

Abstract
Cultural transmission facilitates the spread of behaviours within social groups and may lead to the establishment of stable traditions in both human and non-human animals. The fidelity of transmission is frequently emphasized as a core component of cultural evolution and as a prerequisite for cumulative culture. Fidelity is often considered a synonym of precise copying of observed behaviours. However, while precise copying guarantees reliable transmission in an ideal static world, it may be vulnerable to realistic variability in the actual environment. Here, we argue that fidelity may be more naturally achieved when the social learning mechanisms incorporate trial-and-error; and that the robustness of social transmission is thereby increased. We employed a simple model to demonstrate how culture that is produced through exact copying is fragile in an (even slightly) noisy world. When incorporating a certain degree of trial-and-error, however, cultures are more readily formed in a stochastic environment and are less vulnerable to rare ecological changes. We suggest that considering trial-and-error learning as a stabilizing component of social transmission may provide insights into cultural evolution in a realistic, variable, world.
This article is part of the theme issue ‘Bridging cultural gaps: interdisciplinary studies in human cultural evolution’.
Keywords: cultural evolution, transmission fidelity, imitation, trial-and-error learning, social learning, copying errors
1. Introduction
In studies of cultural evolution, the fidelity of cultural transmission is often considered a core component of the emergence and maintenance of traditions. In this study, we address the assumptions regarding the underlying mechanisms driving transmission fidelity and present a simple model to investigate the influence of trial-and-error on these processes. We first refer to various studies that interpret fidelity as relating to imitation—learning the precise form of demonstrated actions. We suggest that the particular interpretation of imitation common to such studies does not address the role of trial-and-error processes in skill learning and its potential contribution to cultural transmission. In the next section, we focus on the inherent contradiction between the notion of fidelity as a process of exact copying and the necessity for cultural transmission to withstand different sources of variance, namely copying errors and environmental variability. We outline some of the solutions to this tension that have been posited in the literature and present a simple model in which socially mediated trial-and-error learning can lead to successful transmission, especially when copying errors and environmental variability are taken into account. Finally, we discuss the potential benefits of socially mediated trial-and-error learning and suggest that considering it as an inherent part of cultural transmission may advance our understanding of the evolution of culture.
2. Transmission fidelity: accurate imitation and the potential role of trial-and-error
Social learning can lead to the emergence of cultural traditions by facilitating the spread of group-specific behavioural patterns and maintaining them in the population over the course of successive generations [1]. It is often assumed that the establishment and stability of such traditions require certain levels of copying fidelity [2–9]. The logic behind this is rather intuitive: when innovations (the invention of new behaviours or novel solutions [10]) appear within the population, faithful copying allows them to spread and persist. However, when copying is imprecise, such innovations may quickly disappear as each individual may develop its own behavioural pattern, introducing variance that can impede the formation of shared traditions. Theoretical models exploring cultural transmission dynamics suggest that the probability that an innovation will be established as a stable tradition is positively correlated with transmission fidelity [11] and that fidelity has a strong influence on the build-up of cumulative culture [7,12].
The means by which high transmission fidelity is achieved is often attributed to imitation [2,3,5,8,9,13], emphasizing the importance of precisely copying the detailed actions of the demonstrator (reviewed in [8,14]). Imitation is often presented in striking contrast to individual learning processes, and as distinct from other mechanisms of social learning (such as emulation [15,16] and stimulus enhancement [17]), in which learners pay attention to other aspects of the task rather than to the demonstrated behaviour itself, and then fill in the gaps using trial-and-error learning [14,16]. The exact copying of demonstrated actions is assumed to bypass the potential diluting effect embedded in such mechanisms and to allow culture to be sustained over time.
From a mechanistic point of view, the direct link between exact imitation and the fidelity of cultural transmission may not be as straightforward as it often seems. Recent accounts of imitation as an associative mechanism suggest that it develops gradually through experience [18–20]. As such, it may as well be shaped by trial-and-error learning, as attempts to replicate an observed behaviour are likely to involve deviations from it (for instance, due to memory constraints, inaccuracy in performance or even an inherent tendency for exploration). Furthermore, theories addressing the imitation of complex behaviours suggest that the same trial-and-error learning processes with which imitation is often contrasted may actually be embedded in it. Byrne & Russon [21] suggested that imitation may occur at the ‘programme level’, as individuals copy the hierarchic structure of behaviour rather than its surface form [21,22]. The extent of the similarity between observed and performed behaviours can thus vary, depending on the hierarchical level being copied; and trial-and-error learning may be important for fine-tuning the details of performance. Notably, Galef [23] suggested that imitation involves a template-matching process, in which observers create a representation of the demonstrated behaviour in their memory, and then try to match their own behaviour to this stored representation (in a process similar to song learning in birds [23,24], see also [25]). This matching phase is likely to involve trial-and-error learning, especially when the imitated behaviours are novel or complex.
Furthermore, both empirical studies and theoretical models suggest that social learning processes involving trial-and-error learning can lead to the establishment of viable traditions [26–29]. Examples of such traditions in natural populations are accumulating: young rat pups in Israeli pine forests learn from their mothers a specific pinecone stripping technique [30]; young passerine birds copy the mating songs of adults, in a process that leads to the emergence of local dialects ([31,32], also see [33–35] for examples of vocal learning in mammals); tufted capuchins learn nut-cracking procedures socially [36]; and in various chimpanzee populations, the youngsters learn from adults how to use sticks to dig termites out of their mounds [37] or stones to crack nuts [38] (also see [39] and [40] for descriptions of a range of group-specific behaviours in chimpanzee and orangutan populations). While it is often unclear whether such instances involve imitation or emulation, in all of these documented cases individuals acquire the shared behavioural variants in a long process in which the social information is supplemented by trial-and-error learning [23].
Similarly, human skill learning may also depend on trial-and-error. In young children, exploration play may be seen as self-generated opportunities for learning about environmental affordances [41]. As such, trial-and-error learning can be used for generative hypothesis testing, and the imitation of instrumental skills may involve variability and even innovation rather than strict and high-fidelity copying [42]. In adults, proficiency in tool manufacturing is highly dependent on the extent of previous tool-making experience [43,44] and cannot be established through observation alone as it requires deliberate practice and experimentation [45]. Thus, attempts to use or manufacture tools can also involve a prolonged exploration period, in which trial-and-error helps to guide affordance and perceptual learning [44,46]. Finally, laboratory experiments attempting to replicate cumulative cultural learning have indicated that precise imitation may not be a necessary requirement, as exposure to the end-product alone may also lead to high-fidelity copying [47] and cumulative improvements [48,49].
(a). Cultural transmission in a variable world
Under realistic conditions, the notion of precise imitation as fundamental to cultural transmission raises additional challenges. Social learners are constantly exposed to different sources of noise and variance, which can lead to copying errors of various types. Inaccurate copying may occur due to perceptual errors and variability in demonstration quality (and visibility), as well as to differences in body size, or differences in strength and motor-coordination between observers and demonstrators, all of which may cause individuals to imprecisely perceive different aspects of the observed behaviours [50,51]. Thus, a certain level of flexibility in replication is of utmost importance in order to compensate for such errors and ensure successful transmission.
Environmental variability is also likely to contribute to erroneous copying, because it may often lead to differences between the circumstances in which the demonstrated behaviours were observed, and those encountered during subsequent performances by social learners [52]. A young chimpanzee observing its mother as it digs termites out of the mound [37] will later face a slightly different mound or use a slightly different stick for the same purpose. It will thus be required to modify some of the details of the observed actions in order to be able to dig out the termites successfully for itself. A capuchin monkey observing a conspecific cracking a nut [36] will need to use a different stone, on a nut that differs in its size or that may be positioned at a slightly different angle, in its own attempts to crack nuts. Similarly, a hominin attempting to create a hand-axe needs to translate the observation of the construction of an axe by a fellow hominin and adjust it to the stone in hand. An ability to compensate for different sorts of perceptual errors and flexibility in applying observed behaviours to the current state are hence essential for robust and successful transmission.
Attempts to bridge the gap between cultural stability and the effect of copying errors on transmission fidelity have linked such stability to different factors. At the population level, it has been suggested that individual variation, copying errors and low-fidelity transmission may lead to cultural stability and cumulative cultural adaptation, if they are accompanied by transmission biases and demographic factors [50,53–55]. For instance, when individuals copy the behaviour of the majority (conformity) or of the most successful individuals (prestige bias), such biases may balance the potential negative effects of inaccurate copying and, together with population size, dictate whether the culture will be preserved or lost [53,55]. At the individual level, it has been argued that culture is maintained by a process of reconstruction, in which copying errors are corrected through the use of intrinsic attractors (e.g. [56,57], a similar effect may be produced by inductive biases [58]); or, alternatively, that cultural stability is maintained through different pedagogical adaptations [59,60].
Here, we offer a different approach and suggest that transmission fidelity is naturally achieved: not through exact copying but, rather, by compensating for copying errors. This can be realized by integrating trial-and-error into the process of social learning, which countervails the effect of inaccurate copying and leads to robust cultural transmission. We further suggest that social learning that is mediated through trial-and-error provides learners with valuable information regarding the connections between their actions and related consequences. Such information may help them to improve their performance of the specific behaviour being copied and to cope with environmental variability. We exemplify these ideas in a simple model of cultural transmission of skills that entail interaction with the physical world.
3. The model
In the following simulations, we exemplify a process in which culturally transmitted skills are spread in the population through social learning mediated by different degrees of trial-and-error. The learners in our models range from being ‘perfect imitators’—individuals that copy the demonstration precisely—to individuals that apply varying degrees of trial-and-error in their attempts to replicate the observed behaviour.
In the simulations, social learning is modelled as a process in which an observer attempts to copy a successful demonstrator. In doing so, the observer creates a template of the demonstrated behaviour in its memory and then tries to replicate this behaviour by matching its own performance to the stored template. The observer's learning process encompasses two sources of variance: first, the template represented by the observer may be inaccurate, involving some degree of copying error; and second, the actions performed by the observer may also be inaccurate and entail deviations from the stored template, thus creating a trial-and-error range. In our simulations, we explored the interaction between these sources of variation, focusing on the process in which observers attempt to replicate the cultural trait. We allowed each observer to try repeatedly to perform the socially transmitted behaviour until it had reached a threshold criterion of successful skill acquisition. We then measured the resulting rate of spread of this skill in the population.
The socially mediated trial-and-error learning process, through which the learners acquire the observed behaviour, is guided by feedback in the form of value to the learner. The value of the behaviour might be extrinsic—the ability to extract the nut being cracked from its shell or to successfully create a fully functioning hand-axe. However, the value may also be governed by an intrinsic motivation to copy the behavioural variant being demonstrated, for instance—due to its social value, or to an inherent motivation to copy precisely [23,61]. Note that while such a model may not suffice for retaining arbitrary cultural variants [5], under more realistic natural conditions the copying of instrumental behaviours is often goal-directed and involves the attribution of value of some sort [42,62].
(a). Definitions
Let 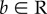 represent a behaviour implementing a certain skill, and Z(b) the value of the behaviour to the performing individual, which can be attributed to the reward obtained (environmental or other, see above). We set the optimal behaviour at 0 (an arbitrarily chosen reference point), and thus, Z receives a maximum value at Z(0) and declines with the distance of b from 0. We assume that when an individual j observes demonstrator i performing bi, it may attempt to copy it if Z(bi) ≥ W (where W is a fixed threshold value that when reached the behaviour is considered successful, see further below). In this case, the observer j will acquire a template bi′ for the copied behaviour (figure 1a). However, bi′ is not a precise copy of bi owing to a copy inaccuracy, and bi′
∼ N(bi, σc). Individual j then repeatedly tries to implement bi′ by performing bj
∼ N(bi′, σI).This is a trial-and-error process that ends when Z(bj) ≥ W (i.e. j has been successful in performing b). Note that for any specific value function Z (with a constant value W, for ‘sufficiently skilled’), two parameters control the dynamics of this process: σc, representing the copy inaccuracy, and σI, representing the learner's trial-and-error range. When σc = 0, the template is completely accurate, and when σI = 0, the template is exactly reproduced.
represent a behaviour implementing a certain skill, and Z(b) the value of the behaviour to the performing individual, which can be attributed to the reward obtained (environmental or other, see above). We set the optimal behaviour at 0 (an arbitrarily chosen reference point), and thus, Z receives a maximum value at Z(0) and declines with the distance of b from 0. We assume that when an individual j observes demonstrator i performing bi, it may attempt to copy it if Z(bi) ≥ W (where W is a fixed threshold value that when reached the behaviour is considered successful, see further below). In this case, the observer j will acquire a template bi′ for the copied behaviour (figure 1a). However, bi′ is not a precise copy of bi owing to a copy inaccuracy, and bi′
∼ N(bi, σc). Individual j then repeatedly tries to implement bi′ by performing bj
∼ N(bi′, σI).This is a trial-and-error process that ends when Z(bj) ≥ W (i.e. j has been successful in performing b). Note that for any specific value function Z (with a constant value W, for ‘sufficiently skilled’), two parameters control the dynamics of this process: σc, representing the copy inaccuracy, and σI, representing the learner's trial-and-error range. When σc = 0, the template is completely accurate, and when σI = 0, the template is exactly reproduced.
Figure 1.

Model value function and illustrated template-matching process. (a) The value function in Models 1 and 2 is a bell-curve encompassing an optimal behaviour and defined as 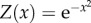 (giving a value Z for a behaviour x). The yellow area depicts a range within which the performance of the behaviour is considered a successful skill performance. Vertical lines illustrate a process of social learning: red: demonstrator's actual performance; green: observer's copied template; grey (dotted): observer's trial-and-error attempts; grey (dashed): observer's successful attempt. (b) Illustration of the value function for Model 3. The value is a function of both the behaviour and the encountered environment and defined as
(giving a value Z for a behaviour x). The yellow area depicts a range within which the performance of the behaviour is considered a successful skill performance. Vertical lines illustrate a process of social learning: red: demonstrator's actual performance; green: observer's copied template; grey (dotted): observer's trial-and-error attempts; grey (dashed): observer's successful attempt. (b) Illustration of the value function for Model 3. The value is a function of both the behaviour and the encountered environment and defined as 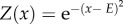 (giving a value Z for a behaviour x and environment E).
(giving a value Z for a behaviour x and environment E).
(b). Simulations
As we are interested in the interplay between σc and σI, we set the value function as the Gaussian: 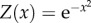 , and set W as 95% of max(Z) (figure 1a). We consider a population of 100 individuals and inspect the spread of a single innovation in that population. In each iteration, each uninformed individual (observer) encounters a randomly selected peer (demonstrator). If the peer is a successful demonstrator (i.e. obtained the value of at least W in the previous iteration), the observer will copy the demonstrator's behaviour, obtaining a template for future implementation. In each iteration, each informed individual, which has already obtained a template, attempts to implement its acquired template. If its last attempt has been successful (rewarded with at least W), it will replicate its previous behaviour (by repeating its previous offset from the template: this constitutes a precise replication, in Models 1 and 2, and will depend on the current environment in Model 3; see below). Otherwise, it will try to replicate its template with a possible error, determined by σI. This trial-and-error process involves stochastic attempts, but may also incorporate learning (in Models 2 and 3), as the observer gradually updates its template according to the reward obtained by its own actions. We measure the assimilation time of the skill: the time until at least 95% of the individuals have demonstrated a skilful behaviour (i.e. were rewarded with at least W for their behaviour) and inspect various combinations of σc and σI.
, and set W as 95% of max(Z) (figure 1a). We consider a population of 100 individuals and inspect the spread of a single innovation in that population. In each iteration, each uninformed individual (observer) encounters a randomly selected peer (demonstrator). If the peer is a successful demonstrator (i.e. obtained the value of at least W in the previous iteration), the observer will copy the demonstrator's behaviour, obtaining a template for future implementation. In each iteration, each informed individual, which has already obtained a template, attempts to implement its acquired template. If its last attempt has been successful (rewarded with at least W), it will replicate its previous behaviour (by repeating its previous offset from the template: this constitutes a precise replication, in Models 1 and 2, and will depend on the current environment in Model 3; see below). Otherwise, it will try to replicate its template with a possible error, determined by σI. This trial-and-error process involves stochastic attempts, but may also incorporate learning (in Models 2 and 3), as the observer gradually updates its template according to the reward obtained by its own actions. We measure the assimilation time of the skill: the time until at least 95% of the individuals have demonstrated a skilful behaviour (i.e. were rewarded with at least W for their behaviour) and inspect various combinations of σc and σI.
We outline three levels of complexity:
(i). Model 1: stochastic trial-and-error
We first simulate the model as described above, where individuals learn socially through inaccurate copying and then apply trial-and-error in a stochastic manner (depending on the value of σI). Repeated attempts are conducted until the learner manages to perform the observed behaviour successfully. Note that in this model, the observer does not learn from its own attempts, but simply continues to try to replicate the socially acquired template, until it reaches the threshold criterion. This model can exemplify, for instance, a young capuchin trying to copy a skilful adult cracking a nut. The template the youngster acquires might represent the strength of the strike, or some other relevant behavioural feature, and could be inaccurate, depending on the value of σc. This inaccuracy (or copying error) can be caused by differences in viewing angle, size appreciation and physical differences between the two capuchins, or the size of the stone being used, etc. The young capuchin then repeatedly attempts to grab a nut and crack it, attempting to reproduce the strike strength represented by the template it has acquired. The similarity between the strength of its strikes and its stored template varies stochastically, depending on the value of σI.
(ii). Model 2: trial-and-error learning with template updating
In the second version of the model, we consider the more realistic possibility that individuals learn during the process of trial-and-error and can thus gradually improve and direct their attempts based on previous experience. Following a short initial sampling period (of 10 attempts), the learners in this model begin to update their template constantly to the mean of their most successful attempts so far (in the simulations: the top-rewarded 25% attempts). If we return to our capuchin example, the learner now gradually adjusts the strength of its strikes according to its most productive attempts. This might be a more realistic scenario than that proposed in Model 1, as individuals constantly learn about the consequences of their actions on the environment.
Note that while this ‘updating’ method may work well in a stable environment in which the task the learner faces is always identical, it may be less efficient when the environment is variable. As ‘no man ever steps into the same river twice’ (Heraclitus of Ephesus), the learner may encounter a slightly different version of the task every time it tries to perform it. In the third, following, version of the model, we sought to test the learners' achievements under such varying environmental conditions.
(iii). Model 3: trial-and-error learning in a variable environment
In the third model, we consider the effect of environmental variability: in each attempt the learner encounters a slightly different variant of the task and needs to adjust its behaviour accordingly. In its attempts to perform the task, the learner can now gain information regarding the relationship between its own behaviour, the relevant environmental features towards which the behaviour is being applied and the consequent reward. Learning about such relationships may help it to direct its future attempts more efficiently.
In this model, the characteristic value function is a two-dimensional function (Z(b, E)) that is also dependent on a varying environmental factor E that is normally distributed (figure 1b; the distribution of E is the same in all simulations). Thus, in each attempt, the learner encounters a slightly different environmental feature. For the sake of simplicity, we model the relationship between the behaviour and its consequences as a linear relationship between the peak of the value function and the environmental factor E. Similarly to Model 2, in its attempts to perform the task the learner can update its template according to its accumulating experience. However, in this model, the template of the behaviour is a linear function of the environmental factor. Thus, rather than simply averaging its top-rewarded attempts, the learner performs a linear regression on these attempts (to estimate the slope a, in 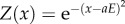 ), and uses the relation between the behaviour and the environment in order to update the template. Note that when the environment is constant, Model 3 reduces to Model 2, i.e. ∀b Z(b, E) = Z(b, 0). Returning to our hypothetical example, as the young capuchin attempts to crack nuts, it may encounter nuts of different sizes. Now, rather than searching for the optimal strike strength, the capuchin should fit each strike to the specific nut at hand (E now represents some relevant feature of the nut, such as its size). In its attempts to solve the task, it may learn that the bigger the nut, the stronger the effective strike strength must be.
), and uses the relation between the behaviour and the environment in order to update the template. Note that when the environment is constant, Model 3 reduces to Model 2, i.e. ∀b Z(b, E) = Z(b, 0). Returning to our hypothetical example, as the young capuchin attempts to crack nuts, it may encounter nuts of different sizes. Now, rather than searching for the optimal strike strength, the capuchin should fit each strike to the specific nut at hand (E now represents some relevant feature of the nut, such as its size). In its attempts to solve the task, it may learn that the bigger the nut, the stronger the effective strike strength must be.
Finally, we used Model 3 to simulate a situation in which, following an initial learning period, the variance of the environmental factor sharply increases. This is analogous to a situation in which the young capuchin learns the nut-cracking skill in a specific location (such as in the vicinity of a limited variety of trees), or at a specific time of the year, and then encounters a greater variety of relevant trees or experiences seasonal changes.
(c). Simulation results
The simulations demonstrate how under even mild copying inaccuracy, a certain degree of trial-and-error is necessary for the spread of a skill in the population. In Models 1 and 2, ‘exact imitators’ (learners whose σI = 0) fail to assimilate the seeded innovation, unless the copying error is exceptionally small (figure 2a,b). In Model 3, exact imitation does not lead to the assimilation of the culture, regardless of the learners' copying error (figure 2c,d). Nevertheless, large trial-and-error deviations also impede the assimilation process.
Figure 2.

The effect of copying inaccuracy and trial-and-error on assimilation rates of the seeded innovation. Assimilation time (number of iterations until 95% of the population acquired the skill) for different combinations of σI (trial-and-error range) and σc (copy inaccuracy) in simulations of (a) Model 1: stochastic trial-and-error; (b) Model 2: trial-and-error with template updating and (c) Model 3: trial-and-error learning in a variable environment. Colours indicate the assimilation time. The white area depicts simulations in which the innovation did not reach the assimilation threshold after 500 iterations. The small circles and their respective regression lines present the fastest assimilation for each value of σc. (d) Assimilation time for different values of σI when σc = 0 in Model 3. Note that, in this model, some degree of trial-and-error produces the most efficient cultural transmission, even without any copying error.
(i). The interplay between copy inaccuracy σc and trial-and-error tendency σI
Intuitively, in all three models, the highest acquisition rates are encountered when σc is correlated with σI (figure 2; regression lines). When σI is too small, the learners fail to correct their copying errors. When σI is too large, the attempts to reproduce the copied behaviour are noisy, reducing the chances of reaching a successful performance. In Model 1, where trial-and-error is applied in a stochastic manner, the highest rate is achieved, not surprisingly, when σI = σc.
(ii). Learning from partial successes
In Model 2, when individuals incorporate learning from their attempts to replicate the desired behaviour, trial-and-error enables faster skill acquisition than in Model 1 (33% shorter assimilation time on average in the simulations) and within a larger range of σI (figure 2b). Specifically, in this scenario, each learner tries to replicate its most beneficial actions, which leads to a constant improvement in its performance. This gradually reduces the gap between the template and the target behaviour, allowing lower values of σI to lead to efficient acquisition. In this learning process, an overly widespread sample (high σI values) encapsulates relatively little information as it includes many unrewarded behaviours, while too small σI values slow down the template updating process. Overall, trial-and-error, in this scenario, provides the learner with a spread-out sample of the possible behavioural variants and their resulting rewards, allowing a rapid convergence towards a successful exhibition of the learned skill.
(iii). Trial-and-error facilitates learning of the physical world
Model 3 demonstrates the power of trial-and-error learning to uncover hidden connections between environmental factors and successful behaviours. When the environment is variable, the learner has an opportunity to relate the outcome of its behaviour to environmental features. In such circumstances, the efficiency of a specific trial-and-error variance (σI) is more dependent on environmental variability (which we maintained the same for all simulations) than on the extent of copy inaccuracy (σc); and the effect of the copying error is reduced (figures 2c and 3a). Even when the template is accurately copied (σc = 0), some degree of trial-and-error is necessary for the culture to be assimilated (figure 2d).
Figure 3.

Simulations of Model 3 under different conditions. (a) Examples of four simulations of Model 3, with different combinations of σI and σc. (b) The effect of changes in environmental variance: examples of three simulations of Model 3, with different combinations of σI and σc, where the variance of the environmental factor increased sharply in iteration no. 250. Note how the top line shows a recovering population, the middle one a partially recovering population, and the lowest line a collapse of the cultural trait. (c) Simulations of Model 3 with different types of learners. Blue: learners that consider environmental variability (as defined in Model 3); red: learners that update their template as in Model 2 (and ignore environmental variability); yellow: a stochastic trial-and-error observer (as in Model 1).
In this scenario, a spread-out sample of the interactions between the behaviour and the environment facilitates a better estimation of their relationship. Note that when the learner does not take into account the environmental variability (as it did in Models 1 and 2), its learning will be inefficient and the skill will not be assimilated into the population (figure 3c).
(iv). Learning by trial-and-error promotes resilience to environmental changes
Populations with different σI levels react differently to a sudden increase in environmental variability (figure 3b). In populations with low levels of trial-and-error variance (σI), most skilful individuals lose their skills owing to the low quality of their constructed templates (i.e. they fail to generalize the relationship between the behaviour and the environment). By contrast, in populations with high σI values, the skilful individuals quickly adapt to the newly encountered environments. This is due to a combination of a higher-quality template (i.e. a better estimation of the underlying environment–behaviour interactions) and the flexibility inherent to their tendency to deviate from the template.
4. Discussion
The outlined models illustrate that trial-and-error can promote robust cultural transmission of seeded innovations as it helps learners to correct copying inaccuracies. When faced with such inaccuracies, certain levels of trial-and-error increase the rate at which the innovation spreads within the population, while ‘exact imitators’ fail to assimilate the observed behaviour. Our models further illustrate that trial-and-error enables individuals to gain information regarding the structure of the cultural trait and can help learners to cope better with environmental variability.
In these models, one of the main contributors to the failure to assimilate the learned behaviours is that of the difficulty encountered in compensating for reduced template quality. In nature, the construction of a poor-quality template may result from a range of potential perceptual errors and differences between observers and demonstrators (e.g. physical attributes or abilities, as noted above). However, the quality of the template is also affected by different cognitive capacities, which may substantially vary between species [63]. Animals may differ in their motivation to copy the behaviour of others, and in the extent to which they pay attention, or give weight, to relevant social stimuli [64,65]. Such input mechanisms will dictate which aspects of the observed behaviour will most attract the learner's attention, perhaps causing its representation of this behaviour to be incomplete. For instance, the learner might only note the body movements of the demonstrator, the manipulated objects, other elements of the behaviour or any combination of these possibilities (e.g. [16], also see [25]). Finally, the quality of the template is also likely to be affected by memory constraints, which may also differ among species [64,65]. As noted above, when trial-and-error learning is incorporated into the social learning process, it can mitigate the effect of copying inaccuracies, at least to some extent. Notably, when copying errors are constantly balanced by trial-and-error, stable cultures may arise even in the absence of social biases such as conformity or prestige-based copying (e.g. [50,53]).
Alongside the compensation for copying inaccuracies, socially mediated trial-and-error can provide learners with additional advantages. First, this process yields a spread-out sample of the possible behavioural variants and their consequences (as illustrated in Model 2), and can enhance the understanding of object affordances and states and their interaction with bodily movements (e.g. [41,66]). Second, having a diverse sample can enable the learner to evaluate the covariance between environmental factors and successful actions. Realization of such dependencies facilitates generalization, and coping with environmental variability and instability (as illustrated in Model 3), and allows individuals to adjust their behaviour to changes in circumstances (e.g. [67]). This may be even more pronounced when the behaviour and the environment are multidimensional, and the covariance may encompass multiple dependencies. Furthermore, trial-and-error may also enable deliberate experimentation [41,45] and provide information about the conditions that involve only partial success, or even failure, in performance. Finally, social learning itself can be more effective when the information is gained through self-experience [68].
Trial-and-error may be especially important when social learning opportunities are confined in time or space. In many species, the period in which social learning of new skills is particularly likely to occur is during the early life stages, when young individuals follow their knowledgeable parents (or other adults). At this stage, they are often exposed to social information that may not be available later on in life (for instance, owing to reduced social tolerance, solitary life stages, etc.; e.g. [30,37,69–71]). These young individuals may also only be exposed to a limited part of the environment, for instance owing to their development during a specific temporal season or within a confined home range. Thus, the environment they experience when opportunities for social learning abound may not encompass the whole range of variability they are likely to encounter later on in life. Social learning that is mediated through trial-and-error may be especially beneficial in such cases, as it can be more robust and expand the learners' acquaintance with relevant environmental features. Such learning can thus be perceived as part of the general tendency of young individuals (including human children) to explore and to engage in play behaviour [72].
Note that the extent of trial-and-error may not be fixed, and may vary between contexts: in human children, it has been shown that the fidelity of imitation decreases when the socially acquired behaviours are presented in instrumental rather than conventional contexts [42,73,74], and changes with age [74,75], or according to demonstration efficacy [75]. Naturally, a too large extent of trial-and-error is likely to diminish the influence of social information and impede the process of cultural transmission.
Furthermore, trial-and-error processes are often assumed to incur energetic costs and may lead individual learners to less profitable outcomes or to potentially dangerous exploration [2,76]. Yet, socially mediated trial-and-error learning can bypass some of these costs, when the template is not too far from the observed behaviour, and the range of attempts is restricted. Such learning promotes exploration but limits it to a specific part of the environment, which helps learners to avoid dangerous situations and refrain from futile behaviours (e.g. [77]).
Finally, accounts of social learning as a process involving trial-and-error may also be important for our understanding of the evolution of cumulative culture. In many of the current models of cumulative culture, advancement appears either through copying errors combined with social learning biases (e.g. [50,53]) or through processes of innovation, modification and the combination of traits (e.g. [7,78–80]). Trial-and-error can facilitate a rich representation of the world and an ability to generalize among contexts, essential for creative processes [81]. As such, it can lead to both accidental innovations and creative modifications, and contribute to the gradual improvement of cumulative culture.
Acknowledgements
We thank Arnon Lotem, Oren Kolodny, Na'ama Aljadeff and Asaf Moran for useful comments and fruitful discussions.
Data accessibility
Data deposited in Dryad: http://dx.doi.org/10.5061/dryad.4m518 [82].
Author contributions
Both authors contributed equally to this work.
Competing interests
The authors have no competing interests.
Funding
We received no funding for this study.
References
- 1.Laland KN, Hoppitt W. 2003. Do animals have culture? Evol. Anthropol. 12, 150–159. ( 10.1002/evan.10111) [DOI] [Google Scholar]
- 2.Boyd R, Richerson PJ. 1985. Culture and the evolutionary process. Chicago, IL: University of Chicago Press. [Google Scholar]
- 3.Tomasello M, Kruger AC, Ratner HH. 1993. Cultural learning. Behav. Brain Sci. 16, 495–511. ( 10.1017/S0140525X0003123X) [DOI] [Google Scholar]
- 4.Whiten A, McGuigan N, Marshall-Pescini S, Hopper LM. 2009. Emulation, imitation, over-imitation and the scope of culture for child and chimpanzee. Phil. Trans. R. Soc. B 364, 2417–2428. ( 10.1098/rstb.2009.0069) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Claidiere N, Sperber D. 2010. Imitation explains the propagation, not the stability of animal culture. Proc. R. Soc. B 277, 651–659. ( 10.1098/rspb.2009.1615) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Mesoudi A. 2011. Cultural evolution: how Darwinian theory can explain human culture and synthesize the social sciences. Chicago, IL: University of Chicago Press. [Google Scholar]
- 7.Lewis HM, Laland KN. 2012. Transmission fidelity is the key to the build-up of cumulative culture. Phil. Trans. R. Soc. B 367, 2171–2180. ( 10.1098/rstb.2012.0119) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dean LG, Vale GL, Laland KN, Flynn E, Kendal RL. 2014. Human cumulative culture: a comparative perspective. Biol. Rev. 89, 284–301. ( 10.1111/brv.12053) [DOI] [PubMed] [Google Scholar]
- 9.Fridland ER. In press. Do as I say and as I do: imitation, pedagogy and cumulative culture. Mind. Lang. [Google Scholar]
- 10.Reader SM, Laland KN. 2003. Animal innovation. Oxford, UK: Oxford University Press. [Google Scholar]
- 11.Enquist M, Strimling P, Eriksson K, Laland K, Sjostrand J. 2010. One cultural parent makes no culture. Anim. Behav. 79, 1353–1362. ( 10.1016/j.anbehav.2010.03.009) [DOI] [Google Scholar]
- 12.Kempe M, Lycett SJ, Mesoudi A. 2014. From cultural traditions to cumulative culture: parameterizing the differences between human and nonhuman culture. J. Theor. Biol. 359, 29–36. ( 10.1016/j.jtbi.2014.05.046) [DOI] [PubMed] [Google Scholar]
- 13.Tennie C, Call J, Tomasello M. 2009. Ratcheting up the ratchet: on the evolution of cumulative culture. Phil. Trans. R. Soc. B 364, 2405–2415. ( 10.1098/rstb.2009.0052) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zentall TR. 2001. Imitation in animals: evidence, function, and mechanisms. Cybernet. Syst. 32, 53–96. ( 10.1080/019697201300001812) [DOI] [Google Scholar]
- 15.Custance D, Whiten A, Fredman T. 1999. Social learning of an artificial fruit task in capuchin monkeys (Cebus apella). J. Comp. Psychol. 113, 13–23. ( 10.1037/0735-7036.113.1.13) [DOI] [Google Scholar]
- 16.Hoppitt W, Laland KN. 2008. Social processes influencing learning in animals: a review of the evidence. Adv. Stud. Behav. 38, 105–165. ( 10.1016/s0065-3454(08)00003-x) [DOI] [Google Scholar]
- 17.Heyes CM. 1994. Social-learning in animals: categories and mechanisms. Biol. Rev. Camb. Phil. Soc. 69, 207–231. ( 10.1111/j.1469-185X.1994.tb01506.x) [DOI] [PubMed] [Google Scholar]
- 18.Heyes CM, Ray ED. 2000. What is the significance of imitation in animals? Adv. Stud. Behav. 29, 215–245. ( 10.1016/S0065-3454(08)60106-0) [DOI] [Google Scholar]
- 19.Cook R, Bird G, Catmur C, Press C, Heyes C. 2014. Mirror neurons: from origin to function. Behav. Brain Sci. 37, 177–192. ( 10.1017/S0140525X13000903) [DOI] [PubMed] [Google Scholar]
- 20.Oostenbroek J, Suddendorf T, Nielsen M, Redshaw J, Kennedy-Costantini S, Davis J, Clark S, Slaughter V. 2016. Comprehensive longitudinal study challenges the existence of neonatal imitation in humans. Curr. Biol. 26, 1334–1338. ( 10.1016/j.cub.2016.03.047) [DOI] [PubMed] [Google Scholar]
- 21.Byrne RW, Russon AE. 1998. Learning by imitation: a hierarchical approach. Behav. Brain Sci. 21, 667–684. ( 10.1017/s0140525X98001745) [DOI] [PubMed] [Google Scholar]
- 22.Byrne RW. 1999. Imitation without intentionality. Using string parsing to copy the organization of behaviour. Anim. Cogn. 2, 63–72. ( 10.1007/s100710050025) [DOI] [Google Scholar]
- 23.Galef BG. 2015. Laboratory studies of imitation/field studies of tradition: towards a synthesis in animal social learning. Behav. Processes 112, 114–119. ( 10.1016/j.beproc.2014.07.008) [DOI] [PubMed] [Google Scholar]
- 24.Galef BG. 2013. Imitation and local enhancement: detrimental effects of consensus definitions on analyses of social learning in animals. Behav. Processes 100, 123–130. ( 10.1016/j.beproc.2013.07.026) [DOI] [PubMed] [Google Scholar]
- 25.Truskanov N, Lotem A. 2017. Trial-and-error copying of demonstrated actions reveals how fledglings learn to ‘imitate’ their mothers. Proc. R. Soc. B 284, 2744 ( 10.1098/rspb.2016.2744) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Franz M, Matthews LJ. 2010. Social enhancement can create adaptive, arbitrary and maladaptive cultural traditions. Proc. R. Soc. B 277, 3363–3372. ( 10.1098/rspb.2010.0705) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Matthews LJ, Paukner A, Suomi SJ. 2010. Can traditions emerge from the interaction of stimulus enhancement and reinforcement learning? An experimental model. Am. Anthropol. 112, 257–269. ( 10.1111/j.1548-1433.2010.01224.x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Alem S, Perry CJ, Zhu XF, Loukola OJ, Ingraham T, Sovik E, Chittka L. 2016. Associative mechanisms allow for social learning and cultural transmission of string pulling in an insect. PLoS Biol. 14, 28 ( 10.1371/journal.pbio.1002564) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.van der Post DJ, Franz M, Laland KN. 2017. The evolution of social learning mechanisms and cultural phenomena in group foragers. BMC Evol. Biol. 17, 49 ( 10.1186/s12862-017-0889-z) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Aisner R, Terkel J. 1992. Ontogeny of pine-cone opening behavior in the black rat, Rattus rattus. Anim. Behav. 44, 327–336. ( 10.1016/0003-3472(92)90038-B) [DOI] [Google Scholar]
- 31.Marler P, Tamura M. 1964. Culturally transmitted patterns of vocal behavior in sparrows. Science 146, 1483–1486. ( 10.1126/science.146.3650.1483) [DOI] [PubMed] [Google Scholar]
- 32.Catchpole CK, Slater PJ. 2003. Bird song: biological themes and variations. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 33.Tyack PL. 2008. Convergence of calls as animals form social bonds, active compensation for noisy communication channels, and the evolution of vocal learning in mammals. J. Comp. Psychol. 122, 319 ( 10.1037/a0013087) [DOI] [PubMed] [Google Scholar]
- 34.Garland EC, Goldizen AW, Rekdahl ML, Constantine R, Garrigue C, Hauser ND, Poole MM, Robbins J, Noad MJ. 2011. Dynamic horizontal cultural transmission of humpback whale song at the ocean basin scale. Curr. Biol. 21, 687–691. ( 10.1016/j.cub.2011.03.019) [DOI] [PubMed] [Google Scholar]
- 35.Prat Y, Azoulay L, Dor R, Yovel Y. 2017. Crowd vocal learning induces vocal dialects in bats: playback of conspecifics shapes fundamental frequency usage by pups. PLoS Biol. 15, e2002556 ( 10.1371/journal.pbio.2002556) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Coelho CG, Falotico T, Izar P, Mannu M, Resende BD, Siqueira JO, Ottoni EB. 2015. Social learning strategies for nut-cracking by tufted capuchin monkeys (Sapajus spp.). Anim. Cogn. 18, 911–919. ( 10.1007/s10071-015-0861-5) [DOI] [PubMed] [Google Scholar]
- 37.Lonsdorf EV. 2006. What is the role of mothers in the acquisition of termite-fishing behaviors in wild chimpanzees (Pan troglodytes schweinfurthii)? Anim. Cogn. 9, 36–46. ( 10.1007/s10071-005-0002-7) [DOI] [PubMed] [Google Scholar]
- 38.Biro D, Inoue-Nakamura N, Tonooka R, Yamakoshi G, Sousa C, Matsuzawa T. 2003. Cultural innovation and transmission of tool use in wild chimpanzees: evidence from field experiments. Anim. Cogn. 6, 213–223. ( 10.1007/s10071-003-0183-x) [DOI] [PubMed] [Google Scholar]
- 39.Whiten A, Goodall J, McGrew WC, Nishida T, Reynolds V, Sugiyama Y, Tutin CEG, Wrangham RW, Boesch C. 1999. Cultures in chimpanzees. Nature 399, 682–685. ( 10.1038/21415) [DOI] [PubMed] [Google Scholar]
- 40.van Schaik CP, Ancrenaz M, Borgen G, Galdikas B, Knott CD, Singleton I, Suzuki A, Utami SS, Merrill M. 2003. Orangutan cultures and the evolution of material culture. Science 299, 102–105. ( 10.1126/science.1078004) [DOI] [PubMed] [Google Scholar]
- 41.Lockman JJ. 2000. A perception–action perspective on tool use development. Child Dev. 71, 137–144. ( 10.1111/1467-8624.00127) [DOI] [PubMed] [Google Scholar]
- 42.Legare CH, Nielsen M. 2015. Imitation and innovation: the dual engines of cultural learning. Trends Cogn. Sci. 19, 688–699. ( 10.1016/j.tics.2015.08.005) [DOI] [PubMed] [Google Scholar]
- 43.Geribàs N, Mosquera M, Vergès JM. 2010. What novice knappers have to learn to become expert stone toolmakers. J. Archaeol. Sci. 37, 2857–2870. ( 10.1016/j.jas.2010.06.026) [DOI] [Google Scholar]
- 44.Nonaka T, Bril B, Rein R. 2010. How do stone knappers predict and control the outcome of flaking? Implications for understanding early stone tool technology. J. Hum. Evol. 59, 155–167. ( 10.1016/j.jhevol.2010.04.006) [DOI] [PubMed] [Google Scholar]
- 45.Stout D. 2011. Stone toolmaking and the evolution of human culture and cognition. Phil. Trans. R. Soc. B 366, 1050–1059. ( 10.1098/rstb.2010.0369) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Stout D, Bril B, Roux V, DeBeaune S, Gowlett J, Keller C, Wynn T, Stout D. 2002. Skill and cognition in stone tool production: an ethnographic case study from Irian Jaya. Curr. Anthropol. 43, 693–722. ( 10.1086/342638) [DOI] [Google Scholar]
- 47.Caldwell CA, Schillinger K, Evans CL, Hopper LM. 2012. End state copying by humans (Homo sapiens): implications for a comparative perspective on cumulative culture. J. Comp. Psychol. 126, 161–169. ( 10.1037/a0026828) [DOI] [PubMed] [Google Scholar]
- 48.Caldwell CA, Millen AE. 2009. Social learning mechanisms and cumulative cultural evolution: is imitation necessary? Psychol. Sci. 20, 1478–1483. ( 10.1111/j.1467-9280.2009.02469.x) [DOI] [PubMed] [Google Scholar]
- 49.Zwirner E, Thornton A. 2015. Cognitive requirements of cumulative culture: teaching is useful but not essential. Sci. Rep. 5, e16781 (doi:1038/srep16781) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Henrich J, Boyd R. 2002. On modeling cognition and culture: why cultural evolution does not require replication of representations. J. Cogn. Cult. 2, 87–112. ( 10.1163/156853702320281836) [DOI] [Google Scholar]
- 51.Eerkens JW, Lipo CP. 2005. Cultural transmission, copying errors, and the generation of variation in material culture and the archaeological record. J. Anthropol. Archaeol. 24, 316–334. ( 10.1016/j.jaa.2005.08.001) [DOI] [Google Scholar]
- 52.Lotem A, Halpern JY, Edelman S, Kolodny O. 2017. The evolution of cognitive mechanisms in response to cultural innovations. Proc. Natl Acad. Sci. USA 114, 7915–7922. ( 10.1073/pnas.1620742114) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Henrich J. 2004. Demography and cultural evolution: How adaptive cultural processes can produce maladaptive losses—the Tasmanian case. Am. Antiquity 69, 197–214. ( 10.2307/4128416) [DOI] [Google Scholar]
- 54.Powell A, Shennan S, Thomas MG. 2009. Late Pleistocene demography and the appearance of modern human behavior. Science 324, 1298–1301. ( 10.1126/science.1170165) [DOI] [PubMed] [Google Scholar]
- 55.Derex M, Beugin M-P, Godelle B, Raymond M. 2013. Experimental evidence for the influence of group size on cultural complexity. Nature 503, 389 ( 10.1038/nature12774) [DOI] [PubMed] [Google Scholar]
- 56.Sperber D. 1996. Explaining culture: a naturalistic approach. Oxford, UK: Blackwell. [Google Scholar]
- 57.Claidière N, Sperber D. 2007. The role of attraction in cultural evolution. J. Cogn. Cult. 7, 89–111. ( 10.1163/156853707X171829) [DOI] [Google Scholar]
- 58.Griffiths TL, Kalish ML, Lewandowsky S. 2008. Theoretical and empirical evidence for the impact of inductive biases on cultural evolution. Phil. Trans. R. Soc. B 363, 3503–3514. ( 10.1098/rstb.2008.0146) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Castro L, Toro MA. 2004. The evolution of culture: from primate social learning to human culture. Proc. Natl Acad. Sci. USA 101, 10 235–10 240. ( 10.1073/pnas.0400156101) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Andersson C. 2013. Fidelity and the emergence of stable and cumulative sociotechnical systems. PaleoAnthropology 2013, 88–103. ( 10.4207/PA.2013.ART81) [DOI] [Google Scholar]
- 61.Gadagkar V, Puzerey PA, Chen R, Baird-Daniel E, Farhang AR, Goldberg JH. 2016. Dopamine neurons encode performance error in singing birds. Science 354, 1278–1282. ( 10.1126/science.aah6837) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Galef BG. 1995. Why behavior patterns that animals learn socially are locally adaptive. Anim. Behav. 49, 1325–1334. ( 10.1006/anbe.1995.0164) [DOI] [Google Scholar]
- 63.Shettleworth SJ. 2010. Cognition, evolution, and behavior, 2nd edn Oxford, UK: Oxford University Press. [Google Scholar]
- 64.Heyes C. 2012. What's social about social learning? J. Comp. Psychol. 126, 193–202. ( 10.1037/a0025180) [DOI] [PubMed] [Google Scholar]
- 65.Lotem A, Halpern JY. 2012. Coevolution of learning and data-acquisition mechanisms: a model for cognitive evolution. Phil. Trans. R. Soc. B 367, 2686–2694. ( 10.1098/rstb.2012.0213) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Visalberghi E, Addessi E, Truppa V, Spagnoletti N, Ottoni E, Izar P, Fragaszy D. 2009. Selection of effective stone tools by wild bearded capuchin monkeys. Curr. Biol. 19, 213–217. ( 10.1016/j.cub.2008.11.064) [DOI] [PubMed] [Google Scholar]
- 67.Tumer EC, Brainard MS. 2007. Performance variability enables adaptive plasticity of ‘crystallized’ adult birdsong. Nature 450, 1240 ( 10.1038/nature06390) [DOI] [PubMed] [Google Scholar]
- 68.Truskanov N, Lotem A. 2015. The importance of active search for effective social learning: an experimental test in young passerines. Anim. Behav. 108, 165–173. ( 10.1016/j.anbehav.2015.07.031) [DOI] [Google Scholar]
- 69.Thornton A, McAuliffe K. 2006. Teaching in wild meerkats. Science 313, 227–229. ( 10.1126/science.1128727) [DOI] [PubMed] [Google Scholar]
- 70.Slagsvold T, Wiebe KL. 2007. Learning the ecological niche. Proc. R. Soc. B 274, 19–23. ( 10.1098/rspb.2006.3663) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Dell'Mour V, Range F, Huber L. 2009. Social learning and mother's behavior in manipulative tasks in infant marmosets. Am. J. Primatol. 71, 503–509. ( 10.1002/ajp.20682) [DOI] [PubMed] [Google Scholar]
- 72.Bateson P, Martin P. 2013. Play, playfulness, creativity and innovation. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 73.Legare CH, Wen NJ, Herrmann PA, Whitehouse H. 2015. Imitative flexibility and the development of cultural learning. Cognition 142, 351–361. ( 10.1016/j.cognition.2015.05.020) [DOI] [PubMed] [Google Scholar]
- 74.Clegg JM, Legare CH. 2016. Instrumental and conventional interpretations of behavior are associated with distinct outcomes in early childhood. Child Dev. 87, 527–542. ( 10.1111/cdev.12472) [DOI] [PubMed] [Google Scholar]
- 75.Carr K, Kendal RL, Flynn EG. 2015. Imitate or innovate? Children's innovation is influenced by the efficacy of observed behaviour. Cognition 142, 322–332. ( 10.1016/j.cognition.2015.05.005) [DOI] [PubMed] [Google Scholar]
- 76.Kendal RL, Coolen I, van Bergen Y, Laland KN. 2005. Trade-offs in the adaptive use of social and asocial learning. In Advances in the study of behavior, vol. 35 (eds Slater PJB, Snowdon CT, Brockmann HJ, Roper TJ, Naguib M), pp. 333–379. San Diego, CA: Elsevier Academic Press. [Google Scholar]
- 77.Derex M, Feron R, Godelle B, Raymond M. 2015. Social learning and the replication process: an experimental investigation. Proc. R. Soc. B 282, 20150719 ( 10.1098/rspb.2015.0719) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Enquist M, Ghirlanda S, Eriksson K. 2011. Modelling the evolution and diversity of cumulative culture. Phil. Trans. R. Soc. B 366, 412–423. ( 10.1098/rstb.2010.0132) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Kolodny O, Creanza N, Feldman MW. 2015. Evolution in leaps: the punctuated accumulation and loss of cultural innovations. Proc. Natl Acad. Sci. USA 112, E6762–E6769. ( 10.1073/pnas.1520492112) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Davis SJ, Vale GL, Schapiro SJ, Lambeth SP, Whiten A. 2016. Foundations of cumulative culture in apes: improved foraging efficiency through relinquishing and combining witnessed behaviours in chimpanzees (Pan troglodytes). Sci. Rep. 6, 35953 ( 10.1038/srep35953) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Kolodny O, Edelman S, Lotem A. 2015. Evolved to adapt: a computational approach to animal innovation and creativity. Curr. Zool. 61, 350 ( 10.1093/czoolo/61.2.350) [DOI] [Google Scholar]
- 82.Truskanov N, Prat Y. 2018. Cultural transmission in an ever-changing world: trial-and-error copying may be more robust than precise imitation Dryad Digital Repository. ( 10.10.5061/dryad.4m518) [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Citations
- Truskanov N, Prat Y. 2018. Cultural transmission in an ever-changing world: trial-and-error copying may be more robust than precise imitation Dryad Digital Repository. ( 10.10.5061/dryad.4m518) [DOI] [PMC free article] [PubMed]
Data Availability Statement
Data deposited in Dryad: http://dx.doi.org/10.5061/dryad.4m518 [82].