

Abstract
Important functions of cornea in the eye include protecting the structures inside the eye, contributing to the refractive power of the eye, and focusing light rays on the retina with minimum scatter and optical degradation. Considerable advances have taken place in understanding the organization of collagen in the corneal stroma and its clinical significance. In this review, the structure and function of various components of cornea and ocular surface are presented.
Keywords: Collagen, cornea, Descemet's membrane, endothelium, epithelium, stroma
Cornea and sclera constitute the outer covering or coat of the eyeball. The main purpose of this coat is to protect structures inside the eye. The cornea is a transparent avascular tissue that acts as a structural barrier and protects the eye against infections.[1] Along with the tear film, it provides proper anterior refractive surface for the eye. Cornea contributes to two-third of the refractive power of the eye.
The cornea is horizontally oval, measuring 11–12 mm horizontally and 9–11 mm vertically. The corneal horizontal diameter (white to white) using ORBSCAN II system has revealed average corneal diameter as 11.71 ± 0.42 mm. The average corneal diameter was 11.77 ± 0.37 in males compared to 11.64 ± 0.47 in females. The corneal diameter ranged from 11.04–12.50 in males and 10.7–12.58 in females.[2] The limbus is widest in superior and inferior cornea. Cornea is convex and aspheric. The anterior curvature is 7.8 mm and posterior curvature is about 6.5 mm. Cornea contributes to about 40–44 D of refractive power and accounts for approximately 70% of total refraction. The refractive index of cornea is 1.376. There is a gradual increase in thickness from central cornea to the periphery[3] [Fig. 1]. Alteration in tissue thickness is due to increase in the amount of collagen in the peripheral stroma. With different methods of evaluation, the central corneal thickness in normal eyes is found to range from 551 to 565 μ and the peripheral corneal thickness from 612 to 640 μ.[4] The corneal thickness is found to decrease with age. Anterior corneal stromal rigidity appears to be particularly important in maintaining the corneal curvature. Anterior curvature resists changes to stromal hydration much more than posterior stroma.[1]
Figure 1.
Picture of cornea showing central and peripheral cornea
The cornea is made up of cellular and acellular components. The cellular components include the epithelial cells, keratocytes, and endothelial cells. The acellular component includes collagen and glycosaminoglycans. The epithelial cells are derived from epidermal ectoderm. The keratocyte and endothelial cells are derived from neural crest. The corneal layers include epithelium, Bowman's layer, stroma, Descemet's membrane, and endothelium [Fig. 2]. Recently, a layer of cornea which is well defined, acellular in pre-Descemet's cornea is getting attention with the development of lamellar surgeries.[5] Table 1 summarizes all layers of cornea with their functions.
Figure 2.
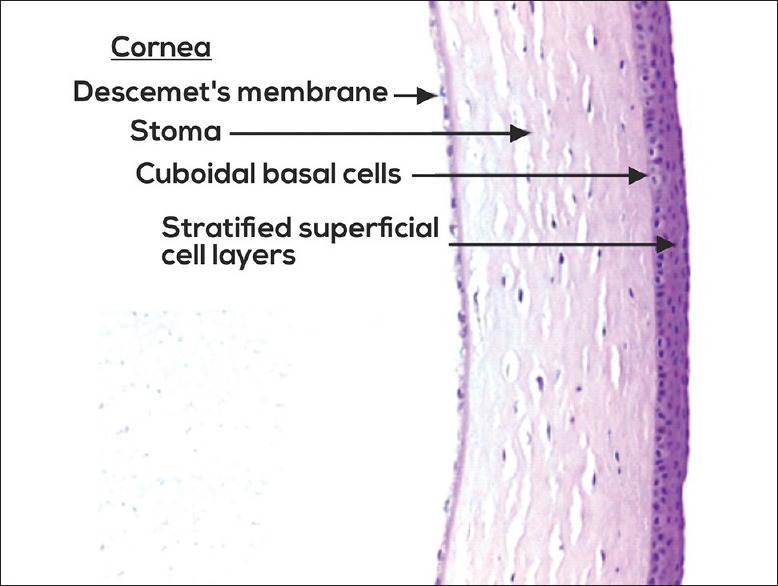
Histopathology of cornea showing corneal epithelium, stroma, and Descemet's membrane
Table 1.
Corneal layers with their functions

The corneal epithelium is composed fairly uniformly of 5–7 layers of cells [Fig. 3]. It is about 50 μ in thickness. The epithelium is uniform to provide a smooth regular surface and is made up of nonkeratinized stratified squamous epithelium. The epithelium is derived from surface ectoderm between 5 and 6 weeks of gestation. The epithelium and the overlying tear film have symbiotic relationship. The mucin layer of the tear film which is in the direct contact with corneal epithelium is produced by the conjunctival goblet cells. It interacts closely with the corneal epithelial cells' glycocalyx to allow hydrophilic spreading of the tear film with each eyelid blink.
Figure 3.
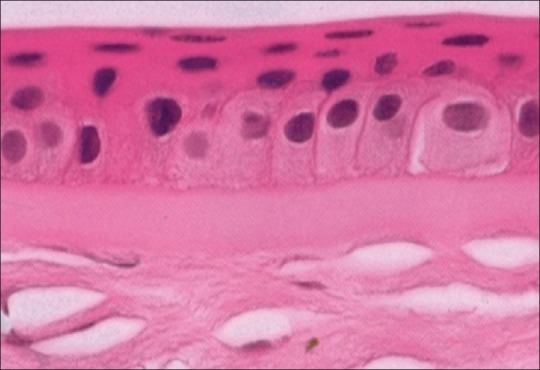
Histopathology of corneal epithelium and Bowman's membrane
Cornea epithelial cells have a lifespan of 7 to 10 days undergoing involution, apoptosis, and desquamation. The presence of high concentrations of intracytoplasmic enzyme crystalline, like in lens epithelial cells, may play an important role in maintaining optical transparency. The epithelium is 5–6 layers structure with three types of cells: superficial cells, wing cells, and the basal cells. The superficial cells are 2–3 layers made up of flat polygonal cells. They have a diameter of 40–60 μ with a thickness of about 2–6 μ. The microvilli on the surface increase the surface area. The epithelial layer is made up of large dark cells and small light cells. Desmosomes form the tight junction in between the superficial cells. Thawing cells are 2–3 layered and are named as they have wing-like shape. They express 64-k dalton keratins. Basal cells are single layer of the epithelium which is cuboidal or columnar. They have abundant organelles and they are active mitotically.
The surface cells maintain tight junction complexes between the neighbors which prohibit tears from entering the intercellular spaces. The deepest cell layer of the epithelium is the basal layer, which compromises the single cell layer of epithelium approximately 20 μ tall. Besides the stem cells and transient amplifying cells, basal cells are the only corneal epithelial cells capable of mitosis. They are the source of wing and superficial cells.
Desmosomes are present along the lateral membranes of all epithelial cells, to keep the epithelial cells adherent to each other. The basal cells are attached to the underlying basement membrane by a hemidesmosomal system. The strong attachment prevents the epithelium getting separated from the underlying layers. Abnormality of this attachment results in corneal erosions and nonhealing epithelial defects.
Tight junctions are present only in the lateral wall of apical cells of epithelium, providing a permeability barrier between the cells at the most superficial level.[3] Adherens junctions are present along the lateral membrane of only the apical cells of the epithelium. They maintain cellular adherence in the region of tight junctions. Gap junctions are permeable channels on the lateral aspects of all epithelial cells, allowing the diffusion of small molecules.
The basement membrane of epithelial cells is 40–60 nm in thickness and is made up of Type IV collagen and laminin secreted by basal cells. The basement membrane is made up of lamina lucida and lamina densa. From the basal epithelial cells, anchoring fibrils pass through the basement membrane and end as anchoring plaques. The anchoring fibrils are made up of Type VII collagen and anchoring plaque is made up of Type I collagen [Fig. 4]. If basement membrane is damaged, fibronectin levels increases and the process of healing can take up to 6 weeks.
Figure 4.
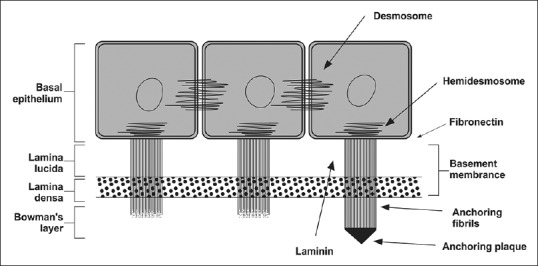
Diagram depicting the junctional complexes of the corneal epithelium
There are differences between epithelium of central and peripheral cornea. In the central cornea, the epithelium has 5–7 layers. The basal cells are columnar. There are no melanocytes or Langerhans cells. There is a smooth basal cell layer with keratan sulfate, and there are no lymphatics. In the peripheral cornea, the epithelium is 7–10 layered. The basal cells are cuboidal. There are melanocytes and Langerhans cells. There are undulating extensions of basal layer. There is no keratan sulfate and there are no lymphatics.
Thoft and Friend proposed the XYZ hypothesis of central epithelial maintenance.[6] X component denotes the basal cell proliferation. The Y component migration of cells from peripheral to central cornea and Z component denotes the loss of cells from the surface. The cells in the limbus are in two compartments. The stem cells and transient amplifying cells are in the proliferative compartment. The postmitotic cells and terminally differentiated cells are in nonproliferative compartment. The palisades of Vogt are undulations in basement membrane which provide increased vascularity, increased surface area for attachment, and also for provide protection to stem cells.
Bowman's membrane is condensation of collagen and proteoglycans. It is a 12 μ structure and is made up of Type I and V collagen as well as proteoglycans. It has no regenerative ability. Bowman's membrane lies just anterior to stroma and is not a true membrane. It is acellular condensate of the most anterior portion of the stroma. This smooth layer helps the cornea maintains its shape. When injured, this layer does not regenerate and may result in a scar.
The corneal stroma forms the bulk of the structural framework of the cornea and comprises approximately 80%–85% of its thickness. Embryologically, it is the result of second wave of neural crest migration that occurs in the 7th week of gestation, after establishment of the primitive endothelium. The stroma is characteristically transparent which is the result of precise organization of stromal fibers and extracellular matrix (ECM). The collagen within corneal fibrils is predominantly Type I. Type VI collagen and Type XII collagen are also found in stroma.[7]
The collagen fibers are arranged in parallel bundles called fibrils. These fibrils are packed in layers or lamellae. The stroma of the human eye contains 200–250 distinct lamellae, each layer arranged at right angles relative to fibers in adjacent lamella. The collagen in the stroma is laid down within lamellae. These structures are of the variable thickness, in humans up to 0.2 mm broad and 2 μm thick. At the center of the human cornea, there is approximately 200 lamellar thickness. The packing density is higher in the anterior lamellae than in the posterior stroma. These anterior lamellae are highly interwoven and most appear to insert into the Bowman's layer.[8] The mid-stromal lamellae are also highly interlaced. The posterior lamellae in the central cornea are more hydrated. The posterior stromal can swell easily whereas the more interwoven cannot.
Cornea lamellae in the deepest stroma have preferred directions which appear to close to inferior superior and nasal-temporal directions. X-rays scattering also has suggested a network of collagen lamella that enters the cornea close to the inferior, superior, nasal, and temporal which probably originate in the adjacent sclera. Collagen fibrils in the prepupillary appear to be more closely packed than in the peripheral cornea.[9] High packing density of stress-bearing collagen fibrils in the prepupillary cornea seems to be necessary for maintaining corneal strength and hence curvature in a region of reduced corneal thickness.
Collagen fibrils within the cornea are narrower than in many other connecting tissues, and there is an important factor for transparency, which is a function of diameter. There are about 300–400 triple-helical molecules within the cross section of the fibril. Ultrastructure within the organization of the lamellar appears to vary based on the depth within the stroma. Deeper layers are more strictly organized than superficial layers. This accounts for care of surgical dissection in a particular plane as one approaches the posterior depths of the corneal stroma.[1] During air injection into stroma, it is a common observation that part of the posterior stroma often adheres to the Descemet's membrane and there is a natural cleavage plane in the stroma almost 10 μ alone Descemet's membrane.[5]
A high refractive index and symmetric curvature are essential for optimal refractive power and minimum astigmatism of the cornea.[10] The architecture of the most anterior part (100–120 μ) of the stroma prevents changes in the morphology of the stromal region even after extreme swelling. The smooth curvature at the anterior side is not affected in conditions with increased corneal hydration. As the diameter of the cornea hardly changes, swelling is compensated by large undulations in Descemet's membrane at the posterior side. The corneal swelling behavior shows a gradual decrease in swelling from posterior to anterior side; this seems to be related to the organization of collagen lamellae and to the presence of different types of proteoglycan. In the posterior part keratan sulfate, a more hydrophilic proteoglycan is prevalent, whereas in the anterior part dermatan sulfate, a much less hydrophilic proteoglycan is prevalent. There is also an elastic fiber network within the stroma. These elastic fibers present throughout the stromal depth, are concentrated below the posterior-most keratocyte layer.
Cornea stroma is made up of keratocytes and ECM. The ECM is composed of collagens (Type I, III, V, VI) and glycosaminoglycans. The glycosaminoglycans constitute keratan sulfate, chondroitin sulfate, and dermatan sulfate. The corneal stroma has keratocytes and about 300 collagen lamellae which are regularly arranged. The glycosaminoglycans are predominantly keratan sulfate. Less contribution is by dermatan sulfate and chondroitin sulfate. Hyaluronan is seen in infancy.
Keratocytes are the major cell type of stroma. They are involved in maintaining the ECM environment. They are able to synthesize collagen molecules and glycosaminoglycans, while also creating matrix metalloproteinases (MMPs), all important in maintaining stromal homeostasis. Most of the keratocytes reside in the anterior stroma.
The Descemet's membrane is a 7 μ structure. It is made up of Type IV collagen and laminin. Descemet's membrane begins in utero at the 8-week stage. Endothelial cells continuously secrete Descemet's membrane. The anterior 3 μ secreted before birth has a distinctive banded appearance. Descemet's membrane produced after birth is unbanded and has an amorphous ultrastructural texture. Descemet's membrane can be up to 10 μ in thickness with age. The Descemet's membrane is elastic and curls on getting severed.
The endothelium is a single layer, 5 μ thick structure [Fig. 5]. The cells are hexagonal and are metabolically active. There is an endothelial pump which regulates water content. Endothelium is a monolayer which appears as a honeycomb-like mosaic when viewed from the posterior side. In early embryogenesis, the posterior surface is lined with a neural crest-derived monolayer of orderly arranged cuboidal cells. The individual cells continue to flatten over time and stabilize at approximately 4 μ in thickness is adulthood. Adjacent cells share extensive lateral interdigitations and possess gap and tight junctions along the lateral borders. The lateral membrane contains the high density of Na+ K+ ATPase pump sites. The two most important ion transport systems are the membrane-bound Na+ K+ ATPase and the intracellular carbonic anhydrase pathway. Activity in both these pathways produces the net flux of ions from the stroma to the aqueous humor. The basal surface of the endothelium contains numerous hemidesmosomes that promote adhesion to Descemet's membrane.
Figure 5.
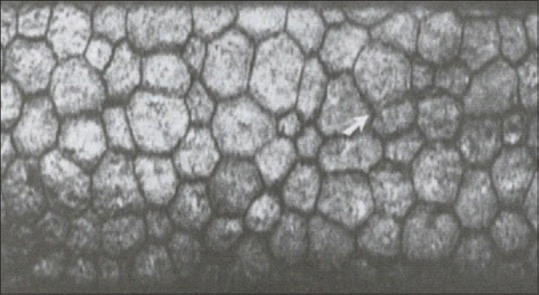
Endothelial cell layer on the specular microscopy
Immediately anterior to the flattened layer of endothelial cell is a discontinuous homogeneous acellular layer Descemet's membrane. At birth, the endothelial layer is approximately 10 μ thick. Descemet's membrane becomes continuous and uniform, fusing peripheral with the trabecular beams. The fusion site, known as Schwalbe line, is a gonioscopic landmark that defines the end of Descemet's membrane and the start of trabecular meshwork.
Endothelial cell density continues to change throughout life, from second to eight decades of life. The cell density declines from 3000 to 4000 cells/mm2 to around 2600 cells/mm2 of hexagonal cells decline from approximately from 75% to 60%. Endothelial cell density at birth is 3500 cells/mm2. Human central endothelial cell density decreases at an average of approximately 0.6% per year in normal corneas throughout adult life, into gradual increase in polymegathism and pleomorphism.[11] Endothelial cells do not regenerate in adults.[12]
A deep corneal layer has been getting attention recently. It is a well-defined, acellular, strong layer in the pre-Descemet's cornea.[5] This layer is found to be 5–8 lamellae of collagen fibers ranging from 6 to 15 μ in thickness. There are no keratocytes in this layer, and it is impervious to air. In donor eyes from eye banks, the bubble can be inflated to pressure about 700 mmHg before it ruptures. It is also found in children.
The cornea is one of the most heavily innervated and most sensitive tissues in the body. The sensation is derived from the nasociliary branch of the first division of (ophthalmic) of the trigeminal nerve. Thick and straight stromal nerve trunks extend laterally and anteriorly to give rise to plexiform arrangements of progressively thin nerve fibers at several levels within the stroma.[13] The nerve fibers perforate Bowman's layer and eventually form a dense nerve plexus just beneath the basal epithelial cell layer characterized by tortuous, thin beaded nerve fibers interconnected by numerous nerve elements. The cornea also contains autonomic sympathetic nerve fibers.
Normal human cornea is avascular. The aqueous humor is the main source of nutrients to the cornea. Blood supply is by tiny vessels at the outer edge of the cornea as well as components supplied by end branches of the facial and ophthalmic arteries through the aqueous humor and the tear film.
Tear film
The minute surface irregularities of the corneal surface epithelium are masked by a smooth and regular overlying tear film. The tear film is the first layer of the cornea with which light comes into contact. In produces lubrication and hydration to the ocular surface. It is also a source of oxygen, immunoglobulins, lysozymes, lactoferrin, and α- and β-defensins. The tear film is traditionally told to be composed of three distinct layers. The most superficial layer is composed of oily secretions of meibomian glands. Because oil is less dense than water, the secretions float to the top of the tear film to form a smooth refractive surface. This oily layer also provides an important barrier against the evaporation of the tear film. The aqueous middle layer of the tear film lies immediately beneath the oily layer. It is produced from the secretion of the lacrimal gland which is located in the superior lateral orbit. The aqueous is secreted onto the ocular surface from ducts in the superior fornix. There are also numerous scattered accessory lacrimal glands embedded within the conjunctiva stroma which contribute to this aqueous layer. The innermost mucin layer is produced primarily by the conjunctival goblet cells although the epithelial cells of the cornea and the conjunctiva also contribute. Current understanding is that all the three layers are in the form of a gel on the ocular surface. Conjunctival goblet cells seem to be important in performing the functions of debris removal and immune surveillance.[14]
Ocular surface
Thoft and Friend first suggested that cornea, conjunctiva, lacrimal glands, and lids work as an integrated unit called ocular surface.[15] The anatomical ocular surface is composed of the mucosa that lines the globe and palpebral surfaces, the corneoscleral limbus, the corneal epithelium, and the tear film.[16]
Sclera, in contrast to cornea, has collagen fibrils which are more haphazard and random. The bulk of stroma is mostly acellular, except for outer episcleral layer.
Conjunctiva extends from the limbus to the fornices, forming a fold at the fornices of the eyelid and then extends back around onto the posterior surface of the eyelids. Plica semilunaris is a crescent-shaped fold of conjunctiva medially with thickened stroma. The caruncle is located medial to the plica and marks the most medial aspect of the interpalpebral fissure. The bulbar conjunctiva is fairly loosely adherent to the underlying anterior Tenon's capsule. On histology, the conjunctiva is composed of a surface layer of nonkeratinizing stratified squamous epithelium overlying vascular stroma composed of loose connective tissue. A unique feature of conjunctiva is the presence of goblet cells in stratified squamous epithelium. These goblet cells are specialized apocrine cells which provide the mucin layer to the tear film.[15] The specific mucin type secreted by these cells has been identified as MUC5AC.
Eyelids provide mechanical protection for the ocular surface. Constant blinking of the lids renews the tear film over the surface of the cornea and mechanically removes away superficially adherent foreign bodies and bacteria. Viewed in cross section, it is composed of five layers. The first layer is the cutaneous epithelium consisting of stratified squamous epithelium. The epithelium rests on very thin dermis. The orbicularis oculi muscle is a striated muscle that surrounds the orbital margins. Deep to the orbicularis lies the orbital septum, a membrane sheet which separates the preseptal space from the posterior orbit. Underneath the septum lie the retractors of the eyelids. The tarsus has a cartilaginous consistency. It is composed of dense connective tissue which molds tightly to the curvature of the eye to keep the eyelid firmly on the surface. Within the tarsus are numerous specialized sebaceous glands called the meibomian glands opening at the lid margin. Their secretions are the main source of superficial lipid layer of the tear film. Each eyelash follicle is also associated with sebaceous glands called the glands of Moll. In addition, there are associated apocrine glands called the glands of Zeis.
Financial support and sponsorship
Nil.
Conflicts of interest
There are no conflicts of interest.
References
- 1.DelMonte DW, Kim T. Anatomy and physiology of the cornea. J Cataract Refract Surg. 2011;37:588–98. doi: 10.1016/j.jcrs.2010.12.037. [DOI] [PubMed] [Google Scholar]
- 2.Rüfer F, Schröder A, Erb C. White-to-white corneal diameter: Normal values in healthy humans obtained with the orbscan II topography system. Cornea. 2005;24:259–61. doi: 10.1097/01.ico.0000148312.01805.53. [DOI] [PubMed] [Google Scholar]
- 3.Fares U, Otri AM, Al-Aqaba MA, Dua HS. Correlation of central and peripheral corneal thickness in healthy corneas. Cont Lens Anterior Eye. 2012;35:39–45. doi: 10.1016/j.clae.2011.07.004. [DOI] [PubMed] [Google Scholar]
- 4.Feizi S, Jafarinasab MR, Karimian F, Hasanpour H, Masudi A. Central and peripheral corneal thickness measurement in normal and keratoconic eyes using three corneal pachymeters. J Ophthalmic Vis Res. 2014;9:296–304. doi: 10.4103/2008-322X.143356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Dua HS, Faraj LA, Said DG, Gray T, Lowe J. Human corneal anatomy redefined: A novel pre-descemet's layer (Dua's layer) Ophthalmology. 2013;120:1778–85. doi: 10.1016/j.ophtha.2013.01.018. [DOI] [PubMed] [Google Scholar]
- 6.Thoft RA, Friend J. The X, Y, Z hypothesis of corneal epithelial maintenance. Invest Ophthalmol Vis Sci. 1983;24:1442–3. [PubMed] [Google Scholar]
- 7.Meek KM, Boote C. The organization of collagen in the corneal stroma. Exp Eye Res. 2004;78:503–12. doi: 10.1016/j.exer.2003.07.003. [DOI] [PubMed] [Google Scholar]
- 8.Meek KM, Knupp C. Corneal structure and transparency. Prog Retin Eye Res. 2015;49:1–6. doi: 10.1016/j.preteyeres.2015.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Boote C, Dennis S, Newton RH, Puri H, Meek KM. Collagen fibrils appear more closely packed in the prepupillary cornea: Optical and biomechanical implications. Invest Ophthalmol Vis Sci. 2003;44:2941–8. doi: 10.1167/iovs.03-0131. [DOI] [PubMed] [Google Scholar]
- 10.Müller LJ, Pels E, Vrensen GF. The specific architecture of the anterior stroma accounts for maintenance of corneal curvature. Br J Ophthalmol. 2001;85:437–43. doi: 10.1136/bjo.85.4.437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bourne WM, Nelson LR, Hodge DO. Central corneal endothelial cell changes over a ten-year period. Invest Ophthalmol Vis Sci. 1997;38:779–82. [PubMed] [Google Scholar]
- 12.Rio-Cristobal A, Martin R. Corneal assessment technologies: Current status. Surv Ophthalmol. 2014;59:599–614. doi: 10.1016/j.survophthal.2014.05.001. [DOI] [PubMed] [Google Scholar]
- 13.Oliveira-Soto L, Efron N. Morphology of corneal nerves using confocal microscopy. Cornea. 2001;20:374–84. doi: 10.1097/00003226-200105000-00008. [DOI] [PubMed] [Google Scholar]
- 14.Gipson IK. The ocular surface: The challenge to enable and protect vision: The friedenwald lecture. Invest Ophthalmol Vis Sci. 2007;48:4390. doi: 10.1167/iovs.07-0770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cher I. Ocular surface concepts: Development and citation. Ocul Surf. 2014;12:10–3. doi: 10.1016/j.jtos.2013.10.004. [DOI] [PubMed] [Google Scholar]
- 16.Gipson IK. Goblet cells of the conjunctiva: A review of recent findings. Prog Retin Eye Res. 2016;54:49–63. doi: 10.1016/j.preteyeres.2016.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]