetic architecture. Annual Review in Ecology, Evolution and Systematics 37, 123–157. [Google Scholar]
- Hansen, T. F. (2013). Why epistasis is important for selection and adaptation. Evolution 67, 3501–3511. [DOI] [PubMed] [Google Scholar]
- Hanski, I. (2011). Eco‐evolutionary spatial dynamics in the Glanville fritillary butterfly. Proceedings of the National Academy of Sciences of the United States of America 108, 14397–14404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanski, I. (2012). Eco‐evolutionary dynamics in a changing world. Annals of the New York Academy of Sciences 1249, 1–17. [DOI] [PubMed] [Google Scholar]
- Hanski, I. , Erälahti, C. , Kankare, M. , Ovaskainen, O. & Sirén, H. (2004). Variation in migration rate among individuals maintained by landscape structure. Ecology Letters 7, 958–966. [Google Scholar]
- * Hanski, I. & Mononen, T. (2011). Eco‐evolutionary dynamics of dispersal in spatially heterogeneous environments. Ecology Letters 14, 1025–1034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanski, I. , Schulz, T. , Chong, S. , Ahola, V. , Ruokolainen, A. & Ojanen, S. P. (2017). Ecological and genetic basis of metapopulation persistence of the Glanville fritillary butterfly in fragmented landscapes. Nature Communications 8, 14504. https://doi.org/10.1038/ncomms14504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- * Hansson, B. , Bensch, S. & Hasselquist, D. (2003). A new approach to study dispersal: immigration of novel alleles reveals female‐biased dispersal in great reed warbles. Molecular Ecology 12, 631–637. [DOI] [PubMed] [Google Scholar]
- Hargreaves, A. L. , Bailey, S. F. & Laird, R. A. (2015). Fitness declines towards range limits and local adaptation to climate affect dispersal evolution during climate‐induced range shifts. Journal of Evolutionary Biology 28, 1489–1501. [DOI] [PubMed] [Google Scholar]
- Hastings, A. (1983). Can spatial variation alone lead to selection for dispersal? Theoretical Population Biology 24, 244–251. [Google Scholar]
- Heino, M. & Hanski, I. (2001). Evolution of migration rate in a spatially realistic metapopulation model. American Naturalist 157, 495–511. [DOI] [PubMed] [Google Scholar]
- Heinz, S. K. & Strand, E. (2006). Adaptive patch searching strategies in fragmented landscapes. Evolutionary Ecology 20, 113–130. [Google Scholar]
- Hendry, A. P. (2013). Key questions in the genetics and genomics of eco‐evolutionary dynamics. Heredity 111, 456–466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hendry, A. P. (2016). Eco‐Evolutionary Dynamics. Princeton University Press, Princeton. [Google Scholar]
- * Henriques‐Silva, R. , Boivin, F. , Calcagno, V. , Urban, M. C. & Peres‐Neto, P. R. (2015). On the evolution of dispersal via heterogeneity in spatial connectivity. Proceedings of the Royal Society of London B 282, 20142879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- * Henry, R. C. , Bocedi, G. , Dytham, C. & Travis, J. M. J. (2014). Inter‐annual variability influences the eco‐evolutionary dynamics of range‐shifting. PeerJ 1, e228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henry, R. C. , Bocedi, G. & Travis, J. M. J. (2013). Eco‐evolutionary dynamics of range shifts: elastic margins and critical thresholds. Journal of Theoretical Biology 321, 1–7. [DOI] [PubMed] [Google Scholar]
- Henry, R. C. , Coulon, A. & Travis, J. M. J. (2016). The evolution of male biased dispersal under the joint selective forces of inbreeding load and demographic and environmental stochasticity. American Naturalist 188 https://doi.org/10.1086/688170. [DOI] [PubMed] [Google Scholar]
- Higgins, K. & Lynch, M. (2001). Metapopulation extinction caused by mutation accumulation. Proceedings of the National Academy of Sciences of the United States of America 98, 2928–2933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- * Hilbish, T. J. , Sasada, K. , Eyster, L. S. & Pechenik, J. A. (1999). Relationship between rates of swimming and growth in veliger larvae: genetic variance and covariance. Journal of Experimental Marine Biology and Ecology 239, 183–193. [Google Scholar]
- Hill, J. K. , Collingham, Y. C. , Thomas, C. D. , Blakeley, D. S. , Fox, R. , Moss, D. & Huntley, B. (2001). Impacts of landscape structure on butterfly range expansion. Ecology Letters 4, 313–321. [Google Scholar]
- Hill, J. K. , Thomas, C. D. & Blakeley, D. S. (1999). Evolution of flight morphology in a butterfly that has recently expanded its geographic range. Oecologia 121, 165–170. [DOI] [PubMed] [Google Scholar]
- Hoffmann, A. A. & Willi, Y. (2008). Detecting genetic responses to environmental change. Nature Reviews Genetics 9, 421–432. [DOI] [PubMed] [Google Scholar]
- * Holt, R. & McPeek, M. A. (1996). Chaotic population dynamics favors the evolution of dispersal. American Naturalist 148, 709–718. [Google Scholar]
- Holt, R. D. (2003). On the evolutionary ecology of species' range. Evolutionary Ecology Research 5, 159–178. [Google Scholar]
- Houck, M. A. & O'Connor, B. M. (1991). Ecological and evolutionary significance of phoresy in the Astigmata. Annual Review of Entomology 36, 611–636. [Google Scholar]
- Hovestadt, T. , Messner, S. & Poethke, H. J. (2001). Evolution of reduced dispersal mortality and “fat‐tailed” dispersal kernels in autocorrelated landscapes. Proceedings of the Royal Society of London B 268, 385–391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang, F. , Peng, S. , Chen, B. , Liao, H. , Huang, Q. , Lin, Z. & Liu, G. (2015). Rapid evolution of dispersal‐related traits during range expansion of an invasive vine Mikania micrantha . Oikos 124, 1023–1030. [Google Scholar]
- Husby, A. , Kawakami, T. , Rönnegård, L. , Smeds, L. , Ellegren, H. & Qvarnström, A. (2015). Genome‐wide association mapping in a wild avian population identifies a link between genetic and phenotypic variation in a life history trait. Proceedings of the Royal Society of London B 282, 1806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imbert, E. (2001). Capitulum characters in a seed heteromorphic plant, Crepis sancta (Asteraceae): variance partitioning and inference for the evolution of dispersal rate. Heredity 86, 78–86. [DOI] [PubMed] [Google Scholar]
- Imbert, E. & Ronce, O. (2001). Phenotypic plasticity for dispersal ability in the seed heteromorphic Crepis sancta (Asteraceae). Oikos 93, 126–134. [Google Scholar]
- Iritani, R. & Iwasa, Y. (2014). Parasite infection drives the evolution of state‐dependent dispersal of the host. Theoretical Population Biology 92, 1–13. [DOI] [PubMed] [Google Scholar]
- Jacob, S. , Wehi, P. , Clobert, J. , Legrand, D. , Schtickzelle, N. , Huet, M. & Chaine, A. (2016). Cooperation‐mediated plasticity in dispersal and colonization. Evolution 70, 2336–2345. [DOI] [PubMed] [Google Scholar]
- Johnson, J. R. , Johnson, B. B. & Bradley Shaffer, H. (2010). Genotype and temperature affect locomotor performance in a tiger salamander hybrid swarm. Functional Ecology 24, 1073–1080. [Google Scholar]
- Jones, C. M. , Papanicolaou, A. , Mironidis, G. K. , Vontas, J. , Yang, Y. , Lim, K. S. , Oakeshott, J. G. , Bass, C. & Chapman, J. W. (2015). Genomewide transcriptional signatures of migratory flight activity in a globally invasive insect pest. Molecular Ecology 24, 4901–4911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jordan, K. W. , Carbone, M. A. , Yamamoto, A. , Morgan, T. J. & Mackay, T. F. (2007). Quantitative genomics of locomotor behavior in Drosophila melanogaster . Genome Biology 8, R172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jordan, K. W. , Craver, K. L. , Magwire, M. M. , Cubilla, C. E. , Mackay, T. F. C. & Anholt, R. R. H. (2012). Genome‐wide association for sensitivity to chronic oxidative stress in Drosophila melanogaster . PLoS One 7, e38722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kardos, M. , Husby, A. , McFarlane, S. E. , Qvarnstrom, A. & Ellegren, H. (2016). Whole genome resequencing of extreme phenotypes in collared flycatchers highlights the difficulty of detecting quantitative trait loci in natural populations. Molecular Ecology Resources 16, 727–741. [DOI] [PubMed] [Google Scholar]
- Karl, I. & Fischer, K. (2008). Why get big in the cold? Towards a solution of a life‐history puzzle. Oecologia 155, 215–225. [DOI] [PubMed] [Google Scholar]
- Kawecki, J. T. (2008). Adaptation to marginal habitats. Annual Review in Ecology, Evolution and Systematics 39, 321–342. [Google Scholar]
- Kawecki, T. J. , Lenski, R. E. , Ebert, D. , Hollis, B. , Olivieri, I. & Whitlock, M. C. (2012). Experimental evolution. Trends in Ecology and Evolution 10, 547–560. [DOI] [PubMed] [Google Scholar]
- Keena, M. A. , Grinberg, P. S. & Wallner, W. E. (2007). Inheritance of female flight in Lymantria dispar (Lepidoptera: Lymantriidae). Environmental Entomology 36, 484–494. [DOI] [PubMed] [Google Scholar]
- * Keil, S. , Gu, H. & Dorn, S. (2001). Response of Cydia pomonella to selection on mobility: laboratory evaluation and field verification. Ecological Entomology 26, 495–501. [Google Scholar]
- Kelman, W. M. & Culvenor, R. A. (2003). The genetic correlation of panicle shattering with the intact rachilla form of seed retention in Phalaris aquatica L . Euphytica 130, 405–411. [Google Scholar]
- * Kent, J. W. & Rankin, M. A. (2001). Heritability and physiological correlates of migratory tendency in the grasshopper Melanoplus sanguinipes . Physiological Entomology 26, 371–380. [Google Scholar]
- Kirchoff, K. B. & Claßen‐Bockhoff, R. (2013). Inflorescences: concepts, function, development and evolution. Annals of Botany 112, 1471–1476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirkpatrick, M. (2010). How and why chromosome inversions evolve. PLoS Biology 8, e1000501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klemme, I. & Hanski, I. (2009). Heritability of and strong single gene (Pgi) effects on life‐history traits in the Glanville fritillary butterfly. Journal of Evolutionary Biology 22, 1944–1953. [DOI] [PubMed] [Google Scholar]
- Knülle, W. (1995). Expression of a dispersal trait in a guild of mites colonizing transient habitats. Evolutionary Ecology 9, 341–353. [Google Scholar]
- Konishi, S. , Izawa, T. , Lin, S. Y. , Ebana, K. , Fukuta, Y. , Sasaki, T. & Yano, M. (2006). A SNP caused loss of seed shattering during rice domestication. Science 312, 1392–1396. [DOI] [PubMed] [Google Scholar]
- Korsten, P. , Mueller, J. C. , Hermannstadter, C. , Bouwman, K. M. , Dingemanse, N. J. , Drent, P. J. , Liedvogel, M. , Matthysen, E. , van Oers, K. , van Overveld, T. , Patrick, S. C. , Quinn, J. L. , Sheldon, B. C. , Tinbergen, J. M. & Kempenaers, B. (2010). Association between DRD4 gene polymorphism and personality variation in great tits: a test across four wild populations. Molecular Ecology 19, 832–843. [DOI] [PubMed] [Google Scholar]
- Korsten, P. , van Overveld, T. , Adriaensen, F. & Matthysen, E. (2013). Genetic integration of local dispersal and exploratory behaviour in a wild bird. Nature Communications 4, 2362. [DOI] [PubMed] [Google Scholar]
- Koskella, B. , Taylor, T. B. , Bates, J. & Buckling, A. (2011). Using experimental evolution to explore natural patterns between bacterial motility and resistance to bacteriophages. The ISME Journal 5, 1809–1817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kubisch, A. , Degen, T. , Hovestadt, T. & Poethke, H. J. (2013). Predicting range shifts under global change: the balance between local adaptation and dispersal. Ecography 36, 873–882. [Google Scholar]
- Kubisch, A. , Holt, R. D. , Poethke, H. J. & Fronhofer, E. A. (2014). Where am I and why? Synthesizing range biology and the eco‐evolutionary dynamics of dispersal. Oikos 123, 5–22. [Google Scholar]
- Kubisch, A. , Hovestadt, T. & Poethke, H. J. (2010). On the elasticity of range limits during periods of expansion. Ecology 91, 3094–3099. [DOI] [PubMed] [Google Scholar]
- Kun, A. & Scheuring, I. (2006). The evolution of density‐dependent dispersal in a noisy spatial population model. Oikos 115, 308–320. [Google Scholar]
- Lande, R. (1979). Quantitative genetic analysis of multivariate evolution, applied to brain: body size allometry. Evolution 33, 402–416. [DOI] [PubMed] [Google Scholar]
- * Larios, E. & Venable, D. L. (2015). Maternal adjustment of offspring provisioning and the consequences for dispersal. Ecology 96, 2771–2780. [DOI] [PubMed] [Google Scholar]
- Le Corre, V. & Kremer, A. (2003). Genetic variability at neutral markers, quantitative trait loci and trait in a subdivided population under selection. Genetics 164, 1205–1219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Le Corre, V. & Kremer, A. (2012). The genetic differentiation at quantitative trait loci under local adaptation. Molecular Ecology 21, 1548–1566. [DOI] [PubMed] [Google Scholar]
- Legrand, D. , Cote, J. , Fronhofer, E. A. , Holt, R. D. , Ronce, O. , Schtickzelle, N. , Travis, J. M. J. & Clobert, J. (2017). Eco‐evolutionary dynamics in fragmented landscapes. Ecography 40, 9–25. [Google Scholar]
- Legrand, D. , Larranaga, N. , Bertrand, R. , Ducatez, S. , Calvez, O. , Stevens, V. M. & Baguette, M. (2016). Evolution of a butterfly dispersal syndrome. Proceedings of the Royal Society of London B 283, 20161533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leimar, O. & Norberg, U. (1997). Metapopulation extinction and genetic variation in dispersal‐related traits. Oikos 80, 448–458. [Google Scholar]
- Lenser, T. & Theißen, G. (2013). Conservation of fruit dehiscence pathways between Lepidium campestre and Arabidopsis thaliana sheds light. The Plant Journal 76, 545–556. [DOI] [PubMed] [Google Scholar]
- Leturque, H. & Rousset, F. (2003). Joint evolution of sex ratio and dispersal: conditions for higher dispersal rates from good habitats. Evolutionary Ecology 17, 67–84. [Google Scholar]
- Levin, S. A. , Cohen, D. & Hastings, A. (1984). Dispersal strategies in patchy environments. Theoretical Population Biology 26, 165–191. [Google Scholar]
- Li, J. & Margolies, D. C. (1993). Quantitative genetics of aerial dispersal behavior and life‐history traits in Tetranychus urticae . Heredity 70, 544–552. [Google Scholar]
- Li, J. & Margolies, D. C. (1994). Responses to direct and indirect selection on aerial dispersal behaviour in Tetranychus urticae . Heredity 72, 10–22. [Google Scholar]
- Li, L.‐F. & Olsen, K. M. (2016). To have and to hold: selection for seed and fruit retention during crop domestication. Current Topics in Developmental Biology 119, 63–109. [DOI] [PubMed] [Google Scholar]
- Li, W. & Gill, B. S. (2006). Multiple genetic pathways for seed shattering in the grasses. Functional and Integrative Genomics 6, 300–309. [DOI] [PubMed] [Google Scholar]
- Lindström, R. , Lindholm, P. , Kallijärvi, J. , Yu, L. Y. , Piepponen, T. P. , Arumäe, U. , Saarma, M. & Heino, T. I. (2013). Characterization of the structural and functional determinants of MANF/CDNF in Drosophila in vivo model. PLoS One 8, e73928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, J. , Wang, J. , Wang, H. , Wang, W. , Zhou, R. , Mei, D. , Cheng, H. , Yang, J. , Raman, H. & Hu, Q. (2016). Multigenic control of pod shattering resistance in Chinese rapeseed Germplasm revealed by genome‐wide association and linkage analyses. Frontiers in Plant Science 7, 1058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lobón, C. M. , Acuña, J. L. , López‐Álvarez, M. & Capitanio, F. L. (2011). Heritability of morphological and life history traits in a pelagic tunicate. Marine Ecology Progress Series 422, 145–154. [Google Scholar]
- Lomáscolo, S. B. , Levey, D. J. , Kimball, R. T. , Bolker, B. M. & Alborn, H. T. (2010). Dispersers shape fruit diversity in Ficus (Moraceae). Proceedings of the National Academy of Sciences of the United States of America 107, 14668–14672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lombaert, E. , Estoup, A. , Facon, B. , Joubard, B. , Grégoire, J. C. , Jannin, A. , Blin, A. & Guillemaud, T. (2014). Rapid increase in dispersal during range expansion in the invasive ladybird Harmonia axyridis . Journal of Evolutionary Biology 27, 508–517. [DOI] [PubMed] [Google Scholar]
- Lowe, W. H. , Likens, G. E. & Cosentino, B. J. (2006). Self‐organisation in streams: the relationship between movement behaviour and body condition in a head water salamander. Freshwater Biology 51, 2052–2062. [Google Scholar]
- Lynch, M. & Walsh, B. (1998). Genetics and Analysis of Quantitative Traits. Sinauer, Sunderland. [Google Scholar]
- Mach, J. (2015). Domesticated versus wild rice? Bring it awn! The Plant Cell 27, 1818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackay, T. F. C. , Richards, S. , Stone, E. A. , Barbadilla, A. , Ayroles, J. F. , Zhu, D. , Casillas, S. , Han, Y. , Magwire, M. M. , Cridland, J. M. , Richardson, M. F. , Anholt, R. R. H. , Barrón, M. , Bess, C. , Blankenburg, K. P. , et al. (2012). The Drosophila melanogaster genetic reference panel. Nature 482, 173–178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackay, T. F. C. , Stone, E. A. & Ayroles, J. F. (2009). The genetics of quantitative traits: challenges and prospects. Nature Reviews Genetics 10, 565–577. [DOI] [PubMed] [Google Scholar]
- * Maes, J. , Damme, R. V. & Matthysen, E. (2013). Individual and among‐population variation in dispersal‐related traits in natterjack toads. Behavioral Ecology 24, 521–531. [Google Scholar]
- Marden, J. H. , Rogina, B. , Montooth, K. L. & Helfand, S. L. (2003). Conditional tradeoffs between aging and organismal performance of Indy long‐lived mutant flies. Proceedings of the National Academy of Sciences of the United States of America 100, 3369–3373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin, A. E. & Fahrig, L. (2015). Matrix quality and disturbance frequency drive evolution of species behavior at habitat boundaries. Ecology and Evolution 5, 5792–5800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Massol, F. & Debarré, F. (2015). Evolution of dispersal in spatially and temporally variable environments: the importance of life cycles. Evolution 69, 1925–1937. [DOI] [PubMed] [Google Scholar]
- * Massol, F. , Duputié, A. , David, P. & Jarne, P. (2011). Asymmetric patch size distribution leads to disruptive selection on dispersal. Evolution 65, 490–500. [DOI] [PubMed] [Google Scholar]
- Matthysen, E. (2005). Density‐dependent dispersal in birds and mammals. Ecography 28, 403–416. [Google Scholar]
- Matthysen, E. , Van de Casteele, T. & Adriaensen, F. (2005). Do sibling tits (Parus major, P. caeruleus) disperse over similar distances and in similar directions? Oecologia 143, 301–307. [DOI] [PubMed] [Google Scholar]
- Mattila, A. L. K. (2015). Thermal biology of flight in a butterfly: genotype, flight metabolism, and environmental conditions. Ecology and Evolution 5, 5539–5551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mattila, A. L. K. & Hanski, I. (2014). Heritability of flight and resting metabolic rates in the Glanville fritillary butterfly. Journal of Evolutionary Biology 27, 1733–1743. [DOI] [PubMed] [Google Scholar]
- * Mazué, G. P. F. , Dechaume‐Moncharmont, F.‐X. & Godin, J.‐G. J. (2015). Boldness‐exploration behavioral syndrome: interfamily variability and repeatability of personality traits in the young of the convict cichlid (Amatitlania siquia). Behavioral Ecology 26, 900–908. [Google Scholar]
- McCleery, R. H. , Pettifor, R. A. , Armbruster, P. , Meyer, K. , Sheldon, B. C. & Perrins, C. M. (2004). Components of variance underlying fitness in a natural population of the Great Tit Parus major . American Naturalist 164, E62–E72. [DOI] [PubMed] [Google Scholar]
- * McGaugh, S. E. , Schwanz, L. E. , Bowden, R. M. , Gonzalez, J. E. & Janzen, F. J. (2010). Inheritance of nesting behaviour across natural environmental variation in a turtle with temperature‐dependent sex determination. Proceedings of the Royal Society of London B 277, 1219–1226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McManus, M. A. & Woodson, C. B. (2012). Plankton distribution and ocean dispersal. Journal of Experimental Biology 215, 1008–1016. [DOI] [PubMed] [Google Scholar]
- McPeek, M. & Holt, R. D. (1992). The evolution of dispersal in spatially and temporally varying environments. American Naturalist 140, 1010–1027. [Google Scholar]
- Meier, C. M. , Starrfelt, J. & Kokko, H. (2011). Mate limitation causes sexes to coevolve towards more similar dispersal kernels. Oikos 120, 1459–1468. [Google Scholar]
- Merckx, T. & van Dyck, H. (2007). Landscape structure and phenotypic plasticity in flight morphology in the butterfly Pararge aegeria . Oikos 113, 226–232. [Google Scholar]
- * Metz, J. A. & Gyllenberg, M. (2001). How should we define fitness in structured metapopulation models? Including an application to the calculation of evolutionarily stable dispersal strategies. Proceedings of the Royal Society of London B 268, 499–508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer, R. S. & Purugganan, M. D. (2013). Evolution of crop species: genetics of domestication and diversification. Nature Reviews Genetics 14, 840–852. [DOI] [PubMed] [Google Scholar]
- Møller, A. (2002). Parent‐offspring resemblance in degree of sociality in a passerine bird. Behavioral Ecology and Sociobiology 51, 276–281. [Google Scholar]
- Monty, A. & Mahy, G. (2010). Evolution of dispersal traits along an invasion route in the wind‐dispersed Senecio inaequidens (Asteraceae). Oikos 199, 1563–1570. [Google Scholar]
- * Moore, C. R. , Gronwall, D. S. , Miller, N. D. & Spalding, E. P. (2013). Mapping quantitative trait loci affecting Arabidopsis thaliana seed morphology features extracted computationally from images. Genes Genomes Genetics 3, 109–118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgan, M. T. (2002). Genome‐wide deleterious mutation favors dispersal and species integrity. Heredity 89, 253–257. [DOI] [PubMed] [Google Scholar]
- Motro, U. (1982a). Optimal rates of dispersal I. Haploid populations. Theoretical Population Biology 21, 394–411. [Google Scholar]
- Motro, U. (1982b). Optimal rates of dispersal II. Diploid populations. Theoretical Population Biology 21, 412–429. [Google Scholar]
- Motro, U. (1983). Optimal rates of dispersal. III. Parent‐offspring conflict. Theoretical Population Biology 23, 159–168. [Google Scholar]
- Motro, U. (1991). Avoiding inbreeding and sibling competition: the evolution of sexual dimorphism for dispersal. American Naturalist 137, 108–115. [Google Scholar]
- * Mousseau, T. A. & Roff, D. A. (1989). Geographic variability in the incidence and heritability of wing dimorphism in the striped ground cricket, Allonemobius fasciatus . Heredity 62, 315–318. [Google Scholar]
- * Murrell, D. , Travis, J. & Dytham, C. (2002). The evolution of dispersal distance in spatially‐structured populations. Oikos 9, 229–236. [Google Scholar]
- Myles‐Gonzalez, E. , Burness, G. , Yavno, S. , Rooke, A. & Fox, M. G. (2015). To boldly go where no goby has gone before: boldness, dispersal tendency, and metabolism at the invasion front. Behavioral Ecology 26, 1083–1090. [Google Scholar]
- Nachappa, P. , Margolies, D. C. , Nechols, J. R. & Morgan, T. (2010). Response of a complex foraging phenotype to selection on its component traits. Evolutionary Ecology 24, 631–655. [Google Scholar]
- Nanoth Vellichirammal, N. , Zera, A. J. , Schilder, R. J. , Wehrkamp, C. , Riethoven, J.‐J. M. & Brisson, J. A. (2014). De novo transcriptome assembly and morph‐specific gene expression profiling of the wing polymorphic cricket Gryllus firmus . PLoS One 9, e82129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neji, M. , Geuna, F. , Taamalli, W. , Ibrahim, Y. , Smida, M. , Badri, M. , Abdelly, C. & Gandour, M. (2015). Morphophenologic diversity among Tunisian populations of Brachypodium distachyon . Journal of Agricultural Science 153, 1006–1016. [Google Scholar]
- * Nelson, R. M. & Greeff, J. M. (2011). Sex ratio dependent dispersal when sex ratios vary between patches. Journal of Theoretical Biology 290, 81–87. [DOI] [PubMed] [Google Scholar]
- Nielsen, R. & Slatkin, M. (2013). An Introduction to Population Genetics: Theory and Applications. Sinauer Associates, Inc, Sunderland. [Google Scholar]
- Niitepõld, K. (2010). Genotype by temperature interactions in the metabolic rate of the Glanville fritillary butterfly. Journal of Experimental Biology 213, 1042–1048. [DOI] [PubMed] [Google Scholar]
- Niitepõld, K. , Mattila, A. L. K. , Harrison, P. J. & Hanski, I. (2011). Flight metabolic rate has contrasting effects on dispersal in the two sexes of the Glanville fritillary butterfly. Oecologia 165, 847–854. [DOI] [PubMed] [Google Scholar]
- Niitepõld, K. & Saastamoinen, M. (2017). A candidate gene in an ecological model species: phosphoglucose isomerase (Pgi) in the Glanville fritillary butterfly (Melitaea cinxia). Annales Zoologici Fennici 54, 259–273. [Google Scholar]
- Niitepõld, K. , Smith, A. D. , Osborne, J. L. , Reynolds, D. R. , Carreck, N. L. , Martin, A. P. , Marden, J. H. , Ovaskainen, O. & Hanski, I. (2009). Flight metabolic rate and Pgi genotype influence butterfly dispersal rate in the field. Ecology 90, 2223–2232. [DOI] [PubMed] [Google Scholar]
- van Noordwijk, A. , Pulido, F. , Helm, B. , Coppack, T. , Delingat, J. , Dingle, H. , Hedenström, A. , van der Jeugd, H. , Marchetti, C. , Nilsson, A. & Pérez‐Tris, J. (2006). A framework for the study of genetic variation in migratory behaviour. Journal of Ornithology 147, 221–233. [Google Scholar]
- van Noordwijk, A. J. (1984). Problems in the analysis of dispersal and a critique on its ‘heritability’ in the Great tit. Journal of Animal Ecology 53, 533–544. [Google Scholar]
- North, A. , Cornell, S. & Ovaskainen, O. (2011). Evolutionary responses of dispersal distance to landscape structure and habitat loss. Evolution 65, 1739–1751. [DOI] [PubMed] [Google Scholar]
- Öckinger, E. & Van Dyck, H. (2012). Landscape structure shapes habitat finding ability in a butterfly. PLoS One 7, e41517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olivieri, I. & Gouyon, P. H. (1997). Evolution of migration rate and other traits: the metapopulation effect In Metapopulation Biology: Ecology, Genetics, and Evolution (eds Hanski I. and Gilpin M. E.), pp. 293–324. Academic Press, San Diego. [Google Scholar]
- * Olivieri, I. , Michalakis, Y. & Gouyon, P.‐H. (1995). Metapopulation genetics and the evolution of dispersal. American Naturalist 146, 202–228. [Google Scholar]
- Orr, H. A. (2005). The genetic theory of adaptation: a brief history. Nature Reviews Genetics 6, 119–127. [DOI] [PubMed] [Google Scholar]
- Orsini, L. , Wheat, C. W. , Haag, C. R. , Kvist, J. , Frilander, M. J. & Hanski, I. (2009). Fitness differences associated with Pgi SNP genotypes in the Glanville fritillary butterfly (Melitaea cinxia). Journal of Evolutionary Biology 22, 367–375. [DOI] [PubMed] [Google Scholar]
- * Paradis, E. (1998). Interactions between spatial and temporal scales in the evolution of dispersal rate. Evolutionary Ecology 12, 235–244. [Google Scholar]
- * Parker, W. E. & Gatehouse, A. G. (1985). Genetic factors controlling flight performance and migration in the African armyworm moth, Spodoptera exempta (Walker) (Lepidoptera: Noctuidae). Bulletin of Entomological Research 75, 49–63. [Google Scholar]
- * Parvinen, K. , Dieckmann, U. , Gyllenberg, M. & Metz, J. A. J. (2003). Evolution of dispersal in metapopulations with local density dependence and demographic stochasticity. Journal of Evolutionary Biology 1, 143–153. [DOI] [PubMed] [Google Scholar]
- * Parvinen, K. & Metz, J. A. J. (2008). A novel fitness proxy in structured locally finite metapopulations with diploid genetics, with an application to dispersal evolution. Theoretical Population Biology 73, 517–528. [DOI] [PubMed] [Google Scholar]
- Pennekamp, F. , Mitchell, K. A. , Chaine, A. & Schtickzelle, N. (2014). Dispersal propensity in Tetrahymena thermophile ciliates – a reaction norm perspective. Evolution 68, 2319–2330. [DOI] [PubMed] [Google Scholar]
- Penteriani, V. & Delgado, M. M. (2011). Birthplace‐dependent dispersal: are directions of natal dispersal determined a priori? Ecography 34, 729–737. [Google Scholar]
- Pereira, H. S. & Sokolowski, M. B. (1993). Mutations in the larval foraging gene affect adult locomotory behavior after feeding in Drosophila melanogaster . Proceedings of the National Academy of Sciences of the United States of America 90, 5044–5046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perkins, A. , Phillips, B. L. , Baskett, M. L. & Hastings, A. (2013). Evolution of dispersal and life history interact to drive accelerating spread of an invasive species. Ecology Letters 16, 1079–1087. [DOI] [PubMed] [Google Scholar]
- Perrin, N. & Mazalov, V. (1999). Dispersal and inbreeding avoidance. American Naturalist 154, 282–292. [DOI] [PubMed] [Google Scholar]
- * Perrin, N. & Mazalov, V. (2000). Local competition, inbreeding, and the evolution of sex‐biased dispersal. American Naturalist 155, 116–127. [DOI] [PubMed] [Google Scholar]
- van Petegem, H. P. K. , Pétillon, J. , Renault, D. , Wybouw, N. , Van Leeuwen, T. , Stoks, R. & Bonte, D. (2015). Empirically simulated spatial sorting points at fast epigenetic changes in dispersal behaviour. Evolutionary Ecology 29, 299–310. [Google Scholar]
- van Petegem, K. , Boeye, J. , Stoks, R. & Bonte, D. (2016). Spatial selection and local adaptation jointly shape‐history evolution during range expansion. American Naturalist 188, 485–498. [DOI] [PubMed] [Google Scholar]
- Phillips, B. L. , Brown, G. P. & Shine, R. (2010). Life‐history evolution in range‐shifting populations. Ecology 91, 1617–1627. [DOI] [PubMed] [Google Scholar]
- Phillips, B. L. , Brown, G. P. , Travis, J. M. J. & Shine, R. (2008). Reid's paradox revisited: the evolution of dispersal kernels during range expansion. American Naturalist 172, S34–S48. [DOI] [PubMed] [Google Scholar]
- Phillips, B. L. , Brown, G. P. , Webb, J. K. & Shine, R. (2006). Invasion and the evolution of speed in toads. Nature 439, 803. [DOI] [PubMed] [Google Scholar]
- Poethke, H. J. , Dytham, C. & Hovestadt, T. (2011a). A metapopulation paradox: partial improvement of habitat may reduce metapopulations persistence. American Naturalist 177, 792–799. [DOI] [PubMed] [Google Scholar]
- Poethke, H. J. , Gros, A. & Hovestadt, T. (2011b). The ability of individuals to assess population density influences the evolution of emigration propensity and dispersal distance. Journal of Theoretical Biology 282, 93–99. [DOI] [PubMed] [Google Scholar]
- Poethke, H. J. & Hovestadt, T. (2002). Evolution of density‐ and patch‐size‐dependent dispersal rates. Proceedings of the Royal Society of London B 269, 637–645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- * Poethke, H. J. , Hovestadt, T. & Mitesser, O. (2003). Local extinction and the evolution of dispersal rates: causes and correlations. American Naturalist 161, 631–640. [DOI] [PubMed] [Google Scholar]
- Post, D. M. & Palkovacs, E. P. (2009). Eco‐evolutionary feedbacks in community and ecosystem ecology: interactions between the ecological theatre and the evolutionary play. Philosophical Transaction of the Royal Society B: Biological Sciences 364, 1629–1640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- * Potti, J. & Montalvo, S. (1991). Return rate, age at first breeding and natal dispersal of pied flycatchers Ficedula hypoleuca in Central Spain. Ardea 79, 419–428. [Google Scholar]
- Pringle, J. M. , Byers, J. E. , Pappalardo, P. , Wares, J. P. & Marshall, D. (2014). Circulation constrains the evolution of larval development modes and life histories in the coastal ocean. Ecology 95, 1022–1032. [DOI] [PubMed] [Google Scholar]
- Pulido, F. (2007). The genetics and evolution of avian migration. BioScience 57, 165–174. [Google Scholar]
- Pulido, F. , Berthold, P. & van Noordwijk, A. . J. (1996). Frequency of migrants and migratory activity are genetically correlated in a bird population: evolutionary implications. Proceedings of the National Academy of Sciences of the United States of America 93, 14642–14647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raman, H. , Raman, R. , Kilian, A. , Detering, F. , Carling, J. , Coombes, N. , Diffey, S. , Kadkol, G. , Edwards, D. , McCully, M. , Ruperao, P. , Parkin, I. A. P. , Batley, J. , Luckett, D. J. & Wratten, N. (2014). Genome‐wide delineation of natural variation for pod shatter resistance in Brassica napus . PLoS One 9, e101673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- * Ramírez‐Valiente, J. A. & Robledo‐Arnuncio, J. J. (2015). Exotic gene flow affects fitness traits values but not levels of heritable trait variation in the Southernmost population of Scots pine (Pinus sylvestris var. nevadensis). Biological Conservation 192, 331–342. [Google Scholar]
- Ravigné, V. , Olivieri, I. , Gonzalez‐Martinez, S. & Roussett, F. (2006). Selective interactions between short‐distance pollen and seed dispersal in self‐compatible species. Evolution 60, 2257–2271. [PubMed] [Google Scholar]
- Reznick, D. N. & Ghalambor, C. K. (2001). The population ecology of contemporary adaptations: what empirical studies reveal about the conditions that promote adaptive evolution. Genetica 112–113, 183–198. [PubMed] [Google Scholar]
- Riba, M. , Mayol, M. , Giles, B. E. , Ronce, O. , Imbert, E. , van der Velde, M. , Chauvet, S. , Ericson, L. , Bijlsma, R. , Vosman, B. , Smulders, M. J. M. & Olivieri, I. (2009). Darwin's wind hypothesis: does it work for plant dispersal in fragmented habitats? New Phytologist 183, 667–677. [DOI] [PubMed] [Google Scholar]
- Richards, C. L. , Bossdorf, O. & Pigliucci, M. (2010). What role does heritable epigenetic variation play in phenotypic evolution? BioScience 60, 232–237. [Google Scholar]
- Robertson, L. K. , Dey, B. K. , Campos, A. R. & Mahaffey, J. W. (2002). Expression of the Drosophila gene disconnected using the UAS/GAL4 system. Genesis 34, 103–106. [DOI] [PubMed] [Google Scholar]
- Roche, E. A. , Brown, C. R. & Brown, M. B. (2011). Heritable choice of colony size in cliff swallows: does experience trump genetics in older birds? Animal Behaviour 82, 1275–1285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rockman, M. V. (2012). The QTN program and the alleles that matter for evolution: all that's gold does not glitter. Evolution 66, 1–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- * Rodrigues, A. M. M. & Johnstone, R. A. (2014). Evolution of positive and negative density‐dependent dispersal. Proceedings of the Royal Society of London B 281, 20141226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roff, D. A. (1975). Population stability and the evolution of dispersal in a heterogeneous environment. Oecologia 19, 217–237. [DOI] [PubMed] [Google Scholar]
- Roff, D. A. (1994). Habitat persistence and the evolution of wing dimorphism in insects. American Naturalist 144, 772–798. [Google Scholar]
- Roff, D. A. (1995). Antagonistic and reinforcing pleiotrophy: a study of differences in development time in wing dimorphic insects. Journal of Evolutionary Biology 8, 405–419. [Google Scholar]
- Roff, D. A. (2002). Life History Evolution. Sinauer Associates, Inc, Sunderland. [Google Scholar]
- * Roff, D. A. & Bradford, M. J. (1996). Quantitative genetics of the trade‐off between fecundity and wing dimorphism in the cricket allonemobius socius . Heredity 76, 178–185. [Google Scholar]
- Roff, D. A. & Bradford, M. J. (1998). The evolution of shape in the wing dimorphic cricket, Allonemobius socius . Heredity 80, 446–455. [Google Scholar]
- Roff, D. A. & Emerson, K. (2006). Epistasis and dominance: evidence for differential effects in life‐history versus morphological traits. Evolution 60, 1981–1990. [PubMed] [Google Scholar]
- Roff, D. A. & Gélinas, M. B. (2003). Phenotypic plasticity and the evolution of trade‐offs: the quantitative genetics of resource allocation in the wing dimorphic cricket, Gryllus firmus . Journal of Evolutionary Biology 16, 55–63. [DOI] [PubMed] [Google Scholar]
- * Roff, D. A. & Simons, A. S. (1997). The quantitative genetics of wing dimorphism under laboratory and ‘field’ conditions in the cricket Gryllus pennsylvanicus . Heridity 78, 235–241. [Google Scholar]
- * Roff, D. A. , Stirling, G. & Fairbairn, D. J. (1997). The evolution of threshold traits: a quantitative genetic analysis of the physiological and life‐history correlates of wing dimorphism in the sand cricket. Evolution 51, 1910–1919. [DOI] [PubMed] [Google Scholar]
- Rollins, L. A. , Richardson, M. F. & Shine, R. (2015). A genetic perspective on rapid evolution in cane toads (Rhinella marina). Molecular Ecology 24, 2264–2276. [DOI] [PubMed] [Google Scholar]
- Ronce, O. (2007). How does it feel to be like a rolling stone? Ten questions about dispersal evolution. Annual Reviews in Ecology, Ecology and Systematics 38, 231–253. [Google Scholar]
- Ronce, O. & Clobert, J. (2012). Dispersal syndromes In Dispersal Ecology and Evolution (eds Clobert J., Baguette M., Benton T. G. and Bullock J. M.), pp. 119–138. Oxford University Press, Oxford. [Google Scholar]
- * Ronce, O. , Gandon, S. & Rousset, F. (2000). Kin selection and natal dispersal in an age‐structured population. Theoretical Population Biology 159, 143–159. [DOI] [PubMed] [Google Scholar]
- Rousset, F. & Gandon, S. (2002). Evolution of the distribution of dispersal distance under distance‐dependent cost of dispersal. Journal of Evolutionary Biology 15, 515–523. [Google Scholar]
- Roze, D. & Rousset, F. (2005). Inbreeding depression and the evolution of dispersal rates: a multilocus model. American Naturalist 166, 708–721. [DOI] [PubMed] [Google Scholar]
- Roze, D. & Rousset, F. (2009). Strong effects of heterosis on the evolution of dispersal rates. Journal of Evolutionary Biology 22, 1221–1233. [DOI] [PubMed] [Google Scholar]
- Russo, S. E. (2003). Responses of dispersal agents to tree and fruit traits in Virola calophylla (Myristicaceae): implications for selection. Oecologia 136, 80–87. [DOI] [PubMed] [Google Scholar]
- Saastamoinen, M. (2008). Heritability of dispersal and other life history traits in the Glanville fritillary butterfly. Heredity 100, 39–46. [DOI] [PubMed] [Google Scholar]
- Saastamoinen, M. , Ikonen, S. & Hanski, I. (2009). Significant effects of Pgi genotype and body reserves on lifespan in the Glanville fritillary butterfly. Proceedings of the Royal Society of London B 276, 1313–1322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saastamoinen, M. A. K. , Brakefield, P. M. & Ovaskainen, O. (2012). Environmentally induced dispersal‐related life‐history syndrome in the tropical butterfly, Bicyclus anynana . Journal of Evolutionary Biology 25, 2264–2275. [DOI] [PubMed] [Google Scholar]
- Saccheri, I. & Hanski, I. (2006). Natural selection and population dynamics. Trends in Ecology and Evolution 21, 341–347. [DOI] [PubMed] [Google Scholar]
- Santure, A. W. , Poissant, J. , De Cauwer, I. , van Oers, K. , Robinson, M. R. , Quinn, J. L. , Groenen, M. A. M. , Visser, M. E. , Sheldon, B. C. & Slate, J. (2015). Replicated analysis of the genetic architecture of quantitative traits in two wild great tit populations. Molecular Ecology 24, 6148–6162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- * Saunders, S. P. , Arnold, T. W. , Roche, E. A. & Cuthbert, F. J. (2014). Age‐specific survival and recruitment of piping plovers Charadrius melodus in the Great Lakes region. Journal of Avian Biology 45, 437–449. [Google Scholar]
- * Scheiner, S. M. , Barfield, M. & Holt, R. D. (2012). The genetics of phenotypic plasticity. XI. Joint evolution of plasticity and dispersal rate. Ecology and Evolution 2, 2027–2039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schielzeth, H. & Husby, A. (2014). Challenges and prospects in genome‐wide quantitative trait loci mapping of standing genetic variation in natural populations. Annals of the New York Academy of Sciences 1320, 35–57. [DOI] [PubMed] [Google Scholar]
- Schtickzelle, N. , Joiris, A. , van Dyck, H. & Baguette, M. (2007). Quantitative analysis of changes in movement behaviour within and outside habitat in a specialist butterfly. BMC Evolutionary Biology 7, 4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schtickzelle, N. , Mennechez, G. & Baguette, M. (2006). Dispersal depression with habitat fragmentation in the Bog fritillary butterfly. Ecology 87, 1057–1065. [DOI] [PubMed] [Google Scholar]
- * Scumacher, P. , Weyeneth, A. , Weber, D. & Dorn, S. (1997). Long flights in Cydia pomonella L. (Lepidoptera: Tortricidae) measured by a flight mill: influence of sex, mated status and age. Physiological Entomology 22, 149–160. [Google Scholar]
- Selonen, V. & Hanski, I. K. (2004). Young flying squirrels (Pteromys volans) dispersing in fragmented forests. Behavioral Ecology 15, 564–571. [Google Scholar]
- Serrano, D. & Tella, J. L. (2007). The role of despotism and heritability in determining settlement patterns in the colonial Lesser kestrel. American Naturalist 169, E53–E67. [DOI] [PubMed] [Google Scholar]
- Shaw, A. K. & Kokko, H. (2014). Mate finding, Allee effects and selection for sex‐biased dispersal. Journal of Animal Ecology 83, 1256–1267. [DOI] [PubMed] [Google Scholar]
- Shaw, A. K. & Kokko, H. (2015). Dispersal evolution in the presence of Allee effects can speed up or slow down invasions. American Naturalist 185, 631–639. [DOI] [PubMed] [Google Scholar]
- Simmons, A. D. & Thomas, C. D. (2004). Changes in dispersal during species' range expansions. American Naturalist 164, 378–395. [DOI] [PubMed] [Google Scholar]
- Sinervo, B. , Calsbeek, R. , Comendant, T. , Both, C. , Adamopoulou, C. & Clobert, J. (2006). Genetic and maternal determinants of effective dispersal: the effect of sire genotype and size at birth in side‐blotched lizards. American Naturalist 168, 88–99. [DOI] [PubMed] [Google Scholar]
- Slate, J. (2005). Quantitative trait locus mapping in natural populations: progress, caveats and future directions. Molecular Ecology 14, 363–379. [DOI] [PubMed] [Google Scholar]
- Soons, M. B. , Heil, G. W. , Nathan, R. & Katul, G. G. (2004). Determinants of long‐distance seed dispersal by wind in grasslands. Ecology 85, 3056–3068. [Google Scholar]
- * Sparkman, A. M. , Adams, J. R. , Steury, T. D. , Waits, L. P. & Murray, D. L. (2012). Evidence for a genetic basis for delayed dispersal in a cooperatively breeding canid. Animal Behaviour 83, 1091–1098. [Google Scholar]
- Stamps, J. , Krishnan, V. V. & Reid, M. L. (2005). Search costs and habitat selection by dispersers. Ecology 86, 510–518. [Google Scholar]
- * Starrfelt, J. & Kokko, H. (2010). Parent‐offspring conflict and the evolution of dispersal distance. American Naturalist 175, 38–49. [DOI] [PubMed] [Google Scholar]
- Stevens, V. , Whitmee, S. , Le Galliard, J.‐F. , Clobert, J. , Bohning‐Gaese, K. , Bonte, D. , Brandle, M. , Dehling, M. , Hof, C. , Trochet, A. & Baguette, M. (2014). A comparative analysis of dispersal syndromes in terrestrial and semi‐terrestrial animals: perspectives for the evolution in a changing world. Ecology Letters 17, 1039–1052. [DOI] [PubMed] [Google Scholar]
- Stevens, V. M. , Trochet, A. , Blanchet, S. , Moulherat, S. , Clobert, J. & Baguette, M. (2013). Dispersal syndromes and the use of life‐histories to predict dispersal. Evolutionary Applications 6, 630–642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stevens, V. M. , Trochet, A. , Van Dyck, H. , Clobert, J. & Baguette, M. (2012). How is dispersal integrated in life histories: a quantitative analysis using butterflies. Ecology Letters 15, 74–86. [DOI] [PubMed] [Google Scholar]
- Sun, S. & Cheptou, P. (2012). Life‐history traits evolution across distribution ranges: how the joint evolution of dispersal and mating system favor the evolutionary stability of range limits? Evolutionary Ecology 26, 771–778. [Google Scholar]
- Taylor, P. (1988). An inclusive fitness model for dispersal of offspring. Journal of Theoretical Biology 130, 363–378. [Google Scholar]
- Taylor, T. B. & Buckling, A. (2011). Selection experiments reveal trade‐offs between swimming and twitching motilities in Pseudomonas aeruginosa . Evolution 65, 3060–3069. [DOI] [PubMed] [Google Scholar]
- * Teyssier, A. , Bestion, E. , Richard, M. & Cote, J. (2014). Partners' personality types and mate preferences: predation risk matters. Behavioral Ecology 25, 723–733. [Google Scholar]
- Therry, L. , Bonte, D. & Stoks, R. (2015). Higher investment in flight morphology does not trade off with fecundity estimates in a poleward range‐expanding damselfly. Ecological Entomology 40, 133–142. [Google Scholar]
- Thomas, C. D. , Bodsworth, E. J. , Wilson, R. J. , Simmons, A. D. , Davies, Z. G. , Musche, M. & Conradt, L. (2001). Ecological and evolutionary processes at expanding range margins. Nature 411, 577–581. [DOI] [PubMed] [Google Scholar]
- Thomas, C. D. , Hill, J. K. & Lewis, O. T. (1998). Evolutionary consequences of habitat fragmentation in a localized butterfly. Journal of Animal Ecology 67, 485–497. [Google Scholar]
- Thomson, F. J. , Moles, A. T. , Auld, T. D. , Ramp, D. , Ren, S. & Kingsford, R. T. (2010). Chasing the unknown: predicting seed dispersal mechanisms from plant traits. Journal of Ecology 98, 1310–1318. [Google Scholar]
- * Thurber, C. S. , Jia, M. H. , Jia, Y. & Caicedo, A. L. (2013). Similar traits, different genes? Examining convergent evolution in related weedy rice populations. Molecular Ecology 22, 685–698. [DOI] [PubMed] [Google Scholar]
- Tien, N. S. H. , Sabelis, M. W. & Egas, M. (2011). Ambulatory dispersal in Tetranychus urticae: an artificial selection experiment on propensity to disperse yields no response. Experimental and Applied Acarology 53, 349–360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- * Toonen, R. J. & Pawlik, J. R. (2001). Foundations of gregariousness: a dispersal polymorphism among the planktonic larvae of a marine invertebrate. Evolution 55, 2439–2454. [DOI] [PubMed] [Google Scholar]
- Travis, J. , Reznick, D. , Bassar, R. D. , Lopez‐Sepulcre, A. , Ferriere, R. & Coulson, T. (2014). Do Eco‐Evo feedbacks help us understand nature? Answers from studies of the Trinidadian guppy. Advances in Ecological Research 50, 1–40. [Google Scholar]
- * Travis, J. M. J. (2001). The color of noise and the evolution of dispersal. Ecological Research 16, 157–163. [Google Scholar]
- Travis, J. M. J. , Delgado, M. , Bocedi, G. , Baguette, M. , Barton, K. , Bonte, D. , Boulengeat, I. , Hodgson, J. A. , Kubisch, A. , Penteriani, V. , Saastamoinen, M. , Stevens, V. M. & Bullock, J. M. (2013a). Dispersal and species response to climate change. Oikos 122, 1532–1540. [Google Scholar]
- Travis, J. M. J. & Dytham, C. (1999). Habitat persistence, habitat availability and the evolution of dispersal. Proceedings of the Royal Society of London B 266, 723–728. [Google Scholar]
- Travis, J. M. J. & Dytham, C. (2002). Dispersal evolution during invasions. Evolutionary Ecological Research 4, 1119–1129. [Google Scholar]
- * Travis, J. M. J. , Münkemüller, T. & Burton, O. J. (2010a). Mutation surfing and the evolution of dispersal during range expansions. Journal of Evolutionary Biology 23, 2656–2667. [DOI] [PubMed] [Google Scholar]
- Travis, J. M. J. , Murrell, D. J. & Dytham, C. (1999). The evolution of density‐dependent dispersal. Proceedings of the Royal Society of London B 266, 1837–1842. [Google Scholar]
- Travis, J. M. J. , Mustin, K. , Bartoń, K. A. , Benton, T. G. , Clobert, J. , Delgado, M. M. , Dytham, C. , Hovestadt, T. , Palmer, S. C. F. , Van Dyck, D. & Bonte, D. (2012). Modelling dispersal: an eco‐evolutionary framework incorporating emigration, movement, settlement behaviour and the multiple costs involved. Methods in Ecology and Evolution 3, 628–641. [Google Scholar]
- Travis, J. M. J. , Mustin, K. , Benton, T. G. & Dytham, C. (2009). Accelerating invasion rates result from the evolution of density‐dependent dispersal. Journal of Theoretical Biology 259, 151–158. [DOI] [PubMed] [Google Scholar]
- * Travis, J. M. J. , Palmer, S. C. F. , Coyne, S. , Millon, A. & Lambin, X. (2013b). Evolution of predator dispersal in relation to spatio‐temporal prey dynamics: how not to get stuck in the wrong place! PLoS One 8, e54453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Travis, J. M. J. , Smith, H. S. & Ranwala, S. M. W. (2010b). Towards a mechanistic understanding of dispersal evolution in plants: conservation implications. Diversity and Distributions 16, 690–702. [Google Scholar]
- Trochet, A. , Legrand, D. , Larranaga, N. , Ducatez, S. , Calvez, O. , Cote, J. , Clobert, J. & Baguette, M. (2013). Population sex ratio and dispersal in experimental, two‐patch metapopulations of butterflies. Journal of Animal Ecology 82, 946–955. [DOI] [PubMed] [Google Scholar]
- Tschirren, B. , Fitze, P. S. & Richner, H. (2007). Maternal modulation of natal dispersal in a passerine bird: an adaptive strategy to cope with parasitism? American Naturalist 169, 87–93. [DOI] [PubMed] [Google Scholar]
- Turlure, C. , Schtickzelle, N. & Baguette, M. (2010). Resource grain scales mobility and adult morphology in butterflies. Landscape Ecology 25, 95–108. [Google Scholar]
- Urban, M. C. , Bocedi, G. , Hendry, A. P. , Mihoub, J.‐B. , Pe'er, G. , Singer, A. , Bridle, J. R. , Crozier, L. G. , De Meester, L. , Godsoe, W. , Gonzalez, A. , Hellmann, J. J. , Holt, R. D. , Huth, A. , Johst, K. , Krug, C. B. , et al. (2016). Improving the forecast for biodiversity under climate change. Science 353, aad8466. [DOI] [PubMed] [Google Scholar]
- van Valen, L. (1971). Group selection and the evolution of dispersal. Evolution 25, 591–598. [DOI] [PubMed] [Google Scholar]
- Van Dyck, H. & Baguette, M. (2005). Dispersal behaviour in fragmented landscapes: routine or special movements? Basic and Applied Ecology 6, 535–545. [Google Scholar]
- Van Dyck, H. , Matthysen, E. & Wiklund, C. (1998). Phenotypic variation in adult morphology and pupal colour within and among families of the speckled wood butterfly Pararge aegeria . Ecological Entomology 23, 465–472. [Google Scholar]
- Venable, D. L. (1985). Ecology of achene dimorphism in Heterotheca latifolia. 3. Consequences of varied water availability. Journal of Ecology 73, 757–763. [Google Scholar]
- Venable, D. L. & Burquez, A. (1989). Quantitative genetics of size, shape, life‐history, and fruit characteristics of the seed heteromorphic composite Heterosperma pinnatum. 1. Variation within and among populations. Evolutionary Ecology 43, 113–124. [DOI] [PubMed] [Google Scholar]
- * Venable, D. L. , Dyreson, E. & Morales, E. (1995). Population dynamic consequences and evolution of seed traits of Heterosperma pinnatum (Asteraceae). American Journal of Botany 82, 410–420. [Google Scholar]
- Violle, C. , Navas, M.‐L. , Vile, D. , Kazakou, E. , Fortunel, C. , Hummel, I. & Garnier, E. (2007). Let the concept of trait be functional! Oikos 116, 882–892. [Google Scholar]
- Vitalis, R. , Rousset, F. , Kobayashi, Y. , Olivieri, I. & Gandon, S. (2013). The joint evolution of dispersal and dormancy in a metapopulation with local extinctions and kin competition. Evolution 67, 1676–1691. [DOI] [PubMed] [Google Scholar]
- Walsh, B. & Blows, M. W. (2009). Abundant genetic variation + strong selection = multivariate genetic constraints: a geometric view of adaptation. Annuals Reviews in Ecology, Evolution and Systematics 40, 41–59. [Google Scholar]
- * Waser, P. M. & Jones, W. T. (1989). Heritability of dispersal in banner‐tailed kangaroo rats. Animal Behaviour 37, 987–991. [Google Scholar]
- * Watkins, T. B. & McPeek, M. A. (2006). Growth and predation risk in green frog tadpoles (Rana clamitans): a quantitative genetic analysis. Copeia 2006, 478–488. [Google Scholar]
- Weeden, N. F. (2007). Genetic changes accompanying the domestication of Pisum sativum: is there a common genetic basis to the ‘domestication syndrome’ for legumes? Annals of Botany 100, 1017–1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wender, N. J. , Polisetty, C. & Donohue, K. (2005). Density‐dependent processes influencing the evolutionary dynamics of dispersal: a functional analysis of seed dispersal in Arabidopsis thaliana (Brassicaceae). American Journal of Botany 92, 88–98. [DOI] [PubMed] [Google Scholar]
- Wheat, C. W. (2012). Dispersal genetics: emerging insights from fruitflies, butterflies, and beyond In Dispersal Ecology and Evolution (eds Clobert J., Baguette M., Benton T. G. and Bullock J. M.), pp. 95–107. Oxford University Press, Oxford. [Google Scholar]
- Wheat, C. W. , Fescemyer, H. W. , Kvist, J. , Tas, E. , Vera, J. C. , Frilander, M. J. , Hanski, I. & Marden, J. H. (2011). Functional genomics of life history variation in a butterfly metapopulation. Molecular Ecology 20, 1813–1828. [DOI] [PubMed] [Google Scholar]
- Wheat, C. W. , Haag, C. R. , Marden, J. H. , Hanski, I. & Frilander, M. J. (2010). Nucleotide polymorphism at a gene (Pgi) under balancing selection in a butterfly metapopulation. Molecular Biology and Evolution 27, 267–281. [DOI] [PubMed] [Google Scholar]
- Wheat, C. W. & Hill, J. (2014). Pgi: the ongoing saga of a candidate gene. Current Opinion in Insect Science 4, 42–47. [DOI] [PubMed] [Google Scholar]
- Wheelwright, N. T. (1993). Fruit size in a tropical tree species: variation, preference by birds, and heritability. Vegetatio 107, 163–174. [Google Scholar]
- Wheelwright, N. T. , Lawler, J. J. & Weinstein, J. H. (1997). Nest‐site selection in Savannah sparrows: using gulls as scarecrows? Animal Behaviour 53, 197–208. [Google Scholar]
- * Wheelwright, N. T. & Mauck, R. A. (1998). Philopatry, natal dispersal, and inbreeding avoidance in an island population of Savannah sparrows. Ecology 79, 755–767. [Google Scholar]
- Williams, J. L. , Kendall, B. E. & Levine, J. M. (2016). Rapid evolution accelerates plant population spread in fragmented experimental landscapes. Science 353, 482–485. [DOI] [PubMed] [Google Scholar]
- Wilson, A. , Reale, D. , Clements, M. N. , Morrisey, M. M. , Postma, E. , Walling, C. A. , Kruuk, L. E. B. & Nussey, D. H. (2010). An ecologits's guide to animal model. Journal of Animal Ecology 79, 13–26. [DOI] [PubMed] [Google Scholar]
- Wolak, M. E. & Keller, L. F. (2014). Dominance genetic variance and inbreeding in natural populations In Quantitative Genetics in the Wild (eds Charmentier A., Garant D. and Kruuk L. E. B.), pp. 104–127. Oxford University Press, Oxford. [Google Scholar]
- Yano, S. & Takafuji, A. (2002). Variation in the life history pattern of Tetranychus urticae (Acari: Tetranychidae) after selection for dispersal. Experimental and Applied Acarology 27, 1–10. [DOI] [PubMed] [Google Scholar]
- Yeaman, S. (2015). Local adaptation by alleles of small effect. American Naturalist 186, S74–S89. [DOI] [PubMed] [Google Scholar]
- Yeaman, S. & Guillaume, F. (2009). Predicting adaptation under migration load: the role of genetic skew. Evolution 63, 2926–2938. [DOI] [PubMed] [Google Scholar]
- Yeaman, S. & Whitlock, M. C. (2011). The genetic architecture of adaptation under migration‐selection balance. Evolution 65, 1897–1911. [DOI] [PubMed] [Google Scholar]
- Yukilevich, R. (2005). Dispersal evolution in fragmented habitats: the interplay between the tendency and the ability to disperse. Evolutionary Ecological Research 7, 973–992. [Google Scholar]
- Závorka, L. , Aldvén, D. , Näslund, J. , Höjesjö, J. & Johnsson, J. I. (2015). Linking lab activity with growth and movement in the wild: explaining pace‐of‐life in a trout stream. Behavioral Ecology 26, 877–884. [Google Scholar]
- Zera, A. J. (2006). Evolutionary genetics of juvenile hormone and ecdysteroid regulation in Gryllus: a case study in the microevolution of endocrine regulation. Comparative Biochemistry and Physiology A‐Molecular & Integrative Physiology 144, 365–379. [DOI] [PubMed] [Google Scholar]
- Zera, A. J. (2016). Juvenile hormone and the regulation of wing polymorphism in insects: new insights from circadian and functional‐genomic studies in Gryllus crickets. Physiological Entomology 41, 313–326. [Google Scholar]
- Zera, A. J. & Brink, T. (2000). Nutrient absorption and utilization by wing and flight muscle morphs of the cricket Gryllus firmus: implications for the trade‐off between flight capability and early reproduction. Journal of Insect Physiology 46, 1207–1218. [DOI] [PubMed] [Google Scholar]
- Zera, A. J. & Brisson, J. A. (2012). Quantitative, physiological, and molecular genetics of dispersal/migration In Dispersal Ecology and Evolution (eds Clobert J., Baguette M., Benton T. G. and Bullock J. M.), pp. 63–82. Oxford University Press, Oxford. [Google Scholar]
- Zera, A. J. & Denno, R. F. (1997). Physiology and ecology of dispersal polymorphism in insects. Annual Review of Entomology 42, 207–230. [DOI] [PubMed] [Google Scholar]
- Zera, A. J. & Harshman, L. G. (2001). The physiology of life history trade‐offs in animals. Annual Review of Ecology and Systematics 32, 95–126. [Google Scholar]
- Zera, A. J. & Larsen, A. (2001). The metabolic basis of life history variation: genetic and phenotypic differences in lipid reserves among life history morphs of the wing‐polymorphic cricket, Gryllus firmus . Journal of Insect Physiology 47, 1147–1160. [DOI] [PubMed] [Google Scholar]
- Zera, A. J. & Zhao, Z. (2003). Life‐history evolution and the microevolution of intermediary metabolism: activities of lipid‐metabolizing enzymes in life‐history morphs of a wing‐dimorphic cricket. Evolution 57, 586–596. [DOI] [PubMed] [Google Scholar]
- Zhang, R. , Jongejans, E. & Shea, K. (2011). Warming increases the spread of an invasive thistle. PLoS One 6, e21725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- * Zheng, C. , Ovaskainen, O. & Hanski, I. (2009). Modelling single nucleotide effects in phosphoglucose isomerase on dispersal in the Glanville fritillary butterfly: coupling of ecological and evolutionary dynamics. Philosophical Transaction of the Royal Society B: Biological Sciences 364, 1519–1532. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Appendix S1. Methods.
Table S1. Examples of heritability (h 2) estimates in the reviewed studies.
Table S2. Models of dispersal evolution and assumptions made on the genetic architecture of the evolving traits.