

Summary
A memory is unstable, making it susceptible to interference and disruption, after its acquisition [1–4]. The function or possible benefit of a memory being unstable at its acquisition is not well understood. Potentially, instability may be critical for the communication between recently acquired memories, which would allow learning in one task to be transferred to the other subsequent task [1, 5]. Learning may be transferred between any memories that are unstable, even between different types of memory. Here, we test the link between a memory being unstable, and the transfer of learning to a different type of memory task. We measured how learning in one task transferred to and so improved learning in a subsequent task. There was transfer from a motor skill to a word-list task, and, vice versa, from a word-list to a motor skill task. What was transferred was a high-level relationship between elements rather than knowledge of the individual elements themselves. Memory instability was correlated with subsequent transfer, suggesting that transfer was related to the instability of the memory. Using different methods, we stabilized the initial memory, preventing it from being susceptible to interference, and found that these methods consistently prevented transfer to the subsequent memory task. This suggests that the transfer of learning across diverse tasks is due to a high-level representation that can only be formed when a memory is unstable. Our work has identified an important function of memory instability.
Graphical abstract
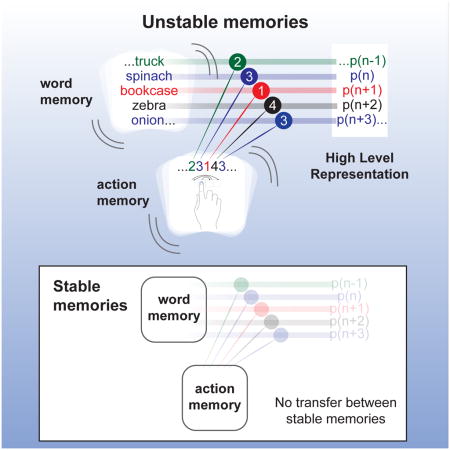
Results & Discussion
We tested the idea that a memory being unstable, and susceptible to interference from a subsequent memory, was necessary for learning to transfer across memory tasks. Learning transfer occurred when learning in the first task lead to enhanced learning in the subsequent novel task, and memory instability was measured as the impairment in performance between testing and subsequent retesting [1, 6]. Participants learnt, at 9am, one memory task, then another, and were retested 12-hrs later, at 9pm, on the initial memory task (please see Supplemental Information Section; Experimental Procedures). The memory tasks had either the same or different high-level structures. A motor sequence and a word-list can share a common structure by assigning each of the positions within the motor sequence (designated 1 to 4) to one of the four semantic categories in the word-list. For example, a specific vegetable can replace each position -1- within the motor sequence so that the first position -1- is replaced by turnip; while the second position -1- is replaced by spinach. Different semantic categories can be assigned to each of the other three positions, and each position replaced by a unique word from that category (for details; please see Supplemental Information Section; Experimental Procedures, and Figure S1). By assigning semantic categories to each of the four positions (1 to 4) we generated a word-list with the same temporal structure as a motor sequence.
Transfer between the different memory tasks
We first tested for the transfer of learning from a word-list to a motor skill task (Figure 1a). As training progressed across the three practice blocks the improvement in motor skill was significantly greater when the earlier word-list and subsequent motor sequence shared a common structure (repeated measures ANOVA, learning x group; F(2,52) = 4.2, p = 0.02; Figure 1b, and Figure S2). By the final block, participants motor skill and recall of the sequence was substantially greater when the word-list and motor sequence shared a common structure than when they had different structures (motor skill; unpaired t-test, 136±17ms vs. 73±12ms, mean±sem; t(26) = 2.96, p = 0.006; sequence recall; unpaired t-test, 4±1.2 vs. 0.6±0.4 items; t(26) = 2.5, p = 0.019; Figure 1b and Figure S3). There was no significant difference in total word recall (unpaired t-test; 10.8±0.4 vs. 10.8±0.3; t(26)<0.1, p>0.9), serial word recall (unpaired t-test; 7.5±0.8 vs. 6.9±1; t(26) = 0.471, p = 0.641) or in the initial motor skill between the two groups (unpaired t-test; 36±11ms vs. 33±10ms; t(26) = 0.171, p = 0.866). Learning two tasks with different structures may have impaired subsequent learning in the motor task; however, when the word-list had no consistent structure (none group; Figure 1b) the subsequent motor skill and sequence recall was still significantly less than when the two tasks shared the same structure (unpaired t-tests; 136±17ms vs. 64±9ms; t(24) = 3.51, p = 0.002; 4±1.2 items vs. 0.5±0.3 items; t(24) = 2.48; p = 0.02; Figure 1b and Figure S3). Thus, learning can transfer from a word-list to a motor skill when they share a common structure.
Figure 1. Learning transfer between different types of memory task.

(a.) In the first set of experiments, participants learnt and then recalled a list of words, acquired skill at performing a motor sequence during a short and then a long training block, had their motor skill tested, and 12-hrs later had their word recall retested. We used three groups to examine learning transfer between tasks: both tasks shared the same structure (same); had different structures (different); or the initial task, the word-list in the first set of experiments, had no consistent structure (none). (b.) The change of motor skill differed significantly across the groups during learning (repeated measures ANOVA; learning x group, F(4,74) = 3.54, p = 0.011). We found a greater increase in skill when the motor and word-list tasks had the same rather than different structures (repeated measures ANOVA; learning x group, F(2,52) = 4.16, p = 0.021) and a greater increase in skill when the tasks had the same structure than when the initial word-list task having no consistent structure (repeated measures ANOVA; learning x group, F(2,48) = 4.5, p = 0.016). There was no significant difference in the initial skill acquired across the three groups (ANOVA, F(2,37) = 0.017, p = 0.983). (c.) In the second set of experiments, participants acquired skill at performing a motor sequence during a short and then a long training block, had their motor skill tested, then learnt and recalled a list of words, and 12-hrs later had their motor skill retested. (d.) During learning, the change in serial word recall differed significantly across the three groups (repeated measures ANOVA; learning x group, F(6.2,156) = 2.95, p = 0.009), and as a consequence, there was a significant difference in serial recall at testing (ANOVA, F(2,39) = 4.046, p = 0.025). We found a greater increase in serial word recall when the word-list and motor task had the same structure rather than different structures (repeated measures ANOVA; learning x group, F(4,104) = 3.885, p = 0.006) and a greater increase when the tasks had the same structure than when the motor task had no consistent structure (repeated measures ANOVA; learning x group, F(3.1,104) = 3.9, p = 0.01). There was no significant difference in initial serial word recall across the three groups (ANOVA, F(2,39) = 1.09, p = 0.344). The bar and circle plots show a mean ± sem.
We then reversed the order of the tasks to test for learning transfer from a motor skill to a word-list task (Figure 1c). As learning progressed across the five iterations of the word-list, serial recall increased significantly more when the motor sequence and word-list had the same rather than a different structure (repeated measures ANOVAs, learning x group; serial recall, F(3.1,104) = 3.885, p = 0.011; although total recall did not differ (F(2.3,104) = 1.47, p = 0.235). At the final recall, there was significantly greater serial but not total recall of the word-list when the motor sequence and word-list shared a common structure than when they had different structures (unpaired t-tests; serial recall, 7.5±0.8 vs. 4.7±0.8; t(26) = 2.24, p = 0.034; total recall; 10.7±0.3 vs. 10±0.4; unpaired t-test, t(26) = 1.537, p = 0.136; Figure 1d). There was no significant difference in the motor skill acquired by participants (unpaired t-test; 78±7ms vs. 70±11ms; t(26) = 0.603, p = 0.552), or in the initial serial (unpaired t-test; 2.8±0.5 vs. 2±0.3; t(26) = 1.385, p = 0.178) and total word recall (unpaired t-test, 5.9±0.4 vs. 6.5±0.4; t(26) = 1, p = 0.322) between the two groups. The greater serial recall was dependent upon the two tasks sharing a common structure. The serial but not total recall decreased significantly when we replaced the motor sequence task with a task that still required motor responses but had no repeating sequential structure (unpaired t-tests; serial recall, 7.5±0.88 vs. 4.8±0.5; t(26) = 2.56, p = 0.016; total recall, 10.7±0.28 vs. 10.0±0.33; t(26) = 1.61, p = 0.12). Thus, learning on a motor sequence can transfer, enhancing the subsequent acquisition of word order, when the two tasks share the same structure.
Transfer and interference between the different memory tasks
When a memory is unstable, susceptible to interference, shown for example, by a decrease in its subsequent recall, it may exchange information with, and so allow learning to be transferred to, another memory task. We found that the transfer of learning from the word-list task to the motor skill task was correlated with a decrease in serial recall (F(1,12)=5.42, p = 0.038, R = 0.558; Figure 2a & Figure 2b; for further analysis see figure legend). By contrast, when there was no transfer of learning to the motor task, we found that motor skill was not correlated with a decrease in serial recall (F(1,12)<0.1, p = 0.994, R= 0.002; Figure 2b). Thus, the instability of a word-list memory, its susceptibility to interference, may be linked to learning transfer. We tested this idea further by preventing interference of the word-list task, and observing its effect on transfer to the motor skill task.
Figure 2. Learning transfer from a word-list memory task to a motor skill task, and its link to the interference of word recall.

(a) There was transfer from the word-list to the subsequent motor task when the tasks shared the same structure shown by the enhanced motor skill at testing compared to when the memory tasks had different structures (unpaired t-test, t(26) = 2.96, p = 0.006; bar plots display, mean ± sem). (b) The transfer of learning to the motor skill correlated with the decrease in serial recall (F(1,12) = 5.42, p =0.038, R = 0.558). This correlation remained even when the when the participant showing the greatest motor skill was removed from the analysis (F(1,11) = 6.2, p = 0.03, R = 0.6), and even without this participant there continued to be greater motor skill when the memory tasks had the same rather than different structures (unpaired t-test, t(25) = 2.9, p = 0.008). By contrast, when there was no learning transfer, when the two tasks had different structures, we found that the decrease in serial recall was not correlated with motor skill (F(1,12)<0.1, p = 0.994, R= 0.002). (c) Inserting a 2-hr interval between the memory tasks prevented interference of the word-list task. The decrease in serial recall was a significantly less than when the tasks had been performed in quick succession (unpaired t-test, t(26) = 2.7, p = 0.012), and there was no significant change in serial recall between testing and retesting (paired t-test, t(13) = 0.234, p = 0.818). We also modified the order of elements within the tasks to prevent interference of the serial recall. There was a significantly smaller decrease in serial recall than without these changes (unpaired t-test, t(24) = 2.37, p = 0.026), and there was no significant change in serial recall between testing and retesting (paired t-test, t(11) = 0.277, p = 0.787). Preventing interference, using either of these methods, also prevented learning transfer to the motor skill task. The enhancement of motor skill learning was significantly less with the insertion of the 2-hr delay (unpaired t-test; t(26) = 3.4, p = 0.002) or with the changes to the order of task elements (unpaired t-test; t(24) = 2.89, p = 0.008) than when the unmodified tasks had been performed in quick succession.
When a 2-hr interval was placed between the word-list and motor sequence tasks, there was no significant decrease in serial word recall (6.1±1 vs. 6.2±1.1 words; paired t-test; t(13) = 0.234, p = 0.818), and the impairment in serial word recall was significantly less than when the tasks had been learnt in quick succession (−1.78±0.36 vs. 0.1±0.6; unpaired t-test; t(26) = 2.7, p = 0.012; Figure 2c). Despite the 2-hr interval participants continued to be retested 12-hrs after initial learning (i.e., at 9pm). Preventing interference of word recall also prevented transfer to the motor skill task. Motor skill and sequence recall were both significantly less when the 2-hr interval was inserted than when the tasks were learnt in quick succession (unpaired t-test; 136±17ms vs. 72±7ms; t(26) = 3.41, p = 0.002; unpaired t-test; 3.9±1.2 items vs. 0.28±0.28 items; t(26) = 2.85, p = 0.008; Figure 2a). Initial serial recall did not differ significantly between the groups (7.5±0.8 vs. 6.1±1; unpaired t-test, t(26) = 1, p = 0.3). Thus, when the word-list memory was stable and no longer susceptible to interference there was no transfer to the motor sequence task.
To further confirm that diminished interference, rather than some other aspect of inserting the 2-hr interval, was responsible for preventing transfer, we used an additional technique to alter interference between the tasks. Subtle changes to the structure of the motor sequence modify the circuits supporting motor learning, from being overlapping with those critical to word-list learning, to being largely independent with less overlap between the circuits. We predicted these changes would diminish the interference between the different memory tasks ([7, 8]; for a review [9]; please also see Supplemental Information Section; Experimental Procedures, Interference between the memory tasks). As predicted, the changes prevented interference. We found no significant change in serial word recall (6.5±0.9 vs. 6.3±0.5; paired t-test, t(11) = 0.227, p = 0.787), and the decrease in serial recall was significantly greater when the tasks had not been modified (−1.78±0.4 vs. −0.16±0.6; unpaired t-test, t(24) = 2.38, p = 0.025; Figure 2c). Preventing interference also prevented the transfer to the motor task. The motor skill and sequence recall were substantially less for the modified tasks, which did not show interference of serial recall, than for the unmodified tasks that did show interference (unpaired t-test; 136±17ms vs. 68±15ms; t(24) = 3.05, p = 0.006; 3.9±1.2 items vs. 1.0±0.5 items; t(24) = 2.16, p = 0.04; Figure 2a & Figure 2c). At initial testing, serial word recall did not differ significantly between the modified and unmodified groups (7.5±0.8 vs. 6.5±0.9; unpaired t-test, t(24) = 0.857, p = 0.4). In sum, we used two different methods to prevent the word-list memory from being susceptible to interference. Despite being very different, both methods offer convergent findings, having the same effect of preventing transfer to the motor sequence task. The instability of the word-list memory was also correlated with the transfer to the motor learning task (see Figure 2b). Thus, multiple lines of evidence come together to demonstrate the importance of a word-list memory being unstable for learning transfer to the motor sequence task.
When we reversed the order of the memory tasks, we found a similar link between learning transfer and interference. We found that the transfer of learning to the word-list task was correlated with the change in motor skill (F(1,12)=6.43, p = 0.026, R = 0.590; Figure 3a and Figure 3b). By contrast, when there was no transfer of learning to the word-list task, we found that the serial word recall was not correlated with the change in motor skill (F(1,12) = 0.12, p = 0.736, R = 0.099; Figure 3b). Thus, the instability of a motor skill memory, its susceptibility to interference, may be linked to learning transfer. We tested for a possible link between the instability of a motor skill memory, and the transfer of learning to a word-list memory task.
Figure 3. Learning transfer from a motor skill task to a word-list memory task, and its link to the interference of the motor skill.

(a) There was transfer from the motor task to the subsequent word-list when the tasks shared the same structure shown by the enhanced serial recall at testing compared to when the memory tasks had different structures (unpaired t-test, t(26) = 2.244, p = 0.034; bar plots display, mean ± sem). (b) The transfer of learning to the word-list, shown by enhanced serial word recall, was correlated with the change in motor skill between testing and retesting (F(1,12)=6.43, p = 0.026, R = 0.590). By contrast, when there was no learning transfer, when the two tasks had different structures, we found that the serial recall was not correlated with the change in motor skill (F(1,12) = 0.12, p = 0.736, R = 0.099). (c) Inserting a 2-hr interval between the memory tasks, or modifying the order of task elements within both tasks prevented interference, which allowed the development of off-line improvements between testing and retesting (both paired t-tests; t(13)> 2.5,p< 0.025). Preventing interference, using either of these methods, also prevented learning transfer to the word-list. Serial word recall was significantly less with the insertion of the 2-hr interval (unpaired t-test; t(26) = 2.73, p = 0.011) or with the changes to the order of task elements (unpaired t-test; t(26) = 2.254, p = 0.033) than when the unmodified tasks had been performed in quick succession.
A motor skill memory can show off-line improvements that develop between testing and subsequent retesting 8–12hrs later; however, because the motor skill memory is still unstable these improvements can be disrupted and impaired due to interference ([10–15]; for reviews [1–4]). We sought to prevent interference of the motor skill task, allowing the development of offline improvements, by inserting a 2-hr interval between the tasks, or modifying the order of elements within the tasks. Both of these approaches had been used successfully to prevent interference in the earlier experiments. Introducing a 2-hr interval between the tasks allowed the development of motor skill improvements between testing and subsequent retesting (paired t-test; 72±9ms vs. 103±9ms; t(13) = 3.835, p = 0.002; [5, 13]), which were significantly greater than when the tasks were learnt in quick succession (unpaired t-test; −2±9ms vs. 30±8ms; t(26) = 2.65, p = 0.013; Figure 3c). At initial testing, motor skill did not differ significantly between the groups (unpaired t-test; 78±8ms vs. 72±9ms; t(26) = 0.517, p = 0.609). Preventing interference of the off-line improvements also prevented learning transfer to the word-list task. The serial word recall was significantly smaller when there was a 2-hr interval than when the tasks were learnt in quick succession (unpaired t-test; 7.5±0.88 vs. 4.7±0.81; t(26) = 2.254, p = 0.033; Figure 3a). Similarly, modifying the order of elements within the tasks prevented interference as it had done in the earlier experiments. Off-line improvements developed between testing and retesting (paired t-test; 72±7.5 vs. 102±12.1; t(13) = 2.626, p = 0.021), and these improvements were significantly greater than in unmodified tasks (unpaired t-test; −2±9ms vs. 29±11ms; t(26) = 2.165, p = 0.04; Figure 3c). Preventing interference of the off-line motor skill improvements also prevented transfer to the word-list task. Serial word recall was significantly greater when the tasks were unmodified rather than modified (unpaired t-test, 7.5±0.88 vs. 4.8±0.53; t(26) = 2.565; p = 0.016; Figure 3c). The initial motor skill did not differ significantly between the modified and unmodified tasks (unpaired t-test, 78±7ms vs. 72±7ms; t(26) = 0.587, p = 0.562). Using two different methods, we prevented the motor skill memory from being susceptible to interference, allowing the development of off-line improvements, and both those methods had the same consistent effect of preventing transfer to the word-list task. The interference of the off-line improvements was also correlated with the subsequent transfer to the word-list learning task (see Figure 3b). In sum, evidence from across all the experiments converges to demonstrate that only when a memory is unstable can learning be transferred to another memory task.
Discussion
By being unstable a newly acquired memory is susceptible to interference, which can impair its subsequent retention. What function this instability might serve has remained poorly understood. Here we show that: (1) a memory must be unstable for learning to transfer to another memory task and (2) that the information transferred is of the high-level or abstract properties of a memory task. We find that transfer from a memory task is correlated with its instability, and that transfer is prevented when a memory is stabilized. Thus, an unstable memory is in privileged state: only when unstable can a memory communicate with and transfer knowledge to affect the acquisition of a subsequent memory. It is the relationship between elements rather, than knowledge of the individual elements themselves (i.e., words vs. actions) that is transferred. These findings suggest that the knowledge transferred is high-level or abstract, which allows learning to be transferred between different types of memory task (i.e., declarative vs. motor memory tasks). Our work shows that an unstable memory is a key component of the mechanism for learning transfer. An unstable memory prevents learning from being rigidly linked to one task; instead, it allows learning to be applied flexibly.
Learning has been shown before to transfer from one task to another, from one hand to another (i.e., intermanual transfer), and from eye to hand (i.e., oculomanual transfer; [11, 16–21]). Yet, these examples of transfer all occur within the same type of memory task. By contrast, here we show that transfer also occurs between different types of memory tasks. The transfer of learning between the word-list and motor skill memory task only occurred when they shared a common sequential structure. Consistent with the importance of the sequence structure, we found that it was specifically the sequential aspect of the tasks that were transferred. For example, the recall of words in the correct sequential order was transferred between tasks; whereas, total recall was not. Thus, during learning it is not just the specific words or actions that are encoded, but also their relationship with the other elements within the sequence. Our work establishes a mechanism that makes it possible for learning to transfer between very different types of memory task. Specifically, using the shared high-level properties of the relationship amongst elements makes transfer possible even when those elements are very different (i.e. words vs. actions).
A memory must be unstable for learning to be transferred to a subsequent memory task. We found a correlation between the susceptibility of a memory to interference, a measure of its instability, and the learning transferred to the other memory task (Figures 2b & 3b). Complementing these findings, we found that preventing a memory from being susceptible to interference also prevented the transfer of learning to the subsequent memory task. The techniques we used to prevent interference differed in many ways, however, they both prevented interference, and they both prevented learning transfer (see Supplemental Information Section; Experimental Procedures, Interference between the memory tasks). Thus, converging lines of evidence demonstrate that a memory must be unstable for learning to be transferred to the subsequent memory task.
Unstable memories are susceptible to interference, which can impair their subsequent retention; yet, instability also gives memories an opportunity to interact and communicate with other memories leading to learning transfer. We show transfer from an initial unstable memory to a subsequent memory task. We also envisage, at least in principle, retrograde transfer occurring from that subsequent memory task to the initial memory task. Yet, substantial learning has already occurred in the initial memory task, and so any benefit from transfer is likely to be negligible. Overall, learning transfer may come at the cost of memory instability; although the expression of that transfer can be affected by other factors such as prior learning.
Instability may allow the interaction, and subsequent transfer of many aspects of a memory, from the high-level features, which we have focused on here, to potentially the low-level features (i.e., specific items). The transfer of low-level features; such as a shared sequence of movements, occurs immediately after an initial memory has been acquired when it is likely to still be unstable [11, 16–21]. Yet, when a memory is stabilized, through consolidation, the subsequent transfer is substantially reduced [22]. Thus, instability is important not only for high-level but also probably for the transfer of low-level information too.
Conclusion
An unstable memory provides a window of opportunity for communication between memories, leading to the construction of a high-level or abstract memory representation, which allows the transfer of knowledge between memory tasks. A link between memory instability and the creation of high-level representation could explain the similarity in the brain areas critical to memory instability and the creation of memory schema (i.e., a framework of knowledge that contains the features common across different learning episodes or tasks; [5, 23–25]). The representation is not of the individual elements or content of the memory (i.e., the words vs. actions); instead, it is of the abstract relationship between the elements. Knowledge is not constrained within independent systems. Instead, knowledge can be organized in the human brain at a higher, more abstract level than has previously been suspected, which allows the transfer of learning between memories for words and actions. The transfer of learning across diverse tasks is due to a high-level representation that can only be formed when a memory is unstable. In sum, we have identified an important functional contribution of memory instability, provided insight into the mechanism of learning transfer, and given a novel perspective on memory organization.
Supplementary Material
Acknowledgments
We are grateful to Daniel Wolpert for his thoughtful comments on aspects of the analysis of this work, and to the National Science Foundation (NSF; Division of Behavioral and Cognitive Sciences; BCS, 0921177; EMR) for supporting this work.
Footnotes
Author Contributions
Conceptualization, E.M.R.; Methodology, E.M.R.; Software, N.M. and E.M.R.; Formal Analysis, N.M. and E.M.R; Investigation, N.M. and E.M.R.; Writing – Original Draft, E.M.R.; Writing – Review & Editing, N.M. & E.M.R; Funding Acquisition, E.M.R.; Supervision, E.M.R.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Robertson EM. New insights in human memory interference and consolidation. Current biology : CB. 2012;22:R66–71. doi: 10.1016/j.cub.2011.11.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Robertson EM, Pascual-Leone A, Miall RC. Current concepts in procedural consolidation. Nat Rev Neurosci. 2004;5:576–582. doi: 10.1038/nrn1426. [DOI] [PubMed] [Google Scholar]
- 3.Walker MP. A refined model of sleep and the time course of memory formation. The Behavioral and brain sciences. 2005;28:51–64. doi: 10.1017/s0140525x05000026. discussion 64–104. [DOI] [PubMed] [Google Scholar]
- 4.Dayan E, Cohen LG. Neuroplasticity subserving motor skill learning. Neuron. 2011;72:443–454. doi: 10.1016/j.neuron.2011.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cohen DA, Robertson EM. Preventing interference between different memory tasks. Nature neuroscience. 2011;14:953–955. doi: 10.1038/nn.2840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Seidler RD. Multiple motor learning experiences enhance motor adaptability. J Cogn Neurosci. 2004;16:65–73. doi: 10.1162/089892904322755566. [DOI] [PubMed] [Google Scholar]
- 7.Curran T. Higher order associative learning in amnesia: Evidence from the serial reaction time task. Journal of Cognitive Neuroscience. 1997;9:522–533. doi: 10.1162/jocn.1997.9.4.522. [DOI] [PubMed] [Google Scholar]
- 8.Schendan HE, Searl MM, Melrose RJ, Stern CE. An FMRI study of the role of the medial temporal lobe in implicit and explicit sequence learning. Neuron. 2003;37:1013–1025. doi: 10.1016/s0896-6273(03)00123-5. [DOI] [PubMed] [Google Scholar]
- 9.Robertson EM. The Serial Reaction Time Task: Implicit Motor Skill Learning? J Neurosci. 2007;27:10073–10075. doi: 10.1523/JNEUROSCI.2747-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Robertson EM, Pascual-Leone A, Press DZ. Awareness modifies the skill-learning benefits of sleep. Current Biology. 2004;14:208–212. doi: 10.1016/j.cub.2004.01.027. [DOI] [PubMed] [Google Scholar]
- 11.Cohen DA, Pascual-Leone A, Press DZ, Robertson EM. Off-line learning of motor skill memory: A double dissociation of goal and movement. PNAS. 2005;102:18237–18241. doi: 10.1073/pnas.0506072102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Spencer RM, Sunm M, Ivry RB. Sleep-dependent consolidation of contextual learning. Current biology : CB. 2006;16:1001–1005. doi: 10.1016/j.cub.2006.03.094. [DOI] [PubMed] [Google Scholar]
- 13.Brown RM, Robertson EM. Off-Line Processing: Reciprocal Interactions between Declarative and Procedural Memories. J Neurosci. 2007;27:10468–10475. doi: 10.1523/JNEUROSCI.2799-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Song S, Howard JH, Jr, Howard DV. Sleep does not benefit probabilistic motor sequence learning. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2007;27:12475–12483. doi: 10.1523/JNEUROSCI.2062-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Abe M, Schambra H, Wassermann EM, Luckenbaugh D, Schweighofer N, Cohen LG. Reward improves long-term retention of a motor memory through induction of offline memory gains. Current biology : CB. 2011;21:557–562. doi: 10.1016/j.cub.2011.02.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bapi RS, Doya K, Harner AM. Evidence for effector independent and dependent representations and their differential time course of acquisition during motor sequence learning. Exp Brain Res. 2000;132:149–162. doi: 10.1007/s002219900332. [DOI] [PubMed] [Google Scholar]
- 17.Japikse K, Negash S, Howard J, Howard D. Intermanual transfer of procedural learning after extended practice of probablistic sequences. Experimental Brain Research. 2003;148:38–49. doi: 10.1007/s00221-002-1264-9. [DOI] [PubMed] [Google Scholar]
- 18.Verwey W, Wright D. Effector-independent and effector-dependent learning in the discrete sequence production task. Psychological Research. 2004;68:64–70. doi: 10.1007/s00426-003-0144-7. [DOI] [PubMed] [Google Scholar]
- 19.Verwey W, Clegg B. Effector dependent sequence learning in the serial RT task. Psychological Research. 2005;69:242–251. doi: 10.1007/s00426-004-0181-x. [DOI] [PubMed] [Google Scholar]
- 20.Perez MA, Wise SP, Willingham DT, Cohen LG. Neurophysiological mechanisms involved in transfer of procedural knowledge. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2007;27:1045–1053. doi: 10.1523/JNEUROSCI.4128-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Perez MA, Tanaka S, Wise SP, Sadato N, Tanabe HC, Willingham DT, Cohen LG. Neural substrates of intermanual transfer of a newly acquired motor skill. Current biology : CB. 2007;17:1896–1902. doi: 10.1016/j.cub.2007.09.058. [DOI] [PubMed] [Google Scholar]
- 22.Stockinger C, Thurer B, Focke A, Stein T. Intermanual transfer characteristics of dynamic learning: direction, coordinate frame, and consolidation of interlimb generalization. Journal of neurophysiology, jn 00727 02015. 2015 doi: 10.1152/jn.00727.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Costanzi M, Saraulli D, Rossi-Arnaud C, Aceti M, Cestari V. Memory impairment induced by an interfering task is reverted by pre-frontal cortex lesions: A possible role for an inhibitory process in memory suppression in mice. Neuroscience. 2009;158:503–513. doi: 10.1016/j.neuroscience.2008.08.026. [DOI] [PubMed] [Google Scholar]
- 24.Tse D, Langston RF, Kakeyama M, Bethus I, Spooner PA, Wood ER, Witter MP, Morris RG. Schemas and memory consolidation. Science. 2007;316:76–82. doi: 10.1126/science.1135935. [DOI] [PubMed] [Google Scholar]
- 25.Tse D, Takeuchi T, Kakeyama M, Kajii Y, Okuno H, Tohyama C, Bito H, Morris RG. Schema-dependent gene activation and memory encoding in neocortex. Science. 2011;333:891–895. doi: 10.1126/science.1205274. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.