

Abstract
Key message
The new stem rust resistance gene Sr60 was fine-mapped to the distal region of chromosome arm 5AmS, and the TTKSK-effective gene SrTm5 could be a new allele of Sr22.
Abstract
The emergence and spread of new virulent races of the wheat stem rust pathogen (Puccinia graminis f. sp. tritici; Pgt), including the Ug99 race group, is a serious threat to global wheat production. In this study, we mapped and characterized two stem rust resistance genes from diploid wheat Triticum monococcum accession PI 306540. We mapped SrTm5, a previously postulated gene effective to Ug99, on chromosome arm 7AmL, completely linked to Sr22. SrTm5 displayed a different race specificity compared to Sr22 indicating that they are distinct. Sequencing of the Sr22 homolog in PI 306540 revealed a novel haplotype. Characterization of the segregating populations with Pgt race QFCSC revealed an additional resistance gene on chromosome arm 5AmS that was assigned the official name Sr60. This gene was also effective against races QTHJC and SCCSC but not against TTKSK (a Ug99 group race). Using two large mapping populations (4046 gametes), we mapped Sr60 within a 0.44 cM interval flanked by sequenced-based markers GH724575 and CJ942731. These two markers delimit a 54.6-kb region in Brachypodium distachyon chromosome 4 and a 430-kb region in the Chinese Spring reference genome. Both regions include a leucine-rich repeat protein kinase (LRRK123.1) that represents a potential candidate gene. Three CC–NBS–LRR genes were found in the colinear Brachypodium region but not in the wheat genome. We are currently developing a Bacterial Artificial Chromosome library of PI 306540 to determine which of these candidate genes are present in the T. monococcum genome and to complete the cloning of Sr60.
Electronic supplementary material
The online version of this article (doi:10.1007/s00122-017-3024-z) contains supplementary material, which is available to authorized users.
Introduction
The current global production of common wheat (Triticum aestivum L., 2n = 2x = 42) exceeds 700 million tons, providing roughly one-fifth of the calories and proteins consumed by the human population (FAOSTAT 2015). As our population continues to grow, further increases in wheat production will be required to meet food demands. One way to contribute to these increases is to minimize disease yield losses caused by pathogens.
Among the wheat pathogens, the rusts have been major threats since the early times of wheat cultivation. Among the three rust diseases, stem rust caused by the fungus Puccinia graminis f. sp. tritici (Pgt) can be the most destructive. In the United States, severe stem rust epidemics occurred between 1919 and 1954 causing significant yield losses (Roelfs 1978). Programs for the eradication of the alternate host barberry and for the deployment of resistant varieties contributed significantly to the reduction of wheat yield losses to this disease in the USA (Singh et al. 2015).
In 1998, a new Pgt race was detected in Uganda that was virulent to the widely deployed stem rust resistance genes Sr31 and Sr38. The corresponding isolate, designated as Ug99 (Pretorius et al. 2000), was typed as race TTKSK using the North American stem rust nomenclature system (Jin et al. 2008; Roelfs and Martens 1988). This race has spread to more than 13 countries, raising global concerns (Singh et al. 2015). During this geographic expansion, new virulences were detected in the Ug99 race group as the pathogen encountered and overcame resistance genes Sr24 (Jin et al. 2008), Sr36 (Jin et al. 2009), Sr9h (Pretorius et al. 2012; Rouse et al. 2014) and SrTmp (Newcomb et al. 2016; Patpour et al. 2016).
Few resistance genes effective against the Ug99 race group are present in common wheat varieties and most of the known Ug99-effective resistance genes are from the secondary and tertiary gene pools (Singh et al. 2015). The diploid wheat species Triticum monococcum (genome Am) is a tractable source of resistance genes. Chromosomes of T. monococcum recombine poorly with the chromosomes of hexaploid wheat in the presence of the Pairing homologous 1 (Ph1) gene. However, once this gene is deleted, the homoeologous chromosomes from these two species recombine normally (Dubcovsky et al. 1995). Five loci conferring resistance to the Ug99 race complex have been postulated in this diploid species (Rouse and Jin 2011a). Among them, Sr35 (Saintenac et al. 2013) and Sr22 (Steuernagel et al. 2016) have been cloned, whereas Sr21 (Chen et al. 2015) and SrTm4 (Briggs et al. 2015) have been genetically mapped.
Triticum monococcum accession PI 306540, collected in Romania, is a source of multiple stem rust resistance genes. Evaluation of progeny derived from the cross between PI 306540 and stem rust-susceptible accession PI 272557 indicated the presence of three genes effective to race TTKSK, two genes effective to race QFCSC, and one gene effective to race TTTTF (Rouse and Jin 2011a). One of the three race TTKSK-effective genes in PI 306540 was predicted to be Sr21, which does not confer resistance to races QFCSC or TTTTF (Chen et al. 2015). A second resistance gene in PI 306540, described as SrTm4, conferred resistance to races TTKSK, TTTTF and QFCSC (Briggs et al. 2015). We refer to the third gene effective to race TTKSK in PI 306540 as SrTm5. Our initial objective in the current study was to genetically map SrTm5. During the mapping of SrTm5, we discovered that this gene does not confer resistance to race QFCSC, and that PI 306540 possesses a fourth stem rust resistance gene. This gene was mapped in a chromosome region that did not include any previously named Sr genes and, therefore, was assigned the new official name Sr60.
Materials and methods
Plant materials and mapping populations
The Pgt-resistant T. monococcum subsp. monococcum cultivated accession PI 306540 was crossed with both cultivated accession PI 272557 and wild accession G3116 (Dubcovsky et al. 1996) to generate two mapping populations. PI 272557 does not carry any known Pgt-resistance genes (Rouse and Jin 2011b), so a population of 108 F2 plants from the cross PI 306540 × PI 272557 was used to separate SrTm5 and Sr60 from all other genes present in PI 306540 using molecular markers. From this population, we selected two lines carrying only SrTm5 (Pi23-15 and Pi54-3) and two lines carrying only Sr60 (Pi57-32 and Pi57-39), both in homozygous state.
Accession G3116 (= PI 427992) is from a different subspecies (T. monococcum subsp. aegilopoides) and is highly polymorphic compared to PI 306540. This characteristic is very useful to accelerate the development of molecular markers and the generation of high-density maps. However, since both G3116 and PI 306540 are homozygous for the presence of Sr21, this population can not be screened with race TTKSK. Instead, we used race QFCSC, which is known to be virulent to Sr21 (Rouse and Jin 2011a) and to SrTm5 but not to SrTm4 and Sr60 (this study). To map Sr60, we first separated it from SrTm4. We used molecular markers developed by Briggs et al. (2015) to identify 18 F2 plants homozygous for the absence of SrTm4 in a population of 98 F2 plants from the cross PI 306540 × G3116. Finally, we selected two families that showed clear phenotypic segregation and fit a 3:1 resistant:susceptible ratio in response to race QFCSC. After the initial mapping of Sr60, we used flanking markers to select plants heterozygous for the Sr60 region and used these plants to develop a second segregating population of 98 plants and a third population of 1811 plants. To generate a precise genetic map of Sr60, we evaluated the progenies from the plants showing informative recombination events between the markers flanking Sr60 with Pgt race QFCSC.
Evaluation for stem rust resistance
All the evaluations for Pgt resistance were performed at the USDA-Agricultural Research Service (USDA-ARS) Cereal Disease Laboratory following procedures described before (Rouse and Jin 2011a). For gene Sr60, P. graminis f. sp. tritici race QFCSC (isolate 06ND76C) was used to inoculate the parents and progeny from the two mapping populations. For gene SrTm5, plants from the mapping population were inoculated with race TTKSK (04KEN156/04). Infection types (ITs) were visually assessed 12–14 days after inoculation using a 0–4 scale (Stakman et al. 1962). The average sporulation areas for selected assays was quantified using the image analysis software ASSESS v.2 from the American Phytopathological Society. To determine the resistance profiles of SrTm5 and Sr60, lines carrying only one of these genes were inoculated with diverse Pgt races TTKSK (04KEN156/04), TTKST (06KEN19v3), MCCFC (59KS19), QFCSC (06ND76C), QTHJC (75ND717C), SCCSC (09ID73-2), TRTTF (06YEM34-1), TTTTF (01MN84A-1-2), and two isolates of TKTTF (13ETH18-1 and 13GER15-1).
Development of molecular markers
For the initial mapping of Sr60, we screened the parental lines PI 306540 and G3116 for polymorphisms using roughly 200 simple sequence repeat (SSR) markers (GrainGenes http://wheat.pw.usda.gov) and used the polymorphic markers to genotype the selected 18 F2 plants lacking SrTm4 (see “Plant materials and mapping populations”). In addition, we genotyped the same plants using the 90,000 SNP iSelect Illumina platform (Jordan et al. 2015). Once the Sr60 region was identified, additional markers were developed from genes located in the colinear regions of the Brachypodium dystachium and the T. aestivum genomes (IWGSC CS WGA v1.0 and Zavitan WEWSeq v.1.0). Primers were designed to amplify intronic regions, and the detected SNPs were used to develop cleaved amplified polymorphic sequence (CAPS) markers and derived cleaved amplified polymorphic sequence (dCAPS) markers (Neff et al. 1998).
The population segregating for SrTm5 was also genotyped with the 90K SNP iSelect Illumina platform. Once we found that SrTm5 was linked to Sr22, we Sanger-sequenced the Sr22 homolog in PI 306540 and compared its sequence with the different haplotypes of Sr22 (Steuernagel et al. 2016).
Construction of genetic linkage maps and comparisons with genomic sequences
Genetic linkage maps were constructed using MAPMAKER EXP3.0 (Lincoln et al. 1992) using the Kosambi function and a LOD threshold of 3.0. The software MAPDRAW V2.1 was used to draw the linkage maps (Liu and Meng 2003).
Sequences flanking the SNP markers were used to BLAST the Brachypodium distachyon genome (JGI Phytozome v11.0, https://phytozome.jgi.doe.gov/). Coordinates in the Chinese Spring wheat genome were based on the IWGSC CS WGA v1.0 assembly (Zimin et al. 2017) and those in tetraploid wheat on the NRGene assembly of the T. turgidum subsp. dicoccoides accession Zavitan (Avni et al. 2017).
Results
Mapping new stem rust resistance gene Sr60 conferring resistance to race QFCSC on chromosome arm 5AmS
The initial mapping of Sr60 was performed with the 18 F2 plants selected for the absence of SrTm4 and their derived F2:3 families from the cross PI 306540 × G3116. Approximately 25 F3 plants from each family were phenotyped with Pgt race QFCSC (virulent to Sr21 and SrTm5 but avirulent to the new gene). We identified three families that were homozygous resistant, 12 that were segregating, and three that were homozygous susceptible (Fig. 1). These numbers were consistent with segregation of a single resistance gene (χ 2 = 2, P = 0.368).
Fig. 1.

Infection types of selected PI 306540 × G3116 F3 families lacking SrTm4 in response to Puccinia graminis f. sp. tritici race QFCSC (isolate 06ND76C) including families that possess Sr60 (1–4), and families that lack Sr60 (5–8). Numbers below leaves are average pustule size. Asterisks indicate significant differences between plants with Sr60 and plants without this gene (***P < 0.001)
Genotyping these 18 F2 plants with all the 90K SNP markers and polymorphic SSR markers revealed one SNP locus (IWB40642) completely linked to this gene (LOD score > 7) and another one (IWB48091) 5.8 cM distal from this gene (LOD score > 4). These two loci were previously mapped on the distal region of chromosome arm 5AS (https://triticeaetoolbox.org/wheat/). Another four SSR markers were loosely linked among each other and to Sr60 through gwm154, which together with gwm415 was located in the short arm (Fig. 2a). This last SSR marker was linked to gwm156, which was linked to gwm186, both on the long arm of chromosome 5Am (Fig. 2a). These data indicated that Sr60 was located on the distal region of the short arm of chromosome 5Am. Because no previously named stem rust resistance gene was mapped on chromosomes 5AS or 5AmS, this gene was assigned the new official name Sr60.
Fig. 2.
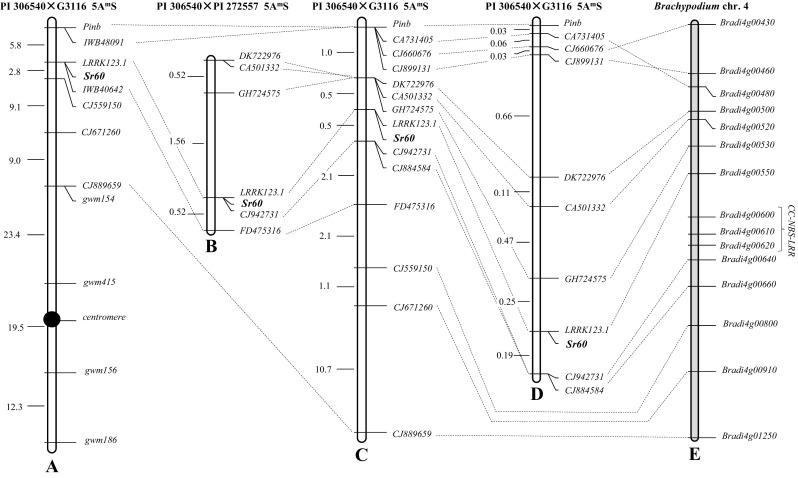
Genetic maps of Sr60 on chromosome arm 5AmS. a, c, d Maps were derived from the cross PI 306540 × G3116 whereas b was derived from the cross PI 306540 × PI 272557. All phenotypes were collected from inoculations with race QFCSC. e Physical map of Brachypodium chromosome 4. a Initial map based on 18 F2:3 families lacking SrTm4. b Map based on 96 F3:2 plants lacking SrTm4 and Sr21. c Map based on 98 F3:2 plants lacking SrTm4. d High-density map based on 1811 F3:2 plants lacking SrTm4. e Colinear region in the sequenced Brachypodium chromosome 4
Using the sequence of SNP marker IWB48091, we selected T. aestivum contig IWGSC_chr5AS_ab_k95_contigs_longerthan_200_1551239 (https://wheat-urgi.versailles.inra.fr/Seq-Repository/BLAST). Within this contig we identified wheat gene TraesCS5A01G003800 (marker CJ660676) that is orthologous to B. distachyon gene Bradi4g00430. We selected close B. distachyon genes (Bradi4g00550, Bradi4g00800, Bradi4g00910 and Bradi4g01250) and developed markers for the corresponding wheat orthologs (LRRK123.1, CJ559150, CJ671260, and CJ889659; Table S1) that were added to the genetic map (Fig. 2a).
Construction of a high-density genetic map of Sr60
The high-density map of Sr60 was constructed in two phases. In phase one, we used a population of 98 F3:2 plants derived from two F3 families heterozygous for flanking markers CJ660676 and CJ889659. Characterization of these plants with the two flanking markers and with Pgt race QFCSC confirmed that Sr60 was located 1.5 cM proximal to CJ660676 and 16.5 cM distal to CJ889659. To define better the position of Sr60, we developed and mapped nine additional molecular markers. First we developed a CAPS marker for the Puroindoline-b (Pinb) gene responsible for grain softness (Tranquilli et al. 1999), which is known to be located in the very distal region of the short arm of homoeologous group 5 chromosomes. This marker was completely linked to CJ660676 (IWB48091), confirming its distal location. We also developed marker FD475316 from a wheat contig selected with the sequence of IWB40642, and mapped it 2.6 cM proximal to Sr60.
We then selected seven more genes from the Sr60 colinear region of B. distachyon chromosome 4 (Fig. 2e), identified the wheat orthologs and developed seven new molecular markers (CA731405, CJ899131, DK722976, CA501332, GH724575, CJ942731 and CJ884584; Table S1) that were added to the genetic map (Fig. 2c). Based on this population, Sr60 was completely linked to marker LRRK123.1, and mapped within a 1 cM interval flanked by markers GH724575 and CJ942731 (Fig. 2c).
In the second phase of the high-density map, we screened 1811 plants (from the same F3 families) segregating for flanking markers Pinb and CJ884584. We found 65 plants with recombination events within this region (1.8 cM) that were used to precisely map all the markers described above (Fig. 2d). In the final high-density map, Sr60 remained completely linked to LRRK123.1 and was flanked on the distal side by GH724575 (0.25 cM) and on the proximal side by CJ942731 (0.19 cM).
Candidate genes for Sr60 within the colinear regions of B. distachyon and wheat genomes
The high-density map of Sr60 showed good colinearity with the genomic sequences of B. distachyon (Fig. 2e), and with the recent NRGene assemblies of tetraploid (T. turgidum subsp. dicoccoides var. Zavitan) and hexaploid wheat (IWGSC CS WGA v1.0). The 0.44 cM candidate region between GH724575 and CJ942731 in the genetic map of T. monococcum corresponded to a 54.6-kb region in B. distachyon (Bradi4g00530–Bradi4g00640) and 425- and 430-kb regions in the A genomes of tetraploid and hexaploid wheat, respectively.
There were several genes present in the B. distachyon genome that were not detected in the genomes of hexaploid or tetraploid wheat. Of particular interest for this project was the presence of three CC–NBS–LRR genes in the Sr60 colinear region of B. distachyon (Bradi4g00600, Bradi4g00610, and Bradi4g00620), since this gene class is frequently associated with pathogen resistance. Although, no CC–NBS–LRR genes were detected in the colinear regions of the tetraploid and hexaploid wheat genomes (Table S2), we cannot currently rule out their presence in T. monococcum PI 306540.
An interesting candidate gene is LRRK123.1 (Shumayla et al. 2016), which is located in the middle of the candidate region in the T. monococcum genetic map and completely linked to Sr60. An ortholog of this gene was found in the colinear position of B. distachyon (Bradi4g00550), which encodes a protein 91% similar to the wheat gene. The candidate wheat gene (TraesCS5A01G005300) is annotated in the genome of Chinese Spring (IWGSC CS WGA v1.0) as a leucine-rich repeat receptor like kinase (LRRK). We sequenced the complete LRRK123.1 gene and found only two amino acid differences between the resistant line PI 306540 and the susceptible line PI 272557 at positions 395 and 484 (T395M and T484A; BLOSUM62 score = − 1 and 0).
Mapping Sr60 in the PI 306540 × PI 272557 population and selecting plants carrying only Sr60
We also mapped Sr60 in a second population (PI 306540 × PI 272557) to identify plants carrying only Sr60 and to determine its race specificity. We first characterized a population of 108 F2 plants from this cross with molecular markers for Sr21 and SrTm4 (Briggs et al. 2015; Chen et al. 2015). We identified six plants homozygous for the absence of these two genes and performed progeny tests (approximately 25 F3 plants per family) using Pgt race QFCSC. Four families segregated for resistance to race QFCSC, one was homozygous susceptible and one, designated as Pi14, was homozygous resistant.
One of the Sr60 segregating families, designated as Pi57, was selected to generate a new mapping population of 96 F3:2 plants that was tested for response to QFCSC. Six of the markers mapped in the previous population (Fig. 2c, d) were polymorphic between PI 306540 and PI 272557 and were used to generate an additional genetic map for Sr60 (Fig. 2b). In this population, Sr60 was mapped 1.6 cM proximal to GH724575 and 0.5 cM distal to FD475316, in the same interval as in the PI 306540 × G3116 population. Using these flanking markers, we confirmed that family Pi14, which was homozygous resistant to QFCSC, was also homozygous for the PI 306540 allele for the two markers flanking Sr60.
When we tested the progeny of Pi14 with race TTKSK, we were surprised to find a clear 3:1 segregation of the resistance response (18 resistant plants with infection type ‘;1’ and 5 susceptible plants with infection type ‘3 +’; χ 2 = 0.13, P = 0.72). Similar results were observed for Pi14 progeny in response to races TTKST and MCCFC. Since SrTm5 was originally postulated based on its resistance to race TTKSK, the observed segregation in the Pi14 progeny indicated that our initial assumption that Sr60 and SrTm5 were the same gene was incorrect.
Genetic map of SrTm5 resistance using race TTKSK (Ug99)
To map SrTm5, we inoculated a population of 63 plants derived from line Pi14 with race TTKSK and identified 49 resistant and 14 susceptible plants. These values fit the 3:1 segregation ratio expected for a single dominant gene (χ 2 = 0.259, P = 0.61). Genotyping of these plants with the 90K SNP iSelect Illumina assay revealed six SNPs (IWB25012, IWB44281, IWB40527, IWB6942, IWB23038 and IWB31237) significantly linked to SrTm5 (Table 1 and Fig. 3a). Based on the linkage results, SrTm5 was mapped to the long arm of chromosome 7Am completely linked to loci IWB25012, IWB44281 and IWB40527. Using the sequences of the linked markers, we determined that their locations in the T. aestivum reference genome of Chinese Spring (IWGSC CS WGA v1.0, 689,920,024–690,055,086 bp) was coincident with the location of the previously cloned gene Sr22 (689,920,136–689,925,980 bp).
Table 1.
SNPs linked with SrTm5 and their previously mapped positions in hexaploid wheat
| SNP ID | SNP name | Chr | Re-scaled distance cMa |
|---|---|---|---|
| IWB25012 | Excalibur_c30730_276 | 7A | 175.67 |
| IWB44281 | Kukri_c34147_152 | 7A | 177.28 |
| IWB40527 | Kukri_c11141_203 | 7B | 113.87 |
| IWB6942 | BS00022169_51 | 7A | 178.42 |
| IWB23038 | Excalibur_c1791_819 | 7A | 178.42 |
| IWB31237 | Excalibur_rep_c74778_252 | 7B | 118.67 |
aDistances for the markers are from https://triticeaetoolbox.org/wheat/
Fig. 3.
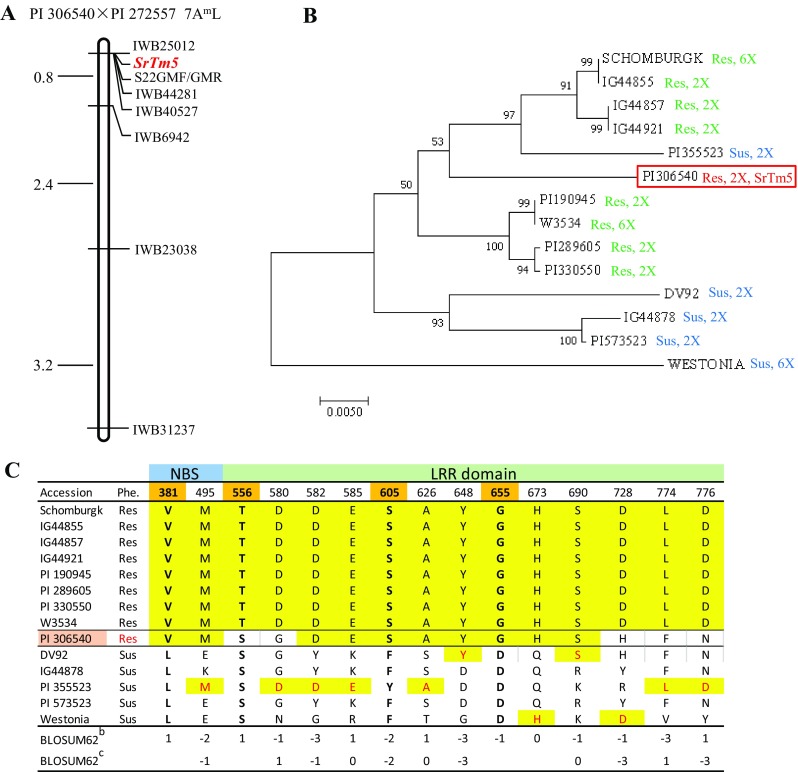
Genetic map of SrTm5 and protein sequence relationships with previously reported Sr22 haplotypes. a Genetic map of chromosome 7AmL. S22GMF/GMR is the Sr22 gene-specific PCR marker and the others are SNP markers. b Neighbor joining phylogenetic tree of protein sequences of the NLR gene from PI 306540 linked to Sr22 and previously characterized Sr22 haplotypes (Steuernagel et al. 2016). Res resistant haplotypes, Sus susceptible haplotypes, 2x T. monococcum, 6x T. aestivum. c Comparison of the PI 306540 Sr22 protein haplotype with protein polymorphisms that discriminate between Sr22 resistant and susceptible haplotypes from Steuernagel et al. (2016). Positions 381, 556, 605 and 655 show a perfect discrimination between haplotypes, whereas other polymorphism included in this table have a maximum of one exception. BLOSUM62 scores are included below each amino acid position (b = score for the most frequent polymorphism and c = score for the second most frequent polymorphism)
To confirm the linkage between SrTm5 and Sr22, we mapped the Sr22 diagnostic marker S22GMF/GMR (Steuernagel et al. 2016) in the 63 F2 plants originally used to map SrTm5 and in another 110 plants also derived from line Pi14. This marker, which amplified a 176 bp product in the resistant parent PI 306540 and no product in the susceptible parents G3116 or PI 272557, was mapped completely linked to SrTm5 in the 173 F2 plants. This result suggests that SrTm5 is either allelic to Sr22 or is less than 1.7 cM from Sr22 (at a 95% confidences interval) based on formulas described by Hanson (1959). We amplified the coding region of Sr22 from PI 306540 using primers reported by Steuernagel et al. (2016). The amplified sequence (GenBank MG018615) was very similar but not identical to the six reported resistant haplotypes of Sr22 (from 98.0 to 98.6% at the cDNA level, Table S3). A phylogenetic analysis showed that the SrTm5 predicted protein is in the middle of a cluster including Sr22 predicted proteins from different haplotypes (Fig. 3b). Only four amino acid positions discriminate perfectly the Sr22 resistant and susceptible protein haplotypes included in Fig. 3b (Fig. 3c). Among these four amino acids, PI 306540 shares three with the Sr22 resistant haplotypes and one with the susceptible haplotypes. The amino acid shared with the susceptible haplotypes (PI 306540 position 556) has a positive BLOSUM62 score of 1, which indicates a non-disruptive amino acid change. By contrast, two of the amino acids shared with the resistant haplotypes (positions 605 and 655 both in the LRR domain) have negative BLOSUM62 scores, which are indicative of changes that are more disruptive to structure and function. For the other 11 amino acid positions that show a maximum of one exception to a perfect separation between R and S haplotypes (Fig. 3c), PI 306540 has the same amino acids as the resistant haplotypes in seven of them. These results suggest that the Sr22 protein sequence present in PI 306540 is more related to the Sr22 protein in resistant haplotypes than in the susceptible haplotypes.
Resistance profiles of Sr60 and SrTm5
Using the markers linked with Sr60 and SrTm5, we identified lines carrying only Sr60 or only SrTm5 from population PI 306540 × PI 272557. Lines Pi23-15 and Pi54-3 homozygous for SrTm5, and lines Pi57-32 and Pi57-39 homozygous for Sr60 were inoculated with Pgt races TTKSK, TTKST (similar to TTKSK with additional virulence to Sr24), MCCFC, QFCSC, QTHJC, SCCSC, TKTTF (two isolates), TRTTF and TTTTF.
Plants carrying only Sr60 displayed resistant infection types (IT = 2–22 +) to Pgt races QFCSC (Fig. 1), QTHJC, and SCCSC and susceptible infection types (IT = 3 +) to the other races (Table 2). We quantified the average sporulation areas using the image analysis software ASSESS v.2 and observed that the average pustule size in the plants with Sr60 was significantly smaller than in the plants without this gene (P < 0.001; Fig. 1).
Table 2.
Resistance profiles of SrTm5 and Sr60 to multiple Puccinia graminis f. sp. tritici races
| Selected line | Resistance gene | Races | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TTKSK | TTKST | MCCFC | QFCSC | QTHJC | SCCSC | TKTTF | TRTTF | TTTTF | ||
| Pi23-15 | Only SrTm5 | ;1 | ; | 1; | 3 + | 3 + | 3 + | 3 + | 3 + | 3 + |
| Pi54-3 | Only SrTm5 | ;1 | ;1 | ;1 | 3 + | 3 + | 3 + | 3 + | 3 + | 3 + |
| Pi57-32 | Only Sr60 | 3 + | 3 + | 33 + | 2 | 2 | 22 + | 3 + | 3 + | 3 + |
| Pi57-39 | Only Sr60 | 3 + | 3 + | 33 + | 2 | 2 | 22 + | 3 + | 3 + | 3 + |
| Pi57-57 | None | 3 + | 3 + | 3 + | 3 + | 3 + | 3 + | 3 + | 3 + | 3 + |
| Pi57-31 | None | 3 + | 3 + | 33 + | 33 + | 3 + | 3 + | 3 + | 3 + | 33 + |
Lines were selected from the population PI 306540 × PI 272557. The two isolates of TKTTF yielded identical results
Plants carrying only SrTm5 showed strong resistance to Pgt races TTKSK, TTKST and MCCFC (IT = ; to ;1) but were susceptible to the other tested races (IT = 3 +) (Fig. 4, Table 2). We also quantified the average sporulation areas in this experiment, and found the average pustule size was significantly smaller in the plants with SrTm5 than in those without the gene (P < 0.001, Fig. 4). Susceptibility of plants with only SrTm5 to races TTTTF, QFCSC, and QTHJC clearly differentiated this gene from Sr22 that previously displayed resistance to these same races (Olson et al. 2010).
Fig. 4.

Infection types of PI 306540 × PI 272557 derivatives lines (1) Pi23-15 (SrTm5), (2) Pi54-3 (SrTm5), (3) Pi57-32 (Sr60), (4) Pi57-39 (Sr60), (5) Pi57-57 (no resistance gene) and (6) Pi57-31 (no resistance gene) in response to Puccinia graminis f. sp. tritici races TTKST and TTKSK (Ug99). R resistance, S susceptible. Numbers below leaves present the average pustule size. Asterisks indicate the existence of significant differences between plants with either SrTm5 or Sr60 compared to plants without either gene (***P < 0.001)
Discussion
Identification of new stem rust resistance gene Sr60
A previous study postulated that the T. monococcum accession PI 306540, which was collected in Romania, possessed three stem rust resistance genes (Sr21, SrTm4, and SrTm5) effective against Pgt race TTKSK (Rouse and Jin 2011a). In this study, we identified a fourth stem rust resistance gene in PI 306540 that was effective against Pgt race QFCSC but not against TTKSK (Table 2). This fourth gene was mapped on the distal region of chromosome arm 5AmS, where no previously named stem rust resistance gene has been mapped. Based on this unique chromosome location and characteristic resistant profile, this gene from PI 306540 was assigned the official designation Sr60.
Using our high-density genetic map, and the recently released genomes of tetraploid and hexaploid wheat (Avni et al. 2017; Zimin et al. 2017), we were able to delimit the Sr60 candidate region to a 430-kb region in hexaploid wheat Chinese Spring and a 425-kb region in tetraploid wheat Zavitan. Within these regions we identified a leucine-rich repeat receptor like kinase LRRK123.1, which was completely linked to Sr60. Members of this gene family have been associated with disease resistance in other organisms (Chinchilla et al. 2007; Halter et al. 2014; Roux et al. 2011; Sun et al. 2004). We detected two amino acid changes in the kinase domain of LRRK123.1 between the resistant line PI 306540 and the susceptible line PI 272557, but we currently do not know if these changes affect the function of this gene. Complementation and mutant experiments will be required to demonstrate if this gene is Sr60.
Another class of genes frequently associated with race-specific resistance mechanisms includes the NLR genes. Many of these genes recognize effectors delivered by the pathogens or the modifications they induce in targeted host proteins and, therefore, mutations or deletions of these effectors result in susceptibility and in differences in virulence among races. Although the Sr60 candidate regions in tetraploid and hexaploid wheat do not include any NLR genes, the presence of three NLR genes in the 54.6-kb colinear region in B. distachyon (Bradi4g00600, Bradi4g00610, and Bradi4g00620, Table S2) raises the possibility of an ancestral NLR cluster in this region that was deleted in tetraploid and hexaploid wheat. If this hypothesis is correct, we cannot rule out the possibility that some NLR genes of this cluster are still present in T. monococcum PI 306540. To address this possibility, we have initiated the development of a Bacterial Artificial Chromosome (BAC) library from PI 306540.
Sr60 conferred intermediate levels of resistance to Pgt races QFCSC, QTHJC and SCCSC, but was susceptible to TTKSK and TTKST of the Ug99 group. This resistance profile complements well the resistance profile of SrTm5 (Table 2), providing a potential explanation for the simultaneous presence of these two genes in PI 306540. Sr60 was not effective against emerging virulent Pgt races such as the Ug99 race group, race TRTTF that was reported in Yemen and Ethiopia (Olivera et al. 2012), and isolates of race TKTTF that caused epidemics of stem rust in Ethiopia and Germany in 2013 (Olivera et al. 2015, 2017). This suggests that Sr60 has limited potential for use in agriculture in regions where these races are present. Sr60 may be useful in combination with other Sr genes if Pgt races emerge that are virulent on the resistance genes deployed in wheat varieties, but avirulent to Sr60. The identification and deployment of multiple resistance genes with an increased diversity of resistance specificities has the potential to extend the durability of wheat resistance to stem rust.
SrTm5 is possibly a new allele of Sr22
Two lines of evidence suggest that SrTm5 is possibly a new allele of Sr22. First, SrTm5 was completely linked to the diagnostic marker for Sr22 (Fig. 3a). However, given the size of the mapping population we can only say that the two genes are less than 1.7 cM apart with a 95% confidence. Second, the sequence of the only NLR gene linked to SrTm5 is very similar (98–99% identical in the coding regions) to Sr22, and the encoded protein is within a cluster including mostly Sr22 resistant alleles (Fig. 3b). In addition, the Sr22 protein haplotype of PI 306540 carries the same amino acids as the resistant alleles at three of the four positions that discriminate the resistant and susceptible Sr22 protein haplotypes (Fig. 3c). Although it is difficult to rule out the possibility that SrTm5 originated from a linked gene paralogous to Sr22, the previous results suggest that the PI 306540 Sr22-like gene may be the one conferring resistance to TTKSK, TTKST and MCCFC.
SrTm5 and Sr22 both confer resistance to Pgt races TTKSK (Ug99), TTKST and MCCFC, but differ in that SrTm5 is susceptible to races QFCSC, TRTTF, and TTTTF (Table 3). We hypothesize that the 23 amino acid differences detected between SrTm5 and the six Sr22 resistant haplotypes (Table S3), which are all located in the LRR region, can affect the ability of the SrTm5 protein to recognize the QFCSC, TRTTF, and TTTTF effectors or the changes produced by these effectors on their target proteins.
Table 3.
Resistance profiles of SrTm5 and Sr22 to multiple Puccinia graminis f. sp. tritici races
| Pgt race | SrTm5, 2x Pi23-15/Pi54-3 | Sr22TB, 6x cv. Schormburgka | Sr22, 2x PI 190945a | No Sr gene, 2x PI 272557a |
|---|---|---|---|---|
| TTKSK | ;1 | 2 − | 1 | 4 |
| TTKST | ; | Not available | Not available | Not available |
| MCCFC | 1; | 2 − | ;1 | 4 |
| QFCSC | 3 + | ;2 − | ;1 − | 4 |
| TRTTF | 3 + | 2 −; | 1; | 3 + |
| TTTTF | 3 + | 2 | 2 − | 4 |
Infection types for SrTm5 are from selected lines Pi23-15 and Pi54-3, whereas Sr22 infection types are from T. aestivum cultivar Schormburgk and T. monococcum ssp. monococcum accession PI 190945. PI 272557 was used as susceptible check. PI 190945 carries Sr22 and does not carry Sr60 or Sr21
2x diploid wheat T. monococcum, 6x hexaploid wheat T. aestivum
aBased on Steuernagel et al. (2016) and Rouse and Jin (2011a). Reductions in the resistance levels have been observed for other resistance genes when transferred from a lower to a higher ploidy level (McIntosh et al. 2011)
We plan to use a PI 306540 BAC library to determine if T. monococcum has close Sr22 paralogs or pseudogenes in the Sr22 region. Complementation experiments with the PI 306540 haplotype of Sr22 (possibly SrTm5) will be required to demonstrate that this gene is the one that confers resistance to races TTKSK, TTKST and MCCFC but not to races QFCSC, TRTTF and TTTTF.
SrTm5 has been postulated to be present also in T. monococcum accession PI 277131-2 (Rouse and Jin 2011a). Indeed, we identified a gene in PI 277131-2 that was 100% identical to the SrTm5 candidate gene in PI 306540.
Conclusions
In summary, we have identified a stem rust resistance locus linked to Sr22 that has a different resistance profile than the previously characterized Sr22 gene. Although it is likely that this gene represents a novel allele of Sr22, we will keep the temporary designation SrTm5 until allelism with Sr22 is more conclusively demonstrated. If SrTm5 is a new allele of Sr22, the new haplotype will be a useful tool to understand the specificity of Sr22 to different effectors. In addition, we identified Sr60, a novel race-specific stem rust resistance gene on chromosome arm 5AmS. With this study, we complete the genetic characterization of the different stem rust resistance genes postulated so far in diploid wheat T. monococcum. This information has the potential to accelerate the deployment of these genes in the polyploid wheat species and to expand our understanding of the role of different resistance genes combinations in the adaptation of diploid wheat to this devastating pathogen.
Author contribution statement
ShisC and YG evaluated the populations, analyzed the data and wrote the first draft. FD contributed to the initial stages of mapping. ShiaC performed the Illumina genotyping and analyzed the data. JB and MR designed and performed the phenotyping experiments of the project. WZ contributed crosses and sequence analyses and supervised ShisC. JD initiated the project, contributed to the genetic map and statistical analyses and generated the final version of the manuscript. All authors revised the manuscript and provided suggestions.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Acknowledgements
Work at JD laboratory was supported by the Howard Hughes Medical Institute and by the Agriculture and Food Research Initiative Competitive Grant 2017-67007-25939 from the USDA National Institute of Food and Agriculture (NIFA). Work at MNR laboratory was supported by USDA-ARS appropriated Project 5062-21220-021-00-D, the USDA-ARS National Plant Disease Recovery System, and NRI Competitive Grant 2017-67007-25939 from the USDA National Institute of Food and Agriculture (NIFA).
Conflict of interest
The authors declare that there are no conflicts of interest.
Human or animal statement
This study does not include human or animal subjects.
References
- Avni R, Nave M, Barad O, Baruch K, Twardziok SO, Gundlach H, Hale I, Mascher M, Spannagl M, Wiebe K. Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science. 2017;357:93–97. doi: 10.1126/science.aan0032. [DOI] [PubMed] [Google Scholar]
- Briggs J, Chen SS, Zhang WJ, Nelson S, Dubcovsky J, Rouse MN. Mapping of SrTm4, a recessive stem rust resistance gene from diploid wheat effective to Ug99. Phytopathology. 2015;105:1347–1354. doi: 10.1094/PHYTO-12-14-0382-R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen S, Rouse MN, Zhang W, Jin Y, Akhunov E, Wei Y, Dubcovsky J. Fine mapping and characterization of Sr21, a temperature-sensitive diploid wheat resistance gene effective against the Puccinia graminis f. sp. tritici Ug99 race group. Theor Appl Genet. 2015;128:645–656. doi: 10.1007/s00122-015-2460-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chinchilla D, Zipfel C, Robatzek S, Kemmerling B, Nürnberger T, Jones JD, Felix G, Boller T. A flagellin-induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature. 2007;448:497. doi: 10.1038/nature05999. [DOI] [PubMed] [Google Scholar]
- Dubcovsky J, Luo M, Dvorak J. Differentiation between homoeologous chromosomes 1A of wheat and 1Am of Triticum monococcum and its recognition by the wheat Ph1 locus. Proc Natl Acad Sci USA. 1995;92:6645–6649. doi: 10.1073/pnas.92.14.6645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dubcovsky J, Luo M-C, Zhong G-Y, Bransteiter R, Desai A, Kilian A, Kleinhofs A, Dvorak J. Genetic map of diploid wheat, Triticum monococcum L., and its comparison with maps of Hordeum vulgare L. Genetics. 1996;143:983–999. doi: 10.1093/genetics/143.2.983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FAOSTAT (2015) Food supply—crops primary equivalent. Food and Agriculture Organization of the United Nations. http://faostat3.fao.org/download/FB/CC/E. Accessed 20 Nov 2017
- Halter T, Imkampe J, Mazzotta S, Wierzba M, Postel S, Bücherl C, Kiefer C, Stahl M, Chinchilla D, Wang X. The leucine-rich repeat receptor kinase BIR2 is a negative regulator of BAK1 in plant immunity. Curr Biol. 2014;24:134–143. doi: 10.1016/j.cub.2013.11.047. [DOI] [PubMed] [Google Scholar]
- Hanson W. Minimum family sizes for the planning of genetic experiments. Agron J. 1959;51:711–715. doi: 10.2134/agronj1959.00021962005100120005x. [DOI] [Google Scholar]
- Jin Y, Szabo LJ, Pretorius ZA, Singh RP, Ward R, Fetch T. Detection of virulence to resistance gene Sr24 within race TTKS of Puccinia graminis f. sp tritici. Plant Dis. 2008;92:923–926. doi: 10.1094/PDIS-92-6-0923. [DOI] [PubMed] [Google Scholar]
- Jin Y, Szabo LJ, Rouse MN, Fetch T, Pretorius ZA, Wanyera R, Njau P. Detection of virulence to resistance gene Sr36 within the TTKS race lineage of Puccinia graminis f. sp tritici. Plant Dis. 2009;93:367–370. doi: 10.1094/PDIS-93-4-0367. [DOI] [PubMed] [Google Scholar]
- Jordan KW, Wang S, Lun Y, Gardiner L, MacLachlan R, Hucl P, Wiebe K, Wong D, Forrest KL, Sharpe AG, Sidebottom C, Hall N, Toomajian C, Close T, Dubcovsky J, Akhunova A, Talbert L, Bansal UK, Bariana HS, Hayden MJ, Pozniak C, Jeddeloh JA, Hall A, Akhunov E. A haplotype map of allohexaploid wheat reveals distinct patterns of selection on homoeologous genomes. Genome Biol. 2015;16:48. doi: 10.1186/s13059-015-0606-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lincoln S, Daly M, Lander E (1992) Constructing genetic maps with MAPMAKER/EXP 3.0. In: Whitehead Institute technical report, 3rd edn
- Liu RH, Meng JL. MapDraw: a Microsoft excel macro for drawing genetic linkage maps based on given genetic linkage data. Yi Chuan. 2003;25:317–321. [PubMed] [Google Scholar]
- McIntosh RA, Zhang P, Cowger C, Parks R, Lagudah ES, Hoxha S. Rye-derived powdery mildew resistance gene Pm8 in wheat is suppressed by the Pm3 locus. Theor Appl Genet. 2011;123:359–367. doi: 10.1007/s00122-011-1589-5. [DOI] [PubMed] [Google Scholar]
- Neff MM, Neff JD, Chory J, Pepper AE. dCAPS, a simple technique for the genetic analysis of single nucleotide polymorphisms: experimental applications in Arabidopsis thaliana genetics. Plant J. 1998;14:387–392. doi: 10.1046/j.1365-313X.1998.00124.x. [DOI] [PubMed] [Google Scholar]
- Newcomb M, Olivera PD, Rouse MN, Szabo LJ, Johnson J, Gale S, Luster DG, Wanyera R, Macharia G, Bhavani S. Kenyan isolates of Puccinia graminis f. sp. tritici from 2008 to 2014: virulence to SrTmp in the Ug99 race group and implications for breeding programs. Phytopathology. 2016;106:729–736. doi: 10.1094/PHYTO-12-15-0337-R. [DOI] [PubMed] [Google Scholar]
- Olivera P, Jin Y, Rouse M, Badebo A, Fetch T, Jr, Singh R, Yahyaoui A. Races of Puccinia graminis f. sp. tritici with combined virulence to Sr13 and Sr9e in a field stem rust screening nursery in Ethiopia. Plant Dis. 2012;96:623–628. doi: 10.1094/PDIS-09-11-0793. [DOI] [PubMed] [Google Scholar]
- Olivera P, Newcomb M, Szabo LJ, Rouse M, Johnson J, Gale S, Luster DG, Hodson D, Cox JA, Burgin L. Phenotypic and genotypic characterization of race TKTTF of Puccinia graminis f. sp. tritici that caused a wheat stem rust epidemic in southern Ethiopia in 2013–14. Phytopathology. 2015;105:917–928. doi: 10.1094/PHYTO-11-14-0302-FI. [DOI] [PubMed] [Google Scholar]
- Olivera P, Newcomb M, Flath K, Sommerfeldt-Impe N, Szabo L, Carter M, Luster D, Jin Y. Characterization of Puccinia graminis f. sp. tritici isolates derived from an unusual wheat stem rust outbreak in Germany in 2013. Plant Pathol. 2017;66:1258–1266. doi: 10.1111/ppa.12674. [DOI] [Google Scholar]
- Olson EL, Brown-Guedira G, Marshall D, Stack E, Bowden RL, Jin Y, Rouse M, Pumphrey MO. Development of wheat lines having a small introgressed segment carrying stem rust resistance gene Sr22. Crop Sci. 2010;50:1823–1830. doi: 10.2135/cropsci2009.11.0652. [DOI] [Google Scholar]
- Patpour M, Hovmøller MS, Shahin AA, Newcomb M, Olivera P, Jin Y, Luster D, Hodson D, Nazari K, Azab M. First report of the Ug99 race group of wheat stem rust, Puccinia graminis f. sp. tritici, in Egypt in 2014. Plant Dis. 2016;100:863. doi: 10.1094/PDIS-08-15-0938-PDN. [DOI] [Google Scholar]
- Pretorius ZA, Singh RP, Wagoire WW, Payne TS. Detection of virulence to wheat stem rust resistance gene Sr31 in Puccinia graminis f. sp. tritici in Uganda. Plant Dis. 2000;84:203. doi: 10.1094/PDIS.2000.84.2.203B. [DOI] [PubMed] [Google Scholar]
- Pretorius Z, Szabo L, Boshoff W, Herselman L, Visser B. First report of a new TTKSF race of wheat stem rust (Puccinia graminis f. sp. tritici) in South Africa and Zimbabwe. Plant Dis. 2012;96:590. doi: 10.1094/PDIS-12-11-1027-PDN. [DOI] [PubMed] [Google Scholar]
- Roelfs AP. Estimated losses caused by rust in small grain cereals in the United States—1918–1976. Washington, D.C.: United States Department of Agriculture; 1978. [Google Scholar]
- Roelfs A, Martens J. An international system of nomenclature for Puccinia graminis f. sp. tritici. Phytopathology. 1988;78:526–533. doi: 10.1094/Phyto-78-526. [DOI] [Google Scholar]
- Rouse M, Jin Y. Genetics of resistance to race TTKSK of Puccinia graminis f. sp. tritici in Triticum monococcum. Phytopathology. 2011;101:1418–1423. doi: 10.1094/PHYTO-05-11-0133. [DOI] [PubMed] [Google Scholar]
- Rouse M, Jin Y. Stem rust resistance in A-genome diploid relatives of wheat. Plant Dis. 2011;95:941–944. doi: 10.1094/PDIS-04-10-0260. [DOI] [PubMed] [Google Scholar]
- Rouse MN, Nirmala J, Jin Y, Chao SM, Fetch TG, Pretorius ZA, Hiebert CW. Characterization of Sr9h, a wheat stem rust resistance allele effective to Ug99. Theor Appl Genet. 2014;127:1681–1688. doi: 10.1007/s00122-014-2330-y. [DOI] [PubMed] [Google Scholar]
- Roux M, Schwessinger B, Albrecht C, Chinchilla D, Jones A, Holton N, Malinovsky FG, Tör M, de Vries S, Zipfel C. The Arabidopsis leucine-rich repeat receptor-like kinases BAK1/SERK3 and BKK1/SERK4 are required for innate immunity to hemibiotrophic and biotrophic pathogens. Plant Cell. 2011;23:2440–2455. doi: 10.1105/tpc.111.084301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saintenac C, Zhang W, Salcedo A, Rousse M, Trick H, Akhunov E, Dubcovsky J. Identification of wheat gene Sr35 that confers resistance to Ug99 stem rust race group. Science. 2013;341:783–786. doi: 10.1126/science.1239022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shumayla Sharma S, Kumar R, Mendu V, Singh K, Upadhyay SK. Genomic dissection and expression profiling revealed functional divergence in Triticum aestivum leucine lich repeat receptor like kinases (TaLRRKs) Front Plant Sci. 2016;7:1374. doi: 10.3389/fpls.2016.01374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh RP, Hodson DP, Jin Y, Lagudah ES, Ayliffe MA, Bhavani S, Rouse MN, Pretorius ZA, Szabo LJ, Huerta-Espino J, Basnet BR, Lan CX, Hovmøller MS. Emergence and spread of new races of wheat stem rust fungus: continued threat to food security and prospects of genetic control. Phytopathology. 2015;105:872–884. doi: 10.1094/PHYTO-01-15-0030-FI. [DOI] [PubMed] [Google Scholar]
- Stakman EC, Stewart DM, Loegering WQ. Identification of physiologic races of Puccinia graminis var. tritici. Washington DC: United States Department of Agriculture Research Service E-617; 1962. [Google Scholar]
- Steuernagel B, Periyannan SK, Hernandez-Pinzon I, Witek K, Rouse MN, Yu G, Hatta A, Ayliffe M, Bariana H, Jones JD, Lagudah ES, Wulff BB. Rapid cloning of disease-resistance genes in plants using mutagenesis and sequence capture. Nat Biotechnol. 2016;34:652–655. doi: 10.1038/nbt.3543. [DOI] [PubMed] [Google Scholar]
- Sun X, Cao Y, Yang Z, Xu C, Li X, Wang S, Zhang Q. Xa26, a gene conferring resistance to Xanthomonas oryzae pv. oryzae in rice, encodes an LRR receptor kinase-like protein. Plant J. 2004;37:517–527. doi: 10.1046/j.1365-313X.2003.01976.x. [DOI] [PubMed] [Google Scholar]
- Tranquilli G, Lijavetzky D, Muzzi G, Dubcovsky J. Genetic and physical characterization of grain texture-related loci in diploid wheat. Mol Gen Genet. 1999;262:846–850. doi: 10.1007/s004380051149. [DOI] [PubMed] [Google Scholar]
- Zimin AV, Puiu D, Hall R, Kingan S, Clavijo BJ, Salzberg SL (2017) The first near-complete assembly of the hexaploid bread wheat genome, Triticum aestivum. bioRxiv:159111 [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.