

Abstract
Purpose
Intratumoral androgen synthesis (IAS) is a key mechanism promoting androgen receptor (AR)reactivation and anti-androgen resistance in castration-resistant prostate cancer (CRPC). However, signaling pathways driving aberrant IAS remain poorly understood.
Experimental Design
The effect of components of the AKT-RUNX2-osteocalcin (OCN)-GPRC6A-CREB signaling axis on expression of steroidogenesis genes CYP11A1 and CYP17A1 and testosterone level were examined in PTEN-null human PCa cell lines. Pten knockout mice were employed to examine the effect of Runx2 heterozygous deletion or abiraterone acetate (ABA), a prodrug of the CYP17A1 inhibitor abiraterone on Cyp11a1 and Cyp17a1 expression, testosterone level and tumor microenvironment (TME) remodeling in vivo.
Results
We uncovered that activation of the AKT-RUNX2-OCN-GPRC6A-CREB signaling axis induced expression of CYP11A1 and CYP17A1 and testosterone production in PTEN-null PCa cell lines in culture. Deletion of Runx2 in Pten homozygous knockout prostate tumors decreased Cyp11a1 and Cyp17a1 expression, testosterone level and tumor growth in castrated mice. ABA treatment also inhibited testosterone synthesis and alleviated Pten loss-induced tumorigenesis in vivo. Pten deletion induced TME remodeling, but Runx2 heterozygous deletion or ABA treatment reversed the effect of Pten loss by decreasing expression of the collagenase Mmp9.
Conclusions
Abnormal RUNX2 activation plays a pivotal role in PTEN loss-induced IAS and TME remodeling, suggesting that the identified signaling cascade represents a viable target for effective treatment of PTEN-null PCa including CRPC.
Keywords: PTEN, AKT, RUNX2, testosterone, abiraterone acetate, extracellular matrix, prostate cancer
Introduction
Most cases of advanced prostate cancer (PCa) are initially responsive to androgen deprivation therapy (ADT); however, the majority relapses and eventually evolves into CRPC. Accumulating evidence suggests that IAS plays a pivotal role in CRPC development and progression (1,2). These findings have led to the development of second-generation anti-AR therapeutics including the cytochrome P450 17A1 (CYP17A1) inhibitor abiraterone that inhibits IAS in prostate tumors (3). Despite the increased benefit of abiraterone in overall survival in clinic, most patients develop resistance after initial responses via unclear mechanisms (4).
Up to 70% of patients with advanced PCa exhibit loss of PTEN function or activation of the PI3K/AKT pathway (5). Aberrant activation of AKT frequently associates with PCa progression, poor prognosis and high risk of recurrence (6). PTEN loss also associates with worse recurrence-free survival in patients treated with radical prostatectomy (7)or ABA (8). These findings imply that PTEN loss may link to IAS. However, the molecular mechanisms by which loss of PTEN promotes IAS remain unexplored.
Invasion into the surrounding stromal niche is a prerequisite for distant metastasis of cancerous cells. The process requires the detachment of cancer cells from the primary tumor and the recapitulation of TME. The stroma-epithelium crosstalk plays a critical role in creating a vicious cycle of positive feedback, thereby facilitating aggressive local cancer growth and metastatic spread to distant organs.
Runt-related transcription factor 2 (RUNX2) is implicated in human PCa. It is frequently over expressed in cancers that metastasize predominantly to bone (9,10). This transcription factor has been shown to promote cancer cell metastasis by increasing expression of various types of matrix metalloproteinases (9-11), thereby causing collagen degradation. It has been shown previously that FOXO1 is a negative regulator of RUNX2 (12,13). Loss of PTEN or growth factor stimulation results in FOXO1 inactivation, which in turn induces aberrant activation of RUNX2 and promiscuous expression of its target genes (e.g. OCN, osteopontin (OPN), vascular endothelial growth factor (VEGF) and interleukin 8 (IL8)) involved in cell migration and invasion (12). To date, however, the exact mechanisms underlying RUNX2-induced PCa cell aggressiveness remain to be elucidated. In the present study, we demonstrated that abnormal activation of RUNX2 caused by PTEN inactivation or AKT activation induces IAS and TME remodeling, thereby promoting PCa tumorigenesis and progression.
Materials and Methods
Cell lines, cell culture and transfection, and RNA interference
PC-3 and LNCaP cell lines were purchased from ATCC, and C4-2 cell line was purchased from UroCorporation. All cell lines were cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum (FBS) (Thermo Fisher Scientific) or otherwise specified plus 100 μg/ml penicillin-streptomycin-glutamine (Thermo Fisher Scientific) at 37°C with 5% CO2. All cell lines were tested and authenticated (karyotyping, RUNX2 expression and PTEN loss status) within 6 months of the submission of the manuscript. Mycoplasma contamination was regularly examined using Lookout Mycoplasma PCR Detection Kit (Sigma-Aldrich). Plasmocin (Thermo Fisher Scientific) was routinely added to the cell culture medium to prevent or eliminate mycoplasma contamination. Cell transfection was performed by electroporation using an Electro Square Porator ECM 830 (BTX) as described previously or lipofectamine 2000 (Thermo Fisher Scientific) according to the manufacturer’s instructions. Transfection efficiencies of approximately 80% to 90% were routinely achieved.
Generation of prostate-specific Runx2 and Pten deletion mice
Runx2 heterozygous (Runx2+/-) mice were reported previously (14,15). Pten conditional knockout (Ptenloxp/loxp or PtenL/L) mice were generated originally in the laboratory of Dr. Hong Wu and purchased from Jackson Laboratory (Bar Harbor, Maine). PB-Cre transgenic mice were generated originally in the laboratory of Dr. Pradip Roy-Burman and acquired from the National Cancer Institute (NCI) Mouse Repository. Cohorts of Runx2+/-;Ptenpc-/-, Runx2+/- and Ptenpc-/- mice were generated from PB-Cre+;Runx2+/-;PtenL/L males and PtenL/L females, which were obtained by cross breeding PB-Cre males with Runx2+/- and PtenL/L females. All procedures were approved by the Mayo Clinic Institutional Animal Care and Use Committee (IACUC) and conform to the legal mandates and federal guidelines for the care and maintenance of laboratory animals in accordance with recommendations as described (16,17).
Genotyping, RNA extraction, and real-time polymerase chain reaction (PCR) analysis
Genomic DNA from mouse tails was purified by phenol/chloroform extraction and used as templates for genotyping PCR reactions. Genotyping of wild-type and knockout allele of Runx2, conditional alleles of Pten, and Cre transgene was performed according to previously described PCR protocols (16,17). Information for primers used is provided in Supplementary Table S1. Total RNA was isolated from cells or tissues with Trizol (Thermo Fisher Scientific) and cDNA was synthesized using the Super-Script kit from Thermo Fisher Scientific. Two-step real-time PCR was performed using the SYBR Green Mix (Bio-Rad) and an iCycler iQTM detection system (Bio-Rad) according to the manufacturers’ instructions. Both forward and reverse primers were used at a final concentration of 200 nM. Expression of Gapdh gene in each sample was used as an internal control.
Abiraterone acetate (ABA) treatment in mice
Four-month-old Ptenpc-/-male mice were castrated for one month. Mice were randomly assigned to two groups (n = 5/group) and treated with intraperitoneal (i.p.) injections of vehicle (5% benzyl alcohol and 95% safflower oil) or ABA (0.5 mmol/kg/d in vehicle) daily for 21 days (18).
Western blotting
Western blotting was performed as described previously (16,17). Antibodies used for western blotting were diluted at 1:1,000 to 1:2,000.
Statistics
Experiments were carried out with three or more replicates. Statistical analyses were performed using Student’s t-test unless otherwise stated. Values with P < 0.05 are considered statistically significant.
Additional methods
Addition methods are provided in Supplementary Information.
Results
AKT-RUNX2-OCN-GPRC6A-CREB signaling axis regulates steroidogenesis gene expression in PTEN-null PCa cells
CYP11A1 and CYP17A1 are members of the cytochrome P450 enzyme family that are critical for steroid biosynthesis. Importantly, CYP17A1 is the target of the next-generation endocrine therapeutic agent ABA. Previous studies in mice showed that Ocn acts as a bone hormone that binds to G-protein coupled receptor family C group 6 member A (Gprc6a) in Leydig cells to stimulate testosterone biosynthesis in the testis by inducing expression of Cyp11a1 and Cyp17a1 (19). OCN is a well-established target gene transcriptionally regulated by RUNX2 (12,20,21). We and others previously showed that FOXO1 binds to and inhibits the transcriptional activities of RUNX2 in PCa cells and osteoblasts, and this effect is abolished by activated AKT due to PTEN loss or insulin-like growth factor-1 (IGF-1)/insulin stimulation (12,13). Osteomimictric properties, such as elevated RUNX2 expression and OCN secretion, have been well documented in PTEN-null PCa cells (22). Furthermore, elevated expression of the GPRC6A gene is common in human PCa cell lines (23) and strongly associated with PCa risk (24). We therefore sought to determine whether expression of CYP11A1 and CYP17A1 genes is regulated by the AKT-RUNX2-OCN-GPRC6A signaling axis in PTEN-deficient PCa cells.
C4-2B is an AR-positive PTEN-null CRPC cell line that expresses high levels of RUNX2 (25,26). We treated C4-2B cells with the PI3K-mTOR dual inhibitor NVP-BEZ235. As expected, NVP-BEZ235 treatment inhibited AKT phosphorylation and importantly, it also decreased CYP11A1 and CYP17A1 mRNA expression (Fig. 1A). Similar results were obtained by treating C4-2B cells with the AKT inhibitor MK2206 (Fig. 1B). Next, we examined whether knockdown of RUNX2 decreases expression of these genes. As shown in Fig. 1C, endogenous RUNX2 was effectively knocked down by a pool of two independent shRNAs in C4-2B cells as demonstrated by western blot analysis. Depletion of RUNX2 not only largely diminished mRNA expression of CYP11A1 and CYP17A1 genes, but also blocked NVP-BEZ235-mediated inhibition of these two genes (Fig. 1C; Supplementary Fig. S1A). In agreement with the finding that FOXO1 acts as a negative upstream regulator of RUNX2 in PCa cells (12), expression of a constitutively active form of FOXO1 (FOXO1-A3), in which three AKT phosphorylation sites are mutated to three alanine residues, significantly decreased CYP11A1 and CYP17A1 mRNA expression in C4-2B cells, but had no overt effect on expression of RUNX2 mRNA and protein (Fig. 1D; Supplementary Fig. S1B).
Figure 1.
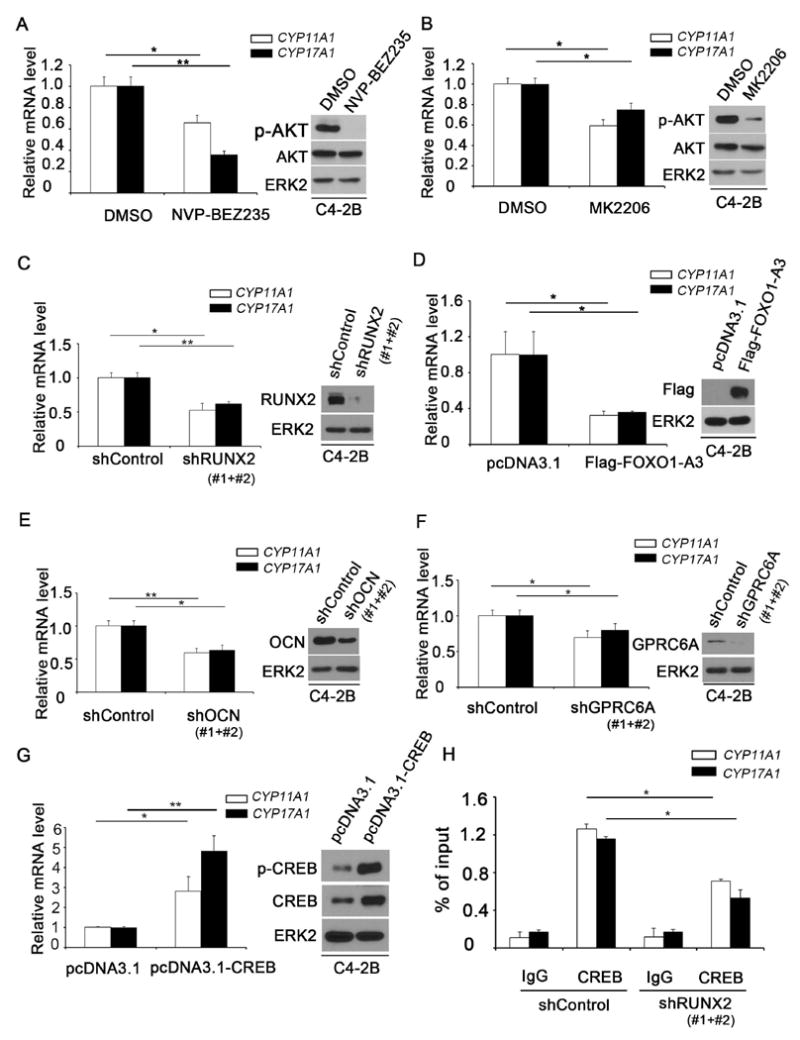
Effect of components of the AKT-RUNX2-OCN-GPRC6A-CREB signaling axis on expression of androgen biosynthesis enzyme genes in C4-2B CRPC cells. A and B, Treatment of C4-2B cells with the PI3K inhibitor NVP-BEZ235 (A) and the AKT inhibitor MK2206 (B) downregulated expression of CYP11A1 and CYP17A1 mRNA. The effectiveness of these drugs was determined by western blot analysis of AKT serine 473 phosphorylation (AKT-S473-p). ERK2 was used as a loading control. C-G, Expression of CYP11A1 and CYP17A1 mRNA in C4-2B cells is regulated by RUNX2 knockdown (C), overexpression of constitutively active FOXO1-A3 (D), OCN knockdown (E), GPRC6A knockdown (F) and ectopic expression of CREB (G). H, Effect of RUNX2 knockdown on CREB binding in the promoter of CYP11A1 and CYP17A1 genes in C4-2B cells. ChIP was performed using non-specific IgG or CREB-specific antibody followed by real-time PCR. A pool of two independent shRNAs (#1 and #2) was used for each gene examined. Data are means±S.D. from experiments with 3 replicates (n = 3). * P< 0.05**; P< 0.01.
The level of testosterone or the ratio of testosterone to estradiol is a crucial factor influencing PCa development during aging of men (27). The finding that mice lacking Ocn and Gprc6a show a significant change in the testosterone/estradiol ratio supports the notion that GPRC6A and OCN are two PCa relevant genes (19,28). OCN is readily expressed in C4-2B cells (29). We therefore examined the effect of OCN knockdown on expression of CYP11A1 and CYP17A1 mRNA in this cell line. Endogenous OCN was effectively knocked down by a pool of two independent shRNAs in C4-2B cells and this effect was correlated with decreased expression of both CYP11A1 and CYP17A1 genes (Fig. 1E). In contrast, overexpression of OCN prevented RUNX2 knockdown-induced downregulation of these two genes (Supplementary Fig. S1C and D).
Previous studies show that GPRC6A is highly expressed in various prostate cancer cell lines (23). While the endogenous GPRC6A protein was only partially (approximately 50%) depleted by infection of C4-2B cells with a pool of two independent shRNA, GPRC6A knockdown substantially decreased CYP11A1 and CYP17A1 mRNA expression (Fig. 1F). Finally, it has been shown that stimulation of mouse Leydig cells with Ocn results in activation of the transcriptional factor Creb and knockout of Creb in Leydig cells decreases expression of testosterone biosynthesis genes (19). We found that ectopic expression of CREB increased expression of CYP11A1 and CYP17A1 in C4-2B cells (Fig. 1G). Chromatin immunoprecipitation coupled quantitative polymerase chain reaction (ChIP-qPCR) assays revealed the binding of CREB protein in the promoter of CYP11A1 and CYP17A1 genes in C4-2B cells (Fig. 1H). Importantly, CREB occupancy at both promoters was largely diminished by RUNX2 knockdown (Fig. 1H), although there is no Runx binding site detected in this region. LNCaP (AR-positive) and PC-3 (AR-negative) are other two PTEN-negative PCa cell lines which also express high level of RUNX2, OCN and GPRC6A (23,25,30). Similar results were obtained in LNCaP and PC-3 cells (Supplementary Fig. S2 and 3). Together, these data indicate that activation of the AKT-RUNX2-OCN-GPRC6A-CREB signaling pathway induces expression of steroidogenesis genes CYP11A1 and CYP17A1 in human PTEN-null PCa cell lines.
Figure 3.

Effect of RUNX2 on the testosterone level and downstream signaling pathways in PCa cells in culture and in mice. A-C, PC-3 cells were infected with lentivirus expression control shRNA (shControl) or a pool of two independent gene-specific shRNAs for endogenous RUNX2 (A) and its downstream effectors OCN (B) and GPRC6A (C). 72 hr after infection, the effect of knockdown of these genes on cellular levels of testosterone was measured. The effectiveness of gene knockdown was confirmed by western blots and ERK2 was used as loading control. D-K, Effect of heterozygous deletion of Runx2 on the level of testosterone (D) and expression of androgen biosynthesis enzyme genes Cyp11a1 and Cyp17a1 (E), Ocn protein (F) and mRNA (G), Creb protein (H) and mRNA (I) and serine 133 (S133)-phosphorylated Creb protein (J and K) in prostate tumor tissues in prostate-specific Pten-knockout mice. Mice at age of 4 months were castrated for a month and testosterone level in prostate tumor tissues was measured using an ELISA-based kit, and protein and mRNA expression was measured by IHC and RT-qPCR, respectively. Data are means±S.D. from experiments with 3 mice per group (n = 3). * P < 0.05; ** P< 0.01.
PTEN-AKT pathway regulates testosterone levels in PCa cells in culture and in mice
An ELISA-based assay (Cayman Chemical) is commonly used for testosterone measurement in Leydig cells, serum and recurrent PCa tissues (1,19,31-33). We employed this well-established approach to determine whether the PTEN-AKT pathway regulates testosterone level in PCa cells. We cultured PC-3 cells in charcoal-stripped (androgen-depleted) medium to minimize the interference of exogenous testosterone from supplemented fetal bovine serum. We found that treatment of PC-3 with the PI3K-mTOR dual inhibitor NVP-BEZ235 significantly decreased testosterone level (Fig. 2A and B). Similar results were obtained when PC-3 cells were treated with the AKT inhibitor MK2206 (Fig. 2A and C). Consistent with a previous report (2), a very low level of testosterone was detected in prostate tissues in castrated wildtype mice (Fig. 2D). However, the testosterone level was significantly elevated in Pten-knockout prostate tumors in castrated mice compared with that in normal prostate tissues from castrated wildtype littermates (Fig. 2D; Supplementary Fig. S4A and B). In agreement with this observation, Pten deletion increased expressions of Cyp11a1 and Cyp17a1 mRNA in prostate tumors in castrated mice (Fig. 2E). We further showed that the testosterone concentration in serum was comparable between castrated Pten knockout mice and castrated wildtype littermates, but was much lower compared with that in serum of uncastrated wildtype mice (Fig. 2F). These data suggest that the source of the increased testosterone levels observed within Pten-null tumors was most likely originated from the tumor itself. Together, our data demonstrate that testosterone is synthesized in PTEN-deficient PCa cells in culture and in mice and this event is regulated by the PTEN-AKT pathway.
Figure 2.

Effect of the PI3K-PTEN-AKT pathway on the testosterone level in human and mouse PCa cells. A-C, Treatment with the PI3K inhibitor NVP-BEZ235 (A and B) and the AKT inhibitor MK2206 (A and C) decreased testosterone levels in PC-3 cells. The effectiveness of these drugs was determined by western blot analysis of AKT serine 473 phosphorylation (AKT-S473-p). ERK2 was used as a loading control. D and E, Homozygous deletion of Pten increased testosterone levels (D) and expression of androgen biosynthesis enzyme genes Cyp11a1 and Cyp17a1 (E) in prostate tumors in mice. F, Comparison of testosterone level in serum of uncastrated wildtype mice, castrated wild-type mice and castrated prostate-specific Pten knockout mice (n = 3/group). Mice at age of 4 months were castrated or sham castrated for a month and testosterone level in serum was measured using an ELISA-based kit. Data are means±S.D. from experiments with 3 replicates (n = 3). NS, not significant; * P< 0.05; **, P< 0.01.
RUNX2 regulates testosterone levels in PTEN-null PCa cells in culture and in mice
Given that the RUNX2-OCN-GPRC6A axis regulates expression of steroidogenesis genes CYP11A1 and CYP17A1 in RUNX2-expressing cell lines (Fig. 1; Supplementary Fig. S2 and 3), we sought to employ a loss-of-function approach to determine whether this pathway affects testosterone levels in PTEN-deficient PCa cells. We found that knockdown of endogenous RUNX2 decreased testosterone levels in PC-3 cells (Fig. 3A). Similar results were obtained in OCN- or GPRC6A-knockdown PC-3 cells (Fig. 3B and C). Importantly, in agreement with the regulation of CYP11A1 and CYP17A1 gene expression by the RUNX2-OCN pathway, OCN overexpression prevented RUNX2 knockdown-induced decrease in testosterone levels in C4-2B cells (Supplementary Fig. S1C-E). Next, we examined the effect of Runx2 knockout on testosterone levels in Pten-knockout tumors in castrated mice. Given that Runx2 homozygous deletion mice die right after birth (14), we generated Runx2 heterozygous mice in the background of prostate-specific Pten homozygous deletion. Although Runx2 was only heterozygously deleted, surprisingly, Runx2 mRNA and protein were largely depleted in the prostates (Supplementary Fig. S4A, C and D), and this result is consistent with the previous finding that Runx2 is a autoregulatory gene (34). In agreement with the findings in cultured human PCa cells, Runx2 heterozygous deletion substantially decreased testosterone levels in Pten-knockout prostate tumors in castrated mice (Fig. 3D). Accordingly, deletion of Runx2 decreased expression of Cyp11a1 and Cyp17a1 genes in prostate tumors in castrated Pten-knockout mice (Fig. 3E). It is worth noting that the expression level of Cyp11a1 was much higher than that of Cyp17a1 in Pten-deficient prostate tumors in castrated mice (Fig. 2E and 3E), but the exact underlying mechanism warrants further investigation. In agreement with these observations, heterozygous deletion of Runx2 also decreased expression of Ocn, a upstream regulator of these two genes in Pten-null prostate tumors in mice (Fig. 3F and G). While Runx2 heterozygous deletion had no overt effect on expression of Creb, it largely decreased the level of phosphorylated (active) Creb in Pten-null prostate tumors (Fig. 3H-K). Together, these data suggest that expression of RUNX2 is required for IAS in both human and mouse PTEN-null PCa cells.
Runx2 promotes Pten-null PCa growth and AR activity in castrated mice
We next examined the causal role of RUNX2 in PTEN loss-induced PCa growth using Pten knockout mice as a model. Consistent with the inhibitory effect on IAS in castrated mice (Fig. 3D), Runx2 heterozygous deletion largely decreased the incidence of high-grade prostatic intraepithelial neoplasia (HGPIN) and cancerous lesions under the castration condition (Fig. 4A-C). We further demonstrated that deletion of Runx2 significantly decreased Ki-67 staining in Pten-deficient prostate tumors in castrated mice (Fig. 4D and E). Runx2 heterozygous deletion in Pten-null prostate tumors from castrated mice also markedly increased cell apoptosis as demonstrated in IHC staining of cleaved caspase 3 (Fig. 4F and G). Thus, Runx2 reduction inhibited tumorigenesis in the Pten-knockout prostate in castrated mice by decreasing IAS and cell proliferation and increasing apoptotic cell death.
Figure 4.

Impact of Runx2 heterozygous deletion on cell proliferation, survival and tumorigenesis of Pten-knockout prostate epithelial cells in mice. A, Design and timeline of castration and histopathological analyses in mice with genotypes indicated. B and C, H&E (B) and quantitative analysis (C) of HGPIN/cancer lesions, LGPIN and nonmalignant acini in the anterior prostate (AP) of 6-month-old mice in the four genotypes indicated (n = 5/group). Scale bar, 200 μm. D-G, Assessment of expression of the proliferation marker Ki-67 (D and E) and the apoptotic marker cleaved caspase 3 (F and G) in the groups of mice at age as indicated in (A). Arrows indicate the positive staining of these markers. Scale bar, 200 μm. Data are means±S.D. from experiments with 3 mice per group (n = 3). * P < 0.05; ** P < 0.01. H, Analysis of Ar target genes by RT-qPCR in the groups of mice at age as indicated in (A). Results were normalized to housekeeping gene (Gapdh). I, IHC for Ar protein in the prostate of Ptenpc-/- and Ptenpc-/-;Runx2+/- castrated mice. Scale bar, 50 μm.
It was shown previously that Pten deletion results in downregulation of Ar protein in the mouse prostate and AR target gene expression (5,17,35). While its expression level is downregulated, Ar protein remains functional and is required for growth of Pten-deficient PCa cells under androgen depleted conditions (36). In agreement with these observations, we demonstrated that expression of AR target genes such as Probasin, Nkx3.1 and Tmprss2 was downregulated in Pten-knockout prostate tissues compared to wildtype counterparts (Fig. 4H). Importantly, expression of these genes was further reduced by co-deletion of Runx2 (Fig.4H). It has been well established that androgen depletion decreases AR protein level (37). Consistent with these findings, Runx2 heterozygous deletion decreased testosterone synthesis gene expression and intratumoral testosterone level in Pten-deficient prostate tumors in castrated mice (Fig. 3D and E) and also decreased Ar protein expression (Fig. 4I). These data demonstrate that Runx2 heterozygous deletion decreases Ar protein level and inhibits its transcriptional activity in Pten-deficient prostate tumors in castrated mice.
ABA treatment inhibits IAS and AR activity in Pten-deficient PCa in castrated mice
ABA impairs AR signaling by inhibiting adrenal and intratumoral androgen biosynthesis in vivo (38). Given that deletion of Runx2 decreased expression of Cyp17a1 mRNA and testosterone synthesis in Pten-knockout PCa in castrated mice, we examined the effect of ABA-mediated inhibition of Cyp17a1 on testosterone synthesis in the same mouse model. As expected, treatment of mice with ABA significantly decreased testosterone levels in PCa tissues in castrated Pten-knockout mice (Fig. 5A and B). Similar to the effect of Runx2 heterozygous deletion, ABA treatment also decreased tumor burden by substantially reducing the incidence of HGPIN and cancerous lesions in castrated Pten-knockout mice (Fig. 5C and D). ABA treatment also largely decreased Ki-67 staining and elevated cleaved caspase 3 expression (Fig. 5E-H). These data suggest that ABA treatment decreases prostate tumor burden in the Pten-knockout model by inhibiting cell proliferation and increasing cell death. In accordance with the results that ABA treatment reduced intratumoral testosterone level (Fig. 5B), ABA treatment also largely decreased expression of Ar protein and its target genes examined in Pten-deficient prostate tumors in castrated mice (Fig. 5I and J), indicating that AR activity is inhibited by ABA treatment in PTEN-null tumors in mice.
Figure 5.

Effort of ABA treatment on cell proliferation, survival and tumorigenesis of Pten-knockout prostate epithelial cells in mice. A, Design and timeline of castration and histopathological analyses in mock-or ABA-treated Pten knockout (Ptenpc-/-) mice. B, Effect of ABA on the level of testosterone in prostate tumor tissues from 6-month-old castrated mice treated with or without ABA as indicated in (A). C and D, quantitative analysis of HGPIN/cancer lesions, LGPIN and nonmalignant acini (C) and H&E analysis (D) of the anterior prostate (AP) of 6-month-old mice in the groups as shown in (A) (n = 5/group). Scale bar, 200 μm. E-H, Assessment of expression of the proliferation marker Ki-67 (E and G) and the apoptotic marker cleaved caspase 3 (F and H) in different groups of mice at age as indicated in (A). Arrows indicate the positive staining of these markers. Scale bar, 200 μm. Data are means±S.D. from experiments with 3 mice (n = 3). * P< 0.05; ** P< 0.01. I, Analysis of Ar target genes by RT-qPCR in the groups of mice at age as indicated in (A). Results were normalized to housekeeping gene (Gapdh). J, IHC for Ar in the prostate of Ptenpc-/- castrated mice with the age and treatment shown in (A). Scale bar, 50 μm.
Runx2 and ABA modulates TME remodeling in Pten-null PCa in castrated mice
Remodeling and stiffening of the stromal microenvironment offer an appropriate niche for cancer cell invasion and progression. As a transcription factor involved in osteoblast differentiation (22), RUNX2 has been studied intensively for its role in tumorigenesis in various types of cancer (9-11). In vitro studies suggest that RUNX2 promotes the aggressiveness of CRPC by enhancing cell invasive capability toward collagen (39). However, the role of RUNX2 in regulating stromal microenvironment of PCa in vivo is poorly undetermined.
A previous report (40) shows that the periacinar fibroblast layers appear to be wider in PIN lesions as an early sign of reactive stroma, implying a local activation of periacinar fibroblasts in response to PIN development. We examined the impact of the PTEN-RUNX2 pathway on stromal microenvironment using castrated Pten/Runx2 double knockout mice as a working system. We first performed IHC for smooth muscle actin (SMA). In the prostate of wildtype mice (Fig. 6A1), we detected a highly condensed layer of SMA-positive stroma, which is regarded as a natural barrier against the invasion of tumor cells into the adjacent stromal tissue. In contrast, we observed much loosened layers of SMA-positive stroma surrounding malignant epithelial cells in the prostate of Pten-knockout mice (Fig. 6A2). Indeed, invasive adenocarcinoma often induces a desmoplastic response, in which collagenous material is diffusely distributed in the surrounding stromal tissue. We performed Masson Trichrome stain which marks keratin and muscle fibers red and collagen blue. We detected a relatively condensed layer of collagen surrounding the stroma in the prostate of wildtype mice (Fig. 6B1). On the contrary, we found the existence of multiple layers of collagen (blue) mingled with layers of small muscle fibers (pink) in the prostate of Pten-knockout mice (Fig. 6B2). Thus, loss of PTEN promotes the recruitment of connective tissues to the diseased areas, which may likely lead to an invasive phenotype of tumor by commutating with the adjacent acini via collagen. SMA IHC showed that Runx2 heterozygous deletion alone had no overt effect on composition of SMA-positive stromal tissues in the prostate of mice without Pten loss (Fig.6A3) and this result is not surprising since Runx2 expression level is very low in the prostate of wildtype mice (26). Notably, Runx2 depletion almost completely reversed Pten knockout-induced formation of multiple layers of SMA-positive stroma surrounding malignant epithelial cells (Fig. 6A4). Similar results were obtained in Trichrome staining (Fig. 6B4). These findings in mice reveal an important role of RUNX2 in modulating PTEN loss-induced remodeling of PCa stromal environment under castration conditions. Similar to the effect of Runx2 heterozygous deletion, ABA treatment also largely inhibited PTEN loss-induced acquisition of loosened layers of SMA-positive stroma surrounding malignant epithelial cells (Fig. 6C). We further showed that ABA administration blocked deposition of collagen in the stromal environment (Fig. 6D). These findings suggest that RUNX2 depletion or ABA treatment prohibits the local invasive potential of PCa in the context of PTEN deletion by reducing extracellular matrix remodeling in the stromal compartment.
Figure 6.

Effort of Runx2 heterozygous deletion or ABA treatment on stromal niche in Pten-knockout prostate tumors in castrated mice. A, IHC for smooth muscle actin (SMA), pointed by red arrows, in the prostate of mice in the four genotypes indicated at age of 6 month as shown in Fig. 4A. Scale bar, 200 μm. B, Masson’s Trichrome staining identified smooth muscle cells (red) and collagen fibers (blue) in prostate stroma in the four indicated groups of mice as shown in Fig. 4A (n = 5/group). Scale bar, 200 μm. C, IHC for smooth muscle actin (SMA), pointed by red arrows, in the prostate of Pten knockout (Ptenpc-/-) mice treated with vehicle or ABA as shown in Fig. 5A (n = 5/group). Scale bar, 200 μm. D, Masson’s Trichrome staining identified smooth muscle cells (red) and collagen fibers (blue) in prostate stroma in Pten knockout (Ptenpc-/-) mice treated with vehicle or ABA as shown in Fig. 5A (n = 5/group). Scale bar, 200 μm.
ABA treatment modulates expression of chemokines and cytokines in the TME niche
The intimate interactions among each cell type in the neoplastic niche, such as epithelium, macrophage, fibroblast and other stromal cells, contribute significantly to tumorigenesis and cancer progression (41). Cross-talk between cytokine and chemokine pathways in the stroma can elicit tumorigenic responses. To investigate the molecular mechanisms by which ABA treatment induces TME remodeling, we examined the effect of ABA on expression of chemokine and cytokine genes in stroma of prostate tumor in the Pten-knockout mouse model. Using the method reported previously (42), we successfully isolated fibroblasts and macrophages from Pten-knockout prostate tumors in mice treated with or without ABA for a month (Supplementary Fig. S5A). The authentication of isolated fibroblasts and macrophages was confirmed by their expression of platelet derived growth factor receptor α (PDGFRα) and colony stimulating factor-1 receptor (CSF-1R), respectively (Supplementary Fig. S5B).
In cancer-associated fibroblasts (CAFs) in Pten-knockout prostate in castrated mice, ABA treatment induced significant downregulation of several cytokine and chemokine genes examined (Fig. 7A and B), including interferon γ-induced protein IP-10 (or termed mCXCL10), macrophage inflammatory protein-1a (MIP-1a or termed mCCL3), macrophage inflammatory protein-1b (MIP-1b or termed mCCL4), regulated upon activation, normal T-cell expressed and secreted (RANTES or termed mCCL5), interleukin-10 (mIL-10), transforming growth factor β1 (mTGFβ1). ABA treatment also significantly upregulated expression of several cytokine and chemokine genes in CAFs (Fig. 7C), including tissue inhibitor of metalloproteinase 1 (mTIMP1), keratinocyte-derived chemokine (KC or termed mCXCL1), chemokine (C-X-C motif) ligand-2 (mCXCL2), thymus and activation regulated chemokine (TARC or termed mCCL17), stromal cell derived factor-1 (mSDF1) and interleukin-1 β (IL-1β). We also detected similar expression changes of these cytokine and chemokine genes in tumor-associated macrophages, such as significant downregulation of several cytokines and chemokines in tumor-associated macrophages (Fig. 7D), including mCXCL10, mCCL3, mCCL4, mCCL5, mIL-10, mCXCL2, and IL-1β. We further showed that ABA induced significant upregulation of several cytokine and chemokine genes in tumor-associated macrophages (Fig. 7E).
Figure 7.

Expression of chemokine and cytokine genes in fibroblasts and macrophages and Mmp9 in epithelial compartments in mock-or ABA-treated Pten knockout prostate tumors in castrated mice. A, Design and timeline of castration and harvest of prostate tissues for FACS analyses from mock-or ABA-treated prostate-specific Pten knockout (Ptenpc-/-) mice. B and C, Downregulation (B) and upregulation (C) of indicated cytokine and chemokine genes in fibroblasts isolated from prostate tumors in castrated mice of the indicated groups as measured by RT-qPCR. D and E, Downregulation (D) and upregulation (E) of indicated cytokine and chemokine genes in macrophages isolated from prostate tumors in castrated mice of the indicated groups as measured by RT-qPCR. F and G, IHC (F) and quantitative (G) analysis for Mmp9 expression in Pten-knockout tumors in the indicated groups of castrated mice at age as shown in (A). Scale bar, 200 μm. Data are means±S.D. from experiments with 3 mice (n = 3). *P< 0.05; ** P< 0.01.
The functional importance of the CAFs in response to neoplastic epithelium can be attributed, at least in part, to the dynamics of expression of cytokines and chemokines in stromal cells and the feedback impact on cancerous epithelium. Matrix metallopeptidase 9 (MMP9) is a collagenase involved in degradation of the extracellular matrix. We demonstrated that Mmp9 protein expression was decreased in ABA-treated Pten-null prostate tumors compared to mock treatment (Fig. 7F and G). Runx2 heterozygous deletion also reduced Mmp9 expression in Pten-knockout prostate tumors in castrated mice (Supplementary Fig. S5C). Thus, both deletion of Runx2 and ABA treatment decrease Mmp9 expression in Pten-null tumors in mice, providing a plausible explanation of the effect of ABA treatment or Runx2 heterozygous deletion on TME remodeling.
Discussion
Increased synthesis of testosterone and further conversion to dihydrotestosterone (DHT) in PCa cells, through either de novo route after castration or from weak androgens produced by the adrenal gland, is one of the major mechanisms that likely contribute to AR reactivation during PCa progression. Findings from multi-institutional integrative clinical sequencing studies reveal that the PTEN gene is frequently lost in advanced PCa (43,44). Moreover, PTEN loss is associated with worse recurrence-free survival after radical prostatectomy (7) and androgen depletion therapy of patients (8). Thus, it can be postulated that PTEN loss in PCa may contribute to IAS. Indeed, Pten inactivation in mice inevitably results in the emergence of castration-resistant, but AR-dependent PCa, although the underlying mechanisms were largely unclear (36,45). We demonstrated previously that PTEN loss-induced inactivation of FOXO1 promotes oncogenic activation of RUNX2, thereby driving promiscuous expression of its downstream target genes involved in cell migration and invasion and PCa progression (12). OCN is a well-established RUNX2 target gene (12,46). Intriguingly, previous studies in mice show that OCN functions as a bone hormone that binds to the G protein-coupled receptor GPRC6A in Leydig cells to promote testosterone biosynthesis in the testis by inducing expression of androgen biosynthesis enzyme genes such as CYP11A1 and CYP17A1 (19). Based upon our findings in vitro and in mice in the present study, we envision a model (Supplementary Fig. S5D) wherein the activation of RUNX2 caused by PTEN loss promotes IAS in PCa via activation of the OCN-GPRC6A-CREB-CYP17A1 axis. Importantly, we provide in vivo evidence that treatment of Pten-null tumors in castrated mice with the CYP17A1 inhibitor ABA not only decreased testosterone levels in tumor tissues, but also caused inhibition of tumor growth and progression by inducing apoptosis in prostate tumors. Thus, our findings identify a signaling cascade that could be a potential target for effective treatment of PTEN-null CRPC.
Invasion of carcinoma cells into the surrounding stromal niche is a prerequisite for distant metastasis. A highly condensed layer of SMA-positive stroma, which is regarded as a natural barrier against the invasion of tumor cells into the adjacent stromal tissue, circuits around the acini in the normal prostate. We found much loosened layers of SMA-positive stroma surrounding malignant epithelial cells in the prostate of Pten-knockout mice. Runx2 heterozygous deletion almost completely reversed Pten knockout-induced formation of multiple layers of SMA-positive stroma outside malignant epithelial cells, suggesting a causal role of Runx2 in Pten loss-induced remodeling of TME. In agreement with the role of Runx2 in testosterone synthesis in Pten-knockout prostate tumors in mice, blockade of testosterone synthesis by ABA also abolishes Pten loss-induced remodeling of stromal niche in castrated mice. Thus, our findings demonstrated for the first time that RUNX2 plays an important role in modulation of TME and this effect is likely mediated through the impact on the stroma compartment imposed by RUNX2-mediated testosterone synthesis in tumor cells.
Our finding that Pten-knockout PCa cells intratumorally produce significant amount of testosterone provides a molecular explanation for the castration-resistance phenotype of prostate tumors in Pten-knockout mice. The importance of IAS in castration-resistant growth of Pten-null tumors is further supported by the observation that ABA treatment substantially reduces tumor burden by decreasing intratumoral testosterone levels. The effectiveness of ABA treatment is also manifested by decreased expression of Ar protein in mouse prostate tumors, a phenomenon consistent with previous finding in cultured cells that androgens induce accumulation of AR protein (37,47).
ADT regulates the state of active stroma and modulates expression of chemokines and cytokines by blocking AR signaling pathway in stromal cells in the context of PTEN haploinsufficiency (48). A number of chemokines and cytokines have been shown to be secreted by different stromal and epithelial cell types and play important roles in microenvironment-mediated promotion of tumorigenesis (49,50). We uncovered that ABA treatment substantially decreased expression of the collagenase MMP9 which plays a major role in degradation of the extracellular matrix. Our data suggest that while ABA treatment abrogates the AR signaling, it also induces TME remodeling by affecting expression of extracellular matrix modulators such as MMP9.
In summary, we demonstrated that PTEN loss or activation of AKT promotes IAS and this process is mediated by hyperactivation of RUNX2 and its downstream OCN-GPRC6A-CREB signaling cascade and increased expression of steroid biosynthesis enzyme genes CYP11A1 and CYP17A1. We also showed that Pten deletion induces TME remodeling in castrated mice.
However, the effects of PTEN loss on androgen biosynthesis and TME remodeling in prostate tumors are completely blocked by Runx2 heterozygous deletion or treatment of mice with ABA, a prodrug of the CYP17A1 inhibitor. Therefore, our findings not only uncover a potential mechanism of emergence of the intrinsic castration-resistant phenotype in PTEN-null PCa cells, but also identify an important role of the interplay between epithelial and stromal cells in development of resistance to abiraterone therapy in PTEN-deficient CRPC.
Supplementary Material
Statement of Translational Relevance.
PTEN is a tumor suppressor that is frequently mutated or deleted in prostate cancer (PCa), especially in castration-resistant prostate cancer (CRPC). Studies using genetic engineering mouse models show that PTEN deletion induces formation of castration-resistant, but androgen receptor (AR)-dependent PCa, although the underlying mechanisms remain poorly understood. In the present study we demonstrate that PTEN loss promotes activation of the AKT-RUNX2-OCN-GPRC6A-CREB signaling axis, which in turn induces expression of steroidogenesis genes CYP11A1 and CYP17A1 and intratumoral androgen synthesis (IAS) in cultured PCa cells. We further show that loss of PTEN augments steroidogenesis gene expression, IAS and tumor microenvironment (TME) remodeling in castrated mice and these effects are largely diminished by Runx2 heterozygous deletion or treatment of mice with abiraterone acetate (ABA), a prodrug of the CYP17A1 inhibitor abiraterone. Thus, our studies not only uncover that loss of PTEN promotes IAS and TME remodeling, but also identify the abnormal activation of the AKT-RUNX2-OCN-GPRC6A-CREB axis as a pivotal mechanisms that drives PTEN loss-induced IAS. Our findings suggest that targeting this signaling cascade can be harnessed for effective treatment of PTEN-null PCa, especially CRPC.
Acknowledgments
This work was supported in part by grants from NIH (CA134514, CA130908, and CA193239 to H. Huang), DOD (W81XWH-14-1-0486 to H. Huang), Natural Science Foundation of China (81270022, 81611130070 and 81771898 to W. Xu), and Natural Science Foundation of Heilongjiang Province of China (No. QC2014C111 to Y. Yang). Abiraterone acetate was kindly provided by the Janssen Research & Development, LLC.
Footnotes
Conflicts of Interest: The authors declare no conflict of interest in this study.
References
- 1.Mohler JL, Gregory CW, Ford OH, 3rd, Kim D, Weaver CM, Petrusz P, et al. The androgen axis in recurrent prostate cancer. Clin Cancer Res. 2004;10(2):440–8. doi: 10.1158/1078-0432.ccr-1146-03. [DOI] [PubMed] [Google Scholar]
- 2.Locke JA, Guns ES, Lubik AA, Adomat HH, Hendy SC, Wood CA, et al. Androgen levels increase by intratumoral de novo steroidogenesis during progression of castration-resistant prostate cancer. Cancer Res. 2008;68(15):6407–15. doi: 10.1158/0008-5472.CAN-07-5997. [DOI] [PubMed] [Google Scholar]
- 3.Attard G, Reid AH, Yap TA, Raynaud F, Dowsett M, Settatree S, et al. Phase I clinical trial of a selective inhibitor of CYP17, abiraterone acetate, confirms that castration-resistant prostate cancer commonly remains hormone driven. Journal of clinical oncology : official journal of the American Society of Clinical Oncology. 2008;26(28):4563–71. doi: 10.1200/JCO.2007.15.9749. [DOI] [PubMed] [Google Scholar]
- 4.Antonarakis ES, Lu C, Wang H, Luber B, Nakazawa M, Roeser JC, et al. AR-V7 and resistance to enzalutamide and abiraterone in prostate cancer. N Engl J Med. 2014;371(11):1028–38. doi: 10.1056/NEJMoa1315815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Mulholland DJ, Tran LM, Li Y, Cai H, Morim A, Wang S, et al. Cell autonomous role of PTEN in regulating castration-resistant prostate cancer growth. Cancer Cell. 2011;19(6):792–804. doi: 10.1016/j.ccr.2011.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bedolla R, Prihoda TJ, Kreisberg JI, Malik SN, Krishnegowda NK, Troyer DA, et al. Determining risk of biochemical recurrence in prostate cancer by immunohistochemical detection of PTEN expression and Akt activation. Clinical cancer research : an official journal of the American Association for Cancer Research. 2007;13(13):3860–7. doi: 10.1158/1078-0432.CCR-07-0091. [DOI] [PubMed] [Google Scholar]
- 7.Lotan TL, Wei W, Morais CL, Hawley ST, Fazli L, Hurtado-Coll A, et al. PTEN Loss as Determined by Clinical-grade Immunohistochemistry Assay Is Associated with Worse Recurrence-free Survival in Prostate Cancer. European urology focus. 2016;2(2):180–8. doi: 10.1016/j.euf.2015.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ferraldeschi R, Nava Rodrigues D, Riisnaes R, Miranda S, Figueiredo I, Rescigno P, et al. PTEN protein loss and clinical outcome from castration-resistant prostate cancer treated with abiraterone acetate. European urology. 2015;67(4):795–802. doi: 10.1016/j.eururo.2014.10.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Brubaker KD, Vessella RL, Brown LG, Corey E. Prostate cancer expression of runt-domain transcription factor Runx2, a key regulator of osteoblast differentiation and function. The Prostate. 2003;56(1):13–22. doi: 10.1002/pros.10233. [DOI] [PubMed] [Google Scholar]
- 10.Pratap J, Javed A, Languino LR, van Wijnen AJ, Stein JL, Stein GS, et al. The Runx2 osteogenic transcription factor regulates matrix metalloproteinase 9 in bone metastatic cancer cells and controls cell invasion. Molecular and cellular biology. 2005;25(19):8581–91. doi: 10.1128/MCB.25.19.8581-8591.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Blyth K, Vaillant F, Hanlon L, Mackay N, Bell M, Jenkins A, et al. Runx2 and MYC collaborate in lymphoma development by suppressing apoptotic and growth arrest pathways in vivo. Cancer Res. 2006;66(4):2195–201. doi: 10.1158/0008-5472.CAN-05-3558. [DOI] [PubMed] [Google Scholar]
- 12.Mullighan CG, Zhang J, Kasper LH, Lerach S, Payne-Turner D, Phillips LA, et al. CREBBP mutations in relapsed acute lymphoblastic leukaemia. Nature. 2011;471(7337):235–9. doi: 10.1038/nature09727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Yang S, Xu H, Yu S, Cao H, Fan J, Ge C, et al. Foxo1 mediates insulin-like growth factor 1 (IGF1)/insulin regulation of osteocalcin expression by antagonizing Runx2 in osteoblasts. J Biol Chem. 2011;286(21):19149–58. doi: 10.1074/jbcM110.197905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Komori T, Yagi H, Nomura S, Yamaguchi A, Sasaki K, Deguchi K, et al. Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell. 1997;89(5):755–64. doi: 10.1016/s0092-8674(00)80258-5. [DOI] [PubMed] [Google Scholar]
- 15.Thalji NM, Hagler MA, Zhang H, Casaclang-Verzosa G, Nair AA, Suri RM, et al. Nonbiased Molecular Screening Identifies Novel Molecular Regulators of Fibrogenic and Proliferative Signaling in Myxomatous Mitral Valve Disease. Circ Cardiovasc Genet. 2015;8(3):516–28. doi: 10.1161/CIRCGENETICS.114.000921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ding L, Chen S, Liu P, Pan Y, Zhong J, Regan KM, et al. CBP Loss Cooperates with PTEN Haploinsufficiency to Drive Prostate Cancer: Implications for Epigenetic Therapy. Cancer Res. 2014;74(7):2050–61. doi: 10.1158/0008-5472.CAN-13-1659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zhong J, Ding L, Bohrer LR, Pan Y, Liu P, Zhang J, et al. p300 acetyltransferase regulates androgen receptor degradation and PTEN-deficient prostate tumorigenesis. Cancer Res. 2014;74(6):1870–80. doi: 10.1158/0008-5472.CAN-13-2485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Mostaghel EA, Marck BT, Plymate SR, Vessella RL, Balk S, Matsumoto AM, et al. Resistance to CYP17A1 inhibition with abiraterone in castration-resistant prostate cancer: induction of steroidogenesis and androgen receptor splice variants. Clinical cancer research : an official journal of the American Association for Cancer Research. 2011;17(18):5913–25. doi: 10.1158/1078-0432.CCR-11-0728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Oury F, Sumara G, Sumara O, Ferron M, Chang H, Smith CE, et al. Endocrine regulation of male fertility by the skeleton. Cell. 2011;144(5):796–809. doi: 10.1016/j.cell.2011.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Merriman HL, van Wijnen AJ, Hiebert S, Bidwell JP, Fey E, Lian J, et al. The tissue-specific nuclear matrix protein, NMP-2, is a member of the AML/CBF/PEBP2/runt domain transcription factor family: interactions with the osteocalcin gene promoter. Biochemistry. 1995;34(40):13125–32. doi: 10.1021/bi00040a025. [DOI] [PubMed] [Google Scholar]
- 21.Geoffroy V, Ducy P, Karsenty G. A PEBP2 alpha/AML-1-related factor increases osteocalcin promoter activity through its binding to an osteoblast-specific cis-acting element. J Biol Chem. 1995;270(52):30973–9. doi: 10.1074/jbc.270.52.30973. [DOI] [PubMed] [Google Scholar]
- 22.Chua CW, Chiu YT, Yuen HF, Chan KW, Man K, Wang X, et al. Suppression of androgen-independent prostate cancer cell aggressiveness by FTY720: validating Runx2 as a potential antimetastatic drug screening platform. Clinical cancer research : an official journal of the American Association for Cancer Research. 2009;15(13):4322–35. doi: 10.1158/1078-0432.CCR-08-3157. [DOI] [PubMed] [Google Scholar]
- 23.Pi M, Quarles LD. GPRC6A regulates prostate cancer progression. Prostate. 2012;72(4):399–409. doi: 10.1002/pros.21442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Takata R, Akamatsu S, Kubo M, Takahashi A, Hosono N, Kawaguchi T, et al. Genome-wide association study identifies five new susceptibility loci for prostate cancer in the Japanese population. Nat Genet. 2010;42(9):751–4. doi: 10.1038/ng.635. [DOI] [PubMed] [Google Scholar]
- 25.Ge C, Zhao G, Li Y, Li H, Zhao X, Pannone G, et al. Role of Runx2 phosphorylation in prostate cancer and association with metastatic disease. Oncogene. 2016;35(3):366–76. doi: 10.1038/onc.2015.91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lim M, Zhong C, Yang S, Bell AM, Cohen MB, Roy-Burman P. Runx2 regulates survivin expression in prostate cancer cells. Lab Invest. 2010;90(2):222–33. doi: 10.1038/labinvest.2009.128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Chamouni A, Oury F. Reciprocal interaction between bone and gonads. Archives of biochemistry and biophysics. 2014;561:147–53. doi: 10.1016/j.abb.2014.06.016. [DOI] [PubMed] [Google Scholar]
- 28.Pi M, Chen L, Huang MZ, Zhu W, Ringhofer B, Luo J, et al. GPRC6A null mice exhibit osteopenia, feminization and metabolic syndrome. PLoS ONE. 2008;3(12):e3858. doi: 10.1371/journal.pone.0003858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Graham TR, Agrawal KC, Abdel-Mageed AB. Independent and cooperative roles of tumor necrosis factor-alpha, nuclear factor-kappaB, and bone morphogenetic protein-2 in regulation of metastasis and osteomimicry of prostate cancer cells and differentiation and mineralization of MC3T3-E1 osteoblast-like cells. Cancer Sci. 2010;101(1):103–11. doi: 10.1111/j.1349-7006.2009.01356.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Yeung F, Law WK, Yeh CH, Westendorf JJ, Zhang Y, Wang R, et al. Regulation of human osteocalcin promoter in hormone-independent human prostate cancer cells. J Biol Chem. 2002;277(4):2468–76. doi: 10.1074/jbc.M105947200. [DOI] [PubMed] [Google Scholar]
- 31.Chen Y, Chi P, Rockowitz S, Iaquinta PJ, Shamu T, Shukla S, et al. ETS factors reprogram the androgen receptor cistrome and prime prostate tumorigenesis in response to PTEN loss. Nat Med. 2013;19(8):1023–9. doi: 10.1038/nm.3216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ellwood-Yen K, Wongvipat J, Sawyers C. Transgenic mouse model for rapid pharmacodynamic evaluation of antiandrogens. Cancer Res. 2006;66(21):10513–6. doi: 10.1158/0008-5472.CAN-06-1397. [DOI] [PubMed] [Google Scholar]
- 33.Chun JY, Nadiminty N, Dutt S, Lou W, Yang JC, Kung HJ, et al. Interleukin-6 regulates androgen synthesis in prostate cancer cells. Clin Cancer Res. 2009;15(15):4815–22. doi: 10.1158/1078-0432.CCR-09-0640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Drissi H, Luc Q, Shakoori R, Chuva De Sousa Lopes S, Choi JY, Terry A, et al. Transcriptional autoregulation of the bone related CBFA1/RUNX2 gene. J Cell Physiol. 2000;184(3):341–50. doi: 10.1002/1097-4652(200009)184:3<341::AID-JCP8>3.0.CO;2-Z. [DOI] [PubMed] [Google Scholar]
- 35.Carver BS, Chapinski C, Wongvipat J, Hieronymus H, Chen Y, Chandarlapaty S, et al. Reciprocal feedback regulation of PI3K and androgen receptor signaling in PTEN-deficient prostate cancer. Cancer cell. 2011;19(5):575–86. doi: 10.1016/j.ccr.2011.04.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Jiao J, Wang S, Qiao R, Vivanco I, Watson PA, Sawyers CL, et al. Murine cell lines derived from Pten null prostate cancer show the critical role of PTEN in hormone refractory prostate cancer development. Cancer Res. 2007;67(13):6083–91. doi: 10.1158/0008-5472.CAN-06-4202. [DOI] [PubMed] [Google Scholar]
- 37.Krongrad A, Wilson CM, Wilson JD, Allman DR, McPhaul MJ. Androgen increases androgen receptor protein while decreasing receptor mRNA in LNCaP cells. Molecular and cellular endocrinology. 1991;76(1-3):79–88. doi: 10.1016/0303-7207(91)90262-q. [DOI] [PubMed] [Google Scholar]
- 38.Watson PA, Arora VK, Sawyers CL. Emerging mechanisms of resistance to androgen receptor inhibitors in prostate cancer. Nat Rev Cancer. 2015;15(12):701–11. doi: 10.1038/nrc4016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Fradet A, Sorel H, Depalle B, Serre CM, Farlay D, Turtoi A, et al. A new murine model of osteoblastic/osteolytic lesions from human androgen-resistant prostate cancer. PLoS ONE. 2013;8(9):e75092. doi: 10.1371/journal.pone.0075092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tuxhorn JA, Ayala GE, Smith MJ, Smith VC, Dang TD, Rowley DR. Reactive stroma in human prostate cancer: induction of myofibroblast phenotype and extracellular matrix remodeling. Clin Cancer Res. 2002;8(9):2912–23. [PubMed] [Google Scholar]
- 41.Barron DA, Rowley DR. The reactive stroma microenvironment and prostate cancer progression. Endocrine-related cancer. 2012;19(6):R187–204. doi: 10.1530/ERC-12-0085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Sharon Y, Alon L, Glanz S, Servais C, Erez N. Isolation of normal and cancer-associated fibroblasts from fresh tissues by Fluorescence Activated Cell Sorting (FACS) Journal of visualized experiments : JoVE. 2013;(71):e4425. doi: 10.3791/4425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.The Molecular Taxonomy of Primary Prostate Cancer. Cell. 2015;163(4):1011–25. doi: 10.1016/j.cell.2015.10.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Robinson D, Van Allen EM, Wu YM, Schultz N, Lonigro RJ, Mosquera JM, et al. Integrative clinical genomics of advanced prostate cancer. Cell. 2015;161(5):1215–28. doi: 10.1016/j.cell.2015.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Shen MM, Abate-Shen C. Pten inactivation and the emergence of androgen-independent prostate cancer. Cancer Res. 2007;67(14):6535–8. doi: 10.1158/0008-5472.CAN-07-1271. [DOI] [PubMed] [Google Scholar]
- 46.Nakashima K, Zhou X, Kunkel G, Zhang Z, Deng JM, Behringer RR, et al. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell. 2002;108(1):17–29. doi: 10.1016/s0092-8674(01)00622-5. [DOI] [PubMed] [Google Scholar]
- 47.Kumar MV, Jones EA, Grossmann ME, Blexrud MD, Tindall DJ. Identification and characterization of a suppressor element in the 5′-flanking region of the mouse androgen receptor gene. Nucleic Acids Res. 1994;22(18):3693–8. doi: 10.1093/nar/22.18.3693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Lai KP, Yamashita S, Huang CK, Yeh S, Chang C. Loss of stromal androgen receptor leads to suppressed prostate tumourigenesis via modulation of pro-inflammatory cytokines/chemokines. EMBO molecular medicine. 2012;4(8):791–807. doi: 10.1002/emmm.201101140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Qian BZ, Li J, Zhang H, Kitamura T, Zhang J, Campion LR, et al. CCL2 recruits inflammatory monocytes to facilitate breast-tumour metastasis. Nature. 2011;475(7355):222–5. doi: 10.1038/nature10138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Zhang XH, Jin X, Malladi S, Zou Y, Wen YH, Brogi E, et al. Selection of bone metastasis seeds by mesenchymal signals in the primary tumor stroma. Cell. 2013;154(5):1060–73. doi: 10.1016/j.cell.2013.07.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.