

Abstract
Aim
Central nervous system (CNS) metastasis is a major obstacle in the treatment of leukemia, and the underlying mechanisms of leukemia CNS metastasis are not fully understood. The present study is an investigation of the role of the CNS microenvironment in leukemia CNS metastasis.
Methods
Analog blood–brain barrier (BBB) was set by coculturing human brain microvascular endothelial cells (HBMVECs) and leukemia cells (U937 and IL-60), as well as HBMVECs and sera from leukemia patients, in vitro. The permeability of the HBMVEC monolayer and the levels of tight junction proteins, cytokines and chemokines (C&Ckines) were measured.
Results
The permeability of HBMVECs increased when cocultured with leukemia sera. The expression of C&Ckines was significantly upregulated in HBMVECs cocultured with leukemia sera or leukemia cells, compared to the normal sera (P<0.05, respectively). Specifically, significantly higher levels of vascular endothelial growth factor A (VEGF-A) and matrix metalloprotease 9 (MMP-9) were found in HBMVECs and leukemia cells/sera coculturing systems.
Conclusion
Both leukemia cells and the molecules in leukemia sera play an important role in leukemia CNS metastasis. VEGF-A and MMPs may be the main factors resulting in the degradation of the BBB and inducing the CNS migration of leukemia cells.
Keywords: CNS leukemia, cytokine, chemokine, U937, IL-60
Introduction
Central nervous system (CNS) relapse is the most important complication and obstacle in the treatment of leukemia. Despite extensive efforts, the underlying mechanisms of leukemia CNS metastasis are yet to be fully elucidated. The metastatic cascade includes cancer cell intravasation, circulation, adhesion, and growth in the CNS.1 Although cancer cells may invade the CNS through different pathways, hematogenous seeding plays a significant role in the spreading of cancer cells to the arachnoid mater.2,3 The blood–brain barrier (BBB) is crucial for insulating the CNS microenvironment from the peripheral circulation, efficiently excluding leukocytes, which are otherwise capable of routinely transmigrating across common endothelial layers.4 Brain microvascular endothelial cells (BMVECs) are the main component of the BBB.5 As the first step in CNS leukemia (CNSL), leukemic cells from the bone marrow (BM) migrate toward the brain and bind to the BBB. To overcome the effects of blood flow, leukemic cells adhere to BMVECs for migration.6 After adhesion, leukemia cells disrupt the extracellular matrix (ECM) and the luminal tight junction (TJ) barrier by releasing active biomolecules (cytokines, chemokines, growth factors, and matrix metalloproteases [MMPs]) and then breach the BBB barrier. Finally, to establish CNS foci, leukemia cells must renew themselves at their new locations.7–10 The intricate structure of the BBB makes metastatic extravasation into the brain significantly slower than in other organs,11 and the process may also require the synergetic input of a panel of molecules.
Molecules involved in tumor metastasis, including cytokines, chemokines, and MMPs, have been extensively studied in leukemia.1,12,13 Chemokines are a family of molecules that govern the homing of leukocytes into different organs, including the CNS.14 CCL2/monocyte chemoattractant protein-1 (MCP-1) is a dominant chemotactic factor that may offer a specific driving stimulus in response to CCR2-expressing leukocytes that migrate across the BBB.15,16 CXCL12 (the ligand for CXCR4) is an important chemokine that attracts and retains leukemia cells within the BM microenvironment. CXCR4 allows the migration and homing of neoplastic cells to sites where stromal cells express CXCL12.17 MMPs hydrolyze the ECM components in tissues surrounding the tumor, which facilitates the invasion of tumor cells through the basement membrane to distant organs, resulting in metastasis.18 In leukemia, MMP-2 and MMP-9 digest the TJs, disrupting the integrity of the luminal aspect of the BBB; therefore, the activity of MMPs is closely correlated with the organ invasiveness of leukemia cells.19 In addition, MMPs play a synergistic role along with vascular endothelial growth factor (VEGF) in promoting angiogenesis and metastasis.20 VEGF, a crucial factor in increasing the permeability of the endothelium, has previously been reported to induce disruption of the BBB, culminating in its breakdown.13,21 Vascular cell adhesion molecule 1 (VCAM-1) promotes cell adhesion and diapedesis of leukocytes.22,23 The interaction of endothelial VCAM-1 with the very late antigen 4 (VLA-4) expressed on leukocytes plays a key role in the transmigration of lymphocytes across the BBB and is implicated in both the capture and strong adhesion of leukocytes to CNS microvessels.6,24 Previous in vivo studies revealed strong, positive associations between the expression of cytokines and chemokines (C&Ckines) and the development of CNSL.1 Moreover, C&Ckines in the CNS microenvironment might offer more insight into the status of CNSL, compared to those in serum.1,13 However, the underlying mechanisms of how C&Ckines affect metastasis are unclear.
Both leukemia cells and BMVECs could secrete C&Ckines in autocrine or paracrine loops, when leukemia cells traverse the BBB.25 Circulating C&Ckine levels may be determined by the degradation of the BMVECs or the consumption during leukocyte transmigration from BM, as well as across the BBB. In this study, to mimic leukemia metastasis to the CNS, a human BMVECs (HBMVECs) culture system was developed in vitro. Subsequently, we attempted to determine whether sera from leukemia patients or leukemic cells in the CNS microenvironment affect the integrity of BMVECs, through the release of abundant C&Ckines into the CNS microenvironment.
Methods
This study was approved by the ethics committee of Shantou University Medical College. Written informed consent was obtained from all patients.
Cell culture and treatment
Leukemia sera were collected from patients with acute myeloid leukemia (AML) and acute lymphoblastic leukemia (ALL), respectively. Detailed patient information is presented in Table 1. Normal sera were collected from four healthy individuals. Sera were inactivated by heating for 30 min, at 56°C, before use. HBMVECs (ScienCell Research Laboratories, Shanghai, People’s Republic of China) were cultured in 24-well tissue culture inserts and 24-well plates from Corning Incorporated (Corning, NY, USA). BMVECs were grown in ECM media (ScienCell Research Laboratories) containing a low concentration of fetal bovine serum (FBS) (5%), in a humidified atmosphere of 5% CO2/air. The cells were seeded in 24-well plates with Transwell inserts, at 6 × 104 cells/cm2. When the cell density surpassed 80%, the basic medium was replaced with Dulbecco’s Modified Eagle Medium (DMEM; ScienCell Research Laboratories), with different ratios of sera (30% AML sera, 30% mixed ALL sera, and 30% sera from healthy people).
Table 1.
General characteristics of leukemia patients
| Parameters | AML | ALL |
|---|---|---|
| Gender (male/female) | 2/2 | 2/2 |
| Age (years) | 31.00 (22.00–50.50) | 33.00 (14.00–45.00) |
| WBCBlood (10E+9/L) | 4.40 (2.52–5.24) | 1.83 (0.51–7.92) |
| HbBlood (g/L) | 96.50 (73.75–120.00) | 111.50 (104.75–121.25) |
| PLTBlood (10E+9/L) | 273.50 (90.25–376.50) | 152.00 (76.25–312.50) |
| WBCCSF (10E+6/L) | 6.00 (3.00–13.50) | 6.00 (2.50–11.50) |
| BBB value | 8.39 (6.50–10.80) | 9.90 (7.03–13.18) |
| sVCAM-1CSF (pg/mL) | 1912.31 (913.69–2536.07) | 1534.97 (939.37–2018.74) |
| sVCAM-1Serum (ng/mL) | 349.95 (226.12–587.45) | 169.80 (96.07–266.06) |
| MMP-2CSF (pg/mL) | 680.96 (541.02–760.00) | 939.58 (825.12–1066.96) |
| MMP-2Serum (pg/mL) | 5363.81 (4320.50–6396.16) | 7064.78 (3426.89–8299.59) |
| MMP-9CSF (pg/mL) | 0.56 (0.51–0.62) | 0.57 (0.46–0.65) |
| MMP-9Serum (pg/mL) | 351.61 (190.44–725.71) | 401.90 (184.37–1395.83) |
| CXCL12CSF (pg/mL) | 318.66 (230.13–399.16) | 383.14 (290.50–447.28) |
| CXCL12Serum (pg/mL) | 435.15 (397.11–514.08) | 520.56 (362.34–823.28) |
| CCL2CSF (pg/mL) | 167.22 (86.48–268.20)* | 778.84 (451.29–996.19) |
| CCL2Serum (pg/mL) | 35.73 (32.27–63.83) | 58.96 (38.93–241.03) |
| VEGF-ACSF (pg/mL) | 127.90 (65.89–133.93) | 95.31 (79.63–152.82) |
| VEGF-ASerum (pg/mL) | 407.12 (272.38–542.58) | 597.48 (388.16–1259.95) |
Note:
P≤0.05 when the ALL group is compared with the AML group. Data shown as n (IQR).
Abbreviations: ALL, acute lymphoblastic leukemia; AML, acute myeloid leukemia; CSF, cerebrospinal fluid; WBC, white blood cell; Hb, hemoglobin; PLT, platelet; WBCCSF, WBC in CSF; IQR, inter-quartile range.
Vascular permeability assay
A total of 6 × 104 of HBMVECs were grown onto 24-well tissue culture inserts (Greiner Bio-One GmbH [Kremsmünster, Austria], transparent polyethylene terephthalate [PET] membrane, 0.4 μm pore size, and 4.0 × 106 pores/cm2) and incubated at 37°C (5% CO2) in the upper and lower chambers. Paracellular permeability was assessed using 0.06% bovine serum albumin (BSA; ScienCell Research Laboratories). The transmittance was measured with 0.06% BSA on days 1, 5, 10, and 15 to determine the permeability of BMVECs.26
Leukemia cells cocultured with BMVECs in vitro
Leukemia cells U937 and HL60 (ScienCell Research Laboratories) were cocultured with HBMVECs, respectively. After reaching confluence, the BMVECs were seeded on the upper chamber of a Transwell plate, while leukemia cells were added in the lower chamber, with a density of 5.0 × 106 cells/L. Supernatants were collected on days 1, 5, 10, and 15 and stored at −80°C.
Immunohistochemistry
HBMVECs were stained with human Zonula occludens-1 (ZO-1) antibody (Thermo Fisher Scientific, Waltham, MA, USA; Dako Denmark A/S, Glostrup, Denmark), according to the kit instructions. Slides were analyzed using Olympus Image Scope Software algorithms for staining intensity and the number of positive cells.
Cytokine measurement
The concentrations of sVCAM-1, MMP-2, MMP-9, VEGF, CXCL-12, and CCL2 in culture supernatants, sera, and cerebrospinal fluid (CSF) were measured by using the Luminex system on a Bioplex (Bio-Rad Laboratories Inc., Hercules, CA, USA).
Data analyses
Data were presented as medians, with interquartile ranges. The Kruskal–Wallis H test, Mann–Whitney U test, and Wilcoxon Rank-Sum test were used to evaluate the differences among each group. A P-value of <0.05 was considered statistically significant.
Results
The effect of leukemia sera/cells on HBMVEC permeability
To investigate the effect of leukemia sera on BMVEC permeability, FBS was replaced with leukemia sera in our culture system. HBMVECs cultured in leukemia sera could be sustained longer than those cultured in FBS, and the longest survival time was ~15 days (Figure 1), indicating that active molecules in leukemia sera may influence HBMVEC survival in vitro.
Figure 1.
Human brain microvascular endothelial cells cultured with different media.
Note: Magnification: 400×.
The baseline permeability was 10%, with HBMVEC permeability increasing to static baselines; on day 5, the permeability reached 30% for the ALL sera group and 40% for the AML sera group vs 20% for the control group. From day 10, the permeability increased sharply in the leukemia sera groups. From day 10 to day 15, the permeability was significantly higher in both the AML and ALL groups than in the control group. Specifically, HBMVECs cultured in AML sera had significantly higher permeability (Figure 2).
Figure 2.


Permeability of HBMVECs and concentration of cytokines in different cell culture media, after exposure to different factors, on different days.
Notes: M, HBMVEC exposure to culture medium containing sera of AML patients; L, HBMVEC exposure to culture medium with sera of ALL patients; C, HBMVEC exposure to culture medium with sera of healthy controls; HL60, HBMVEC exposure to culture medium with HL60 cells; and U937, HBMVEC exposure to culture medium with U937 cells.
Abbreviations: ALL, acute lymphoblastic leukemia; AML, acute myeloid leukemia; BSA, bovine serum albumin; HBMVECs, human brain microvascular endothelial cells.
In the BMVECs and leukemia cells coculture, the permeability increased from day 5 to day 10; however, permeability decreased from day 10 to day 15, which may be caused by the over proliferation of leukemia cells, covering the Transwell membrane filter (Figure 2).
The effect of leukemia sera on C&Ckines production by HBMVECs
The expression levels of sVCAM-1, CCL2, and VEGF-A were significantly increased when HBMVECs were cultured in leukemia sera, compared to those measured in normal sera (P<0.05). Specifically, the concentrations of sVCAM-1, CCL2, MMP-9, and VEGF-A produced by HBMVECs cultured in AML sera were significantly higher than those cultured in ALL sera (P<0.05). Contrary to MMP-9, the expression of MMP-2 was significantly higher in BMVECs cultured in ALL sera than in sera of AML patients and healthy controls (P<0.05). There was no statistically significant difference in the expression of CXCL12 among the three groups (P>0.05) (Table 2 and Figure 2).
Table 2.
Cytokines expression in HBMVECs and leukemia sera cocultured system
| Parameters | AML serum | ALL serum | Control serum |
|---|---|---|---|
| sVCAM-1 (pg/mL) | 11,258.17 (9150.88–11,389.31) | 4803.93 (4567.06–9818.30)* | 4128.01 (3622.83–8764.46)* |
| MMP-2 (pg/mL) | 865.82 (821.64–997.75) | 1802.72 (1422.30–2072.33)** | 860.79 (796.25–1038.08)♦♦ |
| MMP-9 (pg/mL) | 66.72 (50.78–84.86) | 16.33 (11.46–19.84)** | 479.86 (323.93–602.32)**,♦♦ |
| CXCL12 (pg/mL) | 397.20 (381.79–403.47) | 415.86 (399.15–419.00)* | 415.65 (396.03–419.26)* |
| CCL2 (pg/mL) | 1328.26 (1261.81–1345.90) | 154.96 (138.21–162.46)** | 1044.28 (880.23–1170.94)*,♦♦ |
| VEGF-A (pg/mL) | 144.75 (129.08–173.77) | 92.77 (80.69–96.26)** | 68.05 (65.12–70.38)**,♦♦ |
Notes: The concentrations of C&Ckines were measured in the supernatant of HBMVECs and leukemia sera cocultured system.
P<0.05 and
P<0.01 when compared with the AML serum group.
P<0.01 when compared with the ALL serum group. Data shown as n (IQR).
Abbreviations: ALL, acute lymphoblastic leukemia; AML, acute myeloid leukemia; C&Ckines, cytokines and chemokines; HBMVECs, human brain microvascular endothelial cells; IQR, inter-quartile range.
Leukemia cells cocultured with HBMVECs affect the C&Ckines production
To determine whether leukemia cells interact with the HBM-VECs in the CNS microenvironment, we cocultured HBM-VECs with U937 (lymphoid cell line) and HL60 (myeloid cell line). The baseline levels of C&Ckines in coculture supernatant were undetectable or very low. After coculturing with leukemia cells, higher CXCL12 and sVCAM-1 levels were observed in U937 cells, whereas higher MMP-2, MMP-9, VEGF-A, and CCL2 levels were found in HL-60 cells (Table 3). All C&Ckines levels were much lower in sole HBMVECs than those in the cocultured groups (Figure 2). For example, the highest level of VEGF-A was detected in the HL60 cocultured system. The production of VEGF-A was 48.89-fold and that of MMP-9 was 88.4-fold higher than in the leukemia sera. In the U937 cocultured system, the production of VEGF was 35.94-fold and that of MMP-9 was 77.3-fold higher than in the ALL sera (Tables 2 and 3). Our data demonstrate that leukemia cells are more potent in activating the production of MMP-9 and VEGF-A by HBMVECs, compared to leukemia sera. In addition, upon comparing the myeloid and lymphoid cell lines, the former seem to have more strength in inducing higher levels of VEGF-A and MMP-9 (Table 3).
Table 3.
Cytokines expression in HL60/U937 and HBMVECs cocultured system
| Parameters | HL60 | U937 | Control |
|---|---|---|---|
| sVCAM-1 (pg/mL) | 854.77 (448.04–931.71) | 1286.26 (1136.05–1511.09)## | – |
| MMP-2 (pg/mL) | 200.24 (186.57–369.48) | 98.38 (6.25–193.30)# | – |
| MMP-9 (pg/mL) | 5894.30 (5124.31–8637.63) | 1263.17 (628.28–2876.39)## | – |
| CXCL12 (pg/mL) | 426.71 (182.01–721.56) | 986.23 (378.33–1216.69) | 143.32 |
| CCL2 (pg/mL) | 324.21 (167.14–1024.71) | 90.52 (9.59–172.61)# | 4.05 |
| VEGF-A (pg/mL) | 7078.21 (5373.40–17,453.72) | 3334.28 (1076.67–4560.29)## | – |
Notes: The concentrations of C&Ckines were measured in the supernatant of HL60/U937 and HBMVECs cocultured system.
P<0.05 and
P<0.01 when compared with HL60 group. “–”: not detected.
Abbreviations: C&Ckines, cytokines and chemokines; HBMVECs, human brain microvascular endothelial cells.
TJ proteins in the cocultured systems
To further analyze whether higher production of VEGF-A and MMP-9 affects the integrity of the BBB by disrupting the TJs of HBMVEC, fluorescence staining of TJs was performed. The results show that the expression of ZO-1 in HBMVECs decreased after culturing with leukemia cells. ZO-1 (red staining) was restricted to junctional areas and some punctate staining in the cytoplasm (arrows) (Figure 3). Exposure to leukemia cells resulted in a weaker junctional and cytoplasmic pattern of ZO-1 immunoreactivity. Immunohistochemistry showed similar results to those obtained by immunofluorescent staining.
Figure 3.
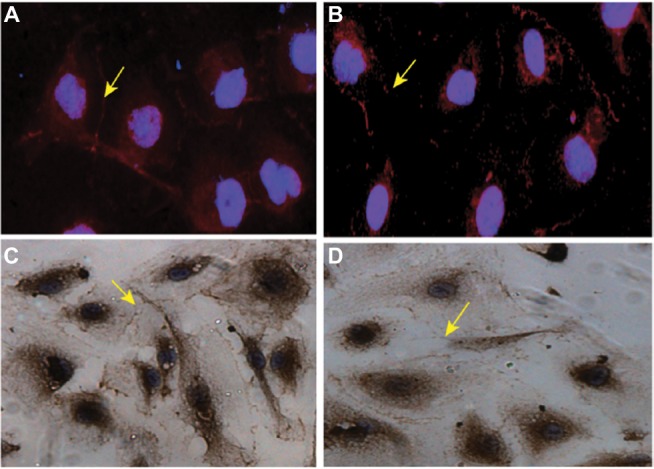
ZO-1 expression in HBMVECs.
Notes: (A) ZO-1 expression in HBMVECs before exposure to leukemia cells, as determined by immunofluorescence microscopy. (B) ZO-1 expression in HBMVECs after exposure to leukemia cells, as determined by immunofluorescence microscopy. (C) ZO-1 expression in HBMVECs before exposure to leukemia cells, as determined by immunohistochemistry. (D) ZO-1 expression in HBMVECs after exposure to leukemia cells, as determined by immunohistochemistry. ZO-1 immunoreactivity (red staining) was restricted to junctional areas and some punctate staining in the cytoplasm (arrows). Exposure to leukemia cells resulted in a weaker junctional and cytoplasmic pattern of ZO-1 immunoreactivity. DAPI (blue staining) was used to visualize the nuclei. Magnification: 1000×.
Abbreviations: HBMVECs, human brain microvascular endothelial cells; ZO-1, Zonula occludens-1; DAPI, 4′,6-diamidino-2-phenylindole.
Discussion
It is now well established that the tumor microenvironment has a major influence on tumor development, invasion, and metastasis.27 Our data show that BBB integrity relies on the orchestrated role of the C&Ckines secreted by leukemic cells and BMVECs in the BBB microenvironment. The crosstalk and cooperation between BMVECs and leukemia cells are critical for the maintenance of functional barrier integrity,28 as well as for maintaining the homeostasis of the microenvironment through their constitutive cytokine release.29–31
The production of CXCL12, CCL2, and sVCAM-1, which are the important chemokines attracting and retaining leukemia cells from the BM to the CNS, significantly increased when HBMVECs were cocultured with leukemia cells. Higher sVCAM-1 may accelerate circulating leukemic cells to target and adhere to HBMVECs, making the interaction with the endothelium possible, whereas higher CCL2 leads to the formation of new foci in the CNS, enhancing the possibility of white blood cells entering the CNS. Our study reveals that HBMVECs, under the influence of leukemia cells, may be the main source of sVCAM-1 and CCL2 in the CNS, which promote the transfer of leukemia cells to the CNS. Therefore, the levels of C&Ckines in the CNS microenvironment, rather than in serum, are more precise in predicting leukemia CNS metastasis.10
The degradation of TJs by MMP-2 and MMP-9 constitutes an important mechanism in the breakdown of the BBB, which contributes to the invasion of the CNS in acute leukemia.19 In our study, leukemia cells activated the HBMVECs to produce significantly higher levels of MMP-2, MMP-9, and VEGF-A, revealing that leukemia cells cross the BBB by stimulating the production of cytokines and then interrupting the ECM and TJs of BMVECs. These results are supported by evidence in vivo, which suggests that higher levels of VEGF-A and MMP-9 in the CSF positively correlate with BBB disruption in patients with CNSL development.1,32 Compared with patients without metastasis, CNSL patients had higher levels of MMP-9 and VEGF-A in the CNS. Our results indicate that C&Ckines in the CNS microenvironment may be the main reason for leukemia CNS metastasis.
FBS likely lacks several important characteristics in mimicking the human physiological microenvironment and, hence, may not be fully adequate for human functional studies, either ex vivo or in vitro. A previous study showed that human and mouse BMVECs cultured in an optimized concentration of growth factors (eg, EGF) gained extended lifetime, as well as good viability and purity.33 Therefore, we presumed that cells cultured in leukemia sera may be provided with more realistic environmental options to investigate the interactions between the peripheral system and the CNS. In our results, the expression levels of sVCAM-1, MMP-2, and VEGF-A increased after culturing in leukemia sera compared with healthy sera, revealing that bioactive substances in leukemia sera may jointly participate in BBB disruption.34 Moreover, a positive relationship exists between the concentrations of VEGF-A and MMP-2, MMP-9, and cell permeability, revealing that interactions between BMVECs and cytokines may occur in a dose-dependent manner. If C&Ckines are produced by leukemic cells and BMVECs, through either autocrine or paracrine loops, C&Ckines in the CNS may not be affected by the periphery.35 This may explain the clinical data suggesting that cytokines in CSF are more potent in influencing BMVECs, than cytokines in serum. C&Ckines produced in the CNS microenvironment may be more potent in recruiting leukemia cells and inducing the breakdown of the BBB.21,36,37 Our study reveals that both leukemia cells and the molecules in leukemia sera play an important role in leukemia CNS metastasis. VEGF-A and MMPs may be the main factors resulting in the degradation of the BBB and inducing the migration of leukemia cells to the CNS.
Acknowledgments
This study was supported by the Department of Education, Guangdong Government, under the top-tier University Development Scheme for Research and Control of Infectious Diseases.
Footnotes
Disclosure
The authors report no conflicts of interest in this work.
References
- 1.Si MY, Fan ZC, Li YZ, Chang XL, Xie QD, Jiao XY. The prognostic significance of serum and cerebrospinal fluid MMP-9, CCL2 and sVCAM-1 in leukemia CNS metastasis. J Neurooncol. 2015;122(2):229–244. doi: 10.1007/s11060-014-1707-8. [DOI] [PubMed] [Google Scholar]
- 2.Tabouret E, Bauchet L, Carpentier AF. Brain metastases epidemiology and biology. Bull Cancer. 2013;100(1):57–62. doi: 10.1684/bdc.2012.1681. [DOI] [PubMed] [Google Scholar]
- 3.Gomes HR. Cerebrospinal fluid approach on neuro-oncology. Arq Neuropsiquiatr. 2013;71(9B):677–680. doi: 10.1590/0004-282X20130149. [DOI] [PubMed] [Google Scholar]
- 4.Ransohoff RM, Kivisakk P, Kidd G. Three or more routes for leukocyte migration into the central nervous system. Nat Rev Immunol. 2003;3(7):569–581. doi: 10.1038/nri1130. [DOI] [PubMed] [Google Scholar]
- 5.Abbott NJ, Patabendige AA, Dolman DE, et al. Structure and function of the blood-brain barrier. Neurobiol Dis. 2010;37(1):13–25. doi: 10.1016/j.nbd.2009.07.030. [DOI] [PubMed] [Google Scholar]
- 6.Basu SK, Remick SC, Monga M, Gibson LF. Breaking and entering into the CNS: clues from solid tumor and nonmalignant models with relevance to hematopoietic malignancies. Clin Exp Metastasis. 2014;31(2):257–267. doi: 10.1007/s10585-013-9623-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Martin TA, Jiang WG. Loss of tight junction barrier function and its role in cancer metastasis. Biochim Biophys Acta. 2009;1788(4):872–891. doi: 10.1016/j.bbamem.2008.11.005. [DOI] [PubMed] [Google Scholar]
- 8.Quintana E, Shackleton M, Sabel MS, et al. Efficient tumour formation by single human melanoma cells. Nature. 2008;456(7222):593–598. doi: 10.1038/nature07567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Civini S, Jin P, Ren J, et al. Leukemia cells induce changes in human bone marrow stromal cells. J Transl Med. 2013;11:298. doi: 10.1186/1479-5876-11-298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Reikvam H, Fredly H, Kittang AO, et al. The possible diagnostic and prognostic use of systemic chemokine profiles in clinical medicine the experience in acute myeloid leukemia from disease development and diagnosis via conventional chemotherapy to allogeneic stem cell transplantation. Toxins (Basel) 2013;5(2):336–362. doi: 10.3390/toxins5020336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lorger M, Lee H, Forsyth JS, Felding-Habermann B. Comparison of in vitro and in vivo approaches to studying brain colonization by breast cancer cells. J Neurooncol. 2011;104(3):689–696. doi: 10.1007/s11060-011-0550-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Horacek JM, Kupsa T, Vasatova M, Jebavy L, Zak P. Biochip array technology and evaluation of serum levels of multiple cytokines and adhesion molecules in patients with newly diagnosed acute myeloid leukemia. Exp Oncol. 2014;36(1):50–51. [PubMed] [Google Scholar]
- 13.Tang YT, Jiang F, Guo L, Si MY, Jiao XY. Expression and significance of vascular endothelial growth factor A and C in leukemia central nervous system metastasis. Leuk Res. 2013;37(4):359–366. doi: 10.1016/j.leukres.2012.10.008. [DOI] [PubMed] [Google Scholar]
- 14.Gomez AM, Martinez C, Gonzalez M, et al. Chemokines and relapses in childhood acute lymphoblastic leukemia: a role in migration and in resistance to antileukemic drugs. Blood Cells Mol Dis. 2015;55(3):220–227. doi: 10.1016/j.bcmd.2015.07.001. [DOI] [PubMed] [Google Scholar]
- 15.Eugenin EA, Osiecki K, Lopez L, Goldstein H, Calderon TM, Berman JW. CCL2/monocyte chemoattractant protein-1 mediates enhanced transmigration of human immunodeficiency virus (HIV)-infected leukocytes across the blood-brain barrier: a potential mechanism of HIV-CNS invasion and NeuroAIDS. J Neurosci. 2006;26(4):1098–1106. doi: 10.1523/JNEUROSCI.3863-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Deshmane SL, Kremlev S, Amini S, Sawaya BE. Monocyte chemoattractant protein-1 (MCP-1): an overview. J Interferon Cytokine Res. 2009;29(6):313–326. doi: 10.1089/jir.2008.0027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Orimo A, Weinberg RA. Stromal fibroblasts in cancer: a novel tumor-promoting cell type. Cell Cycle. 2006;5(15):1597–1601. doi: 10.4161/cc.5.15.3112. [DOI] [PubMed] [Google Scholar]
- 18.Valastyan S, Weinberg RA. Tumor metastasis: molecular insights and evolving paradigms. Cell. 2011;147(2):275–292. doi: 10.1016/j.cell.2011.09.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Feng S, Cen J, Huang Y, et al. Matrix metalloproteinase-2 and -9 secreted by leukemic cells increase the permeability of blood-brain barrier by disrupting tight junction proteins. PLoS One. 2011;6(8):e20599. doi: 10.1371/journal.pone.0020599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Engsig MT, Chen QJ, Vu TH, et al. Matrix metalloproteinase 9 and vascular endothelial growth factor are essential for osteoclast recruitment into developing long bones. J Cell Biol. 2000;151(4):879–889. doi: 10.1083/jcb.151.4.879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Argaw AT, Gurfein BT, Zhang Y, Zameer A, John GR. VEGF-mediated disruption of endothelial CLN-5 promotes blood-brain barrier breakdown. Proc Natl Acad Sci U S A. 2009;106(6):1977–1982. doi: 10.1073/pnas.0808698106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Frenette PS, Wagner DD. Adhesion molecules – Part 1. N Engl J Med. 1996;334(23):1526–1529. doi: 10.1056/NEJM199606063342308. [DOI] [PubMed] [Google Scholar]
- 23.Frenette PS, Wagner DD. Adhesion molecules – Part II: blood vessels and blood cells. N Engl J Med. 1996;335(1):43–45. doi: 10.1056/NEJM199607043350108. [DOI] [PubMed] [Google Scholar]
- 24.Takeshita Y, Ransohoff RM. Inflammatory cell trafficking across the blood-brain barrier: chemokine regulation and in vitro models. Immunol Rev. 2012;248(1):228–239. doi: 10.1111/j.1600-065X.2012.01127.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kornblau SM, McCue D, Singh N, Chen W, Estrov Z, Coombes KR. Recurrent expression signatures of cytokines and chemokines are present and are independently prognostic in acute myelogenous leukemia and myelodysplasia. Blood. 2010;116(20):4251–4261. doi: 10.1182/blood-2010-01-262071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Mitchell RW, Edmundson CL, Miller DW, Hatch GM. On the mechanism of oleate transport across human brain microvessel endothelial cells. J Neurochem. 2009;110(3):1049–1057. doi: 10.1111/j.1471-4159.2009.06199.x. [DOI] [PubMed] [Google Scholar]
- 27.Shuman Moss LA, Jensen-Taubman S, Stetler-Stevenson WG. Matrix metalloproteinases: changing roles in tumor progression and metastasis. Am J Pathol. 2012;181(6):1895–1899. doi: 10.1016/j.ajpath.2012.08.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Alvarez JI, Katayama T, Prat A. Glial influence on the blood brain barrier. Glia. 2013;61(12):1939–1958. doi: 10.1002/glia.22575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Reikvam H, Brenner AK, Hagen KM, et al. The cytokine-mediated crosstalk between primary human acute myeloid cells and mesenchymal stem cells alters the local cytokine network and the global gene expression profile of the mesenchymal cells. Stem Cell Res. 2015;15(3):530–541. doi: 10.1016/j.scr.2015.09.008. [DOI] [PubMed] [Google Scholar]
- 30.Lane SW, Scadden DT, Gilliland DG. The leukemic stem cell niche: current concepts and therapeutic opportunities. Blood. 2009;114(6):1150–1157. doi: 10.1182/blood-2009-01-202606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ito S, Barrett AJ, Dutra A, et al. Long term maintenance of myeloid leukemic stem cells cultured with unrelated human mesenchymal stromal cells. Stem Cell Res. 2015;14(1):95–104. doi: 10.1016/j.scr.2014.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Zhu JC, Si MY, Li YZ, et al. Circulating tight junction proteins mirror blood-brain barrier integrity in leukaemia central nervous system metastasis. Hematol Oncol. 2017;35(3):365–373. doi: 10.1002/hon.2289. [DOI] [PubMed] [Google Scholar]
- 33.Navone SE, Marfia G, Nava S, et al. Human and mouse brain-derived endothelial cells require high levels of growth factors medium for their isolation, in vitro maintenance and survival. Vasc Cell. 2013;5(1):10. doi: 10.1186/2045-824X-5-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Shimizu F, Sano Y, Takahashi T, et al. Sera from neuromyelitis optica patients disrupt the blood-brain barrier. J Neurol Neurosurg Psychiatry. 2012;83(3):288–297. doi: 10.1136/jnnp-2011-300434. [DOI] [PubMed] [Google Scholar]
- 35.Lowenberg B, Touw IP. Hematopoietic growth factors and their receptors in acute leukemia. Blood. 1993;81(2):281–292. [PubMed] [Google Scholar]
- 36.Tham E, Gielen AW, Khademi M, Martin C, Piehl F. Decreased expression of VEGF-A in rat experimental autoimmune encephalomyelitis and in cerebrospinal fluid mononuclear cells from patients with multiple sclerosis. Scand J Immunol. 2006;64(6):609–622. doi: 10.1111/j.1365-3083.2006.01851.x. [DOI] [PubMed] [Google Scholar]
- 37.Chi OZ, Hunter C, Liu X, Weiss HR. Effects of anti-VEGF antibody on blood-brain barrier disruption in focal cerebral ischemia. Exp Neurol. 2007;204(1):283–287. doi: 10.1016/j.expneurol.2006.11.001. [DOI] [PubMed] [Google Scholar]