

Abstract
The aim of the present study is to evaluate plant growth promoting and biocontrol efficacy of a Serratia marcescens strain ETR17 isolated from tea rhizosphere for the effective management of root rot disease in tea. Isolated bacterial culture ETR17 showed significant level of in vitro antagonism against nine different foliar and root pathogens of tea. The phenotypic and molecular characterization of ETR17 revealed the identity of the bacterium as Serratia marcescens. The bacterium was found to produce several hydrolytic enzymes like chitinase, protease, lipase, cellulase and plant growth promoting metabolites like IAA and siderophore. Scanning electron microscopic studies on the interaction zone between pathogen and antagonistic bacterial isolate revealed severe deformities in the fungal mycelia. Spectral analyses (LC-ESI-MS, UV-VIS spectrophotometry and HPLC) and TLC indicated the presence of the antibiotics pyrrolnitrin and prodigiosin in the extracellular bacterial culture extracts. Biofilm formation by ETR17 on polystyrene surface was also observed. In vivo application of talc-based formulations prepared with the isolate ETR17 in tea plantlets under green house conditions revealed effective reduction of root-rot disease as well as plant growth promotion to a considerable extent. Viability studies with the ETR17 talc formulation showed the survivability of the isolate up to six months at room temperature. The sustenance of ETR17 (concentration of 8-9x108 cfu g-1) in the soil after the application of talc formulation was recorded by ELISA. Safety studies revealed that ETR17 did not produce hemolysin as observed in pathogenic Serratia strains. The biocontrol strain reported in this study can be used for field application in order to minimize the use of chemical fungicides for disease control in tea gardens.
Introduction
North-East India especially the sub-Himalayan region of West Bengal and adjoining state of Assam is the largest tea growing region of India. High humidity is required for the growth of tea leaves. High humidity makes the tea plants prone to attack by several fungi as fungi thrive well in humid atmosphere. The varying conditions of climate, soil and several environmental stresses required for tea plantation makes it vulnerable to attack by several fungal pathogens [1–3]. Over the years, plant disease management strategies were mostly based on the use of chemical fungicides. But the uncontrolled and persistent use of chemicals results in harmful residual effect on plants, natural soil microflora and human health. Pathogenic fungus may also develop resistance against fungicides [4, 5, 6]. The rhizosphere inhabiting antagonistic bacteria serve as a promising alternative to synthetic fungicides for controlling several plant diseases without disturbing the natural ecosystem [7, 8, 9].
A comprehensive research in the past few decades has highlighted the role of both gram-positive and gram-negative bacterial strains as promising biological control agents [10–13]. Among the antifungal metabolites produced by biocontrol agents to combat phytopathogens, antibiotics are the most widely studied compounds [14]. In several instances, antibiotics such as pyrrolnitrin, 2, 4- diacetylphloroglucinol (DAPG), prodigiosin, pyoluteorin, phenazine-1-carboxylic acid (PCA), paenimyxin and several others have been shown to be particularly effective in suppressing plant pathogens and reduces the disease incidences they cause [15–20]. The expression and secretion of lytic enzymes like chitinase, cellulase, protease and DNase produced by soil bacteria also result in direct suppression of pathogenic activities by hydrolyzing several polymeric compounds [10, 21, 22]. Furthermore, bacterial siderophores play a major role in plant disease suppression by iron sequestration since fungal siderophores have lower affinity towards iron available in soil [23, 24]. Literature reports indicate that biological control of plant pathogens is a complex process and antagonism against target fungus is often multifaceted [5].
Root associated bacteria can influence plant growth and development and overall plant health [25–27]. Additionally, extensive research has been carried out to provide effective means of in vivo application of biocontrol agents in the form of stable bioformulations and their sustenance in soil environment for enhanced disease control and plant growth promotion [2, 28, 29]. Despite the apparent successes associated with the use of biocontrol agents for combating the plant diseases, their chemical counterparts still succeed over them. The reason lies with the fact that, under field conditions a biocontrol agent largely depends upon its interactions with the natural microbiota, ability to compete for nutrients, effective root colonization and adapting to the changes in environmental conditions [30–32].
The present study aims at the use of a potential antagonistic bacterial strain indigenous to tea rhizosphere for controlling a severe root disease of tea. In an attempt to fulfill the objectives of this study, a potential strain of S. marcescens with multiple bicontrol attributes was isolated from tea rhizosphere and characterized. Biocontrol experiments were conducted against Rhizoctonia root rot in experimental tea seedlings.
Materials and methods
Pathogens
Nine different fungal strains causing several foliar and root diseases in tea (Camellia sinensis) were used in the present study. The pathogens Curvularia eragrostidis (ITCC 4150.2k), Pestalotiopsis theae (PT01), Colletotrichum camelliae (CC01), Lasiodiplodia theobromae (ITCC 5446.02) and Rhizoctonia solani (ITCC 5995.05) were isolated from affected tea plants during earlier studies [1, 33–36] and the identities of the ITCC strains were authenticated by Indian Agricultural Research Institute, New Delhi. Sphaerostilbe repens (SR-01), Fomes lamaoensis (FL-01), Poria hypobrunae (PH-01) and Ustulina zonata (UZ-01) were procured from Tea Research Association, Tocklai Experimental Station, Jorhat, Assam.
Plant material
One month old disease free tea (Camelia sinensis) seedlings of TS-520 variety were obtained from Gayaganga tea estate located in Darjeeling district of West Bengal during April. The soil of the tea estate was sandy loam(pH of 4.8).The organic carbon content was moderate at 20.9 g/kg of soil with electrical conductivity of 0.03 dsm-1. The available N-P-K content of the soil was 122.45 kg/ha-1, 22.12 kg/ha-1and 261.33 kg/ ha-1 respectively [37]. The temperature prevailing was 27 ± 2°C with average rainfall of 94 mm. The procured seedlings were maintained in earthen pots of 16cm diameter and 11cm height in the experimental garden of the Department of Botany, University of North Bengal under normal light and temperature. The tea plantlets were used for experimental purpose after 30 days of acclimatization.
Isolation of bacteria
Rhizosphere soil samples were collected from fifteen different tea gardens located in West Bengal and Assam. The tea plantations of the sampling sites selected for the study remained free from any prior application of biocontrol agents. Roots along with adherent soil were collected from healthy plants aseptically in sterilized bags and transported to the laboratory within an hour. Isolation of bacteria was done following previously described methods [38, 39]. Two hundred bacterial strains were isolated in nutrient agar medium (HiMedia Laboratories Pvt. Ltd., Mumbai) and maintained in nutrient agar (NA) slants. After incubation for 72 h at 30°C in inverted position, isolated single colonies were picked from spread plates and pure cultures were obtained on NA slants. The strains were subsequently maintained in NA slants with periodic transfers to fresh medium.
Test for in vitro antagonism
All isolated strains were initially screened in vitro by dual culture technique for the presence of antagonistic activity against L. theobromae on potato dextrose agar (PDA) medium [39]. A 4 mm diameter mycelial disc of the fungal pathogen was collected from advancing zone of hyphae growing in PDA plates and inoculated at the centre of a 9 cm diameter petriplate containing PDA medium. The bacterial isolate was streaked at a distance of 2–3 cm from the centre in semi-circular/circular pattern. The plates were incubated at 30°C and checked daily for inhibition until the fungal growth on the control plate (inoculated only with the pathogen) reached the edge of the plate. Inhibition of fungal growth along the bacterial line of streaking indicated antagonistic activity of the isolated bacterial strain. Based on the initial screening (S1 Table), the most promising isolate ETR17 was selected for further study. The isolate was further tested for antagonism against eight other fungal pathogens following the same method. Percentage (%) of inhibition of the mycelial growth of test pathogens was calculated as [(Diameter of pathogen growth in control plate (mm)-Diameter of pathogen growth in presence of antagonist(mm)/Diameter of pathogen growth in control plate (mm)] X 100 [38, 39]. The dual culture tests were performed in three replications and the data was averaged. L. theobromae was chosen for initial screening because it is the most common pathogen of tea and can attack all parts of plant [40–42].
Characterization of ETR17
Phenotypic traits of the bacterial isolate ETR17 were examined based on the biochemical, physiological and morphological characteristics [43, 44]. The culture characteristics such as shape, size, colony nature and pigmentation of 3 day old bacterial culture were examined. The isolate was further tested for gram staining, indole, methyl red, Voges Proskauer, citrate utilization, catalase, oxidase, gelatin and starch hydrolysis, oxidation-fermentation, nitrate reduction, decarboxylation of ornithine, arginine and lysine, utilization of different carbon sources such as glucose, lactose, mannitol, sucrose, fructose, arabinose, adonitol, cellobiose, inositol, rhamnose, raffinose, sorbitol, trehalose and xylose and growth at four different temperatures. Results of these tests were scored as either positive or negative.
Further characterization of the antagonistic isolate was performed using 16SrRNA gene sequence. In brief, the total genomic DNA was isolated from ETR17 by CTAB method [45]. Amplification of the 16S rRNA gene was performed in 25μl reaction using Universal primers following specific conditions on a thermal cycler (Applied Biosystems GeneAmp PCR 2400) [39]. The amplified product was purified using PCR purification Kit (Bangalore Genei, India) and sequenced at Bangalore Genei Sequencing Services. The 16S rRNA gene sequence was thereafter deposited in the NCBI GenBank and accession number (JX566992) was assigned. A similarity search of the sequence was carried out and compared with the 16S rRNA sequences of related species available in GenBank databases using the BLAST search program of the National Center for Biotechnology Information (NCBI) [46]. The 16S rRNA gene sequences of the bacterial strains having similarity ranging from 98% to 100% with the target sequence were used for sequence alignment.
Studies on the interaction between biocontrol isolate ETR 17 and pathogen
The fungal mycelia of the pathogen R. solani collected from the interaction zone of the bacterium ETR17 and the pathogen in dual culture PDA plates were used as samples for microscopic studies. The sample was prepared following the method of Saha et al [39] and viewed under a Scanning electron microscope [Model: Hitachi S-530 (Japan) 1986].
Detection of extracellular hydrolytic enzymes produced by ETR17 isolate
Chitinase activity was detected by following the method of Bargabus et al [47]. Positive result was indicated by the presence of non fluorescent lytic zones around the wells under UV light. Production of cellulase and pectinase enzymes was detected following the method of Cattelan et al [48] with some modifications. For assessing cellulase activity, bacterial culture supernatant was added to 4mm agar cups of M9 agar medium [49] supplemented with 10 g L-1 cellulose and 1.2 g L-1 yeast extract. The plate was incubated at 30°C for 5–6 days and thereafter stained with 0.1% Congo red solution overnight and destained thrice with 1M NaCl at 2hrs interval. Formation of clear halo indicated positive result. Pectinase activity was tested in the same medium by amending with 10 g L-1 pectin instead of cellulose. The plate was streak inoculated and incubated for 2 days at 30°C, then it was flooded with 2M HCl. A positive result was indicated by visible clear halos around the colonies. Lipase and protease activities were tested following the method of Smibert and Kreig [50].
Testing for plant growth promoting factors and other biocontrol traits
Production of the phytohormone IAA was investigated spectrophotometrically by using the method of Patten and Glick [51]. The absorbance maxima of IAA in the bacterial culture supernatant were recorded at 535 nm and its concentration was determined by comparison with the standard curve generated from different concentrations of IAA (Merck, India). The ability of S. marcescens strain ETR17 to solubilize phosphate was determined in Pikovskaya’s agar media [52]. Positive result is indicated by the development of a clear zone around the bacterial colonies. Siderophore production was determined using the Universal Chrom azurol S (CAS) assay [53]. Identification of catecholate type of siderophore was carried out following the method of Arnow [54]. Hydroxamate nature was examined by tetrazolium salt test indicated by instant appearance of a deep red colour by addition of siderophore sample to tetrazolium salt under alkaline conditions [55]. Test for HCN production was carried out by the method of Bakkers and Schippers [56] where positive result was indicated by the color shift of a filter paper strip immersed in picric acid from yellow to red. Biofilm formation on microtiter plates by the biocontrol bacterial isolate was assessed in two different medium [Luria Bertani (LB) broth and M9 Yeast extract (M9YE) broth [57].
Extraction, detection and purification of antifungal antibiotic from antagonistic bacterium S. marcescens strain ETR17
Bacterial strain ETR17 was inoculated into semi-solid pigment producing media [58] by pour plate method and incubated at 30°C for 8 d. The total content (180 mL) was crushed in a blender and extracted with 250 mL of 80% aqueous acetone for 24 hours in an orbital shaker [59]. Agar was removed by centrifugation at 15,000 rpm for 20 min at 10°C and the supernatant containing antibiotics was condensed at 40°C in a rotary vacuum evaporator (Eyela CCA-1110, Japan). The aqueous concentrate was filtered through cellulose acetate filter paper (Sartorius, pore size 0.2 μm) and 20 mL portions of the filtrate were extracted twice with 2.5 volumes of diethyl ether. The organic phase containing antibiotics were evaporated to dryness in vacuo at 30°C and the residue was re-extracted with 30 mL of acetone, and finally condensed under vacuum to obtain a red pasty mass [38].
The crude extract (2.5 g) was dissolved in 10 mL methanol, mixed and dried over silica gel (mesh 60–120) and was subjected to silica gel column chromatography. Elution with 100% petroleum ether and petroleum ether -ethyl acetate (95% to 50%) yielded a total of eleven fractions (F1 to F11) of 100 mL each. Each fraction was vacuum concentrated and tested for antifungal activity. The active fractions F3, F4, F5, F10 and F11 were monitored by thin layer chromatography (TLC). Samples were loaded on TLC sheets pre-coated with Silica Gel 60 F254 and co-chromatographed with standard antibiotics pyrrolnitrin, prodigiosin and phenazine (Sigma-Aldrich). The sheets were developed in benzene:acetic acid (9:1) and viewed under UV (254 nm) light. The antibiotics present in the bioactive fractions were further purified by preparative TLC. For this, large volumes (1 mL) of the bioactive fractions were spotted as before on glass-backed preparative TLC plates (prepared manually by coating with silica gel G. The sheets were developed similarly in benzene: acetic acid (9:1) and the zone corresponding to the Rf value obtained on analytical plates were scrapped from the TLC plate, suspended in methanol and centrifuged. The supernatant was dried in vacuo and dissolved in methanol. Partially purified compound in methanol was scanned between 200 to 700 nm on a dual beam Varian Cary 50 Bio UV-Visible spectrophotometer (Varian, Australia) along with standard prodigiosin and pyrrolnitrin. Thus two antibiotics were detected by analytical TLC. High performance liquid chromatographic analysis of the antifungal metabolites purified by TLC was performed in Shimadzu SPD-20A, Japan. Fractions which appeared to contain same antibiotics were combined, dissolved in methanol (50μg per mL) and 20 μl was injected into C18 Reverse Phase column (250 x 4.6mm size and 4μm particle size) (Phenomenex, USA). The pump used was LC-20AD (Shimadzu, Japan). The eluent flow rate was adjusted to 1 mL min-1 and analyzed isocratically in 100% methanol. Standard antibiotics were used at a concentration of 10 μg mL-1. Pyrrolnitrin was detected at 225 nm using a D2 detector (Prominence, Shimadzu, Japan) and prodigiosin was detected at 536 nm using tungsten (W) detector (Prominence, Shimadzu, Japan).
Presence of pyrrolnitrin was further confirmed by LC-ESI-MS analysis using a gradient elution program with solvent A (methanol) and solvent D (ammonium acetate buffer, pH 6.5): 50% solvent A and 50% solvent D from 0 to 10 min; 70% solvent A, 30% solvent D at 10 min and 80% solvent A, 20% solvent D till 30 min at a flow rate of 0.8 mL min-1 and 254 nm. A 20 μl sample was injected into the column without any dilution. The column used was Thermo ODS-2 (250 x 4.6 mm size and 5 μm particle size) (Thermo, India). The electrospray ionization mass spectra were recorded on a Thermo LCQ Advantage Max (Thermo, India) with the following specifications: Source voltage 5.3V, source current 80.0 μA, capillary voltage 3.0 V, tube lens offset 5.0 V and capillary temperature of 300°C.
Test for in vitro antifungal activity
Antifungal activity of the crude extract and column eluted fractions of ETR17 was assessed in vitro against the test pathogens by agar cup bioassay. For this, PDA medium was autoclaved at 121°C for 15 min, cooled to 45°C and 1 mL of pure sporulated mycelial suspensions ofpathogens were mixed with 19 mL of molten medium, and poured into sterile petriplates of 9 cm diameter. Spore suspension of each pathogen except R. solani was prepared in sterile distilled water (1 x 106 conidia mL-1) from 10–12 day- old PDA cultures of each of the pathogens following the method of Saha et al [1]. In case of R. solani, mycelium from a 7 day old culture was scrapped lightly in 2 mL of sterile distilled water with inoculation needle and mixed with 18 mL molten medium. The mixture was then poured into sterile petriplates (9 cm diameter) and allowed to solidify.After solidification of the medium in petriplates, wells (4 mm in diameter) were prepared with sterile cork-borer. Crude metabolite or individual column fractions were diluted separately by dissolving 100 μl of it in 1mL methanol and then loaded into the wells. A well in the same plate was loaded with 100% methanol which served as the control. The plates were incubated at 28°C for 2–4 days and diameter of inhibition zones formed around the wells, if any, was measured.
In vivo biocontrol study for reduction of root rot disease in tea using bacterial formulation
Preparation of talc-based bacterial formulation
Talc-based formulation was prepared with bacterial strain ETR17 following the method of Nandakumar et al [60]. At first, 48 h old nutrient broth (NB) (HiMedia Laboratories Pvt. Ltd. Mumbai, India) culture of the bacterium was used to inoculate 400 mLof sterile NB and grown at 30°C on a rotary shaker operating at 150 rpm. The culture at its stationary phase of growth was centrifuged at 6000 rpm for 10 minutes and bacterial cells were resuspended in 10 mM phosphate buffer (pH 7.0). The concentration was adjusted to 9x108 colony forming unit (cfu) mL-1. Talc powder (used as carrier) was mixed thoroughly with calcium carbonate at the rate of 15 g kg-1 for adjusting its pH to 7.0. Next, carboxymethyl cellulose was added to the talc (at 10 g kg-1), mixed well and the resultant mixture was autoclaved (for 30 min at 15 psi pressure) twice in two consecutive days. Bacterial culture (400 mL) was added to 1 kg of the carrier-cellulose mixture and mixed well under sterile conditions. The mixture was dried aseptically and packed in sterile polypropylene bags, sealed and stored at room temperature (25± 2°C) for future use.
Preparation of fungal inoculum
Pathogen inoculum was prepared following the method of Soares et al [61]. For this, 300 g of rice grains were soaked in 500 mLof distilled water at room temperature for one hour for proper hydration. The excess water was drained off and the grains were distributed equally in three 250 mLconical flasks and autoclaved (for 30 minutes at 15 psi pressure) twice on two consecutive days. Mycelial disc (6 mm) of the pathogen R. solani excised from actively growing regions of a 7 day old fungal culture in PDA plate was used to inoculate the sterilized rice and incubated for 6–9 days at 28°C until the pathogen mycelia covered all the rice grains. The resulting inoculum of mycelium infested rice grains were added at the rate of 10 g per kg soil in the experimental pots.
Biocontrol using talc-based formulation
For in vivo biocontrol study, two month old tea seedlings were transferred to pots (10 cm x 15 cm) each containing 2 kg of either sterile or non-sterile garden soil amended with 25% super phosphate: rock phosphate (1:1) fertilizer. Ten grams of talc based formulation was mixed with 100 mL of sterile distilled water and poured into the experimental pots. Altogether 25 plants per treatment were used and 5 plants were uprooted each day for assessment of disease. A set of control pots received no treatment. A fungicide control was included where the pots were treated with 0.1% thiophanate methyl (Biostadt India Ltd., Mumbai). After three days of treatment, inoculation was done with the root rot pathogen R. solani. For this, the soil was removed carefully, mixed with the rice inoculum and replaced back to the original pots. The whole experiment was done under both sterile and non-sterile soil conditions in order to confirm the biocontrol efficacy of ETR17 and to check the effect of any indigenous microorganism present in the soil in disease control. The soil used had sandy-loam texture and acidic pH of 4.5–5.0. The plants were covered with plastic bags to maintain humid condition and kept in experimental net house under normal light and temperature conditions (light: approximately 13 hours each day; temperature: 22.8 to 32.1°C). The plants were arranged in a randomized block design with five replicates for each assessment and each treatment. The entire experiment was conducted thrice and the results of individual experiments were taken together to determine the mean and expressed as mean disease index (MDI). Assessment of root disease was done at 15, 20, 25, 30 and finally 45 days after pathogen inoculation (dapi). Severity of the symptoms was graded into five disease classes (0–4) [62]. The total of 5 seedlings for each treatment were uprooted carefully and graded as follows: 0 = no disease; 1 = superficial rot affected roots; 2 = moderate rot on roots; 3 = severe rot and 4 = completely damaged roots. Based on the classes, disease index was calculated using the following formula: Disease index = [Ʃ (P x DC) x 100]/ (T x4) where P = plants per class, DC = disease class and T = total number of plants. Percent efficacy of disease control (PEDC) was calculated using the formula: PEDC = [(MDI in untreated control–MDI in treated plants)/MDI in untreated control] x100 [38].
Determination of the survivability of ETR17 in talc formulation
The population of S. marcescens strain ETR17 was assessed in the talc formulated product at an interval of one month for one year. One gram samples of talc formulation were collected aseptically and suitable dilutions (10−1 to 10−5) were prepared in sterile distilled water. An aliquot of 100 μl of the individual dilutions were inoculated in NA medium by spread plate technique. The visible number of colonies formed after incubation at 30°C for 24 h were recorded and the bacterial population was expressed as cfu g-1. The plate count was recorded as mean of three replicates and plotted against time as log cfu g-1.
Sustainability assay of ETR17 in the rhizosphere by ELISA
The sustainability of ETR17 in the rhizosphere when applied as talc based formulation was determined by indirect ELISA [63, 34]. For preparation of bacterial antigen, ETR17 isolate was grown in NB for 48 hours at 30°C on a rotary shaker. The resulting cell culture was centrifuged at 10,000 rpm for 10 min; the pellet was suspended in sterilized 0.15M phosphate buffer saline (PBS) (pH 7.2) and washed thrice by centrifugation using previous conditions. The pellet was resuspended in PBS and bacterial population was inactivated by addition of 1% formaldehyde. The concentration of bacterial suspension was adjusted to an OD545 nm of approximately 1.0 [64]. For raising antiserum against bacterial antigen, equal volume of the cell suspension and Freund’s incomplete adjuvant (Bangalore Genei, India) were mixed and used as antigens for immunizing New Zealand white male rabbit. Antisera were collected using standard methods at regular intervals [34] and stored in sterile cryo-vials at -20°C until required.
For soil antigen preparation, rhizosphere soil samples were collected from the experimental sets containing the bacterial formulation and control sets not treated with bacterial formulation after 45 days of pathogen inoculation. One gram of each soil sample was added in 1 mL of PBS-Tween (phosphate buffered saline, pH 7.0 containing 0.02M phosphate buffer, 0.8% NaCl, 0.02% KCl and 0.05% Tween-20) and mixed by vortexing for 30 seconds. The mixture was allowed to stand at room temperature until the soil particles settled down; the supernatant was filtered using sterile Whatman Grade-I filter paper and the filtrate was used as antigen for indirect ELISA [65].
Indirect ELISA was done as described earlier [34] with necessary modifications. Soil antigen (100 μl) was mixed with an equal volume of 0.2M carbonate buffer (pH 9.6) and coated in the wells of a microtiter plate except the air blank and antigen blank. A homologous antigen-antibody reaction with the whole cell antigen was set as positive control. For this, the bacterial cell suspension in PBS was diluted serially and each dilution was used as antigen for ELISA. For negative control, soil antigen prepared from the control pots containing untreated sterilized and unsterilized soil was allowed to react with ETR17 antiserum. The plate was incubated overnight at 4°C for adsorption. After 24 hour the antigen was poured off, the wells were dried and subsequently washed 4 times with 0.15M PBS (pH 7.2) containing 0.02% sodium azide and 0.05% Tween-20 (v/v). The plate was again air dried and 100 μl of PBS-BSA (0.15M phosphate buffered saline and 1% bovine serum albumin) solution was added to block the unbound sites and incubated for 2 hours at room temperature. The plate was rewashed thoroughly with PBS-Tween and air dried. Two hundred microlitre of antiserum (1:100 dilution; diluted with PBS-Tween) was added to the wells except air blank, antiserum blank and normal serum control where same volume of normal serum was added (1:100 dilution; diluted with PBS-Tween containing 0.5% BSA). Following overnight incubation at 4°C, the sera were poured off and the plate was again thoroughly washed with PBS-Tween and dried. Then, 100 μl of goat anti-rabbit IgG HRP (Horse Raddish Peroxidase) conjugate (1:10,000 dilution) was added to each well except the air blank and antiserum blank and incubated for 2 hours at room temperature. The plate was washed thoroughly with PBS-Tween, air dried and 100 μl of TMB/H2O2 (1:20), a chromogenic substrate, was added to each well except the air blank. It was incubated for 30 minutes in dark at room temperature for development of blue color due to enzyme-substrate reaction. The reaction was stopped by adding 100 μl of 1N H2SO4 and absorbance was recorded at 492 nm in an ELISA reader (Mios Junior; Merck, Darmstadt, Germany). A standard curve was plotted using varying bacterial concentrations of the positive control against the corresponding ELISA values and this curve was used to determine the concentration of bacteria in soil. Concentration of bacteria in the serially diluted antigen preparations was determined by NA plate count method.
Plant growth promotion by bacterial formulation
To determine the plant growth promoting ability of ETR17, two month old tea seedlings were treated twice at an interval of 15 days by talc formulation of the strain by soil drenching method in sterilized and unsterilized soil sets as done during in vivo biocontrol assay. The shoot and root length of the seedlings were measured with a centimeter scale prior to treatment with bacterial formulation and again after 45 days of treatment, by carefully uprooting the seedlings. Plants treated with sterile distilled water grown in both sterilized and unsterilized soil served as control sets. All the treatments were replicated thrice and for each treatment five healthy plants were selected.
Test for hemolytic activity of ETR17
Hemolytic activity of the bacterial isolate ETR17 was assessed on tryptone soya agar (TSA) medium (Himedia Laboratories, India) supplemented with 5% goat blood. For this, the bacterial isolate was streaked on blood supplemented TSA plates and incubated at 30°C for 48 hours. After incubation, the plate was observed against light for halo formation around the bacterial growth. A clear halo indicates β-hemolysin production, dark colored halo indicate α, α′ hemolysin production and green colored halo indicate γ hemolysin production [66].
Statistical analysis
Data were analysed by ANOVA using the statistical software SPSS, version 11.0, SPSS Inc., Chicago, Illinois. Specific differences in means were compared by analysing Duncan’s multiple range test (DMRT).
Results
In vitro screening for antagonistic activity of bacterial isolates
Out of the tested 200 bacterial rhizosphere isolates, 35 strains exhibited antagonism against L. theobromae in PDA plate dual culture assay. The antagonistic strains were further tested against eight other fungal pathogens of tea. Results revealed that all strains were antagonistic to many of the tested fungi (Table 1). The isolate ETR17 was found to exhibit highest antifungal activity against several of the tested pathogens and was therefore selected for further studies. It was found to limit the growth of fungal mycelia to a considerable extent in comparison to the control. The level of antagonism exhibited by ETR17 in PDA plate assay against individual pathogens is summarized in Table 2.
Table 1. List of bacterial isolates from different rhizosphere soil.
| Source of isolation | Code assigned | Total No. of isolates | Antagonistic isolates |
|---|---|---|---|
| Bagdogra T.E. | TBD1–10 | 10 | TBD1-7 |
| Bagrakote T.E. | BTRL1-11 | 11 | BTRL6, BTRL8, |
| Baradighi T.E. | BTR1-24 | 24 | BTR4, BTR8, BTR18, |
| Diana T.E. | D1-10 | 10 | D6, D7 |
| Ellenbarie T.E. | ETR1-24 | 24 | ETR1, ETR17, ETR20, |
| Gayaganga T.E. | TGY1–17 | 17 | TGY1, TGY2, TGY4 |
| Good Hope T.E. | GH1-33 | 30 | GH4, GH6, GH12, GH13, GH21. |
| Kharibarie T.E. | KTR1-18 | 15 | KTR6, KTR18 |
| Labak T.E. | TLB1-11 | 11 | TLB3 |
| Matigara T.E. | TMG1–15 | 15 | TMG1, TMG2, TMG3 |
| Raya T.E. | TR1–20 | 15 | TR1, TR5, TR11, TR19, TR20 |
| Red bank T.E. | TRB1–19 | 18 | TRB1, TRB2, TRB4, TRB7, |
| Total number of isolates | 200 | 35 | |
T.E.: Tea Estate
Table 2. In vitro study of antagonistic activity of bacterial isolate ETR17 against fungal pathogens of tea by dual culture test.
| Fungal Pathogens | Disease produced | Strain identity | Percent inhibition of pathogen by ETR17 over control* |
|---|---|---|---|
| Lasiodiplodia theobromae | Root rot or diplodia | 5446.02 | 51.5 ± 0.8a |
| Rhizoctonia solani | Root rot | 5995.05 | 66.7 ± 0.7b |
| Sphaerostilbe repens | Violet root rot | SR-01 | 71.1± 0.7c |
| Fomes lamaoensis | Soft rot | FL-01 | 76.7 ± 0.4d |
| Ustulina zonata | Stump rot | UZ-01 | 61.1 ± 0.5e |
| Poria hypobrunnae | Poria root rot and stem canker | PH-01 | 71.1 ± 0.7c |
| Pestalotiopsis theae | Grey blight | PT01 | 76.7 ± 0.6d |
| Colletotrichum camelliae | Brown blight | CC01 | 65.5 ± 0.5eb |
| Curvularia eragrostidis | Leaf spot | 4150.2k | 81.5 ± 0.5f |
| LSD (at 5%) | 2.3 | ||
* Inhibition zone was measured when fungal mycelia in the control plate reached the edge of 9 cm petriplates.
Means ± standard error followed by same superscript letters do not differ significantly at P<0.05
Identification and characterization of antagonistic isolate ETR17
Characterization tests revealed that ETR17 was gram-negative, rod-shaped, facultatively anaerobic bacteria which produced smooth, regular edged, round shaped colonies and excessive red pigment in nutrient agar. It showed catalase positive, oxidase negative and VP positive traits, was fermentative in nature and capable of reducing nitrate, hydrolysing gelatin, was positive for lysine and ornithine decarboxylation and utilizing citrate and different carbohydrates such as glucose, mannitol, sucrose, fructose, inositol, sorbitol, trehalose and xylose. The bacterium was capable of growing over a wide temperature range (4°C to 40°C) with optimum growth temperature of 28–30°C. Comparing the results with the keys published in Bergey’s Manual of Systematic Bacteriology (Palleroni, 1984), it was concluded that the bacteria belonged to the genus Serratia. Partial sequence of the 16S rRNA gene (GenBank accession no. JX566992) was 99% similar to such gene sequences of other S. marcescens strains obtained from GenBank. Results of phylogenetic analyses were consistent with the biochemical and physiological traits and the isolate was therefore designated as S. marcescens.
Scanning electron microscopy
For understanding the antagonism involved in the interaction between the fungal pathogen Rhizoctonia solani and bacterial strain ETR17, scanning electron microscopy study was conducted. Microscopic observations revealed severe deformations of the fungal mycelia like surface irregularities, perforations, and bulging and bursting in the hyphae at specific sites (Fig 1). Bursting of the fungal mycelia at some places resulted in release of the protoplasm from fungal cell.
Fig 1. Scanning electron microphotographs showing the antagonistic effect of ETR17 on Rhizoctonia solani; lysis of fungal hyphae is indicated by arrows.

Production of antifungal and plant growth promoting metabolites, hydrolytic enzymes and biofilm
The isolated S. marcescens strain ETR17 was positive for chitinase, lipase and protease activity while it scored negative for the production of pectinase enzyme and HCN. Additionally, the isolate was capable of producing IAA (40 μg mL-1) and siderophore on CAS agar plates but lacked phosphatase activity. Arnow’s test and tetrazolium salt test confirmed the production of both catecholate and hydroxamate type of siderophore. Biofilm formation, which was tested by micro-titre plate assay, depended on the growth medium. It was more pronounced in LB medium (O.D. = 0.254 and 0.448 after 24 and 48 h respectively) than in M9YE medium (O.D. = 0.118 and 0.193 after 24 and 48 h respectively).
Characterization of antifungal metabolite
The crude extract obtained from the culture media produced clear zones of inhibition around the wells against the test pathogens indicating antibiotic production by ETR17. Column chromatography of the crude extract and subsequent bioassay of the fractions revealed that the fractions F3, F4, F5, F10 and F11 possessed antifungal activity. Comparative analysis of each bioactive fraction with standard antibiotics in TLC plates under UV light (254 nm) revealed that the major compound in fractions 10 and 11 produced deep green colored spots at Rf = 0.80 which was similar to the spot produced by standard pyrrolnitrin (Fig 2A). Prodigiosin was detected in fractions F3, F4 and F5 at Rf = 0.6 which produced a visible reddish pink band which was same as the standard (Fig 2B). Phenazine was not detected in any of the fractions. UV-VIS scan revealed that the fractions F10 and F11 produced two sharp peaks at 210 nm and 249 nm (Fig 3) which was similar to the standard pyrrolnitrin (217 nm and 251 nm). Fractions F3, F4 and F5 produced a sharp peak at 536 nm (visible region) which corresponded to the prodigiosin standard (Fig 4). Comparison of the HPLC profiles obtained for the purified compounds recovered from preparative TLC plates with the profiles of standard antibiotics revealed that a single major peak, which was obtained with the fractions F10 and F11 at retention time (RT) of 3.024 min and 3.022 min respectively was comparable to standard pyrrolnitrin (RT: 3.020 min) (Fig 5). The antibiotic, purified from fractions F3, F4 and F5 by preparative TLC, were combined prior to HPLC analysis. The sample produced two major peaks (RT: 8.268 min and 9.024 min) which was similar to that of the standard prodigiosin (RT: 7.813 min; and 9.018 min) (Fig 6). LC-ESI-MS analysis of the combined bioactive fractions F10 and F11 obtained by column chromatography revealed molecular ion peak at m/z 259 (M + 2H) + (Fig 7). This confirmed the presence of pyrrolnitrin in the extracellular culture extract of S. marcescens ETR17.
Fig 2. Detection of antibiotics in extracellular culture extract of the antagonistic S. marcescens strain ETR17 by thin layer chromatography.

(A) Pyrrolnitrin detection under UV light (254 nm) in the partially purified silica gel column eluted fractions F10 and F11. (B) Detection of prodigiosin in the partially purified silica gel column eluted fractions F3, F4 and F5 under visible light. [Standard prodigiosin (PDG) and pyrrolnitrin (PRN) are spotted in the extreme left in both (A) and (B)].
Fig 3. Detection of pyrrolnitrin in extracellular culture extract of the antagonistic S. marcescens strain ETR17 by UV-spectrophotometric analyses (between 200 nm to 400 nm).
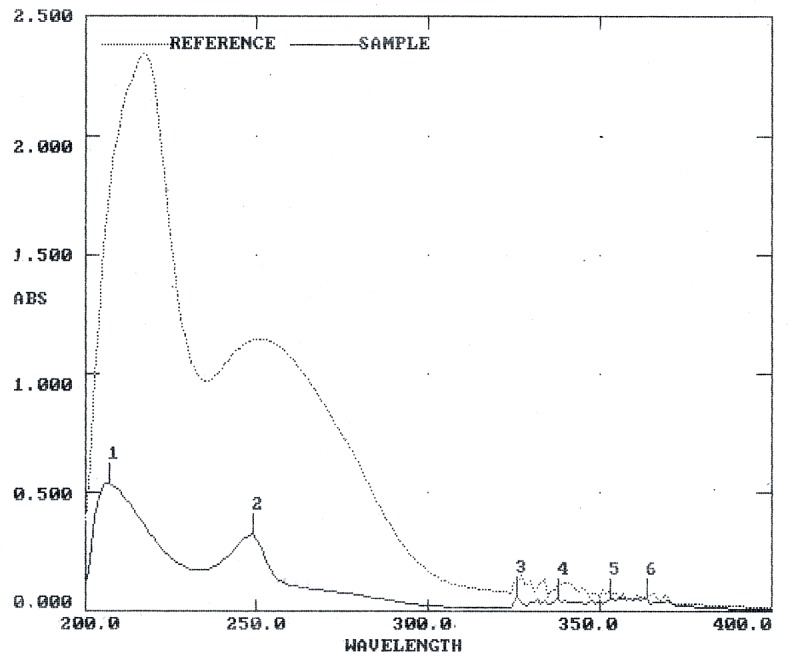
Pyrrolnitrin was purified by preparative thin layer chromatography from fraction F10 obtained previously from silica gel column chromatography of the crude culture extract. Two sharp peaks at 210 nm and 249 nm obtained with the partially purified compound were similar to the pyrrolnitrin standard (217 nm and 251 nm).
Fig 4. Detection of prodigiosin in extracellular culture extract of the antagonistic S. marcescens strain ETR17 by VIS-spectrophotometric analyses (between 500 nm to 600 nm).

Prodigiosin was purified by preparative thin layer chromatography from fraction F3 obtained previously from silica gel column chromatography of the crude culture extract. (A) A peak at 536 nm obtained with the partially purified compound. (B) Prodigiosin standard having absorbance maxima at 536 nm.
Fig 5. Detection of pyrrolnitrin in extracellular culture extract of the antagonistic S. marcescens strain ETR17 by high performance liquid chromatography.

Chromatograms generated by performing HPLC of the antifungal fraction at 225 nm eluted with 100% methanol (A) chromatogram of pyrrolnitrin standard (retention time = 3.020 min) (B) chromatogram of F10 (45% Ethyl Acetate-55% Petroleum Ether) (retention time = 3.024 min).
Fig 6. Detection of prodigiosin in extracellular culture extract of the antagonistic S. marcescens strain ETR17 by high performance liquid chromatography.

Chromatograms generated by performing HPLC of the combined bioactive fractions with absorbance maxima of 536 nm (A) chromatogram of prodigiosin standard (1st peak, retention time = 7.813 min; 2nd peak, retention time = 9.018) (B) chromatogram of prodigiosin sample pooled from F3 (10% Ethyl Acetate-90% Petroleum Ether) (1st peak, retention time = 8.266 min; 2nd peak, retention time = 9.02 min).
Fig 7. Detection of pyrrolnitrin in extracellular culture extract of the antagonistic S. marcescens strain ETR17 by liquid chromatography-electrospray ionization-mass spectrometry (LC-ESI-MS) analysis.

The combined column fractions F10 and F11 obtained previously from silica gel column chromatography of the crude culture extract was analysed using a gradient elution program with methanol and ammonium acetate buffer at a flow rate of 0.8 mL min-1 at 254 nm to obtain a major peak at retention time 14.85 minutes corresponding to pyrrolnitrin. Mass spectra showing molecular ion peak at m/z 259 (M + 2H)+ indicating pyrrolnitrin.
Biocontrol of root rot disease in tea
Treatment of tea seedlings with talc based formulation prepared from the isolate ETR17 three days prior to inoculation with the root rot pathogen R. solani at 10 g per kg soil reduced the disease development significantly under both sterile and non-sterile soil conditions when compared to control (Table 3). In contrast, the progression of disease remained unhindered and finally at 45 dapi, severe disease symptoms appeared in the control plants that were not treated with either ETR17 or thiophanate methyl but inoculated with the pathogen. The control set therefore recorded a high mean disease index value of 78.3 and 81.7 under sterile and non-sterile soil conditions respectively. However, in the ETR17 treated sets, the mean disease index increased slowly and reached its highest on 30th day (16.7 and 20.0 under sterile and non sterile conditions respectively) after which the roots showed signs of recovery which caused the disease index to decrease marginally (15.0 and 18.3 under sterile and non-sterile conditions respectively) by the 45th day. The fungicide recorded lower efficacy of disease control than ETR17 and also did not show any recovery during the period of observation. No significant difference in the overall efficacy of disease control between sterile (80.8%) and non-sterile (77.6%) soil conditions was noted over 45 dapi (Fig 8).
Table 3. Effect of powder formulation of biocontrol bacterial isolate ETR17 on root-rot disease incidence of tea plants (TS-520) during in vivo study.
| Treatments | Mean disease index | PEDC* | ||||
|---|---|---|---|---|---|---|
| 15 dapi** | 20 dapi | 25 dapi | 30 dapi | 45 dapi | 45 dapi | |
| Control (Sterilized soil) | 26.7±0.9a | 36.7±1.0a | 58.3±0.9a | 70.0±0.9a | 78.3±0.9a | 0 |
| Control (Unsterilized soil) | 28.3±0.4a | 50.0±0.8b | 63.3±0.7b | 73.3±0.5b | 81.7±0.9a | 0 |
| Fungicide (Sterilized soil) | 0 | 3.3±0.3c | 12.5±0.3c | 23.3±0.7ce | 25.0±0.6b | 68.1±0.5b |
| Fungicide (Unsterilized soil) | 0 | 6.7±0.4dc | 20.0±0.7d | 25.0±0.7c | 28.3±0.8b | 65.4±0.5b |
| ETR17 (Sterilized soil) | 0 | 5.0±0.5dc | 13.3±0.6ec | 16.7±0.7d | 15.0±0.9c | 80.8±0.8c |
| ETR17 (Unsterilized soil) | 0 | 8.3±0.6d | 15.0±0.8d | 20.0±0.4ed | 18.3±0.5c | 77.6±0.6d |
| LSD | 1.3 | 2.0 | 2.2 | 2.1 | 2.4 | 1.6 |
*PEDC = Percent efficacy of disease control
**dapi = days after pathogen inoculation
Means ± standard errorfollowed by same superscript letters do not differ significantly at P<0.05
Fig 8.

Root rot disease occurrence in tea (TS-520 variety) after 30 days of treatment by talc based formulation prepared with ETR17 in (A1) sterile soil ETR17 treated (A2) sterile soil untreated control (A3) sterile soil fungicide treated and (B1) unsterile soil ETR17 treated (B2) unsterile soil untreated control (B3) unsterile soil fungicide treated.
Plant growth promotion by bacterial formulation
Tea seedlings treated with talc based formulation of the biocontrol bacteria showed approximately 50% higher growth response under both sterile and non-sterile sterile conditions when compared to the untreated controls (Table 4). An increase in shoot and root length of tea seedlings by 6.0 cm and 4.7 cm respectively was recorded in treated plants under sterile soil conditions after 45 days of plantation. But in untreated control plants, shoot length increased by 3.0 cm and root length increased by 2.5 cm only under similar conditions. Similarly, under non-sterile conditions, significantly better shoot and root growth was recorded by the ETR17 treated plants than the respective control.
Table 4. Effect of powder formulation of biocontrol bacterial isolate ETR17 on the growth of tea plants (TS-520) during in vivo study.
| Treatment | Plant response | |||||
|---|---|---|---|---|---|---|
| Shoot length (cm) | Increase in shoot length (cm) | Root length (cm) | Increase in root length (cm) | |||
| (0 day) | (45 days) | (0 day) | (45 days) | |||
| Control (Sterilized soil) | 12.0±0.5a | 15.0±0.4a | 3.0 ±0.3a | 7.5 ±0.3a | 10.0±0.3a | 2.5 ±0.3a |
| Control (Unsterilized soil) | 11.6±0.3a | 15.0±0.6a | 3.4 ±0.5a | 10.0 ±0.5a | 12.0 ±0.4ab | 2.0±0.5a |
| ETR17 (Sterilized soil) | 12.0±0.4a | 18.0±0.5b | 6.0±0.4b | 8.5±0.2a | 13.2 ±0.4b | 4.7 ±0.4b |
| ETR17 (Unsterilized soil) | 12.8±0.5a | 19.2±0.3b | 6.4±0.3b | 10.0±0.7a | 13.8±0.5b | 3.8±0.2ab |
| LSD (0.5%) | 1.4 | 1.5 | 1.2 | 1.5 | 1.3 | 1.2 |
Means ± standard error followed by same superscript letters do not differ significantly at P<0.05
Survivability of bacterial cells in talc formulation
Survivability of ETR 17 cells in the talc based formulation was tested every one month for a period up to one year. The bacterial cell population remained almost stationary for the first six months with no significant changes when stored at room temperature. A reduction in bacterial count was noticed at the 7th month and this reducing trend continued until one year (Fig 9).
Fig 9. Viability of S. marcescens ETR17 isolate in talc formulation.

The bacterial cell population was measured for 12 months at 1 month time interval.
Sustainability of ETR17 in the rhizosphere
Results (Table 5) showed that the soil population was high after 45 days of application of the bacteria as formulation. The bacterial concentration was estimated to be 8x108 cfu g-1 and 9x108 cfu g-1 in the sterilized and unsterilized soil respectively. The polyclonal antibody raised against ETR17 showed negligible cross reactions as evident from the low ELISA values obtained with the unsterile soil antigen (untreated negative control).
Table 5. Determination of S. marcescens ETR17 concentration in soil by ELISA using soil antigens prepared from tea rhizosphere soil, treated with biocontrol formulations and ETR17 antiserum after 45 days of pathogen inoculationa.
| Treatment | Absorbance at 492nm | Bacterial concentration (cfu/g soil)c |
|---|---|---|
| Control (Sterilized soil) | 0.010 | 0 |
| Control (Unsterilized soil) | 0.166 | 1.0 x 108 |
| ETR17 (Sterilized soil) | 0.581 | 8.0 x 108 |
| ETR17 (Unsterilized soil) | 0.660 | 9.0 x 108 |
| Positive controlb ETR17 (whole cell) |
0.744 |
Bacterial concentration (cfu/mL) 1.2 x 109 |
aETR17 application was done three days prior to pathogen inoculation. Primary antiserum dilution: 1:100; secondary antibody (goat anti-rabbit IgG-HRP conjugate) dilution: 1:10,000.
bWhole cell antigen prepared with bacterial cells cultured in NB for 48 hours served as positive control.
cBacterial concentrations were determined using standard curve plotted using varying bacterial concentrations against the corresponding ELISA values of the positive control
This study confirmed the presence of ETR17 in tea rhizosphere on application of talc formulation in both sterilized and unsterilized soil sets for at least 45 days.
Hemolytic activity
The test for hemolytic activity on blood supplemented TSA plates revealed that the isolate ETR17 lacked the hemolytic activity since no halo formation occurred around the bacterial colonies.
Discussion
A total of 200 bacterial strains were isolated from tea rhizosphere from 15 tea growing fields, among which 35 strains showed antifungal activity. The isolate ETR17, which showed best antagonistic activity against eight fungal pathogens of tea, was identified as S. marcescens after analyzing both the phenotypic characters and 16S rRNA gene sequence. S. marcescens strains unlike the S. plymuthica and S. liquefaciens does not produce gas in glucose, shows lysine and ornithine decarboxylation but not arginine decarboxylation and does not ferment L-arabinose and xylose [67]. Additionally, the 16S rRNA gene sequence of ETR17 showed 99% similarity with other S. marcescens gene sequences available in the NCBI GenBank which confirmed the phenotypic data.
Production of antifungal metabolites by biocontrol agents is strongly correlated with their biocontrol activity. Chitinases, which had earlier been considered as a major factor of biocontrol [68] along with other hydrolytic enzymes, degrade fungal cell walls and inhibit fungal growth [69, 70]. ETR17 produced chitinase, protease and lipase but did not produce cellulose and pectinase. Serratia is one of the most competent producers of chitinases, whose production, activity and gene regulation are well studied [71]. Purified chitinases from S. plymuthica [72], S. proteamaculans [73] and S. marcescens [74] are reported to be antagonistic to different pathogenic fungi. But in spite of the strong antifungal activity, factors other than chitinases are considered more essential for biocontrol of plant diseases [75] indicating that chitinases may form only a part of the multi-component mechanism which underlies biocontrol by Serratia strains in the environment [76]. Inability of ETR17 to produce pectinase and HCN may be regarded as an advantageous trait because the production of pectinase is considered as an undesirable feature of plant beneficial bacteria and HCN inhibits plant growth and yield due to the interference with cytochrome oxidation [48, 56].
The role of iron-chelating siderophore produced by the rhizobacteria in biocontrol is well documented [7, 77]. Siderophore producing antagonistic bacteria like Serratia, Bacillus, Pseudomonas etc. exhibit better rhizosphere competence and suppress plant disease by inhibiting the growth or the metabolic activity of plant fungal pathogens by sequestering iron since fungal siderophores have lower affinity [78, 79, 80]. The present isolate was capable of producing both catecholate and hydroxamate type of siderophore. Additionally, ETR17 produced IAA, which is considered as the best-known hormone produced by the plant-associated microorganisms [81].
The nature of antagonism involved in the interaction between ETR 17 and R. solani was observed by scanning electron microscopy. The fungal mycelium at the interaction zone in dual cultures was distorted, showing bulging, lysis and bursting of the hyphae. Abnormal hyphal structures caused due to cell wall degradation were prominent. Ordentlich et al. [82] also observed severe mycelial deformities like holes in the hyphae at the interaction zone of bacterial antagonist S. marcescens and the pathogen Sclerotium rolfsii. The mycelial and spore deformities of Aspergillus parasiticus NFRI-95 caused by S. marcescens JPP1 crude culture filtrate was observed under SEM by Wang et al. [74]. Enzymatic degradation of fungal cell walls leading to loss of protoplasm is considered as a major antagonistic mechanism involved in biocontrol activity of many bacteria [83, 84]. The present strain produced several lytic enzymes, particularly chitinase, responsible for lysis of fungal cell wall. However, further studies are required to ascertain the exact mechanism used by this strain for its biocontrol action. Biofilm production, observed in several biocontrol bacteria, is considered to be a beneficial feature for effective colonization on root surface [85–88]. The present strain ETR 17 formed biofilm when tested after 24 and 48 h of incubation. Better biofilm production was noted in LB medium compared to M9YE medium. Biofilm formation was found to vary depending on the medium in S. plymuthica HRO-C48 [22].
Further studies on biological basis of the antagonism shown by ETR17 revealed that the strain produced the antibiotics pyrrolnitrin and prodigiosin. These were initially detected in the culture filtrate extract by TLC where they showed same migration as the standards. Spectroscopic analysis of the column fractionated extract of the culture filtrate further confirmed the presence of these antibiotics. Pyrrolnitrin is a strong antifungal antibiotic produced by a narrow range of gram negative bacteria such as Pseudomonas, Burkholdaria, Serratia and Enterobacter [88–90]. It has been correlated with the potential of some of these bacteria to control fungal diseases in plants [88, 17]. Pyrrolnitrin has been reported from Serratia marcescens having absorbance maxima between 210–225 nm and being involved in pathogen suppression [91, 16]. Pyrrolnitrin from particular Serratia strains have been shown to be directly linked to its in vitro antifungal activity [92], biocontrol capacity [16] and more important for the antifungal action than its other antagonistic properties [70]. Transcriptional response studies with biocontrol Serratia strain grown in presence of R. solani in vitro, revealed significantly increased expression of genes related to the biosynthesis of the antibiotic pyrrolnitrin [93]. The other antibiotic, prodigiosin, detected during this study in the culture filtrate of ETR17, is well known as the tripyrrole red pigmented antibiotic that is produced by three species of Serratia namely S. marcescens, S. rubidaea and S. plymuthica [94]. It contributes to survival under competition and possesses antifungal, antibacterial, antiprotozoal, immunosuppressive and anticancer properties [76]. Prodigiosin purified from S. marcescens controlled Black Sigatoka in banana by a synergistic action with purified chitinases [95].
Results of the glasshouse experiments conducted by application of talc based formulation of ETR17 were consistent with the in vitro studies. Incidence of root rot in bacteria treated tea plants were considerably lower in comparison to untreated control as well as the fungicide treated sets. The bacterial population in the bioformulation remained unchanged upto six months of storage. Additionally, ETR17 formulation also increased the root and shoot length of the tea seedlings under both sterile and unsterile soil conditions in comparison to the untreated controls. Serratia species are known for their ability to produce a multitude of secondary metabolites and extracellular enzymes that directly contribute to establish themselves by outcompeting other microorganisms when colonizing their respective niche [96]. There are reports on the biocontrol activity of S. marcescens against R. solani causing damping off in cyclamen [97]; Phytophthora parasitica causing root rot disease in citrus [98]; Pyricularia oryzae causing blast disease in rice [28]; Fusarium oxysporum causing wilt in Banana [99]; and Fomes lamaoensis causing brown root rot in tea [100]. Our strain produced chitinase, siderophore and multiple antibiotics in vitro but further studies are required to confirm how these factors either individually or synergistically were actually involved in the biocontrol action in vivo. Characterization of biocontrol strains most often includes evaluation of plant growth promoting traits whose presence increases the overall quality of a strain [76]. S. marcescens have been used widely in several studies in a wide range of crops but such reports involving tea are few. In a previous study, a phosphate solubilizing strain of S. marcescens promoted growth and induced resistance in tea plants against brown root rot caused by Fomes lamaoensis. Our strain did not show phosphate solubilizing ability; but it still promoted plant growth which was not unexpected as the bacteria showed IAA production during in vitro tests. However, further studies are necessary to confirm whether IAA was responsible for the growth promotion in tea plants. Other studies have shown that Serratia strains possessed growth promoting attributes such as phosphate solubilizing ability and IAA production and exhibited beneficial effect on several plants such as coconut, paddy, cowpea and chickpea [101, 102].
Sustainability of ETR17 in the soil environment when applied as talc based formulation was studied by ELISA. ELISA is considered more reliable compared to other techniques because the raised polyclonal antiserum shows no non-specific reactions with other bacterial species in the environment [103]. Literature studies suggest that although ELISA was used for studying the presence of several bacterial populations in soil and bacteria-plant root associations but the use of this technique for detection of biocontrol strain S. marcescens in soil upon in vivo application was done for the first time in our study. Results showed that ETR 17 population remained high in the soil in both sterilized and unsterilized soil treatments for 45 days. In a previous study, S. marcescens B2 which suppressed rice sheath blight caused by Rhizoctonia solani remained in rhizosphere soil at a concentration of 108 cfu g-1soil for more than 4 weeks under glass house conditions [104]. The present strain ETR17 remained in soil at a concentration of 8x108 cfu g-1 and 9x108 cfu g-1 in the sterilized and unsterilized soil respectively which shows that this strain has potential as an effective and persistent biological control agent for root rot in tea.
Some of the bacterial biocontrol agents including Serratia can enter into mutualistic interactions with plant and human hosts [105, 106]. Therefore, it is essential to appraise the risk of every potential biocontrol bacteria in order to avoid using potential human pathogens as agricultural inputs. Pathogenic strains of Serratia are reported to produce hemolysins [107, 108], therefore hemolytic activity was tested in this study which showed negative result.
Conclusions
A pyrrolnitrin and prodigiosin producing S. marcescens strain ETR17, which was isolated from tea rhizosphere was effective in suppressing root rot disease caused by R. solani in tea plants. The isolate exhibited biocontrol traits such as broad spectrum antifungal activity and production of lytic enzymes, HCN, IAA, siderophore and antibiotics. The talc formulations of the bacterial isolate significantly reduced the root rot disease in tea and stimulated plant growth in vivo. In addition, the isolate did not show hemolytic activity which is a general feature of pathogenic S. marcescens strains. Therefore, after considering the safety and biocontrol potential of ETR17, it may be suggested that the bacterium can be used in management of fungal infections of tea after field trials. The use of this bacterial strain as a biocontrol agent to increase plant protection seems to be a promising approach for controlling fungal diseases, as compared to contemporary chemical agents used in agriculture.
Supporting information
(DOCX)
Data Availability
All relevant data are within the paper and its Supporting Information files. All GenBank Sequence files are available from the GenBank database (accession number JX566992).
Funding Statement
This work was supported by: 1. Council of Scientific & Industrial Research (CSIR), India (www.csirhrdg.res.in/) Grant No. 9/285(0064)/2013, EMR I dated 14.03.2013; 2. University Grants Commission (UGC), India (www.ugc.ac.in/) Grant No. F.25-1/2014-15(BSR)/7-132/2007(BSR) dated 07.10.2015. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Saha D, Dasgupta S, Saha A. Antifungal Activity of Some Plant Extracts Against Fungal Pathogens of Tea (Camellia sinensis.).Pharma Biol. 2005; 43(1):87–91. [Google Scholar]
- 2.Chakraborty U, Chakraborty BN, Basnet M, Chakraborty AP. Evaluation of Ochrobactrum anthropi TRS‐2 and its talc based formulation for enhancement of growth of tea plants and management of brown root rot disease. J Appl Microbiol. 2009; 107(2):625–34. doi: 10.1111/j.1365-2672.2009.04242.x [DOI] [PubMed] [Google Scholar]
- 3.Saha D, Kumar R, Ghosh S, Kumari M, Saha A. Control of foliar diseases of tea with Xanthium strumarium leaf extract. Ind Crops Prod. 2012; 37(1):376–82. [Google Scholar]
- 4.Dias MC. Phytotoxicity: An overview of the physiological responses of plants exposed to fungicides. J Bot. 2012; 2012:1–4. doi: 10.1155/2012/135479 [Google Scholar]
- 5.Gerbore J, Benhamou N, Vallance J, Le Floch G, Grizard D, Regnault-Roger C, et al. Biological control of plant pathogens: advantages and limitations seen through the case study of Pythium oligandrum. Environ Sci Pollut Res. 2014; 21(7):4847–60. doi: 10.1007/s11356-013-1807-6 [DOI] [PubMed] [Google Scholar]
- 6.Selim S. Allele-specific real-time PCR for quantification and discrimination of sterol 14α-demethylation-inhibitor-resistant genotypes of Mycosphaerella graminicola. Journal of Plant Pathology 2009; 91(2):391–400. http://dx.doi.org/10.4454/jpp.v91i2.969. [Google Scholar]
- 7.Haas D, Défago G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nature Rev Microbiol. 2005; 3(4):307–19. doi: 10.1038/nrmicro1129 . [DOI] [PubMed] [Google Scholar]
- 8.Lugtenberg B, Kamilova F. Plant-growth-promoting rhizobacteria. Annu Rev Microbiol. 2009; 63:541–56. doi: 10.1146/annurev.micro.62.081307.162918 [DOI] [PubMed] [Google Scholar]
- 9.Samain E., van Tuinen D. and Selim S., The plant-growth-promoting rhizobacterium Paenibacillus sp. strain B2stimulates wheat defense mechanisms against septoria leaf blotch and root colonization by Curtobacterium plantarum. Biological control 2017; 114: 87–96. [Google Scholar]
- 10.Kumar RS, Ayyadurai N, Pandiaraja P, Reddy AV, Venkateswarlu Y, Prakash O, et al. Characterization of antifungal metabolite produced by a new strain Pseudomonas aeruginosa PUPa3 that exhibits broad-spectrum antifungal activity and biofertilizing traits. J Appl Microbiol. 2005; 98(1):145–54. doi: 10.1111/j.1365-2672.2004.02435.x [DOI] [PubMed] [Google Scholar]
- 11.Ge YH, Pei DL, Zhao YH, Li WW, Wang SF, Xu YQ. Correlation between antifungal agent phenazine-1-carboxylic acid and pyoluteorin biosynthesis in Pseudomonas sp. M18. Curr Microbiol. 2007. April 1; 54(4):277–81. doi: 10.1007/s00284-006-0317-x [DOI] [PubMed] [Google Scholar]
- 12.Höfte M, Altier N. Fluorescent pseudomonads as biocontrol agents for sustainable agricultural systems. Res Microbiol. 2010; 161(6):464–71. doi: 10.1016/j.resmic.2010.04.007 [DOI] [PubMed] [Google Scholar]
- 13.Selim S., Negrel J., Wendehenne D., Ochatt S., Gianinazzi S. and van Tuinen D. Stimulation of defence reactions in Medicago truncatula by antagonistic lipopeptides from Paenibacillus sp. strain B2. Applied Environmental Microbiology. 2010; 76(22):7420–7428.doi: 10.1128/AEM.00171-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Raaijmakers JM, Vlami M, de Souza JT. Antibiotic production by bacterial biocontrol agents. Antonie van Leeuwenhoek. 2002; 81(1):537–47. [DOI] [PubMed] [Google Scholar]
- 15.Duffy B, Keel C, De´fago G. 2004. Potential role of pathogen signaling in multitrophic plant-microbe interactions involved in disease protection. Appl Environ Microbiol. 2004; 70(3):1836–42. doi: 10.1128/AEM.70.3.1836-1842.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Liu X, Bimerew M, Ma Y, Muller H, Ovadis M, Eberl L, et al. Quorum-sensing signaling is required for production of the antibiotic pyrrolnitrin in a rhizospheric biocontrol strain of Serratia plymuthica. FEMS Microbiol Lett. 2007; 270(2):299–305. doi: 10.1111/j.1574-6968.2007.00681.x [DOI] [PubMed] [Google Scholar]
- 17.Costa R, Van Aarle IM, Mendes R, Van Elsas JD. Genomics of pyrrolnitrin biosynthetic loci: evidence for conservation and whole-operon mobility within gram negative bacteria. Environ Microbiol. 2009; 11(1):159–75. doi: 10.1111/j.1462-2920.2008.01750.x [DOI] [PubMed] [Google Scholar]
- 18.Someya N, Akutsu K. Indigenous bacteria may interfere with the biocontrol of plant diseases.Naturwissenschaften. 2009; 96(6):743–7. doi: 10.1007/s00114-009-0524-y . [DOI] [PubMed] [Google Scholar]
- 19.Chen Y, Shen X, Peng H, Hu H, Wang W, Zhang X. Comparative genomic analysis and phenazine production of Pseudomonas chlororaphis, a plant growth-promoting rhizobacterium. Genom Data. 2015; 4:33–42. doi: 10.1016/j.gdata.2015.01.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Samain E., van Tuinen D. and Selim S., The plant-growth-promoting rhizobacterium Paenibacillus sp. strain B2stimulates wheat defense mechanisms against septoria leaf blotch and root colonization by Curtobacterium plantarum. Biological control 2017; 114: 87–96 [Google Scholar]
- 21.Someya N, Nakajima A, Hirayae K, Hibi TA, Akutsu K. Synergistic antifungal activity of chitinolytic enzymes and prodigiosin produced by biocontrol bacterium, Serratia marcescens strain BZ against gray mold pathogen, Botrytis cinerea. J Gen Plant Pathol. 2001; 67(4):312–7. [Google Scholar]
- 22.Muller H, Westendorf C, Leitner E, Chernin L, Riedel K, Schmidt S,et al. Quorum-sensing effects in the antagonistic rhizosphere bacterium Serratia plymuthica HRO-C48. FEMS Microbiol Ecol. 2009; 67(3):468–78. doi: 10.1111/j.1574-6941.2008.00635.x [DOI] [PubMed] [Google Scholar]
- 23.Compant S, Duffy B, Nowak J, Clement C, Barka EA. Use of plant growth-promoting bacteria for biocontrol of plant diseases: principles, mechanisms of action, and future prospects. Appl Environ Microbiol. 2005; 71(9):4951–9. doi: 10.1128/AEM.71.9.4951-4959.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Berg G. Plant–microbe interactions promoting plant growth and health: perspectives for controlled use of microorganisms in agriculture. Appl Microbiol Biotechnol. 2009; 84(1):11–8. doi: 10.1007/s00253-009-2092-7 [DOI] [PubMed] [Google Scholar]
- 25.Philippot L, Raaijmakers JM, Lemanceau P, van der Putten WH. Going back to the roots: the microbial ecology of the rhizosphere.Nat Rev Microbiol. 2013; 11(11):789–99. doi: 10.1038/nrmicro3109 [DOI] [PubMed] [Google Scholar]
- 26.Rodrigo Mendes, Garbeva Paolina, Raaijmakers Jos M. The rhizosphere microbiome: significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol Rev. 2013; 37(5): 634–63. doi: 10.1111/1574-6976.12028 [DOI] [PubMed] [Google Scholar]
- 27.Latz E, Eisenhauer N, Scheu S, Jousset A. Plant identity drives the expression of biocontrol factors in a rhizosphere bacterium across a plant diversity gradient. Funct Ecol. 2015; 29(9):1225–34. [Google Scholar]
- 28.Jaiganesh V, Eswaran A, Balabaskar P, Kannan C.2007. Antagonistic activity of Serratia marcescens against Pyricularia oryzae. Not Bot Horti Agrobot Cluj Napoca. 2007; 35(2):48. [Google Scholar]
- 29.Bashan Y, de-Bashan LE, Prabhu SR. Superior Polymeric Formulations and Emerging Innovative Products of Bacterial Inoculants for sustainable Agriculture and the Environment. Chapter Agriculturally Important Microorganisms. 2016; pp 15–46. [Google Scholar]
- 30.Whipps JM. Microbial interactions and biocontrol in the rhizosphere. J Exp Bot. 2001; 52(suppl 1):487–511. . [DOI] [PubMed] [Google Scholar]
- 31.Pal KK, Gardener BM. Biological control of plant pathogens. The Plant Health Instructor. 20062:1117–42. doi: 10.1094/PHI-A-2006-1117-02 APSnet. p.25. [Google Scholar]
- 32.Pang Y, Liu X, Ma Y, Chernin L, Berg G, Gao K. Induction of systemic resistance, root colonisation and biocontrol activities of the rhizospheric strain of Serratia plymuthica are dependent on N-acyl homoserine lactones. Eur J Plant Pathol. 2009; 124(2):261–68. [Google Scholar]
- 33.Saha A, Dasgupta S, Saha D. Discovery of Curvularia eragrostidis on tea (Camellia sinensis (L.) O. Ktze.) leaves from clonal-cutting nurseries in North Bengal. Environ Ecol. 2001; 19(4):846–48. [Google Scholar]
- 34.Dasgupta S, Saha D, Saha A. Levels of common antigens in determining pathogenicity of Curvularia eragrostidis in different tea varieties. J Appl Microbiol. 2005; 98(5):1084–92. doi: 10.1111/j.1365-2672.2005.02538.x [DOI] [PubMed] [Google Scholar]
- 35.Mandal P, Das L, Saha D, Saha A. Screening, isolation and pathogenicity of Rhizoctonia solani associated with seeds of tea. Indian Phytopathol. 2006; 59:384. [Google Scholar]
- 36.Saha A, Mandal P, Dasgupta S, Saha D. Influence of culture media and environmental factors on mycelial growth and sporulation of Lasiodiplodia theobromae (Pat.) Griffon & Maubl. J Environ Biol. 2008; 29(3):407–10. [PubMed] [Google Scholar]
- 37.Ray S.K., Mukhopadhyay D. A study on physical-chemical properties of soils under different tea growing regions of West Bengal (India). Int JAgri.Sci. 2012; 4(8): 325–329. [Google Scholar]
- 38.Purkayastha GD, Saha A, Saha D. Characterization of antagonistic bacteria isolated from tea rhizosphere in sub-Himalayan West Bengal as potential biocontrol agents in tea. J Mycol Plant Pathol. 2010; 40(1):27–37. [Google Scholar]
- 39.Saha D, Purkayastha GD, Ghosh A, Isha M, Saha A. Isolation and characterization of two new Bacillus subtilis strains from rhizosphere of eggplant as potential biocontrol agents. J Plant Pathol. 2012; 94(1):109–18. [Google Scholar]
- 40.Chandramouli B. Global status of tea diseases and management strategies Global advances in tea science (Ed.: Jain NK). Aravali Books International Publication, New Delhi: 1999:647–58. [Google Scholar]
- 41.Sarmah KC. Disease of tea and associated crops in north east Memorandum Indian Tea association, Tocklai Experimental Station, Assam, India: 1960:26. [Google Scholar]
- 42.Singh ID. Planter’s guide to tea culture and manufacture NB Modern Agencies; 2005. [Google Scholar]
- 43.Barrow GI, Feltham RK. Cowan and Steel’s Manual for the Identification of medical bacteria 3rd ed. Cambridge University Press, New York: 1993. [Google Scholar]
- 44.Aneja KR. Experiments in Microbiology Plant Pathology and Biotechnology. 4th ed. New Age International (P) Limited, Publishers, New Delhi, India: 2003. [Google Scholar]
- 45.Gomes LH, Duarte KMR, Andrino FG, Tavares FCA. A simple method for DNA isolation from Xanthomonas spp. Sci Agric. 2000; 57(3):553–5. [Google Scholar]
- 46.Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990; 215(3):403–10. doi: 10.1016/S0022-2836(05)80360-2 . [DOI] [PubMed] [Google Scholar]
- 47.Bargabus RL, Zidack NK, Sherwood JE, Jacobsen BJ. Characterization of systemic resistance in sugar beet elicited by a non-pathogenic, phylosphere-colonizing Bacillus mycoides, biological control agents. Physiol Mol Plant Pathol. 2002; 61(5):289–98. [Google Scholar]
- 48.Cattelan AJ, Hartel PG, Fuhrmann JJ. Screening for plant growth-promoting rhizobacteria to promote early soybean growth. Soil Sci Soc Am J. 1999; 63(6):1670–80. [Google Scholar]
- 49.Miller JH. Experiments in molecular genetics 2nd ed. ColdSpring Harbor, New York: Cold Spring Harbor Laboratory Press; 1974. [Google Scholar]
- 50.Smibert RM, Krieg NR. Phenotypic characterization In: Gerhardt PRG, Murray E, Wood WA and Krieg NR editors. Methods for General and Molecular Biology. Washington DC: American Society for Microbiology; 1994; p. 611–54. [Google Scholar]
- 51.Patten CL, Glick BR. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl Environ Microbiol. 2002; 68(8):3795–801. doi: 10.1128/AEM.68.8.3795-3801.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Pikovskaya RI. Mobilization of phosphorous in soil in connection with vital activity of some microbial species. Mikrobiologiya. 1948; 17:362–70. [Google Scholar]
- 53.Schwyn B, Neilands JB. Universal chemical assay for the detection and determination of siderophores. Anal Biochem. 1987; 160(1):47–56. [DOI] [PubMed] [Google Scholar]
- 54.Arnow LE. Colorimetric determination of the components of 3, 4 dihydroxyphenylalanine-tyrosine mixtures. J Biol Chem. 1937; 118(2):531–7. [Google Scholar]
- 55.Baakza A, Vala AK, Dave AP, Dube HC. A comparative study of siderophore production by fungi from marine and terrestrial habitats. J Exp Mar Biol Ecol. 2004; 311(1):1–9. [Google Scholar]
- 56.Bakker AW, Schippers B. Microbial cyanide production in the rhizosphere in relation to potato yield reduction and Pseudomonas spp-mediated plant growth-stimulation. Soil Biol Biochem. 1987; 19(4):451–7. [Google Scholar]
- 57.Harvey J, Keenan KP, Gilmour A. Assessing biofilm formation by Listeria monocytogenes strains. Food Microbiol. 2007; 24(4):380–92. doi: 10.1016/j.fm.2006.06.006 . [DOI] [PubMed] [Google Scholar]
- 58.Levitch ME, Stadtman ER.A study of the biosynthesis of phenazine-1-carboxylic acid. Arch Biochem Biophys. 1964; 106:194–9. [DOI] [PubMed] [Google Scholar]
- 59.Shanahan P, O’Sullivan DJ, Simpson P, Glennon JD, O’Gara F. Isolation and characterization of an antibiotic-like compound from a fluorescent pseudomonad and investigation of physiological parameters influencing its production. Appl Environ Microbiol.1992; 58:353–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Nandakumar R, Basu S, Viswanathan R, Sheela J, Raguchander T, Samiyappan R. A new bio-formulation containing plant growth promoting rhizobacterial mixture for the management of sheath blight and enhanced grain yield in rice. Biocontrol. 2001; 46(4):493–510. doi: 10.1023/A:1014131131808 [Google Scholar]
- 61.Soares ACF, Sousa CDS, Garrido MDS, Perez JO. Production of Streptomycete inoculum in sterilized rice. Sci Agric. 2007; 64(6):641–4. [Google Scholar]
- 62.Kobriger KM, Hagedorn DJ, Stevenson WR. Analysis of the snap bean root rot potential of Wiscosin fields. Article No. A3242, University of Wisconsin- System Board of Reagents and University of Wisconsin- Extension, Co-operative Extension, Madison, Wisconsin. 1998.
- 63.Elder BL, Boraker DK, Fives-Taylor PM. Whole-bacterial cell Enzyme-Linked Immunosorbent assay for Streptococcus sanguis fimbrial antigens. J Clin Microbiol. 1982; 16(1):141–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Sakai M, Atsuta S, Kobayashi M. Diagnostic application of monoclonal antibodies against the heat extract antigen of Renibacterium salmoninarum, the causative agent of bacterial kidney disease. Asian Fish Sci. 1991; 4:23–30. [Google Scholar]
- 65.Tsuchiya K, Takahashi Y, Shohara K, Homma Y, Suzui T. Rapid and simple ELISA procedure for the specfic detection of Pseudomonas cepacia in soil. Ann Phytopath Soc Jpn. 1991; 57(2):196–202. [Google Scholar]
- 66.Gerhardt P, Murray R, Wood W, Krieg N. In: Methods for General and Molecular Bacteriology. Washington D.C.: American Society of Microbiology; 1994. p. 21–42. [Google Scholar]
- 67.Grimont PAD, Grimont F. The genus Serratia. Annu Rev Microbiol. 1978; 32(1): 221–48. doi: 10.1146/annurev.mi.32.100178.001253 [DOI] [PubMed] [Google Scholar]
- 68.Inbar J, Chet I. The role of recognition in the induction of specific chitinases during mycoparasitism by Trichoderma harzianurn. Microbiology. 1995; 141(11):2823–29. doi: 10.1099/13500872-141-11-2823 [DOI] [PubMed] [Google Scholar]
- 69.Kobayashi DY, Reedy RM, Bick JA, Oudemans PV. Characterization of chitinase gene from Stenotrophomonas maltophilia strain 34S1 and its involvement in biological control. Appl Environ Microbiol. 2002; 68(3):1047–54. doi: 10.1128/AEM.68.3.1047-1054.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Ovadis M, Liu X, Gavriel S, Ismailov Z, Chet I, Chernin L. The global regulator genes from biocontrol strain Serratia plymuthica IC1270: cloning, sequencing, and functional studies. J Bacteriol. 2004; 186(15):4986–93. doi: 10.1128/JB.186.15.4986-4993.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Gutierrez-Roman MI, Dunn MF, Tinoco-Valencia R, Holguin-Melendez F, Huerta-Palacios G, Guillen-Navarro K. Potentiation of the synergistic activities of chitinases Chi A, ChiB and ChiC from Serratia marcescens CFFSUR-B2 by chitobiase (Chb) and chitin binding protein (CBP). World J Microbiol Biotechnol 2014; 30(1):33–42. doi: 10.1007/s11274-013-1421-2 [DOI] [PubMed] [Google Scholar]
- 72.Frankowski F, Lorito M, Scala F, Schmid R. Purification and properties of two chitinolytic enzymes of Serratia plymuthica HRO-C48. Arch Microbiol. 2001; 1176: 421–26. doi: 10.1007/s002030100347 doi [DOI] [PubMed] [Google Scholar]
- 73.Mehmood MA, Xiao X, Hafeez FY, Gai Y,Wang F. Purification and characterization of a chitinase from Serratia proteamaculans. World J Microbiol Biotechnol. 2009; 25(11):1955–61. [Google Scholar]
- 74.Wang K, Yan P-S, Cao L-X, Ding Q-L, Shao C, Zhao T-F. Potential of chitinolytic Serratia marcescens strain JPP1 for biological control of Aspergillus parasiticus and aflatoxin. BioMed Research International. 2013; Article ID 397142, 7, paged 2013. doi: 10.1155/2013/397142 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Kamensky M, Ovadis M, Chet I, Chernin L. Soil-borne strain IC14 of Serratia plymuthica with multiple mechanisms of antifungal activity provides biocontrol of Botrytis cinerea and Sclerotinia sclerotiorum diseases. Soil Biol Biochem. 2003; 35(2):323–31. [Google Scholar]
- 76.Saha D, Purkayastha GD, Saha A. Biological control of plant diseases by Serratia species: A review or a case study In: Goyal Aakash and Maheshwari Priti editors. Frontiers on Recent Developments in Plant Science. Bentham Science Publishers; 2012; 1:99–115 [Google Scholar]
- 77.Sahu GK, Sindhu SS. Disease control and plant growth promotion of green gram by siderophore producing Pseudomonas sp. Res J Microbiol. 2011; 6(10):735–49. [Google Scholar]
- 78.Press CM, Lopper JE, Kloepper JW. Role of iron in rhizobacteria mediated induced systemic resistance of cucumber. Phytopathology. 2001; 91(6):593–8. doi: 10.1094/PHYTO.2001.91.6.593 [DOI] [PubMed] [Google Scholar]
- 79.Sayyed RZ, Badgujar MD, Sonawane HM, Mhaske MM, Chincholkar SB. Production of microbial iron chelators (siderophore) by fluorescent pseudomonads. Indian J Biotechnol. 2005; 4:484–90. [Google Scholar]
- 80.Shanmugam V., Atri K., Gupta S., Kanoujia N., Naruka D.S. Selection and differentiation of Bacillus spp. antagonistic to Fusarium oxysporum f.sp. lycopersici and Alternaria solani infecting tomato. Folia Microbiol. 2011;56(2):170–7. doi: 10.1007/s12223-011-0031-3 [DOI] [PubMed] [Google Scholar]
- 81.Kim YC, Leveau J, Gardener BB, Pierson EA, Pierson LS, Ryu CM. The multifactorial basis for plant health promotion by plant-associated bacteria. Appl Environ Microbiol. 2011; 77(5):1548–55. doi: 10.1128/AEM.01867-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Ordentlich A, Elad Y, Chet I. The role of chitinase of Serratia marcescens in biocontrol of Sclerotium rolfsii. Phytopathology. 1988; 78(1):84–88. [Google Scholar]
- 83.Lim HS, Kim YS, Kim SD. Pseudomonas stutzeri YPL-1 genetic transformation and antifungal mechanism against Fusarium solani, an agent of plant root rot. Appl Environ Microbiol. 1991; 57(2):510–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Kim PI, Chung KC. Production of an antifungal protein for control of Colletotrichum lagenarium by Bacillusamyloliquefaciens MET 0908. FEMS Microbiol Lett. 2004; 234(1):177–83. doi: 10.1016/j.femsle.2004.03.032 [DOI] [PubMed] [Google Scholar]
- 85.Bais HP, Ray F, Vivanco JM. Biocontrol of Bacillus subtilis against infection of arabidopsis roots by Pseudomonas syringae is facilitated by biofilm formation and surfactin production. Plant Physiol. 2004; 134(1):307–19. doi: 10.1104/pp.103.028712 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Timmusk S., Grantcharova N., Wagner EGH. Paenibacillus polymyxa invades plant roots and forms biofilms. Appl. Environ Microbiol. 2005; 71(11):7292–300. doi: 10.1128/AEM.71.11.7292-7300.2005 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Danhorn T, Fuqua C. Biofilm Formation by Plant-Associated Bacteria. Annurev Micro. 2007; 61:401–22. doi: 10.1146/annurev.micro.61.080706.093316 . [DOI] [PubMed] [Google Scholar]
- 88.Chernin L, Brandis A, Ismailov Z, Chet I. Pyrrolnitrin production by an Enterobacter agglomerans strain with a broad spectrum of antagonistic activity towards fungal and bacterial phytopathogens. Curr Microbiol. 1996; 32(4):208–12. [Google Scholar]
- 89.Kirner S, Krauss S, Sury G, Lam S, Ligon J, van Pée K. The non-haem chloroperoxidase from Pseudomonas fluorescens and its relationship to pyrrolnitrin biosynthesis. Microbiology.1996; 142(8):2129–35. doi: 10.1099/13500872-142-8-2129 [DOI] [PubMed] [Google Scholar]
- 90.Hammer PE, Hill DS, Lam ST, Van Pee KH, Ligon JM.Four genes from Pseudomonas fluorescens that encode the biosynthesis of pyrrolnitrin. Appl Environ Microbiol.1997; 63(6): 2147–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Fuente LDL, Thomashow L, Weller D, Bajsa N, Quagliotto L, Chernin L,et al. Pseudomonas fluorescens UP61 isolated from birdsfoot trefoil rhizosphere produces multiple antibiotics and exerts a broad spectrum of biocontrol activity. Eur J Plant Pathol. 2004; 110(7):671–81. [Google Scholar]
- 92.Meziane H, Gavriel S, Ismailov Z, Chet I, Chernin L, Hofte M. Control of green and blue mould on orange fruit by Serratia plymuthica strains IC14 and IC1270 and putative modes of action. Postharvest Biol Technol. 2006; 39(2):125–33. [Google Scholar]
- 93.Neupane S, Finlay RD, Alström S, Elfstrand M, Högberg N. Transcriptional responses of the bacterial antagonist Serratia plymuthica to the fungal phytopathogen Rhizoctonia solani. Environ Microbiol Rep. 2015; 7(1):123–7. doi: 10.1111/1758-2229.12203 [DOI] [PubMed] [Google Scholar]
- 94.Thomson NR, Crow MA, McGowan SJ, Cox A, Salmond GP. Biosynthesis of carbapenem antibiotic and prodigiosin pigment in Serratia is under quorum sensing control. Mol Microbiol. 2000; 36(3):539–56. [DOI] [PubMed] [Google Scholar]
- 95.Gutierrez-Roman MI, Holguin-Melendez F, Dunn MF, Tinoco-Valencia R, Huerta-Palacios G, Guillen-Navarro K. Antifungal activity of Serratia marcesens CFFSUR-B2 purified chitinolytic enzymes and prodigiosin against Mycosphaerella fijiensis, causal agent of black Sigatoka in banana (Musa spp.). Biocontrol. 2015; 60(4):565–72. doi: 10.1007/s10526-015-9655-6 [Google Scholar]
- 96.Petersen LM, Tisa LS. Friend or foe? A review of the mechanisms that drive Serratia towards diverse lifestyles. Can J Microbiol. 2013; 59(9):627–40. doi: 10.1139/cjm-2013-0343 [DOI] [PubMed] [Google Scholar]
- 97.Someya N, Kataoka N, Komagata T, Hirayae K, Hibi T, Akutsu K. Biological control of cyclamen soilborne diseases by Serratia marcescens strain B2. Plant Dis. 2000; 84(3):334–40. [DOI] [PubMed] [Google Scholar]
- 98.Queiroz BPV, Melo IS. Antagonism of Serratia marcescens towards Phytophthora parasitica and its effects in promoting the growth of citrus. Braz J Microbiol. 2006; 37(4): 448–50. [Google Scholar]
- 99.Tan D, Fu L, Han B, Sun X, Zheng P. Identification of an endophytic antifungal bacterial strain isolated from the rubber tree and its application in the biological control of banana Fusarium wilt. PLoS One 2015; 10(7): e0131974 doi: 10.1371/journal.pone.0131974 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Chakraborty U, Chakraborty BN, Chakraborty AP. Influence of Serratia marcescens TRS-1 on growth promotion and induction of resistance in Camellia sinensis against Fomes lamaoensis. J Plant Interact. 2010; 5(4):261–72. [Google Scholar]
- 101.George P, Gupta A, Gopal M, Thomas L, Thomas GV. Multifarious beneficial traits and plant growth promoting potential of Serratia marcescens KiSII and Enterobacter sp. RNF 267 isolated from the rhizosphere of coconut palms (Cocos nucifera L.). World J Microbiol Biotechnol. 2013; 29(1):109–17. doi: 10.1007/s11274-012-1163-6 [DOI] [PubMed] [Google Scholar]
- 102.Zaheer A, Mirza BS, Mclean JE, Yasmin S, Shah TM, Malik KA, et al. Association of plant growth-promoting Serratia spp. with the root nodules of chickpea. Res Microbiol. 2016;167(6):510–20. doi: 10.1016/j.resmic.2016.04.001 . [DOI] [PubMed] [Google Scholar]
- 103.Gyaneshwar P, James EK, Mathan N, Reddy PM, Reinhold-Hurek B, Ladha JK. Endophytic colonization of rice by a diazotrophic strain of Serratia marcescens. J Bacteriol. 2001; 183(8):2634–45. doi: 10.1128/JB.183.8.2634-2645.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Someya N, Nakajima M, Watanabe K, Hibi T, Akutsu K. Potential of Serratia marcescens strain B2 for biological control of rice sheath blight. Biocontrol Sci Technol. 2005; 15(1):105–9. [Google Scholar]
- 105.Berg G, Zachow C, Lottmann J, Gotz M, Costa R, Kornelia S. Impact of plant species and site on rhizosphere-associated fungi antagonistic to Verticillium dahliae Kleb. Appl Environ Microbiol. 2005; 71(8):4203–13. doi: 10.1128/AEM.71.8.4203-4213.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Zachow C, Pirker H, Westendorf C, Tilcher R, Berg G. The Caenorhabditis elegans assay: a tool to evaluate the pathogenic potential of bacterial biocontrol agents. Eur J Plant Pathol. 2009; 125(3):367–76. [Google Scholar]
- 107.König W, Faltin Y, Scheffer J, Schöffler H, Braun V. Role of cell-bound hemolysin as a pathogenicity factor for Serratia infections. Infect Immun 1987; 55(11):2554–61. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Kurz CL, Chauvet S, Andrès E, Aurouze M, Vallet I, Michel GP, et al. Virulence factors of the human opportunistic pathogen Serratia marcescens identified by in vivo screening. EMBO J. 2003; 22(7):1451–1724. doi: 10.1093/emboj/cdg159 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files. All GenBank Sequence files are available from the GenBank database (accession number JX566992).