

Abstract
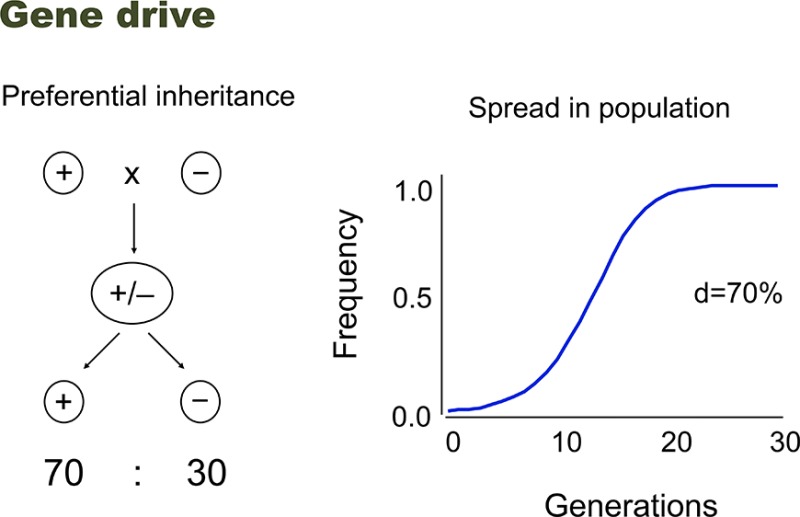
Drive is a process of accelerated inheritance from one generation to the next that allows some genes to spread rapidly through populations even if they do not contribute to—or indeed even if they detract from—organismal survival and reproduction. Genetic elements that can spread by drive include gametic and zygotic killers, meiotic drivers, homing endonuclease genes, B chromosomes, and transposable elements. The fact that gene drive can lead to the spread of fitness-reducing traits (including lethality and sterility) makes it an attractive process to consider exploiting to control disease vectors and other pests. There are a number of efforts to develop synthetic gene drive systems, particularly focused on the mosquito-borne diseases that continue to plague us.
Introduction to Gene Drive
Most genes are thought to spread and persist in populations because they do something useful for the organisms carrying them, increasing survival and/or reproduction, at least on average. That is, most genes spread in populations by positive Darwinian selection and are maintained in the face of recurrent mutation by purifying selection. Some features of a gene may be selectively neutral—such as which nucleotide is found at a particular silent site—but random drift by itself will not lead to long open reading frames and associated control sequences that produce complex proteins performing a particular function.
There are other genes, a minority, which spread and persist not by natural selection, not by increasing organismal survival or reproduction, but instead by distorting their transmission from one generation to the next. For example, some genes manage to be transmitted to more than half of an individual’s gametes, even when that individual only inherited the gene from one of its two parents. In this case, the frequency of the gene increases due to the process of gene transmission from one generation to the next, and it is this unequal genetic transmission that gives the gene its advantage. Genes or genetic elements showing such transmission ratio distortion, or “drive,” include gametic and zygotic killers, meiotic drivers, homing endonuclease genes, B chromosomes, and transposable elements, each of which has evolved several or many times in different taxa.1,2 Moreover, drive has been an important process affecting such genomic features as genome size, base composition, chromosome shape, repeat structure, distribution of recombination hotspots, and centromere structure.1,3,4
Not only can driving genes spread without doing anything useful for the organisms carrying them, they can even spread if they cause some harm, as long as the effect of the transmission distortion is greater than the effect of the reduced survival and reproduction. For this reason, they are often called selfish genes, or selfish genetic elements.1,2 And since they can be harmful to the organism, genes that suppress these genes can themselves spread by natural selection (analogous to the spread of genes suppressing any other parasite), which the selfish gene will then be selected to avoid, potentially leading to an arms race. Occasionally, features of a selfish genetic element may be co-opted to do something useful for the host. Classic examples include mating type switching in yeast, antibody diversification in vertebrates, and telomere maintenance in Drosophila (and in eukaryotes more generally)—the evolution of all of these operations has involved the domestication or co-option of functions of selfish genetic elements.1,3
Synthetic Gene Drive Systems
The fact that gene drive can lead to the spread of fitness-reducing traits makes it potentially useful for controlling disease vectors and other pests. Moreover, the spread can be rapid: in a closed, random mating population, a construct with 100% drive and no fitness effects can increase from 1% to 99% in the population in just nine generations—fast enough to be attractive for public health interventions. Discussions about how to exploit gene drive for pest control date back for decades, long before there was any mechanistic understanding of how they worked, particularly among medical entomologists looking for new ways to control disease vectors.5−10 However, classical genetic approaches were not sufficiently flexible to be able to construct a useful gene drive system. Now, with the recent progress in molecular biology, there is renewed interest in trying to make synthetic gene drive elements,11,12 and preliminary discussions of potential uses have expanded to agriculture and conservation.13
In broad outline, two types of intervention have been considered, either to reduce the size of the target population, or to leave numbers more-or-less intact and genetically modify the population such that it is less harmful (e.g., less able to transmit a pathogen). And three main molecular paradigms are being explored, the use of toxin–antidote systems, chromosomal rearrangements, or sequence-specific nucleases. Toxin-antidote systems and chromosomal rearrangements may be useful for introducing and spreading a new “cargo” gene through a population that makes the population less harmful (e.g., an effector gene that makes mosquitoes unable to transmit a pathogen).14,15 Nuclease-based drive systems may also be used to introduce novel genes, or they may be used for population suppression.16,17
Toxin-Antidote Systems and Chromosomal Rearrangements
Many naturally occurring gene drive systems act as if they produce a toxin and antidote, though often the molecular details are not known. For example, in mice heterozygous for the t-haplotype, and Drosophila heterozygous for Segregation Distorter, these elements somehow act during spermatogenesis to sabotage spermatids or sperm carrying the wild-type allele, with the result that each is transmitted to over 90% of the progeny (compared to the Mendelian 50%).18,19 In Tribolium flour beetles, the medea gene acts in heterozygous females to somehow cause progeny that do not inherit the medea gene to die.20,21 Though the underlying molecular mechanisms are not known, this example stimulated the development of a synthetic gene drive construct in Drosophila with the same logic.22 The construct combined a microRNA-based repressor of myd88 (an important protein normally supplied by the mother into the embryo) with a zygotically expressed myd88 gene that was not affected by the microRNA and supplied the missing protein. As intended, this construct was able to increase in frequency over successive generations in experimental cage populations. Two other medea systems, using different components, have also been developed in Drosophila,23 as have toxin–antidote systems that display maternal-effect lethal underdominance and threshold-dependent invasion into population cages.15,24 Recent descriptions of natural toxin–antidote systems in plants, fungi, and nematodes25−31 may provide further insights into how these sorts of systems can be engineered.
One way for toxin–antidote systems to work is by generating underdominant fitness interactions, in which the heterozygote is less fit than either of the two homozygotes. Underdominant interactions can also be generated with chromosomal rearrangements such as reciprocal translocations, and if these can be introduced at a sufficient frequency into a population (>50% in the simplest scenario), then they can spread to fixation.6 Strains of Drosophila with reciprocal translocations have been engineered, and these showed the expected frequency-dependent spread in lab populations.32
Nuclease-Based Systems: Chromosome Shredding
In Aedes and Culex mosquitoes, there is a naturally occurring driving Y chromosome that, in some crosses, is transmitted to more than 90% of a male’s progeny. First described in the 1960s,5,33 there is still no good understanding of how it works at the molecular level, but cytologically it is associated with breaks of the X chromosome at male meiosis, perhaps having something to do with interrupted crossovers.34,35 This observation led to the idea that cleavage of the X chromosome during male meiosis might lead to drive of the Y.16 In Anopheles gambiae, the most important vector of malaria in Africa, the rRNA genes are found in a single cluster of hundreds of copies on the X chromosome, making it an ideal target,36 and sure enough, production of a nuclease targeting this sequence during spermatogenesis can produce biased sex ratios, up to 95% males, using both an engineered meganuclease and a CRISPR-based nuclease.37,38 A male-biased population sex ratio would be useful because males do not bite people and transmit disease, nor do they contribute as much materially to population productivity, and so total population size is also likely to decline.39 The constructs reported to date do not yet constitute a fully functional gene drive system, because the nuclease genes have been inserted on an autosome, and so are themselves still transmitted in a Mendelian manner; the next step is to put them on the Y chromosome, which is challenging because it is highly repetitive and largely suppressed at meiosis, though some progress has been made.40,41
Nuclease-Based Systems: Homing
Homing endonuclease genes (HEGs) are a class of natural occurring driving elements for which there is a good understanding about the molecular mechanisms, and these are both simple and general enough to potentially be worth exploiting. HEGs encode a nuclease that recognizes and cuts a sequence that typically occurs just once in the genome. The gene is in the middle of its own recognition sequence, disrupting it and protecting the chromosome it is on from being cut. Therefore, in heterozygotes, only the chromosome not containing the gene is cut; it is then repaired using the HEG-containing homologue as a template, with the result that the HEG is copied across to the chromosome where previously it was absent, converting a heterozygote into a homozygote.42,43 This “homing” reaction simply requires a gene encoding a sequence-specific nuclease, with the gene inserted in the middle of its own recognition sequence, and the cell’s DNA repair system takes care of the rest.
HEGs occur naturally in many microbes but have not yet been identified in any insects or vertebrates. Important proof-of-principle experiments demonstrated that the homing reaction can occur in both Drosophila and Anopheles.44−47 These first experiments used meganucleases, and later experiments showed the reaction could also be catalyzed by zinc finger and TALE nucleases,48 and, more recently, CRISPR-based nucleases.49−51 In principle, the homing reaction can be used for both population-wide knockout of target genes, such as those involved in survival, fertility, or pathogen transmission, or for population-wide knock-in of novel effector genes.17,52,53 As with other forms of pest or disease control, due attention must be given to the possibility of resistance evolving.54−57
Prospects
There are a relatively small number of species for which genetic control methods, including gene drive, may be appropriate. Most prominent are those causing or transmitting diseases. Even now, more than 700 000 people die every year from vector-borne diseases, and there is an additional heavy burden of nonlethal morbidity.58 Much of disease control is, ultimately, chemical, with efficacy largely determined by the degree to which production and delivery can be targeted and affordable. Vaccination can be among the most cost-effective of all health interventions because it uses the adaptive immune system to generate and deliver the active agents. The promise of genetic approaches—and gene drive in particular—is again to use biological processes–mating, meiosis, transcription, translation, etc.—for targeted, cost-effective delivery of appropriate chemicals (e.g., nucleases or antimicrobial peptides) that will substantially reduce disease transmission. Indeed, gene drive may take efficiency a step further, with a single release (perhaps with periodic “booster” releases) giving area-wide, population-level control. Important steps have been made toward realizing this potential, though there remains much more to do.
Acknowledgments
We thank the reviewers for useful comments. Funded by grants from the Bill & Melinda Gates Foundation and the Open Philanthropy Project.
The authors declare no competing financial interest.
References
- Burt A., and Trivers R. (2006) Genes in Conflict: the Biology of Selfish Genetic Elements, Belknap Press of Harvard University Press, Cambridge. [Google Scholar]
- Werren J. H.; Nur U.; Wu C. I. (1988) Selfish genetic elements. Trends Ecol. Evol. 3, 297–302. 10.1016/0169-5347(88)90105-X. [DOI] [PubMed] [Google Scholar]
- McLaughlin R. N.; Malik H. S. (2017) Genetic conflicts: the usual suspects and beyond. J. Exp. Biol. 220, 6–17. 10.1242/jeb.148148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Werren J. H. (2011) Selfish genetic elements, genetic conflict, and evolutionary innovation. Proc. Natl. Acad. Sci. U. S. A. 108, 10863–10870. 10.1073/pnas.1102343108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Craig G. B. Jr.; Hickey W. A.; VandeHey R. C. (1960) An inherited male-producing factor in. Science 132, 1887–1889. 10.1126/science.132.3443.1887. [DOI] [PubMed] [Google Scholar]
- Curtis C. F. (1968) Possible uses of translocations to fix desirable genes in insect pest populations. Nature 218, 368–369. 10.1038/218368a0. [DOI] [PubMed] [Google Scholar]
- Curtis C. F.; Grover K. K.; Suguna S. G.; Uppal D. K.; Dietz K.; Agarwal H. V.; Kazmi S. J. (1976) Comparative field cage tests of population suppressing efficiency of 3 genetic-control systems for. Heredity 36, 11–29. 10.1038/hdy.1976.2. [DOI] [PubMed] [Google Scholar]
- Hickey W. A.; Craig G. B. Jr. (1966) Distortion of sex ratio in populations of Aedes aegypti. Can. J. Genet. Cytol. 8, 260–278. 10.1139/g66-033. [DOI] [PubMed] [Google Scholar]
- Hickey W. A.; Craig G. B. Jr. (1966) Genetic distortion of sex ratio in a mosquito Aedes aegypti. Genetics 53, 1177–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinkins S. P.; Gould F. (2006) Gene drive systems for insect disease vectors. Nat. Rev. Genet. 7, 427–435. 10.1038/nrg1870. [DOI] [PubMed] [Google Scholar]
- Adelman Z. N. (2016) Genetic Control of Malaria and Dengue, Academic Press, London. [Google Scholar]
- Champer J.; Buchman A.; Akbari O. S. (2016) Cheating evolution: engineering gene drives to manipulate the fate of wild populations. Nat. Rev. Genet. 17, 146–159. 10.1038/nrg.2015.34. [DOI] [PubMed] [Google Scholar]
- NASEM . (2016) Gene Drives on the Horizon: Advancing Science, Navigating Uncertainty, and Aligning Research with Public Values, The National Academies Press, Washington, DC. [PubMed] [Google Scholar]
- Adelman Z. N., Basu S., and Myles K. M. (2016)Engineering pathogen resistance in mosquitoes, in Genetic Control of Malaria and Dengue (Adelman Z. N., Ed.), pp 277–304, Academic Press, London. [Google Scholar]
- Marshall J. M., and Akbari O. S. (2016) Gene drive strategies for population replacement, in Genetic Control of Malaria and Dengue (Adelman Z. N., Ed.), pp 169–200, Academic Press, London. [Google Scholar]
- Burt A. (2003) Site-specific selfish genes as tools for the control and genetic engineering of natural populations. Proc. R. Soc. London, Ser. B 270, 921–928. 10.1098/rspb.2002.2319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burt A. (2014) Heritable strategies for controlling insect vectors of disease. Philos. Trans. R. Soc., B 369, 20130432. 10.1098/rstb.2013.0432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larracuente A. M.; Presgraves D. C. (2012) The selfish Segregation Distorter gene complex of Drosophila melanogaster. Genetics 192, 33–53. 10.1534/genetics.112.141390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyon M. F. (2003) Transmission ratio distortion in mice. Annu. Rev. Genet. 37, 393–408. 10.1146/annurev.genet.37.110801.143030. [DOI] [PubMed] [Google Scholar]
- Beeman R. W.; Friesen K. S.; Denell R. E. (1992) Maternal-effect selfish genes in flour beetles. Science 256, 89–92. 10.1126/science.1566060. [DOI] [PubMed] [Google Scholar]
- Lorenzen M. D.; Gnirke A.; Margolis J.; Garnes J.; Campbell M.; Stuart J. J.; Aggarwal R.; Richards S.; Park Y.; Beeman R. W. (2008) The maternal-effect, selfish genetic element Medea is associated with a composite Tc1 transposon. Proc. Natl. Acad. Sci. U. S. A. 105, 10085–10089. 10.1073/pnas.0800444105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen C. H.; Huang H. X.; Ward C. M.; Su J. T.; Schaeffer L. V.; Guo M.; Hay B. A. (2007) A synthetic maternal-effect selfish genetic element drives population replacement in Drosophila. Science 316, 597–600. 10.1126/science.1138595. [DOI] [PubMed] [Google Scholar]
- Akbari O. S.; Chen C.-H.; Marshall J. M.; Huang H.; Antoshechkin I.; Hay B. A. (2014) Novel synthetic medea selfish genetic elements drive population replacement in Drosophila; a theoretical exploration of medea-dependent population suppression. ACS Synth. Biol. 3, 915–928. 10.1021/sb300079h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akbari O. S.; Matzen K. D.; Marshall J. M.; Huang H.; Ward C. M.; Hay B. A. (2013) A synthetic gene drive system for local, reversible modification and suppression of insect populations. Curr. Biol. 23, 671–677. 10.1016/j.cub.2013.02.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ben-David E.; Burga A.; Kruglyak L. (2017) A maternal-effect selfish genetic element in Caenorhabditis elegans. Science 356, 1051–1055. 10.1126/science.aan0621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grognet P.; Lalucque H.; Malagnac F.; Silar P. (2014) Genes that bias Mendelian segregation. PLoS Genet. 10, e1004387. 10.1371/journal.pgen.1004387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammond T. M.; Rehard D. G.; Xiao H.; Shiu P. K. T. (2012) Molecular dissection of Neurospora Spore killer meiotic drive elements. Proc. Natl. Acad. Sci. U. S. A. 109, 12093–12098. 10.1073/pnas.1203267109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu W.; Jiang Z. D.; Suo F.; Zheng J. X.; He W. Z.; Du L. L. (2017) A large gene family in fission yeast encodes spore killers that subvert Mendel’s law. eLife 6, e26057. 10.7554/eLife.26057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nuckolls N. L.; Bravo Nunez M. A.; Eickbush M. T.; Young J. M.; Lange J. J.; Yu J. S.; Smith G. R.; Jaspersen S. L.; Malik H. S.; Zanders S. E. (2017) wtf genes are prolific dual poison-antidote meiotic drivers. eLife 6, e26033. 10.7554/eLife.26033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seidel H. S.; Ailion M.; Li J. L.; van Oudenaarden A.; Rockman M. V.; Kruglyak L. (2011) A novel sperm-delivered toxin causes late-stage embryo lethality and transmission ratio distortion in C. elegans. PLoS Biol. 9, e1001115. 10.1371/journal.pbio.1001115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang J. Y.; Zhao X. B.; Cheng K.; Du H. Y.; Ouyang Y. D.; Chen J. J.; Qiu S. Q.; Huang J. Y.; Jiang Y. H.; Jiang L. W.; Ding J. H.; Wang J.; Xu C. G.; Li X. H.; Zhang Q. F. (2012) A killer-protector system regulates both hybrid sterility and segregation distortion in rice. Science 337, 1336–1340. 10.1126/science.1223702. [DOI] [PubMed] [Google Scholar]
- Buchman A. B., Ivy T., Marshall J. M., Akbari O., and Hay B. A. (2016) Engineered reciprocal chromosome translocations drive high threshold, reversible population replacement in Drosophila, bioRxiv, DOI: 10.1101/088393. [DOI] [PubMed] [Google Scholar]
- Wood R. J.; Newton M. E. (1991) Sex-ratio distortion caused by meiotic drive in mosquitos. Am. Nat. 137, 379–391. 10.1086/285171. [DOI] [Google Scholar]
- Newton M. E.; Wood R. J.; Southern D. I. (1976) Cytogenetic analysis of meiotic drive in mosquito, Aedes aegypti (L). Genetica 46, 297–318. 10.1007/BF00055473. [DOI] [Google Scholar]
- Sweeny T. L.; Barr A. R. (1978) Sex-ratio distortion caused by meiotic drive in a mosquito, Culex pipiens L. Genetics 88, 427–446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Windbichler N.; Papathanos P. A.; Catteruccia F.; Ranson H.; Burt A.; Crisanti A. (2007) Homing endonuclease mediated gene targeting in Anopheles gambiae cells and embryos. Nucleic Acids Res. 35, 5922–5933. 10.1093/nar/gkm632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galizi R.; Doyle L. A.; Menichelli M.; Bernardini F.; Deredec A.; Burt A.; Stoddard B. L.; Windbichler N.; Crisanti A. (2014) A synthetic sex ratio distortion system for the control of the human malaria mosquito. Nat. Commun. 5, 3977. 10.1038/ncomms4977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galizi R.; Hammond A.; Kyrou K.; Taxiarchi C.; Bernardini F.; O’Loughlin S. M.; Papathanos P. A.; Nolan T.; Windbichler N.; Crisanti A. (2016) A CRISPR-Cas9 sex-ratio distortion system for genetic control. Sci. Rep. 6, 31139. 10.1038/srep31139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deredec A.; Godfray H. C. J.; Burt A. (2011) Requirements for effective malaria control with homing endonuclease genes. Proc. Natl. Acad. Sci. U. S. A. 108, E874–E880. 10.1073/pnas.1110717108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernardini F.; Galizi R.; Menichelli M.; Papathanos P. A.; Dritsou V.; Marois E.; Crisanti A.; Windbichler N. (2014) Site-specific genetic engineering of the Anopheles gambiae Y chromosome. Proc. Natl. Acad. Sci. U. S. A. 111, 7600–7605. 10.1073/pnas.1404996111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall A. B.; Papathanos P. A.; Sharma A.; Cheng C. D.; Akbari O. S.; Assour L.; Bergman N. H.; Cagnetti A.; Crisanti A.; Dottorini T.; Fiorentini E.; Galizi R.; Hnath J.; Jiang X. F.; Koren S.; Nolan T.; Radune D.; Sharakhova M. V.; Steele A.; Timoshevskiy V. A.; Windbichler N.; Zhang S. M.; Hahn M. W.; Phillippy A. M.; Emrich S. J.; Sharakhov I. V.; Tu Z. J.; Besansky N. J. (2016) Radical remodeling of the Y chromosome in a recent radiation of malaria mosquitoes. Proc. Natl. Acad. Sci. U. S. A. 113, E2114–E2123. 10.1073/pnas.1525164113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colleaux L.; D'Auriol L.; Betermier M.; Cottarel G.; Jacquier A.; Galibert F.; Dujon B. (1986) Universal code equivalent of a yeast mitochondrial intron reading frame is expressed into Escherichia coli as a specific double strand endonuclease. Cell 44, 521–533. 10.1016/0092-8674(86)90262-X. [DOI] [PubMed] [Google Scholar]
- Dujon B. (1989) Group I introns as mobile genetic elements – facts and mechanistic speculations – a review. Gene 82, 91–114. 10.1016/0378-1119(89)90034-6. [DOI] [PubMed] [Google Scholar]
- Chan Y.-S.; Huen D. S.; Glauert R.; Whiteway E.; Russell S. (2013) Optimising homing endonuclease gene drive performance in a semi-refractory species: the Drosophila melanogaster experience. PLoS One 8, e54130. 10.1371/journal.pone.0054130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan Y.-S.; Naujoks D. A.; Huen D. S.; Russell S. (2011) Insect population control by homing endonuclease-based gene drive: an evaluation in Drosophila melanogaster. Genetics 188, 33–44. 10.1534/genetics.111.127506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan Y.-S.; Takeuchi R.; Jarjour J.; Huen D. S.; Stoddard B. L.; Russell S. (2013) The design and in vivo evaluation of engineered I-OnuI-based enzymes for HEG gene drive. PLoS One 8, e74254. 10.1371/journal.pone.0074254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Windbichler N.; Menichelli M.; Papathanos P. A.; Thyme S. B.; Li H.; Ulge U. Y.; Hovde B. T.; Baker D.; Monnat R. J. Jr.; Burt A.; Crisanti A. (2011) A synthetic homing endonuclease-based gene drive system in the human malaria mosquito. Nature 473, 212–215. 10.1038/nature09937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simoni A.; Siniscalchi C.; Chan Y.-S.; Huen D. S.; Russell S.; Windbichler N.; Crisanti A. (2014) Development of synthetic selfish elements based on modular nucleases in Drosophila melanogaster. Nucleic Acids Res. 42, 7461–7472. 10.1093/nar/gku387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gantz V. M.; Bier E. (2015) The mutagenic chain reaction: a method for converting heterozygous to homozygous mutations. Science 348, 442–444. 10.1126/science.aaa5945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gantz V. M.; Jasinskiene N.; Tatarenkova O.; Fazekas A.; Macias V. M.; Bier E.; James A. A. (2015) Highly efficient Cas9-mediated gene drive for population modification of the malaria vector mosquito Anopheles stephensi. Proc. Natl. Acad. Sci. U. S. A. 112, E6736–E6743. 10.1073/pnas.1521077112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammond A.; Galizi R.; Kyrou K.; Simoni A.; Siniscalchi C.; Katsanos D.; Gribble M.; Baker D.; Marois E.; Russell S.; Burt A.; Windbichler N.; Crisanti A.; Nolan T. (2016) A CRISPR-Cas9 gene drive system-targeting female reproduction in the malaria mosquito vector Anopheles gambiae. Nat. Biotechnol. 34, 78–83. 10.1038/nbt.3439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eckhoff P. A.; Wenger E. A.; Godfray H. C. J.; Burt A. (2017) Impact of mosquito gene drive on malaria elimination in a computational model with explicit spatial and temporal dynamics. Proc. Natl. Acad. Sci. U. S. A. 114, E255–E264. 10.1073/pnas.1611064114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Godfray H. C. J.; North A.; Burt A. (2017) How driving endonuclease genes can be used to combat pests and disease vectors. BMC Biol. 15, 81. 10.1186/s12915-017-0420-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beaghton A.; Beaghton P. J.; Burt A. (2017) Vector control with driving Y chromosomes: modelling the evolution of resistance. Malar. J. 16, 286. 10.1186/s12936-017-1932-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beaghton A.; Hammond A.; Nolan T.; Crisanti A.; Godfray H. C. J.; Burt A. (2017) Requirements for driving antipathogen effector genes into populations of disease vectors by homing. Genetics 205, 1587–1596. 10.1534/genetics.116.197632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Champer J.; Reeves R.; Oh S. Y.; Liu C.; Liu J. X.; Clark A. G.; Messer P. W. (2017) Novel CRISPR/Cas9 gene drive constructs reveal insights into mechanisms of resistance allele formation and drive efficiency in genetically diverse populations. PLoS Genet. 13, e1006796. 10.1371/journal.pgen.1006796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammond A. M.; Kyrou K.; Bruttini M.; North A.; Galizi R.; Karlsson X.; Kranjc N.; Carpi F. M.; D’Aurizio R.; Crisanti A.; Nolan T. (2017) The creation and selection of mutations resistant to a gene drive over multiple generations in the malaria mosquito. PLoS Genet. 13, e1007039. 10.1371/journal.pgen.1007039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WHO Vector-borne diseases . http://www.who.int/mediacentre/factsheets/fs387/en/ (accessed Nov 26, 2017).