

Abstract
Mitogen-activated protein kinase (MAPK or MPK) cascades play key roles in responses to various biotic stresses, as well as in plant growth and development. However, the responses of MPK and MPK kinase (MKK) in Chinese cabbage (Brassica rapa ssp. pekinensis) to Plasmodiophora brassicae, a causal agent of clubroot disease in Brassica crops, are still not clear. In the present study, a total of 11 B. rapa MKK (BraMKK) and 30 BraMPK genes were identified and unevenly distributed in 6 and 10 chromosomes, respectively. The synteny analysis indicated that these genes experienced whole-genome triplication and segmental and tandem duplication during or after the divergence of B. rapa, accompanied by the loss of three MKK and two MPK orthologs of Arabidopsis. The BraMKK and BraMPK genes were classified into four groups with similar intron/exon structures and conserved motifs in each group. A quantitative PCR analysis showed that the majority of BraMKK and BraMPK genes were natively expressed in roots, hypocotyls, and leaves, whereas 5 BraMKK and 16 BraMPK genes up-regulated in the roots upon P. brassicae infection. Additionally, these 5 BraMKK and 16 BraMPK genes exhibited a significantly different expression pattern between a pair of clubroot-resistant/susceptible near-isogenic lines (NILs). Furthermore, the possible modules of MKK-MPK involved in B. rapa-P. brassicae interaction are also discussed. The present study will provide functional clues for further characterization of the MAPK cascades in B. rapa.
Introduction
Biotic stresses are the main threat factors for plant survival and crop production. To defend against pathogen attack, plants have evolved a variety of defense mechanisms at the molecular and cellular levels [1]. One of the earliest defense responses is the activation of mitogen-activated protein kinase (MAPK) cascades when the invading pathogens are sensed by the pattern recognition receptors (PRRs) and resistant (R) proteins [2]. The MAPK cascades are highly conserved in all eukaryotes, defining the key functional signaling modules that usually consist of three gene families—MAPKs or MPKs, MPK kinases (MKKs or MEKs), and MKK kinases (MAPKKKs or MEKKs) [3]. The components of MAPK cascades are sequentially activated via phosphorylation mediated by their upstream kinases [4]. MEKK, the first component of the MAPK cascades, phosphorylates the threonine and serine residues in the conserved S/T-X3–5-S/T domain of MKKs. Finally, the cascades are terminated through the MKK-mediated phosphorylation of tyrosine and threonine residues in the TEY/TDY motifs of MPKs [5]. Subsequently, the activated MPKs phosphorylate the transcription factors and enzymes that participate in the synthesis of antimicrobial metabolites, reprogramming of host genes, and synthesis of phytohormones, such as jasmonic acid (JA) and salicylic acid (SA), triggered by necrotrophic and biotrophic pathogens, respectively [6]. Thus, the stress signals are amplified and transferred into appropriate intracellular responses through MAPK cascades [7].
The genome sequencing projects enable the identification of all members of the MAPK cascades in plant species, including Arabidopsis, rice, pepper, and other crops, allowing for intensive functional studies. A total of 20 MPKs, 10 MKKs, and 80 MEKKs were identified in Arabidopsis [3, 8], whereas 17 MPKs, 8 MKKs, and 74 MEKKs were found in rice [9–11]. However, the expansion of MPK gene families is found in Chinese cabbage, Brassica rapa [12]. Generally, the number of MKKs is much less than that of their upstream and downstream kinases in the cascades. Plant MPKs are phylogenetically classified into four subfamilies, in which groups A, B, and C of MPK contain the conserved TEY domain and group D contains more MPKs sharing the TDY domain. All members of four MKK subfamilies contain the S/T-X3–5-S/T domain. Although the MAPK cascades are evolutionally conserved, sequence diversities exist between the members of both MPK and MKK gene families [8]. Recently, the emergence of a new version of B. rapa genome sequences allowed for the systematic characterization of gene diversity and further MAPK cascades in B. rapa [12].
Previous studies have shown that the transcripts of MPKs, MKKs, and MEKKs are specifically modified by biotic and abiotic stimuli. For instance, the MPK3, MPK4, and MPK6 genes can be activated by pathogen elicitors and regulate innate immune responses in Arabidopsis [13–15]. The AtMKK3 gene can enhance the tolerance against the bacteria Pseudomonas syringae by regulating a pathogenesis-related (PR) gene [14]. OsMPK5 can be induced by various pathogens and abiotic stresses in rice, and has been found to positively regulate drought, cold, and salt tolerance, but negatively regulate the PR gene expression and resistance to fungal (Magnaporthe grisea) and bacterial (Burkholderia glumae) pathogens [16]. Several MAPK cascades involved in plant-pathogen interactions and abiotic stress responses have been functionally characterized. For example, MEKK1-MKK4/5-MPK3/6 and MEKK1-MKK1/2-MPK4 promote the expression of Arabidopsis resistance genes against Pseudomonas syringae; MEKK1-MKK1-MPK4 can transmit drought and wounding signals [15,17]. MAPKKKα/ε-MEK2-SIPK/WIPK/NTF4 and NPK-MEK1-NTF6 induce a hypersensitive response in tobacco [18]; MAPKKKα-MKK2-MPK1/MPK2/MPK3 is involved in disease resistance response in tomato [19]; MKK4-MPK1 is involved in wounding in rice [20]. Furthermore, a single MAPK cascade might be involved in multiple environmental stress responses. For example, the activation of CRLK1-MEKK1/MKK2-MPK4/MPK6 cascade has been reported in salt and cold stress responses in Arabidopsis [21].
Brassica rapa includes a variety of valuable vegetable crops, such as Chinese cabbage, pak choi, turnip, etc., as well as oilseed crops, such as turnip rape and yellow sarson. These and other cruciferous species are seriously challenged by the clubroot disease caused by Plasmodiophora brassicae, a soil-borne, obligate biotrophic parasite. Unlike other plant pathogens, P. brassicae belongs to Rhizaria, a supergroup of protists, and has a distinct intracellular lifestyle with two infection stages, including the primary infection of root hair and the secondary infection of root cortex [22, 23]. After successful infection, P. brassicae leads to the abnormal hyperplasia of roots and subsequent yield losses. Breeding resistant cultivars is considered to be the most efficient approach to control the clubroot disease of Brassica species. So far, several clubroot resistance (CR) genes have been identified and used for resistance breeding [24, 25], and two CR genes with toll interleukin-1 receptor (TIR)–nucleotide binding (NB)–Leu-rich repeat (LRR) domain have been cloned in B. rapa [26, 27]. However, the resistance mechanism remains to be clarified. The functional characterization of P. brassicae-triggered MAPK cascades is a feasible approach for understanding the CR mechanism since MAPK cascades usually act as important signaling modules in plant innate immune systems. However, the mechanism by which the component of MAPK cascades is transcriptionally modified by P. brassicae and the roles of these cascades in B. rapa-P. brassicae interaction remain to be identified.
In this study, we aimed to identify the MKK and MPK family genes of B. rapa to characterize their gene structures and evolution and to analyze gene expression pattern in different tissues of B. rapa. The MKK and MPK genes involved in the interaction between B. rapa and P. brassicae were also identified based on their expression profiles in a pair of Chinese cabbage near-isogenic lines (NILs), carrying the clubroot-resistant or -susceptible genes at the CRb locus. The results obtained in this study have paved the way for a functional study of the P. brassicae-triggered MAPK cascades in B. rapa.
Materials and methods
Identification of MKK and MPK family genes in B. rapa
To identify the MKK and MPK genes in B. rapa, 10 Arabidopsis thaliana MKK and 20 MPK protein sequences were used as the seed sequences to search against B. rapa genome database BRAD (http://brassicadb.org version 1.5 and version 2.0) by using the basic local alignment search tool (BLASTP) program with a cut-off expect value of 100. The genomic and protein sequences of Arabidopsis were downloaded from TAIR (http://www.arabidopsis.org). The predicted B. rapa MKK and MPK genes were nominated as BraMKK1 to BraMKK9 and BraMPK1 to BraMPK20, respectively, as suggested in Arabidopsis nomenclature (S1 Table). To ensure that no more related MPK and MKK genes existed in B. rapa, the identified BraMPKs and BraMKKs were used as query sequences to search against BRAD (http://brassicadb.org) using the BLASTP program. Similarly, B. napus and B. oleracea MPK and MKK genes were identified using the BLAST-like alignment tool (BLAT) CNS-Genoscope database (http://www.genoscope.cns.fr/brassicanapus/) and B. oleracea database (http://brassicadb.org/brad/blastPage.php), and named as in B. rapa.
The databases NCBI-CDD (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi), SMART (http://smart.embl-heidelberg.de/), and Pfam (http://pfam.sanger.ac.uk/) were used to find the conserved domains of BraMKK and BraMPK. The molecular weights (kDa) and isoelectric points (pI) of BraMKKs and BraMPKs were calculated by the Compute pI/Mw tool of ExPASy (http://web.expasy.org/compute_pi/). Their subcellular locations were predicted using TargetP and PProwler in WoLF PSORT (http://www.genscript.com/wolf-psort.html) and BaCelLo (http://gpcr.biocomp.unibo.it/bacello/pred.htm).
Gene structure, conserved domain, motif analysis of BraMKK and BraMPK
The exon/intron structures of BraMKK and BraMPK genes were analyzed by their coding sequences with corresponding genomic sequences and obtained figures using the Gene Structure Display Server (GSDS; http://gsds.cbi.pku.edu.cn/) and adorned by Adobe Photoshop CC (http://www.photoshop.com). The conserved motif structures of BraMKK and BraMPK genes were identified by using the online analysis tool MEME (http://meme-suite.org/tools/meme).
Sequence alignment and phylogenetic analysis of Brassica MPK and MKK
The multiple alignments of all BraMKK, BnaMKK, BolMKK, BraMPK, BnaMPK, and BolMPK protein sequences were fulfilled using Clustal Omega (http://www.ebi.ac.uk/Tools/msa/clustalo/) and tinted by using BoxShade (http://www.ch.embnet.org/software/BOX_form.html). The phylogenetic trees were constructed using the neighbor-joining (NJ) statistical method in MEGA7 program for Brassica MKKs and MPKs. In the phylogenetic tree, the confidence level of each branch was estimated as 0~100 by using 1000 bootstrap replications [28].
Chromosomal location of BraMKK and BraMPK genes
The positional information of all identified MPK and MKK genes in B. rapa, B. napus, B. oleracea and A. thaliana was obtained from the above-mentioned corresponding databases and mapped to their chromosomal locations using Circos (http://circos.ca/). In addition, the BraMKK and BraMPK genes were mapped onto the B. rapa chromosomes using the Mapchart (Version 2.1) software. The evolutionary relationships between BraMKKs, BnaMKK, and BolMKK and between BraMPKs, BnaMPKs, and BolMPKs were analyzed based on their chromosomal locations and compared with Arabidopsis MKKs and MPKs.
Plant materials and treatments with P. brassicae
In our previous study, a pair of Chinese cabbage near-isogenic lines (NILs) carrying either the clubroot-resistant allele of CRbCRb (CR BJN3-2) or the clubroot-susceptible allele of crbcrb (CS BJN3-2) were developed [29]. The CR BJN3-2 and CS BJN3-2 plants were sown in 50-hole plugs containing a sterile substrate and cultivated in a culture room maintained at 24°C under a photoperiod of 16 h light/8 h dark. Previous studies showed that P. brassicae can penetrate the root hairs at 12 hours after infection (hai) [30]. Therefore, the 21-day-old seedlings were inoculated with P. brassicae (107 spores/mL), and then the second true leaves, hypocotyls, and roots were collected at 0, 10, 12, 13, 14, 16, and 20 hai from both CR BJN3-2 and CS BJN3-2 plants. Each treatment was biologically replicated thrice, with each replicate containing five plants. Plants treated with sterile water were sampled at 0 hai and used as controls for the real-time (RT) PCR analysis. All samples were frozen immediately in liquid nitrogen and stored at −80°C until use.
RNA isolation and semi-quantitative and quantitative RT-PCR analyses
Total RNA was extracted from 100 mg of each sample using the TRIZOL reagent (Invitrogen, USA), according to the manufacturer’s instructions. The concentration of RNA samples was determined by using 1% agarose gel electrophoresis and NanoDrop Spectrophotometer (Thermo Scientific, USA). The first strand of cDNA was synthesized using the FastQuant RT Kit (with gDNase) (TIANGEN Biotech, China). The primers used for RT-PCR were designed using the software Primer Premier 5.0 (S4 Table). For BraMKK1-1 and BraMKK1-2, the same primer pair was picked up because of their highly similar sequences.
To select the possible P. brassicae-triggered BraMKK and BraMPK genes, the cDNA pools from the leaves, hypocotyls, and roots of the CS BJN3-2 plants, were made with different samples collected at different time points to perform a semi-quantitative RT-PCR. The expression patterns at different tissues were analyzed. Each reaction was carried out in a volume of 20 μL with 2 μL diluted cDNA, 2 μL specific primers (1.0 μM), 10 μL 2X Premix Taq (TaKaRa Taq Version 2.0 plus dye, Japan), and 6 μL RNase-free ddH2O. The following PCR program was used: initial denaturation at 95°C for 3 min, followed by 35 cycles of denaturation at 95°C for 30 s, annealing at 60°C for 30 s, and extension at 72°C for 30 s, and a final extension at 72°C for 5 min. The actin and 18S rRNA genes of B. rapa were used as the reference genes.
The BraMKK and BraMPK genes, which were differentially expressed in the roots of P. brassicae-treated CS BJN3-2 plants, were used for further analysis by a quantitative RT-PCR. Each reaction was carried out in three independent biological and technical replicates, and contained 2 μL of diluted cDNA, 2 μL of specific primers (1.0 μM), 10 μL of 2X SuperReal PreMix Plus (with SYBR Green I) (TIANGEN Biotech, China), and 6 μL of RNase-free ddH2O in a total volume of 20 μL. The qRT-PCR reactions were conducted in a Bio-Rad CFX96 real-time PCR system (Bio-Rad, USA) using the following reaction conditions: initial denaturation at 95°C for 4 min, followed by 40 cycles of denaturation at 95°C for 10 s, annealing at 63°C for 20 s, and extension at 73°C for 25 s. Following amplification, the melting curves were obtained by increasing the temperatures from 55 to 95°C at the intervals of 0.5°C every 10 s to confirm the specificity of PCR amplification. The actin and 18S rRNA genes of B. rapa were used as reference genes and internal controls for the comparative CT method of quantitation. The relative expression levels of BraMPK and BraMKK genes were calculated by the comparative cycle threshold (ΔΔCT) method.
Results
Identification and annotation of MKK and MPK genes in B. rapa
A total of 11 MKK genes were identified in B. rapa by applying 10 A. thaliana MKK protein sequences as the reference sequences to search against the local Brassica Database (BRAD version 2.0). And then, the identified 11 MKK genes in B. rapa were used as query sequences to search against BRAD (http://brassicadb.org) using the BLASTP program. The two analysis steps suggested an identical result. The orthologous genes of AtMKK7, -8, and -10 were not detectable in B. rapa. In addition, only 30 B. rapa MPK genes homologous to 20 AtMPKs were identified, not 32 as reported in a previous study [12]. The MPK11 and MPK14 genes were missing in B. rapa.
The physicochemical parameters were also predicted for each BraMKK and BraMPK gene. The 11 identified BraMKK proteins ranged from 308 (BraMKK9) to 544 (BraMKK2) amino acids in length. Their relative molecular weights ranged from 34.29 (BraMKK9) to 60.84 (BraMKK2) kDa and their pIs ranged from 5.48 (BraMKK3) to 9.33 (BraMKK4-1). The physicochemical parameters of all proteins are listed in Table 1. The BraMKK proteins showed variable subcellular locations, with four of them located in the cytoplasm, two in the nucleus, two in the mitochondria, and one each in the plasma membrane, peroxisomes, and chloroplasts. Two sets of MKK paralogs (BraMKK1-1/1-2 and BraMKK4-1/4-2) were distributed in different subcellular locations and two sets (BraMKK5-1/5-2 and BraMKK6-1/6-2) in the same subcellular location. Among the 30 BraMPK genes, 11 are located in the nucleus, 10 in cytoskeleton, five in peroxisomes, two in cytoplasm and one each in chloroplasts and vacuoles. Five sets of MPK paralogs (BraMPK6-1/6-2, -7-1/7-2, -12-1/12-2, -17-1/17-2/17-3, and -19-1/19-2) were located in the same subcellular location and five sets (BraMPK8-1/8-2, -10-1/10-2/10-3, -16-1/16-2, -18-1/18-2, and -20-1/20-2) in different locations (S1 Table). These results indicate the functional diversity of MKK and MPK genes in B. rapa.
Table 1. MKK genes in B. rapa genome and their sequence characteristics and physicochemical properties.
| Name | Gene ID | A.thaliana ID | Group | Gene Length (bp) | Protein Length (aa) | Mol. Wt. (KD) | PI | Intron Number | Subcellular location | |
|---|---|---|---|---|---|---|---|---|---|---|
| PProwler | TargetP | |||||||||
| BrMKK1-1 | BraA03005649 | AT4G26070 | A | 2083 | 310 | 34.54361 | 6.19 | 7 | Other | cyto |
| BrMKK1-2 | BraA03005651 | AT4G26070 | A | 2216 | 355 | 39.36718 | 6.71 | 7 | Other | nucl |
| BrMKK2 | BraA01000849 | AT4G29810 | A | 3641 | 544 | 60.84307 | 7.61 | 13 | Other | cyto |
| BrMKK3 | BraA08002528 | AT5G40440 | B | 2265 | 518 | 57.56042 | 5.48 | 8 | Other | nucl |
| BrMKK4-1 | BraA08000717 | AT1G51660 | C | 1066 | 355 | 39.29151 | 9.33 | 0 | CTP | plas |
| BrMKK4-2 | BraA05000831 | AT1G51660 | C | 1012 | 337 | 37.24358 | 9.17 | 0 | Other | pero |
| BrMKK5-1 | BraA01002775 | AT3G21220 | C | 994 | 331 | 36.65592 | 8.12 | 0 | CTP | mito |
| BrMKK5-2 | BraA03004232 | AT3G21220 | C | 991 | 330 | 36.58185 | 8.99 | 0 | MTP | mito |
| BrMKK6-1 | BraA03001347 | AT5G56580 | A | 1874 | 358 | 39.92869 | 5.95 | 7 | MTP | cyto |
| BrMKK6-2 | BraA10001670 | AT5G56580 | A | 1893 | 356 | 39.74451 | 5.94 | 7 | MTP | cyto |
| BrMKK9 | BraA07002948 | AT1G73500 | D | 925 | 308 | 34.28858 | 6.99 | 0 | MTP | chlo |
Phylogenetic and domain analysis of Brassica MPKs and MKKs
To reveal the evolutionary relationships of Brassica MKKs and MPKs, these proteins were also searched in B. oleracea (2n = 18, C genome) and B. napus (2n = 38, AC genome). The above-mentioned nomenclature of MKKs and MPKs was followed. In total, 13 BolMKKs and 25 BolMPKs and 21 BnaMKKs and 54 BnaMPKs were identified from B. oleracea and B. napus, respectively. The number of BnaMKKs and BnaMPKs retrieved in this study was more than that from a previous study [28].
The amino acid sequences of Brassica MKKs or MPKs and the corresponding sequences of Arabidopsis MKKs or MPKs were used to construct a phylogenetic tree. In agreement with Arabidopsis, the Brassica MKKs and MPKs were classified into A, B, C, and D groups (Figs 1 and 2). Notably, the Brassica MKKs in group C show divergence because the branch lengths of group C are shorter than those of the other groups. The multiple alignments of amino acid sequences showed that BraMKKs contained the conserved motif S/T-X3–5-S/T, which is known as the phosphorylation site (S1 Fig), while BraMPKs with a highly conserved TEY motif were classified in groups A, B, and C and those with a TDY motif were classified in group D (S2 Fig).
Fig 1. Phylogenetic tree of MKK genes from B. rapa, B. napus, B. oleracea and A. thaliana.

The unrooted trees were constructed by using ClustalW in MEGA6 via the neighbor-joining (NJ) methods. Different species are indicated in different labels. A. thaliana: ■; B.rapa: ●; B.napus: ◆; B.oleracea: ▲. MKKs of B. rapa, B. napus, B. olearacea and A. thaliana were divided into four groups (A-D) highlighted by different colors, respectively.
Fig 2. Phylogenetic tree of MPK genes from B. rapa, B. napus, B. oleracea and A. thaliana.

The unrooted trees were constructed by using ClustalW in MEGA6 via the neighbor-joining (NJ) methods. Different species are indicated in different labels. A. thaliana: ■; B.rapa: ●; B.napus: ◆; B.oleracea: ▲. MKKs of B. rapa, B. napus, B. olearacea and A. thaliana were divided into four groups (A-D) highlighted by different colors, respectively.
Ten conserved motif domains of BraMKKs and BraMPKs were investigated by aligning protein sequences using the MEME program. All BraMKKs were conserved in their main motifs (1, 2, 4, and 6), while motif diversity was also found between different groups. Motif 7 and 9 belonged to groups A and C, respectively. Group B is more unique; it contains only four main motifs. The paralogous pairs in group C are consistent, while the imparities in group A (BraMKK1-1/1-2 and BraMKK6-1/6-2) are due to the lack of motifs 3 and 7 in BraMKK1-1 and BraMKK6-2, respectively. Groups B and D are imperfect in motif 5 compared to groups A and C (Fig 3A and S3A Fig). BraMPKs possess 8–10 motifs. Motif 6 is specific to group D. Both motifs 6 and 9 were lost in groups A and C, but only motif 6 was lost in group B. These motifs can be used as the indicators of each group of BraMPKs (Fig 3B and S3B Fig).
Fig 3. Motif analysis of MKK (A) and MPK (B) gene families in B. rapa.

Protein sequences of BraMKKs and BraMPKs were used for alignment by MEME online program. BraMKKs and BraMPKs are grouped to the phylogenetic classification (A-D). Different colors of the squares represent different motifs and the grey lines indicate non-conserved sequences. The length of proteins and motifs can be estimated by scale at the bottom.
Structural analysis of BraMKK and BraMPK genes
To further study the gene structural evolution and analyze the structural characteristics, the coding sequences of MKK and MPK genes were aligned to the genomic sequences. The results showed that the BraMKK genes in groups A and B contained 7–8 introns, except for BraMKK2 that contained 13 introns, while the genes belonging to groups C and D had no introns (Fig 4A). The protein lengths of all BraMKKs are similar, indicating that the difference in their gene structures is mainly due to the variation in their intron numbers and lengths. All paralogous gene pairs showed the same pattern of the number of exons/introns and similarity in the length of exons/introns. The intron/exon structures of most BraMPK genes were consistent with the previous study [12]. However, the intron numbers of the BraMPK8-2, -10-1, and -18-2 genes showed differences in version 2.0, and were corrected to be 13, 9, and 6, respectively (Fig 4B).
Fig 4. The exon/intron structure of MKK (A) and MPK (B) genes in B. rapa.

Exons are represented by colorful boxes, and introns are reprensented by black lines. Introns and exons of BraMKKs and BraMPKs are grouped according to the phylogenetic classification (A-D). The exon and intron sizes can be estimated by the scale at the bottom.
Chromosomal location and synteny analysis of Brassica MKK and MPK genes
Based on the physical positions of BraMKK and BraMPK genes, their chromosomal locations were determined on 10 chromosomes of B. rapa (Fig 5). The BraMKK genes were distributed on six chromosomes: four genes on ChrA03, two genes each on ChrA01 and ChrA08, one gene each on ChrA05, -07, and -10. The MKK genes located on ChrA03 showed tandem duplication, while the other paralogous pair of genes was dispersed on two chromosomes. Thirty BraMPK genes were distributed among 10 chromosomes: six genes on ChrA05, five genes on ChrA09, four genes each on ChrA06 and ChrA03, three genes each on ChrA04 and ChrA07, and two genes on ChrA01. Three chromosomes (ChrA02, -08, and -10) contain only one gene. Compared to the Arabidopsis MKK and MPK genes, a majority of BraMKK and BraMPK genes showed segmental duplication and only two genes (BraMPK10-2/10-3 and BraMKK1-1/1-2) showed tandem duplication. These results indicated that segmental duplication played an important role in the gene evolution of BraMKK and BraMPK.
Fig 5. Distribution of BraMPKs and BraMKKs on B. rapa chromosomes.

Paralogous genes were represented by same colors and single-copy genes were colorless. Black arrows indicated the direction of genes in corresponding chromosomes. Chromosome locations of BraMKKs and BraMPKs can be estimated by scale in the left. 11 BraMKKs were mapped to 8 out of 10 chromosomes. 30 BraMPKs were mapped to 6 out of 10 chromosomes.
To comprehensive clarify the mutual relationships of MKK and MPK genes in the A and C genomes of Brassica species, the chromosomal location and orientation of MKK and MPK genes from B. rapa (A genome), B. oleracea (C genome), and B. napus (A, C genomes) were compared with A. thaliana (S2 and S3 Tables). Among the 21 BnaMKK genes, 10 are located in the A genome and the remaining are located in the C genome (Fig 6 and S4 Fig). Out of the 54 BnaMPK genes, 29 and 26 are distributed in the A and C genomes of B. napus, respectively (Fig 7 and S5 Fig). The synteny map revealed that the MKK and MPK family genes are conserved, along with the duplication or loss of some genes (S4 and S5 Figs). The orthologous genes of AtMKK7 were not found in all Brassica crops examined. Further, it was found that the AtMKK8 and AtMKK10 genes are lost in B. rapa and B. oleracea. However, the duplicated genes of AtMKK8 and AtMKK10 are present in both A and C genomes of B. napus. This similar phenomenon was also found in the MPK gene family. MPK11 and MPK14 are absent in both A and C genomes of Brassica species. MPK10 and MPK12 were not detected in B. oleracea, but were found in both B. napus and B. rapa genome.
Fig 6. Synteny mapping of MKK genes in B. rapa, B. napus and A. thaliana chromosomes.

Circle represents each chromosome of B. rapa, B. napus and A. thaliana. Synteny relationships were lined by Circos (http://circos.ca/). Lines with four different colors indicated four groups (A-D) of MKK gene family. Genes located on B. napus A genome are syntenic with genes of B. rapa and A. thaliana.
Fig 7. Synteny mapping of MPK genes in B. rapa, B. napus and A. thaliana chromosomes.

Synteny relationships were lined by Circos (http://circos.ca/). Lines with four different colors indicated four groups (A-D) of MPK gene family. Genes located on B. napus A genome are syntenic with genes of B. rapa and A. thaliana.
Expression profiles of BraMPK and BraMKK genes in different tissues of B. rapa
To elucidate the expression pattern of BraMKK and BraMPK genes in natural condition and identify valuable P. brassicae-triggered MKK and MPK genes, a semi-quantitative RT-PCR was performed (Fig 8). It was found that 10 BraMKK genes were expressed in the roots, hypocotyls, and leaves of B. rapa. BraMKK4-1 was not detected in the leaves. The expression levels of BraMKK genes in groups A and B were weaker than those of the BraMKK genes in groups C and D in all tissues. The expression of BraMKKs showed no difference between different tissues. After inoculation with P. brassicae, four genes (BraMKK4-2, 5–1, 5–2, 9) were up-regulated in roots and hypocotyls. BraMKK4-1 was up-regulated and induced in roots and leaves, respectively, but was down-regulated in hypocotyls. These five BraMKK genes showing up-regulation in roots were selected for further qRT-PCR analysis.
Fig 8. Expression patterns of MKK and MPK genes in different tissues of Chinese cabbage.

Semi-quantitative RT-PCR was used for analyzing tissue specific expressions of MKK (A) and MPK (B) genes in B. rapa (CS NIL). BraMKKs and BraMPKs are grouped to the phylogenetic classification (A-D). Actin and 18s rRNA were used as internal control. CK: The cDNA pools of control samples. Pb: The cDNA pools of treated samples with P. brassicae. R: Root, H: hypocotyl, L: Leaf.
Out of the 30 BraMPKs, one gene was ubiquitously expressed in the roots, hypocotyls, and leaves of B. rapa. Three genes (BraMPK16-1, 18–2, 20–1) were not expressed in the leaves, but were induced after P. brassicae infection. Four genes (BraMPK10-1, 10–2, 10–3, 15) were undetectable in any of the tissues tested, even after treatment with P. brassicae. The pair-wise genes in groups A and B showed similar expression patterns, while different expression patterns were observed in groups C and D. After P. brassicae infection, 16 BraMPK genes showed up-regulation only in roots, and were used for further expression survey with the samples collected at different time points. Among the 16 up-regulated genes, five genes (BraMPK2, 5, 6–1, 16–2, and 19–2) were up-regulated in all tissues, five (BraMPK3, -4, -7-2, -9, and 17–3) were up-regulated in roots and hypocotyls, BraMPK1 and BraMPK6-2 showed up-regulation in roots and leaves. BraMPK7-1, -16-1, -17-2, and -19-1 were specifically up-regulated in roots. Additionally, BraMPK13 and BraMPK20-2 were up-regulated in hypocotyls, but four genes (BraMPK6-2, 17–2, 18–1, 18–2) were down-regulated. Three genes were up-regulated (BraMPK12-1, -12-2, and -20-2) and three genes (BraMPK4, -9, -13) were down-regulated in leaves.
Expression profiles of BraMAPK and BraMKK genes after P. brassicae infection
To determine the possible biological effects of BraMKK and BraMPK genes after stimulation by P. brassicae, 5 BraMKK and 16 BraMPK genes showing up-regulation in the roots upon P. brassicae infection were selected for further expression analysis in the roots of CR and CS NILs at the early-infection stages by real-time PCR. Compared to the control, the majority of BraMKK and BraMPK genes were up-regulated after inoculation, even though some differential expression tendencies were observed between the two NILs.
After P. brassicae infection, the expression levels of all BraMKK genes were higher in CS NIL than in CR NIL (Fig 9, S5 Table). The CS BraMKK genes maintained up-regulation after 12 hai. Although the CR BraMKK genes showed high levels of expression after 12 hai, they recovered previous levels after 14 hai. Interestingly, BraMKK4-1 and BraMKK9 were strongly induced in the CS BJN3-2 plants, but they showed no difference in the CR BJN3-2 plants. It is worth noting that the pair-wise genes (BraMKK4-1/4-2 and BraMKK5-1/5-2) showed different expression trends in the CR BJN3-2 plants, indicating their functional diversity in response to P. brassicae.
Fig 9. Expression levels of MKK (A) and MPK (B) genes in roots after treatment with P. brassicae in both NILs.

Actin and 18s rRNA expression levels were used to normalize the data. All data is the mean of 3 biological replicates ± S.E. Significant differences are showed by * (p< 0.05) and ** (p< 0.01).
The CS BraMPK genes were strongly induced at 13 hai and maintained the high level of expression even though the folds of expression were fluctuating. After 12 hai, the relative expression levels of the pair-wise genes (BraMPK17-2/17-3 and BraMPK19-1/19-2) were different in each period, while those of the other paralogous pairs were similar. In addition, the CR BraMPK genes impetuously reacted at 13 and 14 hai, and restored to normal condition in the later period compared to the CR BraMKK genes. The changes in the relative expression quantities of most BraMPK genes were higher than those of the CS BraMKK genes.
Discussion
The MAPK cascades play important roles not only in plant growth and development, but also in generating response to biotic and abiotic stresses [31]. Some of the MAPK cascades were found to be involved in plant-virus, -bacteria and -fungus interactions [32]. However, there is no systematic study concerning the effect of MAPK cascades during the interaction between B. rapa and P. brassicae. In this study, we identified 11 BraMKK and 30 BrMPK genes in B. rapa; these genes were divided into four groups as in A. thaliana, rice, and other plant species [3, 11, 33, 34]. The use of a pair of near isogenic lines, CR BJN3-2 and CS BJN3-2, allowed us to identify the probable BraMKK and BrMPK genes participating in B. rapa-P. brassicae interaction.
Evolution of BraMPK and BraMKK genes in B. rapa
Since Brassica genomes have undergone whole-genome triplication (WGT) after speciation from A. thaliana [35, 36], the B. rapa genome theoretically contains three copies of AtMPK and AtMKK genes. However, only 11 BraMKK and 30 BraMPK genes homologous to 10 MKK and 20 MPK genes in Arabidopsis genome were found in the B. rapa genome. Only AtMPK10 and AtMPK17 showed triplication, four AtMKK and eight AtMPK genes showed duplication, and five genes (AtMKK7, -8, and -10, AtMPK11, and -14) showed no counterpart in the B. rapa genome. This suggests that more than half of the MPK and MKK genes have been lost during the triplication events of B. rapa genome evolution [37]. Moreover, MKK8 and MKK10 were only present in A and C subgenomes of B. napus, and both MPK10 and MPK12 were detected in B. rapa and B. napus but not in B. oleracea genome. This result indicated that these genes were preserved before speciation, but may lost during evolution or artificial selection. The loss of MPK and MKK genes was also found in other plant species. For example, the homologous genes of AtMKK7, -8 and AtMPK11, -14 were commonly lost in B. rapa, B. oleracea, B. napus, Piper nigrum (pepper), Oryza sativa (rice), and other species [8, 28, 33, 34, 38–40], indicating that they were not selected during the evolution process. Interestingly, AtMKK7, which has no orthologs in B. rapa, has a function similar to AtMKK4 in Arabidopsis [8]. The retention and duplication of AtMKK4 in these three species suggests that MKK4 was probably strongly selected during evolution. In addition, MKK9 was the only gene retained in group D in both Brassica and some other species [11, 39]. In previous studies, MKK9 was reported to directly interact with a numbr of MPKs, including BnaMPK1, -2, -5, -9, -19, and -20 in B. napus [28] and AtMPK5, -10, -17, and -20 in Arabidopsis [41, 42], suggesting a possible functional divergence of MKK9 in Brassica species. Our further results suggest that BraMKK9 might be a key member of the MAPK cascade in B. rapa-P. brassicae interaction since its expression was strongly induced after P. brassica infection in a clubroot-susceptible NIL.
Most of the BraMKK genes have similar number of introns compared to other plant species [11, 33]. However, BraMKK3 contains relatively more introns than the MKK3 genes in other species, such as Malus domestica (8), A. thaliana (7), Zea mays (maize) (8–9) and Capsicum annuum (7). It is commonly accepted that the genes responding to stresses contain a small number of introns [43]. AtMKK4 and AtMKK5, which respond to biotic and abiotic stresses, have no intron [8]. The BraMKK4-1, -4-2, -5-1, -5-2, and -9 genes, which are up-regulated after P. brassicae treatment, also have no intron, the same as C. annuum [33] and B. napus [28]. In addition, AtMPK3, -4, and -6 involved in biotic, abiotic, and hormone responses contain relatively few introns[44, 45]. However, the BraMPK genes with more introns were also found be induced by P. brassicae infection.
Expression profiles of BraMPK and BraMKK genes in different tissues and in response to P. brassicae
In this study, we found that the majority of BraMKK and BraMPK genes are expressed in the leaves, hypocotyls, and roots of Chinese cabbage, suggesting that they are involved in plant growth and development as in other plant species [41, 46]. The expression of BraMPK10-1/-2/-3, and BraMPK15 were undetectable under normal conditions, even after P. brassica infection, indicating that these genes functioned as pseudogenes. The most expressed BraMKK and BraMPK genes showed no differential expression pattern between different organs, although a few genes showed relatively low or no expression in leaves. The low-level expression of MKK and MPK genes was also detected in the leaves of other plant species [33, 34]. The MKK genes in group A showed relatively low expression in the leaves, hypocotyls, and roots of B. rapa. However, the precise biological significance of the uneven expression of these genes remains to be further studied.
The transcriptional levels of MAPK cascades are largely regulated after environmental stimulation, and the genes transcriptionally modified against the challenge of stress might play important roles in plants’ response to stress [47–49]. In general, the components of MAPK cascades are immediately activated in response to the biotic and abiotic stimuli before 12 hai [11, 28, 33]. In this study, half of the BraMKK and BraMPK genes were constantly induced in the roots of the clubroot-susceptible genotype ‘CS BJN3-2’ at 13 hai. This is due to the fact that the spores of P. brassicae can penetrate the root hair at 12 hai [30]. The induction of MKK and MPK genes by the SA treatment and biotic stresses was also found in other plant species [11, 33, 50]. For instance, the AtMKK4 and AtMKK9 genes were activated by the SA treatment, which can regulate the responses to biotrophic pathogens [13]. In agreement, their orthologous genes BraMKK4-1/4-2 and BraMKK9 were highly induced in the CS BJN3-2 plants, but were not changed or slightly repressed in the CR BJN3-2 plants. The up-regulation of these genes suggested that they act as the positive regulators of the B. rapa-P. brassicae interaction. The distinctive expression patterns of the BraMKK and BraMPK genes in the two NILs might have resulted from the expression of the CRb gene. In CR BJN3-2 plants, the R gene could recognize the P. brassicae effectors and transmit the signals to the downstream components. Thereafter, the signals might modulate the defense hormone synthesis and/or signaling, defense gene activation, cell wall modification, and phytoalexin accumulation to defend against pathogens. Subsequently, the expression levels of MKK and MPK genes might recover to the normal level because of the activation of downstream reactions. Remarkably, BraMPK16-2 was strongly induced from 12 hai and constantly expressed at high levels in the CS BJN3-2 plants, suggesting its key role in the B. rapa-P. brassicae interaction. However, no information is available to prove that it was a response to pathogens in any other plant species. Thus, further analysis is needed to determine which BraMKKs and BraMPKs are the direct upstream components of BraMPK16-2.
Possible modules of MKKs and MPKs involved in B. rapa-P. brassicae interactions
Plants have evolved a delicate and complicated immune system to protect plants against the attack by microbial pathogens [7]. Plant PRR proteins can recognize pathogen/microbe-associated molecular patterns (PAMPs) to trigger pattern-triggered-immunity (PTI) signaling, the first layer of inducible immunity. PTI provides the basal resistance to a wide range of pathogens, including fungi, bacteria, oomycetes, nematodes, and viruses [7, 51, 52]. After the recognition of a PAMP by PRR, a complex defense network is activated at the early cellular stages, such as the production of reactive oxygen species (ROS), MAPK cascades phosphorylation, and hormone signaling events [53]. Finally, transcription factors, such as WRKY [54, 55], are phosphorylated to promote the expression of stress resistance-related genes [56].
Numerous studies have functionally characterized several important MAPK cascades involved in plant-microbe interactions. It is well known that the modules of MEKK1-MKK4/5-MPK3/6 and MEKK1-MKK1/MKK2-MPK4 and their downstream targets WRKY22/29 regulate plant immune responses against Pseudomonas syringae in Arabidopsis [13, 15]. The Arabidopsis mutant lines of mkk1mkk2 and mpk4 showed enhanced resistance to biotrophic pathogens and increased susceptibility to necrotrophic fungi [17, 57]. In this study, we found that three pair-wise genes (BraMKK4-1/4-2, BraMKK5-1/5-2, and BraMPK6-1/6-2) and BraMPKs, MPK3 and MPK4, were strongly and continuously activated in the roots of the CS BJN3-2 plants, but BraMKK4-1 and BraMKK5-2 showed weak up-regulation in the CR BJN3-2 plants. Furthermore, the up-regulation of WRKY22/29 was found in our previous study [30]. Therefore, it could be speculated that the elicitors secreted by P. brassicae, a biotrophic pathogen, can induce the activation of BraMKK4-1/4-2 and BraMKK5-1/5-2, which in turn activates BraMPK3, -4, and -6, leading to gall formation in the roots.
The cascade of LeMAPKKKα-LeMKK2-LeMPK1/LeMPK2/LeMPK3 has been identified in tomato-Hyaloperonospora parasitica Noco2 interaction [58]. The expression level of BraMKK2 was not changed after P. brassicae infection, indicating that this cascade was not related to the B. rapa-P. brassicae interaction. Except for the above-mentioned BraMPK genes that are involved in the well-known plant-microbe interaction, several other BraMPKs were found to respond to P. brassicae. In B. napus, BnaMKK9-BnaMPK1, -2, -5, -9, -19, -20, and BnaMKK9-BnaMPK5, -9, -19, -20 were associated with response to P. brassicae. Furthermore, the signaling module of MKK9-MPK19-WRKY20 was reported to be responsible to Sclerotinia sclerotiorum in B. napus [28]. The BraMKK9 and BraMPK19-1/19-2 genes were also significantly induced in the P. brassicae-infected CS BJN3-2. The AtMKK9 gene was reported to induce the biosynthesis of camalexin, which is related to the defense response to P. brassicae in Arabidopsis [59]. In the modules of MKK9-MPK1/2-WRKY53, MKK9-MPK5, and MKK9-MPK9/19/20, the transcripts of BraMKK9, BraMPK1, -2, -5, -9, -19, and -20-1 were increased in B. rapa after P. brassicae infection. The induction of CaMPK3, -4-1, -6-1, -6-2, -7, and -9-1 was revealed in the interaction between C. annuum and Ralstonia solanacearum. However, in the MEKK1-MKK1/2-MPK4 module found in the Arabidopsis-Pseudomonas syringae interaction [60], only BraMPK4-1/4-2 showed up-regulation. The results obtained in this study indicated that the MEKK1-MKK4/5-MPK3/6-WRKY22/29- and MKK9- mediated modules might be involved in the defense response to P. brassicae in B. rapa. Further studies are needed to unveil the precise MAPK cascades involved in B. rapa-P. brassicae interactions by using yeast two hybridization assays.
Conclusion
So far, some components of the MAPK cascades have been studied in some plant species, such as Arabidopsis, B. napus, B. rapa, O. sativa, and others, while no systematic study has been carried out on the interaction of B. rapa with P. brassicae, the causal agent of clubroot disease in Brassicas. In this study, we identified a total of 11 BraMKK and 30 BraMPK genes, which were unevenly distributed in 6 and 10 chromosomes, respectively. Both BraMKKs and BraMPKs were classified into 4 clades, which were identical to the Arabidopsis genes; however, they showed gene duplication and losses. The gene expression analysis revealed that most of the genes were expressed in different tissues. Further analysis indicated that half of the BraMPK and BraMKK genes were expressed in response to P. brassicae infection, and their expression patterns were different between the clubroot-resistant and -susceptible genotypes. We speculated that the possible MAPK cascades involved in P. brassicae-B. rapa interaction were BraMKK4/5-BraMPK1/3/6/7/19 and/ or BraMKK9-BraMPK1/2/5/9/19. The results obtained in this study will provide information to uncover the biological roles of MAPK cascades involved in B. rapa-P. brassicae interaction.
Supporting information
Alignment is performed using Clustal Omega and marked by using BoxShade. Duplicate and similar amino acids are shaded in black and grey, respectively. Amino acids in red box (VGTxxYMSPER) are typical motif of MKKs.
(JPG)
{kind=link}
Alignment is performed using Clustal Omega and marked by using BoxShade. Duplicate and similar amino acids are shaded in black and grey, respectively. Amino acids in red box (T(E/D)YVxTRWYRAPE(L/V)) are typical motif of MPKs.
(JPG)
{kind=link}
(JPG)
{kind=link}
Synteny relationships were lined by Circos. Lines with four different colors indicate four groups (A-D) of MKK gene family. Genes located on B. napus C genome are syntenic with genes of B. oleracea and A. thaliana.
(JPG)
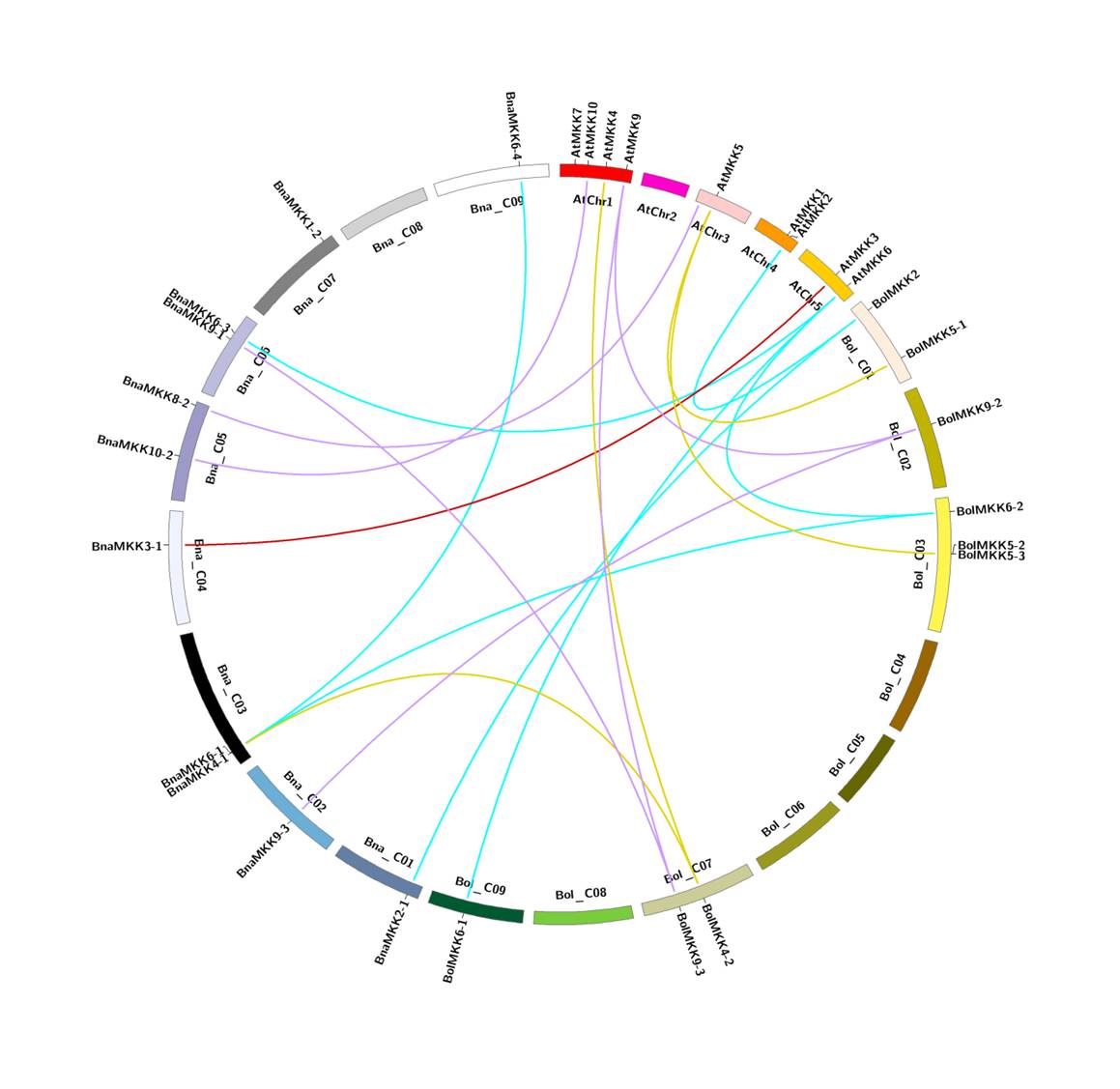{kind=link}
Synteny relationships were lined by Circos. Lines with four different colors indicate four groups (A-D) of MPK gene family. Genes located on B. napus C genome are syntenic with genes of B. oleracea and A. thaliana.
(JPG)
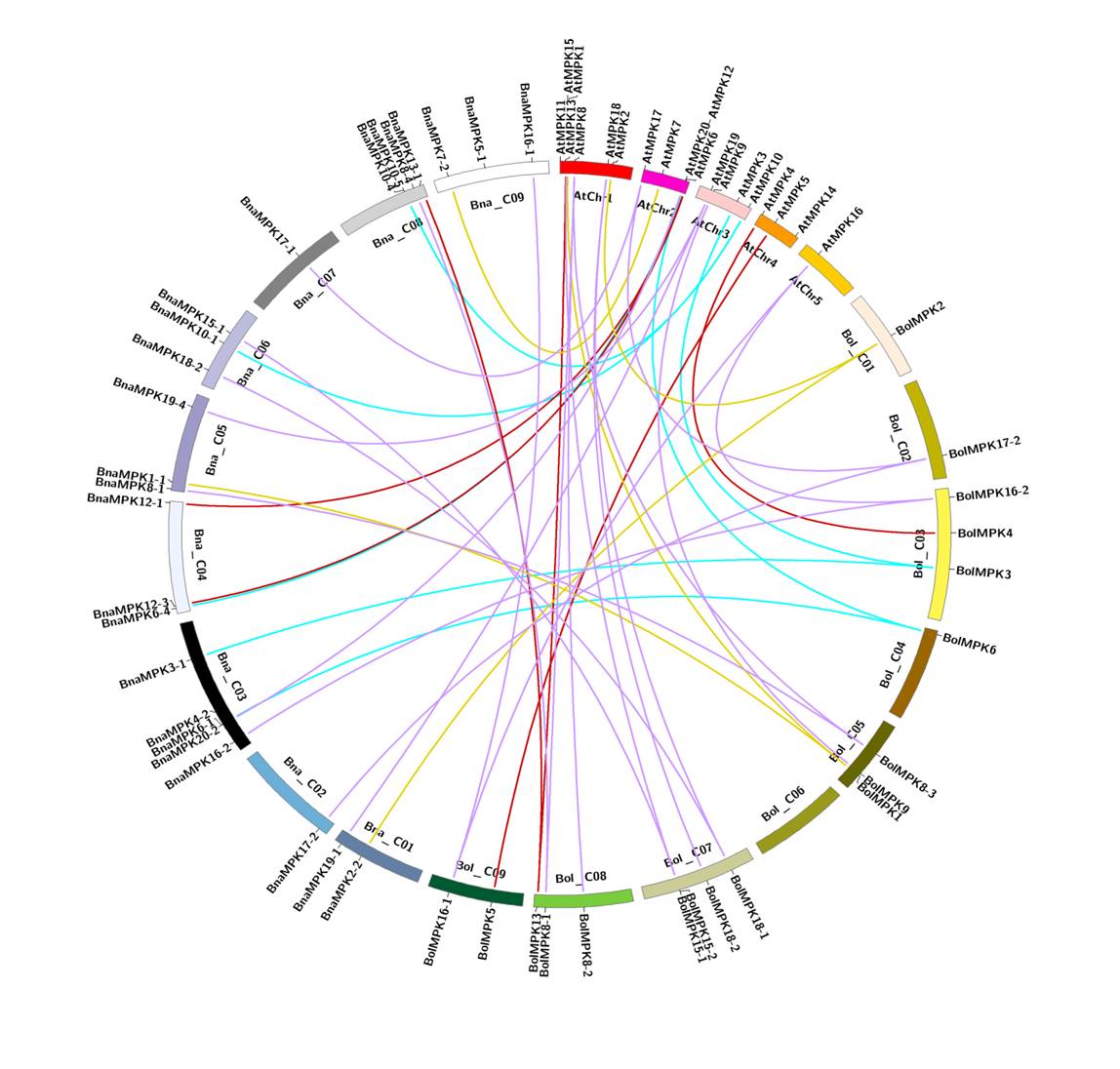{kind=link}
(XLSX)
(XLSX)
(XLSX)
(XLSX)
(XLSX)
Acknowledgments
This work was supported by the National Key Research and Development Program of China (Grant No. 2016YFD0100202-19 and No.2017YFD0101802), the earmarked fund for China Agriculture Research System (CARS-12), and National Natural Science Foundation of China (Grant No. 31471882).
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by the National Key Research and Development Program of China (Grant No. 2016YFD0100202-19 and No.2017YFD0101802), the earmarked fund for China Agriculture Research System (CARS-12), and National Natural Science Foundation of China (Grant No. 31471882). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Chisholm ST, Coaker G, Day B, Staskawicz BJ. Host-microbe interactions: shaping the evolution of the plant immune response. Cell. 2006;124(4):803–14. doi: 10.1016/j.cell.2006.02.008 [DOI] [PubMed] [Google Scholar]
- 2.Dixon RA. Natural products and plant disease resistance. Nature. 2001;411(6839):843–7. doi: 10.1038/35081178 [DOI] [PubMed] [Google Scholar]
- 3.Ichimura K, Shinozaki K, Tena G, Sheen J, Y H. Mitogen-activated protein kinase cascades in plants: a new nomenclature. Trends Plant Sci. 2002;7(7):301–8. [DOI] [PubMed] [Google Scholar]
- 4.Jonak C, Okresz L, Bogre L, Hirt H. Complexity, cross talk and integration of plant MAP kinase signalling. Curr Opin Plant Biol. 2002;5(5):415–24. [DOI] [PubMed] [Google Scholar]
- 5.Chang L, Karin M. Mammalian MAP kinase signalling cascades. Nature. 2001;410(6824):37–40. doi: 10.1038/35065000 [DOI] [PubMed] [Google Scholar]
- 6.Glazebrook J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu Rev Phytopathol. 2005;43:205–27. doi: 10.1146/annurev.phyto.43.040204.135923 [DOI] [PubMed] [Google Scholar]
- 7.Jones JDG, Dangl JL. The plant immune system. Nature. 2006;444(7117):323–9. doi: 10.1038/nature05286 [DOI] [PubMed] [Google Scholar]
- 8.Hamel LP, Nicole MC, Sritubtim S, Morency MJ, Ellis M, Ehlting J, et al. Ancient signals: comparative genomics of plant MAPK and MAPKK gene families. Trends Plant Sci. 2006;11(4):192–8. doi: 10.1016/j.tplants.2006.02.007 [DOI] [PubMed] [Google Scholar]
- 9.Reyna NS, Yang Y. Molecular analysis of the rice MAP kinase gene family in relation to Magnaporthe grisea infection. Mol Plant Microbe Interact. 2006;19(5):530–40. doi: 10.1094/MPMI-19-0530 [DOI] [PubMed] [Google Scholar]
- 10.Rao KP, Richa T, Kumar K, Raghuram B, Sinha AK. In silico analysis reveals 75 members of mitogen-activated protein kinase kinase kinase gene family in rice. DNA Res. 2010;17(3):139–53. doi: 10.1093/dnares/dsq011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Yang Z, Ma H, Hong H, Yao W, Xie W, Xiao J, et al. Transcriptome-based analysis of mitogen-activated protein kinase cascades in the rice response to Xanthomonas oryzae infection. Rice. 2015;8:4 doi: 10.1186/s12284-014-0038-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lu K, Guo W, Lu J, Yu H, Qu C, Tang Z, et al. Genome-wide survey and expression profile analysis of the mitogen-activated protein kinase (MAPK) gene family in Brassica rapa. PLoS One. 2015;10(7):e0132051 doi: 10.1371/journal.pone.0132051 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Asai T, Tena G, Plotnikova J, Willmann MR, Chiu WL, Gomez-Gomez L, et al. MAP kinase signalling cascade in Arabidopsis innate immunity. Nature. 2002;415(6875):977–83. doi: 10.1038/415977a . [DOI] [PubMed] [Google Scholar]
- 14.Dóczi R, Brader G, Pettkó-Szandtner A, Rajh I, Djamei A, Pitzschke A, et al. The Arabidopsis mitogen-activated protein kinase kinase MKK3 is upstream of group C mitogen-activated protein kinases and participates in pathogen signaling. Plant Cell. 2007;19(10):3266–79. doi: 10.1105/tpc.106.050039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gao M, Liu J, Bi D, Zhang Z, Cheng F, Chen S, et al. MEKK1, MKK1/MKK2 and MPK4 function together in a mitogen-activated protein kinase cascade to regulate innate immunity in plants. Cell Res. 2008;18(12):1190–8. doi: 10.1038/cr.2008.300 [DOI] [PubMed] [Google Scholar]
- 16.Xiong L, Yang Y. Disease resistance and abiotic stress tolerance in rice are inversely modulated by an abscisic acid-inducible mitogen-activated protein kinase. Plant Cell. 2003;15(3):745–59. doi: 10.1105/tpc.008714 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Qiu J-L, Zhou L, Yun B-W, Nielsen HB, xf, rn, et al. Arabidopsis Mitogen-activated protein kinase kinases MKK1 and MKK2 have overlapping functions in defense signaling mediated by MEKK1, MPK4, and MKS1. Plant Physiol. 2008;148(1):212–22. doi: 10.1104/pp.108.120006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jin H, Axtell MJ, Dahlbeck D, Ekwenna O, Zhang S, Staskawicz B, et al. NPK1, an MEKK1-like mitogen-activated protein kinase kinase kinase, regulates innate immunity and development in plants. Dev Cell. 2002;3(2):291–7. [DOI] [PubMed] [Google Scholar]
- 19.Melech-Bonfil S, Sessa G. Tomato MAPKKKε is a positive regulator of cell-death signaling networks associated with plant immunity. Plant J. 2010;64(3):379–91. [DOI] [PubMed] [Google Scholar]
- 20.Yoo SJ, Kim SH, Kim MJ, Ryu CM, Kim YC, Cho BH, et al. Involvement of the OsMKK4-OsMPK1 cascade and its downstream transcription factor oswrky53 in the wounding response in rice. Plant Pathol J. 2014;30(2):168–77. doi: 10.5423/PPJ.OA.10.2013.0106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yang T, Shad Ali G, Yang L, Du L, Reddy AS, Poovaiah BW. Calcium/calmodulin-regulated receptor-like kinase CRLK1 interacts with MEKK1 in plants. Plant Signal Behav. 2010;5(8):991–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ingram DS, Tommerup IC. The life history of plasmodiophora brassicae woron. Roy Soc London Proc Ser B Biol Sci. 1972;180(1058):103–12. [Google Scholar]
- 23.Naiki T, Dixon GR, Ikegami H. Quantitative estimation of spore germination of Plasmodiophora brassicae. Trans Br Mycol Soc. 1987;89(4):569–72. [Google Scholar]
- 24.Piao Z, Ramchiary N, Lim YP. Genetics of clubroot resistance in Brassica species. J Plant Growth Regul. 2009;28(3):252–64. [Google Scholar]
- 25.Chu M, Yu F, Falk KC, Liu X, Zhang X, Chang A, et al. Identification of the clubroot resistance gene Rpb1 and introgression of the resistance into canola breeding lines using a marker-assisted approach. Acta Hortic. 2013;1005(1005):599–605. [Google Scholar]
- 26.Ueno H, Matsumoto E, Aruga D, Kitagawa S, Matsumura H, Hayashida N. Molecular characterization of the CRa gene conferring clubroot resistance in Brassica rapa. Plant Mol Biol. 2012;80(6):621–9. doi: 10.1007/s11103-012-9971-5 [DOI] [PubMed] [Google Scholar]
- 27.Hatakeyama K, Suwabe K, Tomita RN, Kato T, Nunome T, Fukuoka H, et al. Identification and characterization of Crr1a, a gene for resistance to clubroot disease (Plasmodiophora brassicae Woronin) in Brassica rapa L. PLoS One. 2013;8(1):e54745 doi: 10.1371/journal.pone.0054745 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Liang W, Yang B, Yu BJ, Zhou Z, Li C, Jia M, et al. Identification and analysis of MKK and MPK gene families in canola (Brassica napus L.). BMC Genomics. 2013;14:392 doi: 10.1186/1471-2164-14-392 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Piao Z, Di W, Miao W, Teng Z. Marker-assisted selection of near isogenic lines for clubroot resistant gene in Chinese cabbage. Acta Hortic Sinica. 2010;37(8):1264–72. [Google Scholar]
- 30.Chen J, Pang W, Chen B, Zhang C, Piao Z. Transcriptome analysis of Brassica rapa near-isogenic lines carrying clubroot-resistant and -susceptible alleles in response to plasmodiophora brassicae during early infection. Front Plant Sci. 2016;6:1183 doi: 10.3389/fpls.2015.01183 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Robinson MJ, Cobb MH. Mitogen-activated protein kinase pathways. Curr Opin Cell Biol. 1997;9(2):180–6. [DOI] [PubMed] [Google Scholar]
- 32.Zhang S, Klessig DF. MAPK cascades in plant defense signaling. Trends Plant Sci. 2001;6(11):520–7. [DOI] [PubMed] [Google Scholar]
- 33.Liu Z, Shi L, Liu Y, Tang Q, Shen L, Yang S, et al. Genome-wide identification and transcriptional expression analysis of mitogen-activated protein kinase and mitogen-activated protein kinase kinase genes in Capsicum annuum. Front Plant Sci. 2015;6:780 doi: 10.3389/fpls.2015.00780 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Yan Y, Wang L, Ding Z, Tie W, Ding X, Zeng C, et al. Genome-Wide Identification and expression analysis of the mitogen-activated protein kinase gene family in Cassava. Front Plant Sci. 2016; 7:1294 doi: 10.3389/fpls.2016.01294 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Cheng F, Liu S, Wu J, Fang L, Sun S, Liu B, et al. BRAD, the genetics and genomics database for Brassica plants. BMC Plant Biol. 2011;11(1):136 doi: 10.1186/1471-2229-11-136 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Cheng F, Mandakova T, Wu J, Xie Q, Lysak MA, Wang X. Deciphering the diploid ancestral genome of the Mesohexaploid Brassica rapa. Plant Cell. 2013;25(5):1541–54. doi: 10.1105/tpc.113.110486 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wang X, Wu J, Liang J, Cheng F, Wang X. Brassica database (BRAD) version 2.0: integrating and mining Brassicaceae species genomic resources. Database (Oxford). 2015; doi: 10.1093/database/bav093 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Liu Q, Xue Q. Computational identification and phylogenetic analysis of the MAPK gene family in Oryza sativa. Plant Physiol Biochem. 2007;45(1):6–14. doi: 10.1016/j.plaphy.2006.12.011 [DOI] [PubMed] [Google Scholar]
- 39.Kong F, Wang J, Cheng L, Liu S, Wu J, Peng Z, et al. Genome-wide analysis of the mitogen-activated protein kinase gene family in Solanum lycopersicum. Gene. 2012;499(1):108–20. doi: 10.1016/j.gene.2012.01.048 [DOI] [PubMed] [Google Scholar]
- 40.Liu Y, Zhang D, Wang L, Li D. Genome-wide analysis of mitogen-activated protein kinase gene family in maize. Plant Mol Biol Rep. 2013;31(6):1446–60. [Google Scholar]
- 41.Lee JS, Wang S, Sritubtim S, Chen JG, Ellis BE. Arabidopsis mitogen-activated protein kinase MPK12 interacts with the MAPK phosphatase IBR5 and regulates auxin signaling. Plant J. 2009;57(6):975–85. doi: 10.1111/j.1365-313X.2008.03741.x [DOI] [PubMed] [Google Scholar]
- 42.Popescu SC, Popescu GV, Bachan S, Zhang Z, Gerstein M, Snyder M, et al. MAPK target networks in Arabidopsis thaliana revealed using functional protein microarrays. Genes Dev. 2009;23(1):80–92. doi: 10.1101/gad.1740009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Xie DW, Wang XN, Fu LS, Sun J, Zheng W, Li ZF. Identification of the trehalose-6-phosphate synthase gene family in winter wheat and expression analysis under conditions of freezing stress. J Genet. 2015;94(1):55–65. [DOI] [PubMed] [Google Scholar]
- 44.Kong Q, Qu N, Gao M, Zhang Z, Ding X, Yang F, et al. The MEKK1-MKK1/MKK2-MPK4 kinase cascade negatively regulates immunity mediated by a mitogen-activated protein kinase kinase kinase in Arabidopsis. Plant Cell. 2012;24(5):2225–36. doi: 10.1105/tpc.112.097253 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zhang Z, Wu Y, Gao M, Zhang J, Kong Q, Liu Y, et al. Disruption of PAMP-induced MAP kinase cascade by a Pseudomonas syringae effector activates plant immunity mediated by the NB-LRR protein SUMM2. Cell Host Microbe. 2012;11(3):253–63. doi: 10.1016/j.chom.2012.01.015 [DOI] [PubMed] [Google Scholar]
- 46.Xu J, Zhang S. Mitogen-activated protein kinase cascades in signaling plant growth and development. Trends plant sci. 2015;20(1):56–64. doi: 10.1016/j.tplants.2014.10.001 [DOI] [PubMed] [Google Scholar]
- 47.Ramonell K, Berrocal-Lobo M, Koh S, Wan J, Edwards H, Stacey G, et al. Loss of function mutation in chitin responsive genes show increased susceptibility to the powdery mildew pathogen Erysiphe cichoracearum. Plant Physiol. 2005;138(2):1027–36. doi: 10.1104/pp.105.060947 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Bartsch M, Gobbato E, Bednarek P, Debey S, Schultze JL, Bautor J, et al. Salicylic Acid-Independent ENHANCED DISEASE SUSCEPTIBILITY1 Signaling in Arabidopsis Immunity and Cell Death Is Regulated by the Monooxygenase FMO1 and the Nudix Hydrolase NUDT7. Plant Cell. 2006;18(4):1038–51. doi: 10.1105/tpc.105.039982 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Knoth C, Eulgem T. The oomycete response gene LURP1 is required for defense against Hyaloperonospora parasitica in Arabidopsis thaliana. Plant J. 2008;55(1):53–64. doi: 10.1111/j.1365-313X.2008.03486.x [DOI] [PubMed] [Google Scholar]
- 50.Wang G, Lovato A, Liang Y-H, Wang M, Chen F, Tornielli Gb, et al. Validation by isolation and expression analyses of MAPK gene family in grapevine (Vitis vinifera). Australian Journal of Grape & Wine Research 2014;20(2):255–62 [Google Scholar]
- 51.Nurnberger T, Brunner F, Kemmerling B, Piater L. Innate immunity in plants and animals: striking similarities and obvious differences. Immunol Rev. 2004;198:249–66. [DOI] [PubMed] [Google Scholar]
- 52.Muthamilarasan M, Prasad M. Plant innate immunity: an updated insight into defense mechanism. Journal of biosciences. 2013;38(2):433–49. [DOI] [PubMed] [Google Scholar]
- 53.Meng X, Zhang S. MAPK cascades in plant disease resistance signaling. Annu Rev Phytopathol. 2013;51(1):245–66. doi: 10.1146/annurev-phyto-082712-102314 [DOI] [PubMed] [Google Scholar]
- 54.Rushton PJ, Somssich IE, Ringler P, Shen QJ. WRKY transcription factors. Trends Plant Sci. 2010;15(5):247–58. doi: 10.1016/j.tplants.2010.02.006 [DOI] [PubMed] [Google Scholar]
- 55.Chi Y, Yang Y, Zhou Y, Zhou J, Fan B, Yu JQ, et al. Protein-protein interactions in the regulation of WRKY transcription factors. Mol Plant. 2013;6(2):287–300. doi: 10.1093/mp/sst026 [DOI] [PubMed] [Google Scholar]
- 56.Eulgem T, Somssich IE. Networks of tWRKY transcription factors in defense signaling. Curr Opin Plant Bio. 2007;10(4):366–71. [DOI] [PubMed] [Google Scholar]
- 57.Petersen M, Brodersen P, Naested H, Andreasson E, Lindhart U, Bo J, et al. Arabidopsis MAP kinase 4 negatively regulates systemic acquired resistance. Cell. 2000;103(7):1111–20. [DOI] [PubMed] [Google Scholar]
- 58.Ekengren SK, Liu Y, Schiff M, Dinesh-Kumar SP, Martin GB. Two MAPK cascades, NPR1, and TGA transcription factors play a role in Pto-mediated disease resistance in tomato. Plant J. 2003;36(6):905–17. [DOI] [PubMed] [Google Scholar]
- 59.Séverine L, Alexandre RS, Christine L, Jocelyne L, Nathalie M, Anne L, et al. Camalexin contributes to the partial resistance of Arabidopsis thaliana to the biotrophic soilborne protist Plasmodiophora brassicae. Front Plant Sci. 2015;6:539 doi: 10.3389/fpls.2015.00539 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Underwood W, Zhang S, He SY. The Pseudomonas syringae type III effector tyrosine phosphatase HopAO1 suppresses innate immunity in Arabidopsis thaliana. Plant J. 2007;52(4):658–72. doi: 10.1111/j.1365-313X.2007.03262.x [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Alignment is performed using Clustal Omega and marked by using BoxShade. Duplicate and similar amino acids are shaded in black and grey, respectively. Amino acids in red box (VGTxxYMSPER) are typical motif of MKKs.
(JPG)
Alignment is performed using Clustal Omega and marked by using BoxShade. Duplicate and similar amino acids are shaded in black and grey, respectively. Amino acids in red box (T(E/D)YVxTRWYRAPE(L/V)) are typical motif of MPKs.
(JPG)
(JPG)
Synteny relationships were lined by Circos. Lines with four different colors indicate four groups (A-D) of MKK gene family. Genes located on B. napus C genome are syntenic with genes of B. oleracea and A. thaliana.
(JPG)
Synteny relationships were lined by Circos. Lines with four different colors indicate four groups (A-D) of MPK gene family. Genes located on B. napus C genome are syntenic with genes of B. oleracea and A. thaliana.
(JPG)
(XLSX)
(XLSX)
(XLSX)
(XLSX)
(XLSX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.