

Key Points
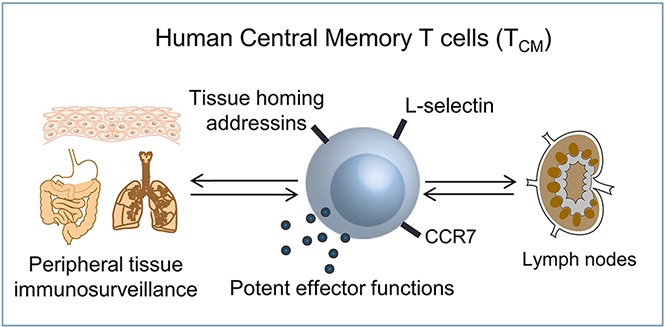
Human TCM are tissue tropic, have impressive effector functions, and are found in noninflamed human tissues.
TCM can act alone to induce inflammation in human skin–grafted mice; results suggest a role for human TCM in primary immunosurveillance.
Abstract
Central memory T cells (TCM) patrol lymph nodes, providing central immunosurveillance against known pathogens, but have not been described as conducting primary tissue immunosurveillance. We analyzed the expression of tissue-homing addressins in human TCM vs effector memory T cells (TEM) from the same donors. In humans, the majority of human TCM were tropic for either skin or gut, and the overall tissue tropism of TCM was comparable to that of TEM. TCM were present in healthy, noninflamed human skin, lung, colon, and cervix, suggesting a role for TCM in the primary immunosurveillance of peripheral tissues. TCM also had potent effector functions; 80% of CD8+ TCM produced TC1/TC2/TC17/TC22 cytokines. TCM injected into human skin–grafted mice migrated into skin and induced inflammatory eruptions comparable to TEM-injected mice. In summary, human TCM express peripheral tissue-homing receptors at levels similar to their effector memory counterparts, are found in healthy human tissues, have impressive effector functions, and can act alone to induce skin inflammation in human engrafted mice. Our studies support a novel role for human TCM in primary immunosurveillance of peripheral tissues and highlight the important role of this long-lived cell type in tissue-based immune responses.
Visual Abstract
Introduction
Central memory T cells (TCM) coexpress L-selectin and CCR7 and provide central immunosurveillance by patrolling the lymph nodes draining peripheral tissue sites.1,2 In mice and nonhuman primates, TCM provide effective long-term protection because they persist long term in the circulation, have a high proliferative potential, and can give rise to both effector and effector memory T cells (TEM) after antigen reencounter.3-6 Although they can be drawn into inflamed tissues, TCM have not been identified in animals as providing primary tissue-based immunosurveillance.7 We report that human TCM express tissue-homing receptors, are found in noninflamed human tissues, and have potent effector functions, supporting a role for these cells in primary tissue immunosurveillance.
Methods
Samples
All studies were performed in accordance with the Declaration of Helsinki. Approval of the Partners Institutional Review Board committee was obtained for all studies. Deidentified blood, skin, foreskin, lung, and colon were obtained from Brigham and Women’s Hospital, and cervix was obtained from Johns Hopkins. Lung and colon were obtained from distant, uninvolved tissue obtained from patients undergoing resection of small isolated tumors. Normal cervix was obtained from patients undergoing hysterectomy for nonmalignant disorders. T cells were isolated by collagenase digestion or short-term explant culture.8,9
Flow cytometry and CyTOF
A list of antibodies used to immunostain cells is included in the supplemental Methods. Flow cytometry samples were run on a Becton Dickinson FACSCanto instrument, and CyTOF samples were analyzed on a Fluidigm CyTOF 2 mass cytometer. Data were analyzed with FCS Express 5.0 or FACSDiva 8.0. Cells were sorted on a FACSAria cell sorter.
Human engrafted mouse model
Human engrafted mice were prepared as described previously.9 Briefly, neonatal foreskins were grafted onto the backs of 6- to 8-week-old nonobese diabetic/severe combined immunodeficiency/interleukin-2 (IL-2) receptor γ chainnull mice (Jackson Laboratories). One week later, 3 × 106 flow-sorted TCM, TEM, or naive T cells from an unrelated adult human blood donor were injected IV. The gating strategy and flow cytometry profiles of infused cells are included in supplemental Figure 1. Skin grafts were harvested after 3 weeks for analysis.
Statistical analyses
Primary methods of data analysis included descriptive statistics (means, medians, and standard deviation). Differences between 2 sample groups were detected using the 2-tailed Wilcoxon-Mann-Whitney test, α = 0.05. For comparisons of multiple groups, a Kruskal-Wallis 1-way analysis of variance with a Bonferroni-Dunn’s posttest for multiple means test was used, α = 0.05.
Results
We compared the expression of tissue-homing addressins in TCM vs TEM from healthy human blood donors by CyTOF (Figure 1). TEM are generated in response to tissue-based infections and should all be tropic for peripheral tissues.10,11 Skin (cutaneous lymphocyte antigen, CLA)12 and gut (α4β7 integrin)13,14 homing receptors have been identified, but receptors that direct T cells to lung, brain, and other peripheral tissues remain uncharacterized. We therefore measured the expression of skin and gut-homing addressins in TCM vs TEM from healthy human donors. Subsets of TCM and TEM expressed either gut-homing or skin-homing addressins, with very few cells expressing both (Figure 1A). Both CD8+ and CD4+ TCM contained skin-tropic and gut-tropic populations (Figure 1A-B,G). The expression of skin and gut-homing receptors was comparable in TCM and TEM, except that significantly more CD4+ TEM expressed gut-homing receptors and significantly more CD8+ TCM expressed skin-homing receptors (Figure 1C-D). When the total number of T cells expressing skin or gut-homing receptors were compared, TCM and TEM expressed tissue-homing addressins at similar levels (Figure 1E-F).
Figure 1.

Human TCMexpress tissue-homing addressins and are found in healthy human peripheral tissues. (A-B) Comparison of the expression of skin-homing (CLA) and gut-homing (α4β7) addressins in TCM and TEM by CyTOF analyses from a representative human donor. The gating strategy used to identify TCM and TEM among human peripheral blood CD45RO+ CD45RA− FOXP3− memory T cells is shown. (C-D) Aggregate data of skin and gut addressin expression by (C) CD4+ and (D) CD8+ TCM (white bars) and TEM (black bars). T cells that expressed neither CLA nor α4β7 were designated as unassigned. The mean and standard error of the mean (SEM) of 4 donors are shown. (E-G) TCM and TEM have comparable expression of skin- and gut-homing addressins. The mean and SEM of the total assignable tissue-tropic populations (total tropism, the sum of skin and gut-tropic populations) for (E) CD4+ and (F) CD8+ T cells are shown. The mean and SEM of 6 donors are shown. (H) TCM are present in noninflamed human peripheral tissues. T cells were isolated from noninflamed human tissues and analyzed by flow cytometry. TCM as a percentage of the total T-cell population are shown for (H) individual representative samples and (I) pooled data. The mean and SEM of 5 skin, 2 lung, 3 colon, and 6 cervix samples are shown.
These results suggest human TCM have intrinsic tissue tropism and may play a role in primary immunosurveillance of peripheral tissues. To evaluate this possibility, we isolated T cells from noninflamed human skin, lung, colon, and cervix. We observed populations of TCM in each of these healthy peripheral tissues, confirming that TCM do gain access to noninflamed human tissues (Figure 1H-I).
TCM are known for their ability to persist in the circulation, to proliferate, and to give rise to effector T cells in animal models, but their production of cytokines that can directly combat infection has not been characterized in humans. We studied cytokine production of TCM and TEM and found that both CD4 and CD8 TCM had impressive effector functions (Figure 2A-J). Interferon-γ (IFN-γ) was the most highly produced signature cytokine in TCM, as it was in TEM. CD8+ TCM were particularly cytokine rich, with a mean 70% and 97% of CD8+ TCM producing signature cytokines among the skin-tropic and gut-tropic populations, respectively. The proportion of CD8+ TCM producing signature cytokines was not significantly different from TEM from the same donors (Figure 2G). Gut-tropic CD8+ TCM produced significantly more IL-2 than CD8+ TEM, but levels produced by all other subsets were comparable (Figure 2C,H). Between 40% and 55% of CD4+ TCM also produced signature cytokines, and IFN-γ was the most frequently produced cytokine in both CD4+ and CD8+ TCM (Figure 2A-B,F). Tumor necrosis factor-α (TNF-α) was produced by the majority of T cells tested, and IL-10 was produced at low levels in all subsets (Figure 2D-E,I-J). These results demonstrate that all TCM, and CD8+ TCM in particular, have considerable effector functions that endow them with the capacity to contribute to frontline antipathogen responses in peripheral tissues.
Figure 2.
Human TCMhave potent effector functions, home to skin, and induce dermatitis in human engrafted mice. (A,F) Both CD4+ and CD8+ TCM produced T-cell signature inflammatory cytokines. Cytokine production was assayed by CyTOF following stimulation with phorbol 12-myristate 13-acetate/ionomycin. Production of TH1/TC1 (IFN-γ), TH2/TC2 (IL-4, IL-13), TH17/TC17 (IL-17A), TH22/TC22 (IL-22) signature cytokines by TCM and TEM is shown. Figures represent the mean of 6 donors. (B,G) Total production of signature cytokines by TCM (white bars) compared with TEM from the same donors (black bars) for (B) CD4+ and (G) CD8+ T cells. (C-E,H-J) Production of IL-2, TNF-α, and IL-10 by (C-E) CD4+ and (H-J) CD8+ T cells is shown. The mean and SEM of 6 donors are shown. (K-W) Human TCM home to skin and induce an inflammatory dermatitis comparable to TEM in human engrafted mice. (K) The human engrafted mouse experimental model. (L-P) Hematoxylin and eosin evaluation of human skin grafts 3 weeks after injection of (L-M) saline, (N) TEM, (O) TCM, or (P) naive T cells. TCM homed to human skin grafts and induced T-cell–mediated inflammatory dermatitis. (P) Injection of naive T cells led to minimal inflammation. (Q-S) The inflammatory patterns induced by purified TCM included (Q) interface dermatitis, (R) spongiotic dermatitis, and (S) epidermal necrosis. Results shown are representative of those obtained with 6 different human blood cell donors. (T) T-cell migration into the skin as assessed by NanoString CD3/CD4/CD8 gene expression analysis was comparable in TCM- and TEM-injected mice. (U-V) The production of inflammatory T-cell cytokines in skin (TNF-α, IFN-γ, IL-17A, and IL-22) and cytotoxic effector molecules (PRF1, perforin; GZMA, granzyme A; GZMB, granzyme B) was comparable in TCM- and TEM-injected mice. (W) Production of inflammatory chemokines in skin was comparable in TCM- and TEM-injected mice. For panels T-W, the mean and SEM of messenger RNA copies detected by NanoString analyses from 6 different human TCM/TEM donors are shown. Scale bars, 100 μm. GVHD, graft-versus-host disease; NS, normal skin; ns, not significant.


We used a human engrafted mouse model to measure the relative ability of TCM and TEM to enter skin and induce inflammation. NSG mice were grafted with neonatal human foreskin, a tissue that lacks T cells, and infused IV with allogeneic purified peripheral blood TEM, TCM, or naive T cells isolated from healthy adult human donors (Figure 2K; supplemental Figure 1).9 A robust inflammatory dermatitis was observed in both TEM- and TCM-injected mice (Figure 2L-S). TCM effectively entered human skin and induced inflammatory changes, including interface dermatitis, epidermal spongiosis, and epidermal necrosis (Figure 2Q-S). One caveat is that TCM may differentiate into effector cells within the skin graft, and we cannot rule out a contribution of newly generated effector cells to skin inflammation. In contrast, injection of naive T cells led to minimal infiltration of T cells into skin and little if any visible inflammation (Figure 2P). Very few human T cells migrated into mouse skin adjacent to the grafts, and no inflammation in mouse skin was appreciated (supplemental Figure 2). Although this is not a model of immunosurveillance per se, it does measure in vivo the ability of human T cells to enter human skin and induce inflammation.
NanoString-based gene expression profiling of the skin grafts demonstrated comparable levels of T cells in the skin of TCM- and TEM-injected mice (Figure 2T). Moreover, the levels of TNF-α, IFN-γ, IL-17A, IL-22, perforin, granzyme A, granzyme B, and inflammatory chemokines were similar in the skin of TCM- and TEM-injected mice (Figure 2U-W). These studies demonstrate and confirm the ability of TCM to enter the skin and initiate inflammation in the absence of other T-cell subsets.
Discussion
TCM provide effective long-term memory responses because they have the capacity to persist long term in the circulation, have a high proliferative capacity, and can replenish other memory T-cell subsets, including TEM.3-6,15 The role of TCM in immunosurveillance has been assumed to be limited to patrolling the lymph nodes for evidence of pathogen exposure. The initial description of human TCM characterized these cells as having poor effector functions and little tissue tropism.1 However, these studies did not evaluate the expression of the gut-homing addressin α4β7, used a different antibody to detect CLA than the one that identifies cutaneous T cells (HECA-205 vs HECA-45212), and did not study the cytokine production of polyclonally stimulated TCM. With the benefit of updated and more comprehensive approaches, it is clear that TCM express tissue-homing addressins at levels similar to TEM, and indeed, these cells are present in healthy human peripheral tissues. This is consistent with a prior report that CCR7 is expressed by the majority of CLA- and α4β7-expressing T cells in human blood.16 Human TCM, particularly CD8+ TCM, also have significant effector functions. TCM alone were capable of entering human skin and initiating inflammation comparable to that induced by TEM.
These findings demonstrate that TCM express tissue-homing receptors, are found in healthy human peripheral tissues, have potent effector functions, and can migrate into and initiate tissue-based inflammation. Our findings suggest that human TCM, much like TEM, are imprinted with both tissue-homing addressin expression and specialized programs of cytokine production and likely participate directly in the immunosurveillance of peripheral tissues.
Tissue-tropic TCM have not yet been described in animal models, perhaps because young mice kept in pathogen-free conditions, the animals used in most experiments, lack the large numbers of pathogen-specific recirculating TCM that human patients have accumulated over decades of pathogen exposures. Alternatively, there may be key differences in homing of human vs mouse TCM.
Our work demonstrates that human TCM enter and have the capacity to provide primary immunosurveillance of peripheral tissues. Added to their known abilities to persist long term in the circulation, proliferate, and give rise to additional memory T-cell subsets, our work supports a critical role for TCM in providing long-term protection against known pathogens.
Supplementary Material
The full-text version of this article contains a data supplement.
Acknowledgments
Michael Yaremchuk, Jeffrey Darrow, George Volpe, Dax Guenther, and Raffi Der Sarkissian of the Boston Center for Plastic Surgery and Bohdan Pomahac and Simon Talbot of Brigham and Women’s Hospital generously provided adult human skin samples. CyTOF samples were acquired with the assistance of Nicole Paul of the Dana-Farber Cancer Institute. Eugene Butcher kindly provided the ACT-1 antibody.
This work was supported by National Institutes of Health (NIH), National Institute of Arthritis and Musculoskeletal and Skin Diseases grants R01 AR063962 (R.A.C.), R01 AR056720 (R.A.C.), P30 AR069625 (R.A.C.), T32 AR-07098-36 (T.S.K.; supplied salary for J.T.O.), NIH, National Institute of Allergy and Infectious Diseases grant R01 AI097128 (T.S.K. and R.A.C.), NIH, National Cancer Institute grant R01 CA203721 (R.A.C. and T.S.K.), and the AstraZeneca Foundation/Faculty of Medicine of the University of Lisbon Research Grant (T.R.M.).
Authorship
Contribution: A.G., J.E.T., and T.R.M. carried out the experiments and assisted in analyzing data; V.H., C.Y., and R.W. participated in developing the human engrafted mouse model; J.T.O. helped in analyzing gene expression data; R.A.C. designed experiments, analyzed data, and drafted the manuscript and figures; C.L.T. provided human cervix; and T.S.K. provided access to human lung and colon.
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Correspondence: Rachael A. Clark, Department of Dermatology, Brigham and Women’s Hospital, Room 501A, 221 Longwood Ave, Boston, MA 02115; e-mail: rclark1@partners.org.
References
- 1.Sallusto F, Lenig D, Förster R, Lipp M, Lanzavecchia A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature. 1999;401(6754):708-712. [DOI] [PubMed] [Google Scholar]
- 2.Sallusto F, Geginat J, Lanzavecchia A. Central memory and effector memory T cell subsets: function, generation, and maintenance. Annu Rev Immunol. 2004;22(1):745-763. [DOI] [PubMed] [Google Scholar]
- 3.Zaph C, Uzonna J, Beverley SM, Scott P. Central memory T cells mediate long-term immunity to Leishmania major in the absence of persistent parasites. Nat Med. 2004;10(10):1104-1110. [DOI] [PubMed] [Google Scholar]
- 4.Wherry EJ, Teichgräber V, Becker TC, et al. . Lineage relationship and protective immunity of memory CD8 T cell subsets. Nat Immunol. 2003;4(3):225-234. [DOI] [PubMed] [Google Scholar]
- 5.Berger C, Jensen MC, Lansdorp PM, Gough M, Elliott C, Riddell SR. Adoptive transfer of effector CD8+ T cells derived from central memory cells establishes persistent T cell memory in primates. J Clin Invest. 2008;118(1):294-305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gerlach C, Moseman EA, Loughhead SM, et al. . The chemokine receptor CX3CR1 defines three antigen-experienced CD8 T cell subsets with distinct roles in immune surveillance and homeostasis. Immunity. 2016;45(6):1270-1284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wakim LM, Gebhardt T, Heath WR, Carbone FR. Cutting edge: local recall responses by memory T cells newly recruited to peripheral nonlymphoid tissues. J Immunol. 2008;181(9):5837-5841. [DOI] [PubMed] [Google Scholar]
- 8.Clark RA, Chong B, Mirchandani N, et al. . The vast majority of CLA+ T cells are resident in normal skin. J Immunol. 2006;176(7):4431-4439. [DOI] [PubMed] [Google Scholar]
- 9.Watanabe R, Gehad A, Yang C, et al. . Human skin is protected by four functionally and phenotypically discrete populations of resident and recirculating memory T cells. Sci Transl Med. 2015;7(279):279ra39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Campbell DJ, Butcher EC. Rapid acquisition of tissue-specific homing phenotypes by CD4(+) T cells activated in cutaneous or mucosal lymphoid tissues. J Exp Med. 2002;195(1):135-141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Robert C, Kupper TS. Inflammatory skin diseases, T cells, and immune surveillance. N Engl J Med. 1999;341(24):1817-1828. [DOI] [PubMed] [Google Scholar]
- 12.Picker LJ, Michie SA, Rott LS, Butcher EC. A unique phenotype of skin-associated lymphocytes in humans. Preferential expression of the HECA-452 epitope by benign and malignant T cells at cutaneous sites. Am J Pathol. 1990;136(5):1053-1068. [PMC free article] [PubMed] [Google Scholar]
- 13.Hamann A, Andrew DP, Jablonski-Westrich D, Holzmann B, Butcher EC. Role of alpha 4-integrins in lymphocyte homing to mucosal tissues in vivo. J Immunol. 1994;152(7):3282-3293. [PubMed] [Google Scholar]
- 14.Erle DJ, Briskin MJ, Butcher EC, Garcia-Pardo A, Lazarovits AI, Tidswell M. Expression and function of the MAdCAM-1 receptor, integrin alpha 4 beta 7, on human leukocytes. J Immunol. 1994;153(2):517-528. [PubMed] [Google Scholar]
- 15.Roberts AD, Ely KH, Woodland DL. Differential contributions of central and effector memory T cells to recall responses. J Exp Med. 2005;202(1):123-133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Campbell JJ, Murphy KE, Kunkel EJ, et al. . CCR7 expression and memory T cell diversity in humans. J Immunol. 2001;166(2):877-884. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.