

Abstract
Parkinson’s disease (PD) is a progressive neurodegenerative disorder clinically characterized by cardinal motor deficits including bradykinesia, tremor, rigidity and postural instability. Over the past decades, it has become clear that PD symptoms extend far beyond motor signs to include cognitive, autonomic and psychiatric impairments, most likely resulting from cortical and subcortical lesions of non-dopaminergic systems. In addition to nigrostriatal dopaminergic degeneration, pathological examination of PD brains, indeed, reveals widespread distribution of intracytoplasmic inclusions (Lewy bodies) and death of non-dopaminergic neurons in the brainstem and thalamus. For that past three decades, the MPTP-treated monkey has been recognized as the gold standard PD model because it displays some of the key behavioral and pathophysiological changes seen in PD patients. However, a common criticism raised by some authors about this model, and other neurotoxin-based models of PD, is the lack of neuronal loss beyond the nigrostriatal dopaminergic system. In this review, we argue that this assumption is largely incorrect and solely based on data from monkeys intoxicated with acute administration of MPTP. Work achieved in our laboratory and others strongly suggest that long term chronic administration of MPTP leads to brain pathology beyond the dopaminergic system that displays close similarities to that seen in PD patients. This review critically examines these data and suggests that the chronically MPTP-treated nonhuman primate model may be suitable to study the pathophysiology and therapeutics of some non-motor features of PD.
Keywords: Parkinson’s disease, MPTP monkey, Nigrostriatal dopamine, Extra-striatal dopamine, Norepinephrine, Serotonin, Acetylcholine, α-synuclein
Introduction
Parkinson’s disease (PD) is a progressive movement disorder clinically characterized by bradykinesia, tremor, rigidity and postural instability. The pathological hallmarks of PD are the loss of nigrostriatal dopaminergic (DA) neurons and the widespread distribution of cytoplasmic inclusions composed of pre-synaptic α-synuclein protein, termed Lewy bodies. The motor signs appear when ~50–60% of DA cells in the substantia nigra par compacta (SNC) and ~60–80% of striatal DA terminals have been lost (Bernheimer et al. 1973; Hornykiewicz 1975; Hornykiewicz 1998; Hornykiewicz and Kish 1987). Apart from lesion of the nigrostriatal dopamine system, the PD pathology involves profound loss of brainstem noradrenergic, serotonergic and cholinergic neurons (Braak et al. 2003b; Chan-Palay 1991; Del Tredici et al. 2002; German et al. 1992b; Patt and Gerhard 1993; Zarow et al. 2003b). Loss of these non-dopaminergic neuronal groups likely contributes to some of the cognitive, autonomic and psychiatric impairments that are commonly seen in PD patients (Aarsland et al. 2004; Aarsland et al. 1999; Grimbergen et al. 2004; Langston 2006; Zesiewicz et al. 2003). However, because of the lack of animal models to study the pathophysiology of these non-motor signs, advances in the development of therapeutics have been limited and remain a major challenge in PD research for years to come.
Since the discovery that nigral dopamine loss is the key pathological hallmark of PD, various neurotoxin- and genetic-based models of PD that display profound nigrostriatal dopaminergic degeneration have been developed in rodents. Previous reviews have discussed in detail the strengths and weaknesses of these models and their relevance towards our understanding of the pathophysiology of motor and non-motor signs of PD (Bove and Perier 2012; Cenci et al. 2015; Duty and Jenner 2011; Jackson-Lewis et al. 2012; Jagmag et al. 2015; Lindgren and Dunnett 2012; Morin et al. 2014; Solari et al. 2013; Tolwani et al. 1999; Yue 2012). Although these models have made significant contribution to the field of PD research and therapeutics, their acute nature and the limited pathology to the nigrostriatal dopaminergic system limits their use for studying non-motor signs in PD (Dunnett and Lelos 2010; Fifel et al. 2016; Jenner 2002; Lindgren and Dunnett 2012; Taylor et al. 2010). The recent attempt at developing mice models that display chronic exposure to neurotoxin or genetically induced reduced expression of vesicular monoamine transporters have shown promising results in regards to pathology of non-dopaminergic cell groups and the development of some non-motor deficits commonly seen in PD (Betarbet et al. 2000; Fornai et al. 2005; Hoglinger et al. 2003b; Johnson and Bobrovskaya 2015; McNaught et al. 2004; Taylor et al. 2009).
The discovery of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) as a contaminant of street drugs in the mid-1980s had a major impact in the field of PD because of its use as potent dopaminergic neurotoxin and its impact on the epidemiology of environmental toxins in the development of PD (Langston et al. 1983). Since then, the MPTP-treated nonhuman primate model is recognized as the “gold standard” model of PD (Bezard and Przedborski 2011; Dauer and Przedborski 2003; Emborg 2007; Fox and Brotchie 2010; Johnston and Fox 2015; Morissette and Di Paolo 2017; Porras et al. 2012). However, like most neurotoxin-based models of PD, a common criticism put forward by some authors in recent years was the lack of pathological neuronal loss beyond the nigrostriatal dopaminergic system in this model (Blandini and Armentero 2012; Dauer and Przedborski 2003; Forno et al. 1993). Although this may be the case for monkeys that undergo acute exposure to the toxin, evidence from the literature suggests that chronic systemic administration of MPTP leads to pathological neuronal loss that extends far beyond the ventral midbrain dopaminergic cell groups (Forno et al. 1993; Fox and Brotchie 2010; Karachi et al. 2010; Masilamoni et al. 2011b; Masilamoni et al. 2016; Villalba et al. 2014). Furthermore, various groups have shown that MPTP-treated monkeys display early cognitive impairments and other non-motor features of PD that likely result from non-dopaminergic lesions (Pessiglione et al. 2004; Roeltgen and Schneider 1991; Schneider and Kovelowski 1990; Slovin et al. 1999; Taylor et al. 1990; Vezoli et al. 2011).
In this review, we will critically examine the current state of knowledge of the brain pathology and related motor and non-motor deficits reported from nonhuman primate studies using various regimens of MPTP as neurotoxin-based models of PD. We will argue that monkeys treated chronically with low doses of MPTP should be considered as reliable animal models to study the pathophysiology and therapeutics of some non-motor features of PD.
MPTP: Discovery and Relevance for PD
MPTP was first identified as a PD-related drug in the 1980s after Californian heroin addicts displayed severe parkinson-like motor syndrome following self-administration of a potent pethidine derivative contaminated with MPTP (Langston et al. 1983; Langston and Ballard 1983). These victims displayed symptoms of bradykinesia and rigidity almost identical to those seen in idiopathic PD patients (Langston et al. 1983; Langston and Ballard 1983). Furthermore, their positive response to Levodopa therapy and the subsequent development of motor and non-motor side effects commonly associated with long term exposure to such treatment provided compelling evidence that these individuals had been rendered parkinsonian through MPTP exposure. Postmortem pathological studies of brains from some of these individuals confirmed the profound dopaminergic neuronal loss in the ventral midbrain and striatal dopamine denervation (Langston et al. 1999).
Since its discovery, the mechanisms of action of MPTP towards dopaminergic cell loss have been characterized and discussed in detail in previous reviews (Dauer and Przedborski 2003; Forno et al. 1995; Fox and Brotchie 2010; Jenner and Marsden 1986; Johannessen 1991; Langston et al. 1999; Meissner et al. 2003; Porras et al. 2012; Rappold and Tieu 2010). In brief, once it has crossed the blood brain barrier and got to the brain, MPTP is rapidly converted into the toxic MPP+ metabolite by glial monoamine oxidase-B (MAO-B). Because of its high affinity for MPP+, dopamine transporters (DAT), heavily expressed on nigrostriatal dopamine terminals, become the main gateway for MPP+ to preferentially access and kill nigral dopamine neurons (Dauer and Przedborski 2003; Jenner and Marsden 1986). Albeit a lower affinity for norepinephrine and serotonin transporters (NET, SERT), MPP+ is also picked up by these transporters, providing a substrate for MPTP-induced noradrenaline and serotonin cell loss in some PD models (Boulet et al. 2008; D’Amato et al. 1986; Fornai et al. 2005; Herkenham et al. 1991; Masilamoni et al. 2011b; Perez-Otano et al. 1991; Pifl et al. 1996; Unguez and Schneider 1988). Because of differential ratios of intracerebral versus systemic MAO-B expression between rodents and primates, the MPTP dosages used to induce cell death in the monkey brain are many orders of magnitude lower than those used in mice (Duty and Jenner 2011; Giovanni et al. 1994b; Jenner 2002; Johannessen et al. 1985). In rats, MPTP is largely ineffective at permanently killing brain cells (Giovanni et al. 1994a; Jenner 2002).
Non-human primates (NHP) treated with MPTP develop motor abnormalities that closely resemble those seen in PD patients, including bradykinesia, rigidity and postural abnormalities. These animals display a strong positive response to L-DOPA (Burns et al. 1983) which, after chronic exposure, lead to human-like treatment-related motor complications; in particular, the L-DOPA-associated dyskinesias (LID) (Boyce et al. 1990; Pearce et al. 1995; Potts et al. 2014). Recent evidence also suggests that MPTP-treated monkeys display early cognitive impairments and other non-motor deficits reminiscent of those described in PD patients. In the following account, the brain pathology and behavioral changes reported in MPTP-treated monkeys will be critically examined and their relevance towards PD will be discussed.
Do MPTP-treated Monkeys Display a Pattern of Nigrostriatal Dopaminergic Denervation Similar to PD?
The major sources of DA in the central nervous system are in the ventral midbrain comprising areas A8 (retrorubral field; RRF), A9 (SNc), and A10 (ventral tegmental area; VTA). The SNc neurons are further segregated into a dorsal (SNCd) and ventral (SNCv) tiers based on the expression or not of the calcium binding protein, calbindin D28k (CB) (Masilamoni et al. 2010; Masilamoni et al. 2011b) (Fig.1A, B). Although all dopaminergic cell groups undergo degeneration in PD, the SNCv is recognized as the most sensitive region, while the VTA is the least affected ventral midbrain area (Damier et al. 1999a; Damier et al. 1999c; German et al. 1992a; Lu et al. 2006), (see Table 1). The presence of CB, low level expression of DAT, VMAT2 and more profuse norepinephrine innervation may contribute to the differential sensitivity of VTA/SNCd vs SNCv neurons to PD pathology (Dopeso-Reyes et al. 2014; Iacopino et al. 1992a; Kilbourn et al. 1998; Liang et al. 1996b; Masilamoni et al. 2010; Masilamoni et al. 2016; Mejias-Aponte et al. 2009; Miller et al. 1999a; Rommelfanger et al. 2004; Sanghera et al. 1997; Yuan et al. 2013). However, the importance of these features on the overall pattern of MPTP-induced neuronal degeneration can be assessed only if monkeys are treated with slowly progressive MPTP regimen (Airaksinen et al. 1997; Porras et al. 2012). Using CB as specific marker of SNCd and VTA neurons, monkeys administered chronically with weekly low doses of MPTP (0.2–0.5mg/kg MPTP, once a week for 21 weeks) display the same pattern of midbrain DA degeneration as shown in PD patients (Damier et al. 1999b; Damier et al. 1999d; Fearnley and Lees 1991); i.e. a profound loss of CB-negative SNCv neurons over CB-positive SNCd, VTA and RRF neurons (Lavoie and Parent 1991; Masilamoni et al. 2010; Masilamoni et al. 2011b) (Table 1). In contrast, when monkeys are made parkinsonian through a more acute and aggressive regimen of MPTP, the extent of DA degeneration is the same in both SNCv and SNCd (Collier et al. 2007). Thus, although CB may be neuroprotective against the toxic effects of low doses of MPTP in chronically treated animals (Dopeso-Reyes et al. 2014; Iacopino et al. 1992b; Liang et al. 1996a; Yuan et al. 2013), these properties are overcome by high doses of MPTP. Other features that have been put forward to explain the relative sparing of SNCd/VTA neurons versus SNCv cells in response to MPTP (or other neurotoxins) include the relatively low levels of DAT expression on VTA/SNCd striatal dopamine terminals compared with SNCv neurons (less uptake sites for the dopaminergic neurotoxin MPP+) and/or the neuroprotective effects of a much more profuse norepinephrine innervation of VTA/SNCd neurons than SNCv neurons (Kilbourn et al. 1998; Masilamoni et al. 2016; Mejias-Aponte et al. 2009; Rommelfanger et al. 2004; Sanghera et al. 1997). Thus, the biochemical differences between the various groups of midbrain DA neurons and their respective extent in NE innervation may contribute to the differential sensitivity of SNCd/VTA vs SNCv neurons to PD pathology and MPTP toxicity.
Figure 1.

Photomicrographs of adjacent calbindin (CB) and tyrosine hydroxylase (TH) -immunostained coronal sections at the level of ventral midbrain of control (A,C) and MPTP-treated parkinsonian monkey (B,D). CB immunostaining was used to delineate TH/CB-positive neurons in the dorsal tier of the SNC (SNCd) and the ventral tegmental area (VTA) from TH-positive/CB-negative neurons in ventral tier of the SNC (SNCv). (E) Percentage loss of TH-positive neurons in SNCv, SNCd and VTA regions of three MPTP-treated monkeys (M1, M2 and M3) based on unbiased stereological estimates of total TH-positive cell counts. (see Masilamoni et al., 2011b for additional details)
Table 1.
| Species | Age | Sex | PD patients/MPTP dose | MPTP regimen | Parkinsonian clinical symptom | Pathology observation (Nigral dopamine cell/striatal DA levels loss in %) | Reference | |
|---|---|---|---|---|---|---|---|---|
| Midbrain dopaminergic neurons | ||||||||
| Human | 62–83 | M,F | PD | – | Fully Parkinsonian | VM – 71 VL - 91 PL – 61 DM – 47 DL - 57 |
(Fearnley and Lees 1991) | |
| Human | 72 ± 4 | – | PD | – | Fully Parkinsonian | SNC v nigrosomes – 95 SNC v matrix - 80 SNC d - 57 |
(Damier et al. 1999b; Damier et al. 1999d) |
|
| Callithrix jacchus | – | F | 1–4 mg/kg/day for 5 days,i.p | Acute high dose treatment | Fully Parkinsonian | SNC 82 VTA 59 |
(Waters et al. 1987) | |
| Papio papio | 0.2–0.5 mg/kg i.v once a week until a full parkinsonian | Chronic low dose treatment | Fully Parkinsonian | VM - 73.8 VL - 90.9 PL - 89.5 DM – 46 DL - 56.6 VTA - 53 |
(Varastet et al. 1994) | |||
| Cercopithec us aethiops | 4 – 6 | M | 0.4 – 0.6 mg/kg i.m Progressive intoxication: (injections spaced 4 to 5 days) | Chronic low/high dose treatment | Fully Parkinsonian monkeys | SNC - 71 RRF - 35 VTA - 34 |
(Mounayar et al. 2007) | |
| Macaca mulatta | 8–9 Young 15–17 Middle 21–31 Old |
F | 2.3–3.00 mg, unilateral intracarotid administration | Single dose | Hemiparkinsonia monkey | SNC - 81.8 SNC - 82.8 SNC-81.0 |
(Collier et al. 2007) | |
| Macaca mulatta | 3–6 | F | 0.2–0.5 mg/kg i.m once a week until a full parkinsonian | Chronic low dose treatment | Moderate Parkinsonian monkeys Fully Parkinsonian monkeys |
SNC v - 64 SNC d - 53 VTA – 42 SNC v - 85 SNC d - 73 VTA – 60 |
(Masilamoni et al. 2011b) | |
| Macaca fascicularis | 4–5 | M | 0.5 mg/kg i.v once in 2 weeks | Chronic low dose treatment | Moderate Parkinsonian monkeys Fully Parkinsonian monkeys |
SNC - 74 RRF - 40 VTA – 34 SNC - 84 RRF - 45 VTA - 33 |
(Blesa et al. 2012) | |
| Nigrostriatal Dopamine Innervation | ||||||||
| Human | - | - | Fully Parkinsonian | Pre-commissural CA-90 Commissural CA - 82 Post-commissural CA- 70 Pre-commissural PU-93 Commissural PU – 97 Post-commissural PU-98 |
(Kish et al. 1988) | |||
| Human | – | – | Fully Parkinsonian | CA-84 PU-98 |
(Pifl et al. 1988b) | |||
| Human | – | – | Fully Parkinsonian | CA-80 PU-98 |
(Kish et al. 2008) | |||
| Macaca mulatta | 10–17 | 0.15 mg/kg for the first 3 days and consecutively 0.24 – 0.4 mg/kg per day until severe Parkinson-like symptoms | Acute low dose treatment | Fully Parkinsonian monkeys | CA-99 PU-99 |
(Pifl et al. 1988b) | ||
| Cercopithec us- aethiops subueus | 0.4mg/kg for 5 days | Acute low dose treatment | Fully Parkinsonian monkeys | CA 98 PU 96 |
(Elsworth et al. 1989) | |||
| Callithrix jaccus | 1–2Y | M,F | 1.25–2.5 mg/kg s.c. was given twice a week up to development of symptom | Chronic high dose treatment | Fully Parkinsonian monkeys | CA 95 PU 95 |
(Perez-Otano et al. 1991) | |
| Macaca fascicularis | daily injections (0.25–0.50 mg/kg) | Acute low dose treatment | Fully Parkinsonian monkeys | PU 97 CA 98 |
(Alexander et al. 1992) | |||
| Macaca fascicularis | – | 3 consecutive i.v injection at 0.7mg/day | Acute high dose treatment | Fully Parkinsonian monkeys | CA 94 PU 93 | (Hantraye et al. 1992) | ||
| Pupio pupio | 0.4–0.6 mg/kg i.v once a week until a full parkinsonian | Chronic low dose treatment | Fully Parkinsonian monkeys | CA 70 PU 90 |
(Hantraye et al. 1993) | |||
| Saimiri sciureus | – | M,F | single subcutaneous injection of 2 mg/kg MPTP received multiple subcutaneous injections |
Single dose Acute intoxication |
no behavioral signs of parkinsonism Fully Parkinsonian monkeys |
CA 63 PU 70 CA 95 PU 99 |
(Di Monte et al. 2000) | |
| Cercopithec us- aethiops sabaeus | – | 0.4mg/kg for 5 days | Acute low dose treatment | Asymptomatic Mild Moderate Severe |
CA 70; PU 58 CA 90; PU 68 CA 94; PU 92 CA99; PU 99 |
(Elsworth et al. 2000) | ||
| Caca fascicularis | 3.1 ± 0.3 | F | 0.2 mg/kg, i.v. daily after 6 injections; D6 after 15 injections; D15 after 15 injections; D25 | Acute low dose treatment | Pre-symptomatic Appearance of symp. Fully Parkinsonian monkeys |
Loss of dopamine CA 50; PU 40 PU 85%; CA 90 CA 95; PU 95% |
(Bezard et al. 2001) | |
| Mucucu fusciculuris | – | M | Intracarotid injections Single dose of 1.25 mg Single dose of 2.5 mg |
Acute low dose treatment | hemiparkinsonism | ipsilateral DA loss CA 99.4; PU, 99.8; AC 74.2 CA 99.3; PU 99.5; AC 90.1 |
(Guttman et al. 1990) | |
| Macaca fascicularis | 2–4 | 5–15 injections of 0.5–1.0 mg/kg spaced one week apart | Chronic high dose treatment | Fully Parkinsonian | Pre-commissural CA-98 Commissural CA - 94 Post-commissural CA- 90 Pre-commissural PU-98 Commissural PU – 99 Post-commissural PU-99 |
(Perez-Otano et al. 1994a) | ||
| Macaca mulatta | 3–6 | F | 0.2–0.5 mg/kg i.m once a week until full parkinsonian | Chronic low dose treatment | Moderate Parkinsonian monkeys Fully Parkinsonian monkeys |
CA - 63 PU - 80 CA - 80 PU - 90 |
(Masilamoni et al. 2011b) | |
| Macaca fascicularis | 4–5 | M | 0.5 mg/kg i.v once in 2 weeks | Chronic low dose treatment | Moderate Parkinsonian monkeys Fully Parkinsonian monkeys |
CA - 78 PU - 79 CA - 84 PU - 83 |
(Blesa et al. 2012) | |
VM – Ventral medial, VL - Ventral lateral, PL – Pars lateralis, DM – Dorsal medial part, DL - lateral part, CA- Caudate, PU- Putamen, SNC - Substantial nigra, SNC v- ventral, SNC-d-dorsal, VTA – Ventral tegmental area, RRF – Retrorubral area
This pattern of DA neuronal degeneration holds true for susceptibility to MPTP-induced degeneration. This differential degeneration at the cell body level is represented in the striatum by a more severe loss of dopaminergic innervation in the dorsal (targeted mainly by SNCv neurons) than the ventral striatum (targeted by VTA neurons). In the dorsal striatum, the dopamine loss is not homogeneous, but rather displays a progressive caudo-rostral gradient, ie it affects preferentially the post-commissural putamen (known as the sensorimotor striatal territory) over the anterior putamen and the caudate nucleus (known as associative striatal regions) (Davis et al. 2003; Kish et al. 1988). Although there is no clear explanation as to why SNc neurons that project to the sensorimotor putamen are more sensitive to PD degeneration than those that innervate the cognitive striatal regions, some hypotheses have been put forward ranging from a different ratio of DAT/VMAT2 on dopamine terminals in the caudate nucleus vs putamen (Miller et al. 1999a; Miller et al. 1999b) to a differential synaptic innervation of dopamine neurons that project to the caudal striatum compared with other dopamine-projecting neurons in the mouse SNc (Menegas et al. 2015).
There has been various reports indicating that the preferential dopamine denervation of the post-commissural putamen over other dorsal striatal regions was not found in monkeys treated with acute or a high dose regimen of MPTP (Alexander et al. 1992; Bezard et al. 2001; Di Monte et al. 2000; Elsworth et al. 1989; Fernagut et al. 2010; Hantraye et al. 1992; McCallum et al. 2006; Pifl et al. 1988a). However, data from our laboratory and others using chronic low-dose MPTP exposure revealed that the progressive loss of striatal dopamine terminals reported in PD patients, i.e. a greater DA depletion in the post-commissural putamen than ventral striatum and caudate (Fig. 2 A–F), can also be seen in parkinsonian monkeys (Hantraye et al. 1993; Iravani et al. 2005; Masilamoni et al. 2010; Masilamoni et al. 2011b; Moratalla et al. 1992; Mounayar et al. 2007). Thus, dopamine terminals in the post-commissural putamen are more sensitive to degeneration than any other nigrostriatal afferents in PD and in monkeys chronically intoxicated with low doses of MPTP.
Figure 2.
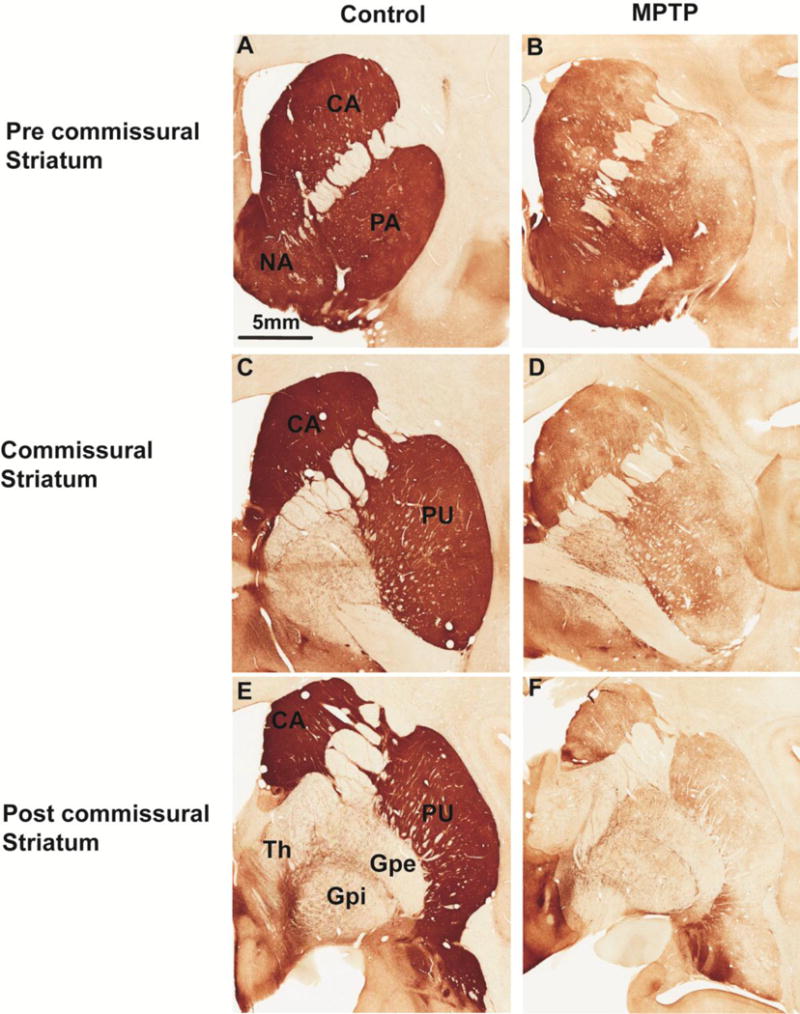
Photomicrographs of TH-immunostained coronal sections at the level of the pre-commissural (A, B), commissural (C, D), post-commissural (E,F) striatum of a control (left column) and a MPTP-treated (right column) monkey. Abbreviations: CA: caudate nucleus; PU: putamen; GPe: globus pallidus, external segment; GPi: globus pallidus, internal segment; Th: thalamus.
Early Dopaminergic Denervation of Striosomes over Matrix in Chronically MPTP-treated Monkeys
During brain development and early postnatal period, striatal DA innervation is compartmentally organized, giving rise to the so-called “dopamine islands” which, in adults, corresponds to the striosomal striatal compartment (Graybiel et al. 1981; Prensa et al. 2000). Striosomes and matrix can also be differentiated from each other neurochemically or anatomically based on the heterogeneous distribution of various neuronal markers and afferents from specific cortical and sub-cortical regions (Gerfen 1984; Gimenez-Amaya and Graybiel 1991; Graybiel and Ragsdale 1978; Herkenham and Pert 1981). Because of its abundance and homogeneous distribution in both compartments, markers of nigrostriatal dopaminergic axons and terminals (TH or DAT immunostaining; Fig. 3A, B) do not display any heterogeneity in the normal adult striatum (Graybiel et al. 1981; Prensa et al. 2000). However, in monkeys chronically treated with low doses of MPTP, dopamine denervation of striosomes in the caudate nucleus and anterior putamen precedes that of the matrix (Iravani et al. 2005; Lin et al. 2015; Masilamoni et al. 2011b) (see Fig 3 C–F). In this model, early striosomal dopamine loss occurs in parallel with the denervation of the post-commissural putamen (Iravani et al. 2005; Lin et al. 2015; Vingerhoets et al. 1994), suggesting that SNCv nigrostriatal neurons that project to the caudal putamen and the striosomes are most sensitive to MPTP-induced neurodegeneration. Based on recent findings that striatonigral GABAergic projections from striosomes form tight connections with clusters of SNCv dopaminergic cells (Crittenden et al., 2016), one could speculate that the striosomal striato-nigro-striatal loop is a functional network that displays increased sensitivity to degeneration in PD. Tract-tracing studies showing that anterograde tracer injections into the SNCv produce labeling that predominantly target the striosomes, whereas dorsal injections label preferentially the striatal matrix are consistent with this hypothesis (Fujiyama et al. 2011; Gerfen et al. 1987; Jimenez-Castellanos and Graybiel 1987; Joel and Weiner 2000; Langer and Graybiel 1989). Because striosomes are in a position to modulate and transmit signals to the SNc from limbic-related cortical regions (Crittenden et al. 2016; Eblen and Graybiel 1995; Friedman et al. 2015), early striosomal dopamine loss may affect the control of mood, attention and motivation, non-motor signs that are commonly seen in PD patients. It is noteworthy that imbalanced activity between striosomes and matrix has been proposed as the underlying substrate of a wide variety of motor and non-motor signs in a wide array of basal ganglia disorders (Crittenden and Graybiel, 2011).
Figure 3.
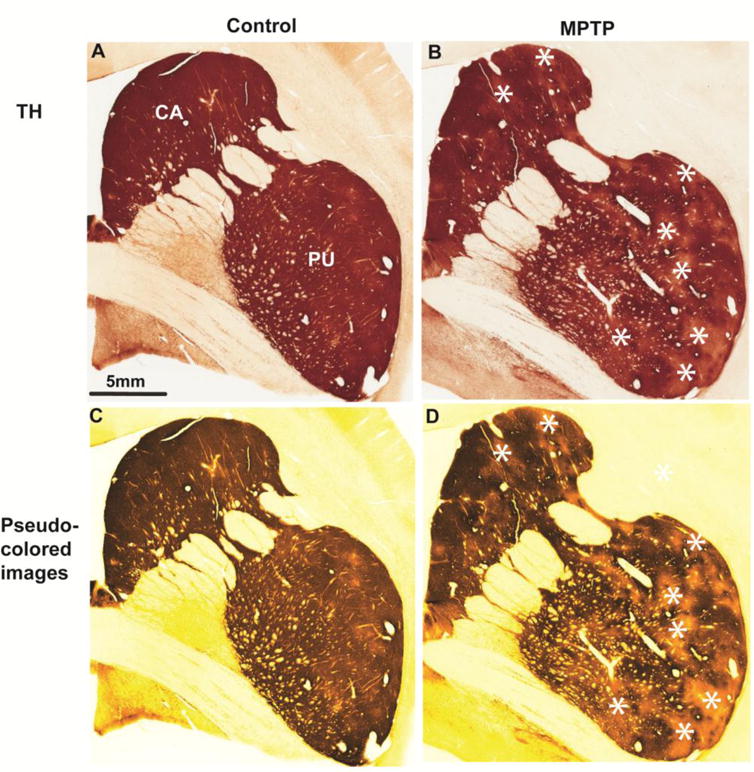
Photomicrographs of TH-immunostained sections of the striatum in a control (A) and MPTP-treated asymptomatic monkey (B). (C,D) Pseudo-colored images (NIH ImageJ program) of TH-immunostained sections showed in A and B. MPTP treatment preferentially reduced TH immunoreactivity in presumed striatal patches (white asterisks). Abbreviations: CA: caudate nucleus; PU: putamen.
Extra-striatal DA Denervation in PD vs MPTP-treated Monkeys
Although the dorsal striatum is the most severely dopamine-denervated brain region in PD and MPTP-treated monkeys, non-striatal brain nuclei also undergo dopamine depletion (Benazzouz et al. 2014; Lee et al. 2014; Parent and Cossette 2001; Rommelfanger and Wichmann 2010; Smith and Villalba 2008). In contrast to the nigrostriatal dopamine degeneration, which contributes to the pathophysiology of the key parkinsonian motor signs, very little is known about the functional significance of extrastriatal dopamine loss in PD (Smith and Kieval, 2000; Rommelfanger and Wichmann, 2010). However, because the cortical and subcortical regions affected are involved in broad range motor and non-motor functions, a comparative analysis of the overall pattern of extrastriatal DA degeneration between PD patients and MPTP-treated monkeys is critical for the assessment of the reliability of this animal model towards studies of extrastriatal DA pathology in PD non-motor signs.
Dopamine Denervation of the Globus pallidus
Immunohistochemical and tract-tracing studies reported that the two pallidal segments (GPe - external and GPi- internal) receive DA innervation from the ventral midbrain dopamine cell groups in both humans and NHPs (Cossette et al. 1999; Hedreen 1999; Nobin and Bjorklund 1973). In normal humans, DA levels in the GPe are nearly six times greater than in the GPi. In PD, the mean loss of DA is severe (82%) in GPe and moderate (51%) in GPi (Rajput et al. 2008). Electrophysiological and pharmacological studies supporting these observations showed that dopamine receptor-related drugs induce significant changes in GPe and GPi neurons through pre- and post-synaptic mechanisms in control and parkinsonian monkeys (Boraud et al. 1998; Filion et al. 1991; Hadipour-Niktarash et al. 2012; Kliem et al. 2010; Rommelfanger and Wichmann 2010; Smith and Kieval 2000). Several studies have shown that DA inputs are differentially distributed in GPe and GPi, with DA fibers arborizing more profusely in the medial “limbic-related” pole of GPi than in other functional sectors of GPe and GPi (Hedreen 1999; Lavoie et al. 1989; Parent and Smith 1987; Parent et al. 1989). In monkeys, the nigropallidal projection originates in part from a subset of dopaminergic neurons different from those that give rise to the nigrostriatal system (Smith and Kieval 2000). The importance of the nigropallidal dopaminergic system in the development of parkinsonian motor symptoms is further supported by pharmacological and electrophysiological data gathered in normal and MPTP-treated monkeys. Thus, infusion of D1 or D2 dopamine receptor antagonists into the GPe of normal rats induces akinesia (Hauber and Lutz 1999). Local intrapallidal delivery of D1 or D2 receptor-related drugs into GPe or GPi of normal and parkinsonian monkeys affect the firing rate and pattern of pallidal neurons (Hadipour-Niktarash et al. 2012; Kliem et al. 2007).
Data from MPTP-treated monkeys support the human studies suggesting that the PD-related pathology of the nigropallidal dopaminergic system is less severe than the nigrostriatal projection, particularly during the asymptomatic or early stages of the disease (Lin et al. 2015; Mounayar et al. 2007; Pifl et al. 1990). In fact, an increase in 18F-dopa uptake was demonstrated in the GPi of early PD patients compared to aged match controls, suggesting a compensatory response of the nigro-GPi system to nigrostriatal dopamine denervation (Moore et al. 2008; Pavese et al. 2011; Rakshi et al. 1999; Whone et al. 2003). Thus, data from both human and MPTP-treated monkeys suggest a relative sparing or up-regulation of the nigro-pallidal dopamine projection in the early stages of PD representing adaptive mechanism to preserve functionality.
Dopamine Denervation of the Subthalamic nucleus
Anatomical and tracing studies have demonstrated the existence of SNC-STN projections in both monkeys and humans (Augood et al. 2000; Cossette et al. 1999; Francois et al. 2000; Gauthier et al. 1999; Hassani et al. 1997; Hedreen 1999; Lavoie et al. 1989). However, very few vesicle-filled dopamine axonal profiles and terminals were reported in electron microscopic analysis of the monkey STN, raising questions about the source and mechanism of release of dopamine by nigrosubthalamic axons (Smith and Kieval 2000). The situation is different in the rodent STN, which is far more enriched in dopamine terminals than the primate STN (Cragg et al. 2004). Various DA receptor subtypes are expressed pre- and post-synaptically in the primate and rodent STN (Augood et al. 2000; Flores et al. 1999; Galvan et al. 2014; Parry et al. 1994; Rommelfanger and Wichmann 2010; Smith and Villalba 2008). Intra-subthalamic applications of D1 or D2 receptor agonists alter the firing rate and pattern of STN neurons in control and MPTP-treated monkeys (Galvan et al. 2014).
There is a substantial loss of dopamine innervation of the STN in MPTP-treated monkeys (Francois et al. 2000; Pifl et al. 1990; Pifl et al. 1991) and in human PD patients (Hornykiewicz 1998; Pavese et al. 2011). In contrast to the nigropallidal projection that is relatively spared in motor asymptomatic MPTP-treated monkeys, the nigrosubthalamic system is severely degenerated in these animals (Pifl et al. 1991), thereby suggesting that the nigropallidal, but not the nigrosubthalamic, system might functionally compensate striatal dopamine denervation in early parkinsonian state. Thus, one would speculate that the DA loss in the STN may concomitantly contribute to the abnormal increased activity of these glutamatergic neurons and eventually lead to the expression of motor signs in PD (Bergman et al. 1994; Fox and Brotchie 2010; Galvan et al. 2014; Rommelfanger and Wichmann 2010; Wichmann and DeLong 2003).
Dopamine Denervation of the Cerebral Cortex
Dopamine innervation of prefrontal cortices is well established and heavily studied because of its possible role in psychiatric disorders (Akil et al. 1999; Arnsten et al. 1995; Arnsten et al. 2015; Devoto et al. 2012; Dreher and Burnod 2002; Drouot et al. 2004; Erickson et al. 2000; Fallon et al. 2015; Goldman-Rakic 1998; Ko et al. 2013; Naneix et al. 2009; Narayanan et al. 2013; Sutoo et al. 2001; Thompson et al. 2014; Venator et al. 1999; Winterer and Weinberger 2004). Imaging and tract tracing studies have shown that the mesocortical dopamine system is far more extensive in primates than in rodents (Akil and Lewis 1994; Berger et al. 1991; Lewis et al. 1987; Lewis et al. 1994; Lewis et al. 2001; Preuss 1995; Sesack et al. 1995; Williams and Goldman-Rakic 1998a). Layer I is a widespread target of dense dopaminergic innervation in all areas of the macaque and human neocortex, while it is mainly confined to prefrontal cortical regions in rats (Berger et al. 1992; Berger et al. 1988; Gaspar et al. 1989; Petrides and Pandya 1999; Zilles et al. 1996) (Preuss 1995). Significant interspecies differences have also been demonstrated in the origin of the mesocortical system between rodents and primates, such that it arises almost exclusively from the VTA in rats (Berger et al. 1991; Deutch et al. 1988; Hosp et al. 2011; Luft and Schwarz 2009), while neurons in the SNc and the retrorubral area (RRA) significantly contribute to that system in monkeys (Gaspar et al. 1992; Williams and Goldman-Rakic 1998b).
Using 18F-dopa PET imaging, (Brooks and Piccini 2006) showed a significant loss of DA terminals in M1 and PFC of PD patients compared with age-matched control subjects. Postmortem data have revealed a significant degeneration of the dopaminergic inputs throughout M1 in humans, most strongly affecting dopaminergic fibers in layer I in PD patients (Gaspar et al. 1991; Gaspar et al. 1992). These observations are consistent with previous anatomical data (Jan et al. 2003), and with biochemical findings showing a 30–40% decrease of dopamine levels in M1 of MPTP-treated parkinsonian monkeys (Jan et al. 2003; Pifl et al. 1991; Scatton et al. 1983). Abnormal cortical metabolic activity was also noticed in both PD patients and MPTP-treated monkeys (Carbon et al. 2003; Dogali et al. 1994; Eckert et al. 2008; Eidelberg et al. 1994; Huang et al. 2007) with an earlier occurrence and greater magnitude in the M1 than in the PFC (Huang et al. 2007). Knowing that pre-motor and prefrontal cortical areas are under the influence of the dopamine system directly through the mesocortical network, one would expect that changes in these networks may contribute to these cortical metabolic dysfunctions. It has, indeed, been demonstrated that even very subtle cortical dopamine depletion may lead to a severe loss of action selection ability (Leblois et al. 2006), highlighting the important role of cortical dopamine in mediating these regulatory effects.
Thus, because of the evolutionary increase in cortical dopamine innervation between rodents and primates, and the comparable extent of degeneration of this system in motor and non-motor cortices in PD patients and MPTP-treated monkeys, nonhuman primates are the species of choice to study the role of cortical dopamine degeneration in the development of motor and non-motor parkinsonian symptoms.
Dopamine Denervation of the Thalamus
Anatomical studies have shown that various thalamic nuclei receive a significant dopaminergic innervation in humans and NHP (Brown et al. 1979; Garcia-Cabezas et al. 2009; Garcia-Cabezas et al. 2007; Goldman-Rakic and Brown 1981; Oke and Adams 1987; Pifl et al. 1990; Pifl et al. 1991; Sanchez-Gonzalez et al. 2005). The innervation of midline limbic nuclei, the mediodorsal and lateral posterior association nuclei, and the ventral lateral/ventral anterior motor nuclei (Brown et al. 1979; Garcia-Cabezas et al. 2009; Garcia-Cabezas et al. 2007; Goldman-Rakic and Brown 1981; Oke and Adams 1987; Pifl et al. 1990; Pifl et al. 1991; Sanchez-Gonzalez et al. 2005) is particularly dense. This distribution suggests that thalamic dopamine may play important roles in regulating emotion, attention, cognition, complex somatosensory and visual processing, and motor control. In contrast to primates, the dopamine innervation of the rodent thalamus is very scant (Papadopoulos and Parnavelas 1990; Voorn et al. 1988). The functional significance of this important species difference is unknown, but combined with the more widespread dopamine innervation of the cerebral cortex (see above), it suggests that dopamine modulatory functions of thalamocortical and corticothalamic relationships may be more complex in primates than non-primates (Berger et al. 1991; Papadopoulos and Parnavelas 1990).
Dopaminergic cell groups in the VTA, retrorubral field, periacqueductal gray and hypothalamus are the main sources of the primate thalamus innervation (Sanchez-Gonzalez et al. 2005), although another study suggested that collaterals of the nigrostriatal dopamine projection innervate the primate thalamus (Freeman et al. 2001).
In PD patients and MPTP-treated parkinsonian monkeys, DAT binding is significantly decreased throughout the thalamus (Brownell et al. 2003; Freeman et al. 2001; Pavese et al. 2010; Pifl et al. 1990; Pifl et al. 1991). Postmortem studies showed that the dopaminergic innervation of the MD and CnMd nuclei is most affected in PD patients and MPTP-treated parkinsonian monkeys (Freeman et al. 2001; Pavese et al. 2010; Pifl et al. 1990; Pifl et al. 1991) Although the role of dopamine in the primate thalamus remains to be established, it is reasonable to suggest that thalamic dopamine denervation may contribute to the pathophysiology of abnormal activity and transmission of motor and non-motor innervation through the basal ganglia-thalamocortical loops.
MPTP-induced Dopamine Denervation in Nonhuman Primates: Concluding Remarks
In PD, post-mortem studies have demonstrated that dopaminergic lesion affects cortical and sub-cortical regions beyond the nigrostriatal system. Similarly, chronic MPTP administration in adult monkeys results in a broad degeneration of dopaminergic projections to non-striatal regions. Extrastriatal basal ganglia nuclei such as the globus pallidus and subthalamic nucleus are both affected, but in opposite directions. Most importantly, widespread cortical dopamine pathology is also induced in this animal model. Because the mesocortical dopamine innervation of sensorimotor cortices is far more extensive in primates than rodents, the MPTP-treated monkey model is essential to assess the importance of cortical dopamine denervation on sensorimotor dysfunctions associated with parkinsonism. Another dopaminergic system that underwent a significant expansion in primates, and was found to be affected in MPTP-treated monkeys and PD patients is the meso-thalamic dopaminergic network. Although the function of this system remains unknown, its widespread termination in thalamic nuclei involved in the processing of motor, sensory, limbic and cognitive information suggests that degeneration of the meso-thalamic system may have broad consequences on the pathophysiology of the thalamocortical and corticothalamic relationships that underlie some of the motor and non-motor symptoms of PD.
Thus, the dopaminergic brain pathology induced in the chronic MPTP-treated monkey model is closely similar to that reported in PD patients, indicating that chronic MPTP-treated monkeys can be used to assess the importance of dopamine denervation beyond the nigrostriatal dopaminergic system that potentially contribute to a wide variety of non-motor symptoms in PD.
Chronic MPTP Administration in Non-human Primates Induces Severe Loss of Non-dopaminergic Neurons Relevant to Parkinson’s disease
In this section, we will review reports about the loss (or not) of non-DAergic cell groups in MPTP-treated monkeys, and discuss their relevance towards the pathophysiology and the development of non-motor signs in Parkinson’s disease.
Norepinephrine pathology-Locus Coeruleus and related brainstem noradrenergic cell groups
There is strong agreement across postmortem pathological studies that the loss of locus coeruleus (LC) noradrenergic neurons in PD patients is as prominent as that of the nigral dopaminergic neurons (Braak et al. 2003b; Chan-Palay 1991; Del Tredici and Braak 2013; German et al. 1992b; Patt and Gerhard 1993; Zarow et al. 2003a). However, reports about noradrenergic cell loss in LC of MPTP-treated monkeys have been inconsistent (Forno et al. 1986; Gibb et al. 1989; Herrero et al. 1993a), most likely because of differences in MPTP treatment regimens. Acute MPTP administration induces parkinsonian motor symptoms with massive nigrostriatal DA loss, but without significant LC neuronal death (Forno et al. 1993; Gibb et al. 1989). On the other hand, monkeys that undergo chronic systemic MPTP administration display prominent LC neurodegeneration (Herrero et al. 1993b; Masilamoni et al. 2011b)(Fig 4 A, B). In light of these findings, it appears that the regimen of MPTP administration may account for the differences in norepinephrine cell loss reported in these studies, such that chronic administration regimen is essential to induce significant LC neuronal death.
Figure 4.

Photomicrographs of TH-immunostained coronal sections at the level of noradrenergic cell groups (A, B), 5HT-immunostained serotonergic cell groups (C, D) and ChAT-immunostained pedunculopontine nucleus (E, F) of a control (A,C,E) and a MPTP-treated (B,E,F) monkey. Abbreviations: A5,A7: Noradrenergic cell groups A5 and A7; PPN: pedunculopontine nucleus. (see Masilamoni et al., 201b for more details).
Another noticeable feature of the noradrenergic system degeneration in MPTP-treated monkeys is that NE terminal loss in the midbrain and cerebral cortex is more pronounced than the magnitude of LC neuronal loss (Buddhala et al. 2015; Masilamoni et al. 2016; Nayyar et al. 2009), suggesting that NE nerve terminals are the primary target of the degenerative process after MPTP administration and in PD patients.
Thus, chronic MPTP injections recreate a pathological state of the noradrenergic system that mimics human PD more accurately than acute MPTP bolus injections. These observations suggest that severe and prolonged stress may trigger the onset of sporadic PD, thereby contributing to its progression (Herkenham et al. 1991). It is noteworthy that similar observations were reported for rotenone, i.e. only continuous administration of this neurotoxin induces LC neuronal loss (Hoglinger et al. 2003a; Hoglinger et al. 2003b). Even when administered at high doses, acute administration of rotenone damages only the nigrostriatal DA system (Ferrante et al. 1997; Heikkila et al. 1985).
Because LC plays a central role in cognitive performance (Coull et al. 1999; Sterpenich et al. 2006), sleep-waking cycle (Jouvet 1972) mood regulation and various autonomic functions, the degeneration of the LC and its projections likely contribute to a wide variety of non-motor symptoms in PD patients. Thus, the chronic MPTP-treated monkey model provides a unique tool needed to better understand the pathophysiology and refine therapy for these parkinsonian non-motor signs (Abbott et al. 2005; Barraud et al. 2009; Lees and Smith 1983; Schneider et al. 1999)
Serotonergic pathology
Serotonergic cell loss in raphe neurons has been well documented in advanced PD patients (D’Amato et al. 1987; Halliday et al. 1990; Ohama and Ikuta 1976). Consistent with these postmortem pathological observations, PET imaging studies have reported profound and widespread neocortical and striatal decrease of 5-HT transporter ligand binding in advanced PD patients (Albin et al. 2008; Guttman et al. 2007; Politis et al. 2010). However, in early PD state the serotonergic system is involved in compensatory phenomenon with an up-regulation or no change in the expression of 5-HT receptors in the striatum (Kaasinen et al. 2001; Kumakura et al. 2010; Moore et al. 2008; Rakshi et al. 1999; Whone et al. 2003). This compensatory mechanism fades away as the disease progresses so that advanced PD patients show reduced 5HT tracer uptake in cortical and striatal regions (Moore et al. 2008; Rakshi et al. 1999). Because dysfunction of the serotonergic system is involved in the pathophysiology of depression (Cummings 1992; Kostic et al. 1987; McCance-Katz et al. 1992), cognitive impairments (Brown and Marsden 1988; Pillon et al. 1989) and other psychiatric disorders (Cooney and Stacy 2016; Lenka et al. 2016; Reichmann et al. 2016), dysregulation of this system may contribute to some of the commonly encountered non-motor psychiatric and sleep disorders in PD (Abbott et al. 2005; Barraud et al. 2009; Lees and Smith 1983; Schneider et al. 1999). Mood changes in PD are, indeed, responsive to antidepressants including serotonin and norepinephrine (NE)-selective reuptake inhibitors (Blier 2006; Brooks and Doder 2001; Lemke 2008; Miyasaki et al. 2006; Zesiewicz and Hauser 2002).
Reports about serotonergic pathology in MPTP-treated monkeys have been variable (Ballanger et al. 2016; Beaudoin-Gobert et al. 2015; Boulet et al. 2008; Gaspar et al. 1993; Mihatsch et al. 1991; Mounayar et al. 2007; Perez-Otano et al. 1994a; Perez-Otano et al. 1994b; Pifl et al. 1991; Zeng et al. 2010). Increased serotonergic fibers (Boulet et al. 2008; Gaspar et al. 1993; Mounayar et al. 2007; Zeng et al. 2010) and serotonin levels (Boulet et al. 2008) have been reported in both MPTP-treated monkeys that recovered from transient motor symptoms and symptomatic animals. These reports suggested that this serotonin up-regulation may compensate for the loss of dopamine and delay the appearance of motor symptoms. There is evidence from animal models of PD that the relative sparing of striatal serotonin innervation over the nigrostriatal dopamine system may contribute to the development of L-DOPA-induced dyskinesia via storage and non-physiological release of dopamine following L-DOPA administration (Carta et al. 2007). On the other hand, we and others have recently shown significant neuronal loss in the raphe nuclei (Fig. 4 C, D) (Masilamoni et al. 2011a; Unguez and Schneider 1988) and a corresponding decrease in serotonin innervation of the prefrontal cortex and striatum (Masilamoni et al. 2011a; Pifl et al. 1991; Russ et al. 1991) in chronically MPTP-treated monkeys. The variations of MPTP dosage schedules and survival times after intoxication probably account, at least in part, for the differences in the pattern of serotonin loss between these studies.
Cholinergic pathology
Three major sources of acetylcholine display significant pathology in PD. Loss of basal forebrain cholinergic neurons in the nucleus basalis of Meynert (nbM), which provide inputs to the entire cortical mantle, is well documented in PD brains (Candy et al. 1983; Nakano and Hirano 1984; Rogers et al. 1985; Tagliavini et al. 1984; Whitehouse et al. 1983). A significant correlation between the extent of cortical cholinergic denervation and the severity of cognitive dysfunction has been reported in some PD patients (Bohnen and Albin 2011; Korczyn 2001; Perry et al. 1985). However, very limited data has been collected about the state of the nbM cholinergic neurons in MPTP-treated monkeys. To our knowledge, the only significant report comes from (Garvey et al. 1986), who observed no significant cholinergic cell loss in nbM of acutely MPTP-treated marmosets. However, low dose chronic MPTP administration to macaque monkeys significantly reduced cortical cholinergic innervation, and induced changes in nicotinic AChR expression in the frontal cortex, similar to those observed in human PD patients (Aubert et al. 1992; Court and Clementi 1995; Court et al. 2000; Forgacs and Bodis-Wollner 2004; Lange et al. 1993; Mundinano et al. 2013). Furthermore, stable early cognitive deficits that can be improved with the β4*-selective nAChR agonist SIB-1553A treatment were reported in macaque monkeys chronically treated with low doses of MPTP (Schneider and Kovelowski 1990; Schneider et al. 1999). Additional pathological studies are needed to further assess the state of the basal forebrain cholinergic cell group and its cortical innervation in chronically MPTP-treated monkeys.
The pedunculopontine nucleus-laterodorsal tegmental complex (PPN) is another cholinergic cell group of potential importance in PD pathology. Cholinergic neurons in this region provide widespread ascending and descending inputs to the all basal ganglia nuclei, the basal forebrain, thalamus, cerebellum, several brainstem nuclei, and the spinal cord (Heckers et al. 1992). Importance of these systems in regulating sleep-wake cycle, vigilance, attention, gait and other motor functions has been suggested (Datta et al. 2002; Delwaide 2001; Gilman et al. 2003; Gut and Winn 2016; Munro-Davies et al. 1999; Nandi et al. 2002; Steriade 2005; Takakusaki et al. 2004; Thevathasan et al. 2010). Various neuropathological studies reported that around 50% of the cholinergic neurons are degenerated in the lateral part of the PPN, pars compacta in PD (Gai et al. 1991; Hirsch et al. 1987; Jellinger 1988; Zweig et al. 1989). Recently, (Karachi et al. 2010) reported that aged, but not young, MPTP-treated monkeys display significant PPN cholinergic cell loss. Preliminary studies from our laboratory confirmed the lack of significant cholinergic pathology in the PPN/LDT of chronically MPTP-treated young adult rhesus monkeys (Masilamoni et al., unpublished observations; Fig. 4 E–F). Thus, chronic MPTP intoxication can recapitulate some aspect of the brainstem cholinergic pathology in aged, but not young adult monkeys. Further studies are needed to determine the extent of non-cholinergic cell loss in the PPN/LDT region of both PD patients and MPTP-treated monkeys.
Another major group of cholinergic neurons known to play an important role in basal ganglia function in normal and diseased states are the striatal cholinergic interneurons (Tanimura et al. 2017). Although there is evidence for dysregulation of the intrastriatal cholinergic function in PD, very few studies have assessed changes in the morphology or number of striatal cholinergic interneurons in PD patients (Gonzales and Smith 2015; Kharkwal et al. 2016). Two recent studies addressed this issue in MPTP-treated monkeys and came up to different conclusions. On one hand, some authors reported that the total number of striatal cholinergic neurons is not changed in the striatum of parkinsonian monkeys, but that a significantly larger number of cholinergic neurons co-express calretinin immunoreactivity in parkinsonian animals than in controls (Petryszyn et al. 2016). On the other hand, recent findings from our laboratory showed a 30–40% increase in the total number of ChAT-IR neurons in the head of the caudate nucleus of chronically MPTP-treated monkeys, with no significant change in other striatal regions (caudate body, pre- and post-commissural putamen) (Villalba and Smith 2017). Although, there is no clear explanation for the discrepancy between these studies, the different MPTP regimen, monkeys were exposed to may be a contributing factor.
Quantitative studies of the number and density of striatal cholinergic interneurons in the striatum of PD patients are needed to validate the significance of these data from MPTP-treated monkeys towards our understanding of striatal cholinergic pathology in PD.
Olfactory pathology
Deficits in odor detection, discrimination and identification are commonly seen during the prodromal stage of PD (Berendse and Ponsen 2009; Mesholam et al. 1998). This olfactory dysfunction does not respond to L-DOPA therapy (Doty et al. 1992; Hawkes et al. 1997). Various groups have reported an increased number of DA cells in the olfactory bulb (OB) of PD patients with olfactory deficits (Berendse et al. 2001; Hawkes et al. 1997; Huisman et al. 2004). This increased DA activity in the OB of PD patients may lead to a suppression of olfactory information due to the inhibitory effect of DA on the transmission between olfactory receptor cells and mitral cells within the olfactory glomeruli (Doty and Risser 1989; Duchamp-Viret et al. 1997; Hsia et al. 1999; Koster et al. 1999; Mundinano et al. 2011; Wilson and Sullivan 1995). Recent findings revealed that DA levels are also increased in the OB of chronically MPTP-treated monkeys irrespective of their state of parkinsonism, such that motor asymptomatic and moderate or severe parkinsonian animals display comparable increases in OB DA levels (Pifl et al. 2017). These findings are consistent with the previous report of an increased number of intrinsic DA neurons in the OB of chronically MPTP-treated monkeys (Belzunegui et al. 2007). However, because olfactory tests were not performed in these monkeys, the significance of these anatomical observations remains unclear and warrants further studies.
Thalamus CM/PF pathology
A profound degeneration (~40–50% cell loss) of the caudal intralaminar thalamic nuclei, namely the centromedian (CM) and parafascicular nuclei (Pf) has been reported in idiopathic PD patients (Henderson et al. 2000a; Henderson et al. 2000b). The extent of CM/Pf cells loss is not related to the severity of parkinsonian motor signs. Even patients in early stages of the disease harbor major thalamic pathology (Henderson et al. 2000a; Henderson et al. 2000b), suggesting that CM/Pf degeneration is an early event that likely starts during the prodromal period of PD, even before the degeneration of midbrain dopaminergic neurons (See (Smith et al. 2014) for discussion). Because CM/Pf neurons are the main sources of the thalamostriatal glutamatergic system (Galvan and Smith 2011; Smith et al. 2014; Smith et al. 2009; Smith et al. 2004; Smith et al. 2011), early degeneration of these neurons may significantly disrupt the glutamatergic drive of striatal projection neurons and cholinergic interneurons. In light of recent studies showing that CM/Pf neurons are sensitive to salient sensory stimuli and may play an important role in cognitive processes related to attention and reinforcement, it has been suggested that the loss of CM/Pf neurons may contribute to early cognitive impairments in attentional set-shifting or behavioral switching commonly found in PD patients (Smith et al. 2014).
We have recently shown that CM/Pf degeneration can be induced in chronically MPTP-treated rhesus monkeys (Villalba et al. 2014). Although the mechanisms by which MPTP kills these glutamatergic thalamic cells remain to be established, there is evidence that CM/Pf loss occurs early during the course of MPTP intoxication, an reach its maximum extent even before the development of parkinsonian motor signs. As expected, this thalamic cell loss results in decreased thalamic innervation of striatal neurons (Villalba and Smith 2011). Behavioral studies are currently in progress to assess the potential role of CM/Pf pathology on cognitive and motor functions in these monkeys. Thus, the chronic MPTP-treated monkey model of PD is a suitable tool to assess the potential contribution of the thalamostriatal degeneration to early attentional cognitive deficits in PD (Decamp and Schneider 2004; Decamp and Schneider 2006; Ko et al. 2016; Roeltgen and Schneider 1994; Schneider 2006).
Lewy Body Pathology in Monkeys
In addition to the profound degeneration of various dopaminergic and non-dopaminergic cell groups, another key element of PD pathology is the development of cytoplasmic inclusions called Lewy bodies. These inclusions are made up of protein aggregates that are found throughout the whole brain of PD patients, but are particularly abundant in lower and upper brainstem regions and the ventral midbrain dopaminergic cell groups (Baba et al. 1998; Devine et al. 2011; Irizarry et al. 1998; Spillantini et al. 1997). One of the key component protein of Lewy bodies is α-synuclein, a ubiquitous pre-synaptic protein that likely plays an important role in the pre-synaptic regulation of neurotransmission throughout the brain. Because of its abundance in Lewy bodies (Niu et al. 2015), alpha-synuclein has become a major target of PD pathobiology. Its abnormal aggregation throughout the parkinsonian brain may lead to disruption of axonal transport and contribute to a wide array of motor and non-motor symptoms of PD. Recent evidence that these aggregates may be transported throughout the brain in a prion-like fashion has generated significant interest in the field (Braak and Braak 2000; Braak et al. 2003a; Chu and Kordower 2015; Dunning et al. 2013; Dunning et al. 2012; Rey et al. 2016). However, significant controversy exists in understanding the mechanisms by which these aggregates are transported from one structure to another in the mammalian CNS (Surmeier et al. 2017).
Because of the increased significance of alpha-synuclein pathology in the etiology, degeneration process and possible development of PD motor and non-motor symptoms, efforts at developing a nonhuman primate model of PD that displays alpha-synuclein aggregates and Lewy body-like pathology could be of utmost importance for future therapeutic and neuroprotective studies in PD. As discussed in another review of this Special Issue (Marmion and Kordower 2017), attempts at developing such model are currently in progress in various laboratories (Collier et al. 2016; Low and Aebischer 2012; Niu et al. 2015; Recasens et al. 2014; Yang et al. 2015). Although transgenic models offer insights into the causes of PD pathogenesis or LB-like formation, the absence of consistent neuronal loss in the SNc remains a major limitation for these models. On the other hand, viral transduction of wild type or mutant α-synuclein protein has been shown to induce loss of DA neurons, and cause motor impairment in both mice and rats (Oliveras-Salva et al. 2013) but it is limited to the region targeted (Eslamboli et al. 2007; Koprich et al. 2016; Yang et al. 2015). Recently, Shimozawa et al. (2017) reported that intracerebral injection of synthetic α-synuclein fibrils into wild-type marmoset brains (caudate nucleus and/or putamen) results in the formation of abundant α-synuclein-positive inclusions in midbrain TH-positive neurons, suggesting the retrograde spreading of abnormal α-synuclein from the striatum to the SNC. Moreover, the authors reported a significant decrease in the number of nigral DA neurons ipsilateral to the striatal injection sites of α-synuclein fibrils. However, despite such pathology, these monkeys did not exhibit any apparent motor deficits up to 3 months after inoculation. Further studies are needed to establish the relationship between pathologies and symptoms in wild-type marmosets.
A common criticism raised about the MPTP-treated NHP model of PD is that it fails to produce cytoplasmic inclusion bodies in DA neurons and other brain regions. Although protein aggregates structurally similar to Lewy bodies are, indeed, not found in the brain of MPTP-treated monkeys, there is strong evidence that chronic MPTP treatment induces the formation of alpha-synuclein-positive inclusion bodies or aggregates. Early reports have described eosine-positive inclusion bodies in the SNC, LC, nucleus basalis of Meynert, dorsal motor nucleus of the vagus, and raphe nucleus of MPTP-treated aged squirrel monkeys (Forno et al. 1993; Forno et al. 1986). More recently, we and others have demonstrated α-synuclein aggregation in midbrain DA neurons and dystrophic nigrostriatal axons in chronically MPTP-treated young adult squirrel and rhesus monkeys (Masilamoni and Smith 2011; McCormack et al. 2008) (Fig. 5 A–D). Although this issue remains to be addressed in a more systematic fashion, these observations suggest that MPTP treatment induces alpha-synuclein inclusions only when mild and prolonged inhibition of the mitochondrial respiratory chain causes a chronic decrease of the ubiquitin-proteasome activity.
Figure 5.

Photomicrographs of α-synuclein-immunostained coronal sections at the level of the ventral midbrain (A–D) of a control (left column) and a MPTP-treated (right column) monkey. Alpha-synuclein-positive aggregates in SNCd and VTA of chronically MPTP-treated monkeys are shown in B and D.
Concluding Remarks
In conclusion, the MPTP-treated monkey model has been recognized as the gold standard model of PD because the degeneration of the dopaminergic nigrostriatal system leads to parkinsonian motor signs that closely mimic those seen in PD patients. This model has been instrumental to our understanding of the pathophysiology of the basal ganglia-thalamocortical loops in PD and to the rejuvenation and refinement of surgical therapies for the disease.
However, as is the case for most neurotoxin-based models of PD, it is recognized by many investigators as a mere model of nigrostriatal dopaminergic lesion that does not involve other monoaminergic brain systems affected in PD. In this review, we discussed series of data published by various groups over the years that negate this misconception. It is clear from the literature that monkeys treated chronically with low doses of MPTP display brain pathological changes that extend far beyond the nigral dopaminergic neurons. Among the various systems that are known to be partly degenerated in PD, chronically MPTP-treated monkeys exhibit loss of noradrenergic neurons in the locus coeruleus and related brainstem regions, serotonergic neurons in the dorsal raphe, glutamatergic neurons in the thalamic CM/Pf nuclear complex and loss of cholinergic neurons in the pedunculopontine region (only in aged animals). Because these systems are involved in a wide range of cognitive and limbic functions and that their disruption leads to various neuropsychiatric and sleep disorders, their sensitivity to chronic MPTP intoxication adds some significant value to the MPTP-treated monkey model as a tool to study the pathophysiology and develop new therapies for non-motor features of PD.
Acknowledgments
The authors acknowledge Mrs. Susan Jenkins and Jean-Francois Pare for their technical support. This work was supported by NIH/ORIP grant P51-OD011132 to the Yerkes National Primate Research Center, grants from P50NS098685-NINDS (UDALL PD Center) and R01NS083386-NINDS.
Footnotes
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
References
- Aarsland D, Andersen K, Larsen JP, Perry R, Wentzel-Larsen T, Lolk A, Kragh-Sorensen P. Archives of neurology. 2004;61:1906–1911. doi: 10.1001/archneur.61.12.1906. [DOI] [PubMed] [Google Scholar]
- Aarsland D, Larsen JP, Lim NG, Janvin C, Karlsen K, Tandberg E, Cummings JL. Range of neuropsychiatric disturbances in patients with Parkinson’s disease. Journal of neurology, neurosurgery, and psychiatry. 1999;67:492–496. doi: 10.1136/jnnp.67.4.492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abbott RD, et al. Excessive daytime sleepiness and subsequent development of Parkinson disease. Neurology. 2005;65:1442–1446. doi: 10.1212/01.wnl.0000183056.89590.0d. [DOI] [PubMed] [Google Scholar]
- Airaksinen MS, Thoenen H, Meyer M. Vulnerability of midbrain dopaminergic neurons in calbindin-D28k-deficient mice: lack of evidence for a neuroprotective role of endogenous calbindin in MPTP-treated and weaver mice. Eur J Neurosci. 1997;9:120–127. doi: 10.1111/j.1460-9568.1997.tb01360.x. [DOI] [PubMed] [Google Scholar]
- Akil M, Lewis DA. The distribution of tyrosine hydroxylase-immunoreactive fibers in the human entorhinal cortex. Neuroscience. 1994;60:857–874. doi: 10.1016/0306-4522(94)90268-2. [DOI] [PubMed] [Google Scholar]
- Akil M, Pierri JN, Whitehead RE, Edgar CL, Mohila C, Sampson AR, Lewis DA. Lamina-specific alterations in the dopamine innervation of the prefrontal cortex in schizophrenic subjects. Am J Psychiatry. 1999;156:1580–1589. doi: 10.1176/ajp.156.10.1580. [DOI] [PubMed] [Google Scholar]
- Albin RL, Koeppe RA, Bohnen NI, Wernette K, Kilbourn MA, Frey KA. Spared caudal brainstem SERT binding in early Parkinson’s disease. J Cereb Blood Flow Metab. 2008;28:441–444. doi: 10.1038/sj.jcbfm.9600599. [DOI] [PubMed] [Google Scholar]
- Alexander GM, Schwartzman RJ, Brainard L, Gordon SW, Grothusen JR. Changes in brain catecholamines and dopamine uptake sites at different stages of MPTP parkinsonism in monkeys. Brain research. 1992;588:261–269. doi: 10.1016/0006-8993(92)91584-2. [DOI] [PubMed] [Google Scholar]
- Arnsten AF, Cai JX, Steere JC, Goldman-Rakic PS. Dopamine D2 receptor mechanisms contribute to age-related cognitive decline: the effects of quinpirole on memory and motor performance in monkeys. J Neurosci. 1995;15:3429–3439. doi: 10.1523/JNEUROSCI.15-05-03429.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnsten AF, Wang M, Paspalas CD. Dopamine’s Actions in Primate Prefrontal Cortex: Challenges for Treating Cognitive Disorders. Pharmacol Rev. 2015;67:681–696. doi: 10.1124/pr.115.010512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aubert I, Araujo DM, Cecyre D, Robitaille Y, Gauthier S, Quirion R. Comparative alterations of nicotinic and muscarinic binding sites in Alzheimer’s and Parkinson’s diseases. J Neurochem. 1992;58:529–541. doi: 10.1111/j.1471-4159.1992.tb09752.x. [DOI] [PubMed] [Google Scholar]
- Augood SJ, Hollingsworth ZR, Standaert DG, Emson PC, Penney JB., Jr Localization of dopaminergic markers in the human subthalamic nucleus. J Comp Neurol. 2000;421:247–255. [PubMed] [Google Scholar]
- Baba M, et al. Aggregation of alpha-synuclein in Lewy bodies of sporadic Parkinson’s disease and dementia with Lewy bodies. Am J Pathol. 1998;152:879–884. [PMC free article] [PubMed] [Google Scholar]
- Ballanger B, et al. Imaging Dopamine and Serotonin Systems on MPTP Monkeys: A Longitudinal PET Investigation of Compensatory Mechanisms. J Neurosci. 2016;36:1577–1589. doi: 10.1523/JNEUROSCI.2010-15.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barraud Q, et al. Sleep disorders in Parkinson’s disease: the contribution of the MPTP non-human primate model. Exp Neurol. 2009;219:574–582. doi: 10.1016/j.expneurol.2009.07.019. [DOI] [PubMed] [Google Scholar]
- Beaudoin-Gobert M, et al. Behavioural impact of a double dopaminergic and serotonergic lesion in the non-human primate. Brain. 2015;138:2632–2647. doi: 10.1093/brain/awv183. [DOI] [PubMed] [Google Scholar]
- Belzunegui S, et al. The number of dopaminergic cells is increased in the olfactory bulb of monkeys chronically exposed to MPTP. Synapse. 2007;61:1006–1012. doi: 10.1002/syn.20451. [DOI] [PubMed] [Google Scholar]
- Benazzouz A, Mamad O, Abedi P, Bouali-Benazzouz R, Chetrit J. Involvement of dopamine loss in extrastriatal basal ganglia nuclei in the pathophysiology of Parkinson’s disease. Front Aging Neurosci. 2014;6:87. doi: 10.3389/fnagi.2014.00087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berendse HW, Booij J, Francot CM, Bergmans PL, Hijman R, Stoof JC, Wolters EC. Subclinical dopaminergic dysfunction in asymptomatic Parkinson’s disease patients’ relatives with a decreased sense of smell. Ann Neurol. 2001;50:34–41. doi: 10.1002/ana.1049. [DOI] [PubMed] [Google Scholar]
- Berendse HW, Ponsen MM. Diagnosing premotor Parkinson’s disease using a two-step approach combining olfactory testing and DAT SPECT imaging. Parkinsonism Relat Disord. 2009;15(Suppl 3):S26–30. doi: 10.1016/S1353-8020(09)70774-6. [DOI] [PubMed] [Google Scholar]
- Berger B, Gaspar P, Verney C. Dopaminergic innervation of the cerebral cortex: unexpected differences between rodents and primates. Trends Neurosci. 1991;14:21–27. doi: 10.1016/0166-2236(91)90179-x. [DOI] [PubMed] [Google Scholar]
- Berger B, Gaspar P, Verney C. Colocalization of neurotensin in the mesocortical dopaminergic system. Restricted regional and laminar distribution in rat, lack of colocalization in human. Ann N Y Acad Sci. 1992;668:307–310. doi: 10.1111/j.1749-6632.1992.tb27359.x. [DOI] [PubMed] [Google Scholar]
- Berger B, Trottier S, Verney C, Gaspar P, Alvarez C. Regional and laminar distribution of the dopamine and serotonin innervation in the macaque cerebral cortex: a radioautographic study. J Comp Neurol. 1988;273:99–119. doi: 10.1002/cne.902730109. [DOI] [PubMed] [Google Scholar]
- Bergman H, Wichmann T, Karmon B, DeLong MR. The primate subthalamic nucleus. II. Neuronal activity in the MPTP model of parkinsonism. J Neurophysiol. 1994;72:507–520. doi: 10.1152/jn.1994.72.2.507. [DOI] [PubMed] [Google Scholar]
- Bernheimer H, Birkmayer W, Hornykiewicz O, Jellinger K, Seitelberger F. Brain dopamine and the syndromes of Parkinson and Huntington. Clinical, morphological and neurochemical correlations. J Neurol Sci. 1973;20:415–455. doi: 10.1016/0022-510x(73)90175-5. [DOI] [PubMed] [Google Scholar]
- Betarbet R, Sherer TB, MacKenzie G, Garcia-Osuna M, Panov AV, Greenamyre JT. Chronic systemic pesticide exposure reproduces features of Parkinson’s disease. Nat Neurosci. 2000;3:1301–1306. doi: 10.1038/81834. [DOI] [PubMed] [Google Scholar]
- Bezard E, et al. Relationship between the appearance of symptoms and the level of nigrostriatal degeneration in a progressive 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-lesioned macaque model of Parkinson’s disease. J Neurosci. 2001;21:6853–6861. doi: 10.1523/JNEUROSCI.21-17-06853.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bezard E, Przedborski S. A tale on animal models of Parkinson’s disease. Mov Disord. 2011;26:993–1002. doi: 10.1002/mds.23696. [DOI] [PubMed] [Google Scholar]
- Blandini F, Armentero MT. Animal models of Parkinson’s disease. FEBS J. 2012;279:1156–1166. doi: 10.1111/j.1742-4658.2012.08491.x. [DOI] [PubMed] [Google Scholar]
- Blesa J, et al. The nigrostriatal system in the presymptomatic and symptomatic stages in the MPTP monkey model: a PET, histological and biochemical study. Neurobiol Dis. 2012;48:79–91. doi: 10.1016/j.nbd.2012.05.018. [DOI] [PubMed] [Google Scholar]
- Blier P. Psychopharmacology for the clinician. Treating depression with selective norepinephrine reuptake inhibitors. J Psychiatry Neurosci. 2006;31:288. [PMC free article] [PubMed] [Google Scholar]
- Bohnen NI, Albin RL. The cholinergic system and Parkinson disease. Behav Brain Res. 2011;221:564–573. doi: 10.1016/j.bbr.2009.12.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boraud T, Bezard E, Guehl D, Bioulac B, Gross C. Effects of L-DOPA on neuronal activity of the globus pallidus externalis (GPe) and globus pallidus internalis (GPi) in the MPTP-treated monkey. Brain Res. 1998;787:157–160. doi: 10.1016/s0006-8993(97)01563-1. [DOI] [PubMed] [Google Scholar]
- Boulet S, et al. Behavioral recovery in MPTP-treated monkeys: neurochemical mechanisms studied by intrastriatal microdialysis. J Neurosci. 2008;28:9575–9584. doi: 10.1523/JNEUROSCI.3465-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bove J, Perier C. Neurotoxin-based models of Parkinson’s disease. Neuroscience. 2012;211:51–76. doi: 10.1016/j.neuroscience.2011.10.057. [DOI] [PubMed] [Google Scholar]
- Boyce S, Rupniak NM, Steventon MJ, Iversen SD. Characterisation of dyskinesias induced by L-dopa in MPTP-treated squirrel monkeys. Psychopharmacology (Berl) 1990;102:21–27. doi: 10.1007/BF02245739. [DOI] [PubMed] [Google Scholar]
- Braak H, Braak E. Pathoanatomy of Parkinson’s disease. J Neurol. 2000;247(Suppl 2):II3–10. doi: 10.1007/PL00007758. [DOI] [PubMed] [Google Scholar]
- Braak H, Del Tredici K, Rub U, de Vos RA, Jansen Steur EN, Braak E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol Aging. 2003a;24:197–211. doi: 10.1016/s0197-4580(02)00065-9. [DOI] [PubMed] [Google Scholar]
- Braak H, Del Tredici K, Rub U, de Vos RA, Jansen Steur EN, Braak E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiology of aging. 2003b;24:197–211. doi: 10.1016/s0197-4580(02)00065-9. [DOI] [PubMed] [Google Scholar]
- Brooks DJ, Doder M. Depression in Parkinson’s disease. Curr Opin Neurol. 2001;14:465–470. doi: 10.1097/00019052-200108000-00006. [DOI] [PubMed] [Google Scholar]
- Brooks DJ, Piccini P. Imaging in Parkinson’s disease: the role of monoamines in behavior. Biol Psychiatry. 2006;59:908–918. doi: 10.1016/j.biopsych.2005.12.017. [DOI] [PubMed] [Google Scholar]
- Brown RG, Marsden CD. Internal versus external cues and the control of attention in Parkinson’s disease. Brain. 1988;111(Pt 2):323–345. doi: 10.1093/brain/111.2.323. [DOI] [PubMed] [Google Scholar]
- Brown RM, Crane AM, Goldman PS. Regional distribution of monoamines in the cerebral cortex and subcortical structures of the rhesus monkey: concentrations and in vivo synthesis rates. Brain Res. 1979;168:133–150. doi: 10.1016/0006-8993(79)90132-x. [DOI] [PubMed] [Google Scholar]
- Brownell AL, et al. Mapping of brain function after MPTP-induced neurotoxicity in a primate Parkinson’s disease model. Neuroimage. 2003;20:1064–1075. doi: 10.1016/S1053-8119(03)00348-3. [DOI] [PubMed] [Google Scholar]
- Buddhala C, Loftin SK, Kuley BM, Cairns NJ, Campbell MC, Perlmutter JS, Kotzbauer PT. Dopaminergic, serotonergic, and noradrenergic deficits in Parkinson disease. Ann Clin Transl Neurol. 2015;2:949–959. doi: 10.1002/acn3.246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burns RS, Chiueh CC, Markey SP, Ebert MH, Jacobowitz DM, Kopin IJ. A primate model of parkinsonism: selective destruction of dopaminergic neurons in the pars compacta of the substantia nigra by N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Proc Natl Acad Sci U S A. 1983;80:4546–4550. doi: 10.1073/pnas.80.14.4546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Candy JM, Perry RH, Perry EK, Irving D, Blessed G, Fairbairn AF, Tomlinson BE. Pathological changes in the nucleus of Meynert in Alzheimer’s and Parkinson’s diseases. J Neurol Sci. 1983;59:277–289. doi: 10.1016/0022-510x(83)90045-x. [DOI] [PubMed] [Google Scholar]
- Carbon M, Edwards C, Eidelberg D. Functional brain imaging in Parkinson’s disease. Adv Neurol. 2003;91:175–181. [PubMed] [Google Scholar]
- Carta M, Carlsson T, Kirik D, Bjorklund A. Dopamine released from 5-HT terminals is the cause of L-DOPA-induced dyskinesia in parkinsonian rats. Brain. 2007;130:1819–1833. doi: 10.1093/brain/awm082. [DOI] [PubMed] [Google Scholar]
- Cenci MA, Francardo V, O’Sullivan SS, Lindgren HS. Rodent models of impulsive-compulsive behaviors in Parkinson’s disease: How far have we reached? Neurobiol Dis. 2015;82:561–573. doi: 10.1016/j.nbd.2015.08.026. [DOI] [PubMed] [Google Scholar]
- Chan-Palay V. Alterations in the locus coeruleus in dementias of Alzheimer’s and Parkinson’s disease. Progress in brain research. 1991;88:625–630. doi: 10.1016/s0079-6123(08)63839-x. [DOI] [PubMed] [Google Scholar]
- Chu Y, Kordower JH. The prion hypothesis of Parkinson’s disease. Curr Neurol Neurosci Rep. 2015;15:28. doi: 10.1007/s11910-015-0549-x. [DOI] [PubMed] [Google Scholar]
- Collier TJ, et al. Aging-related changes in the nigrostriatal dopamine system and the response to MPTP in nonhuman primates: diminished compensatory mechanisms as a prelude to parkinsonism. Neurobiol Dis. 2007;26:56–65. doi: 10.1016/j.nbd.2006.11.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collier TJ, Redmond DE, Jr, Steece-Collier K, Lipton JW, Manfredsson FP. Is Alpha-Synuclein Loss-of-Function a Contributor to Parkinsonian Pathology? Evidence from Non-human Primates Front Neurosci. 2016;10:12. doi: 10.3389/fnins.2016.00012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooney JW, Stacy M. Neuropsychiatric Issues in Parkinson’s Disease. Curr Neurol Neurosci Rep. 2016;16:49. doi: 10.1007/s11910-016-0647-4. [DOI] [PubMed] [Google Scholar]
- Cossette M, Levesque M, Parent A. Extrastriatal dopaminergic innervation of human basal ganglia. Neurosci Res. 1999;34:51–54. doi: 10.1016/s0168-0102(99)00029-2. [DOI] [PubMed] [Google Scholar]
- Coull JT, Buchel C, Friston KJ, Frith CD. Noradrenergically mediated plasticity in a human attentional neuronal network. Neuroimage. 1999;10:705–715. doi: 10.1006/nimg.1999.0513. [DOI] [PubMed] [Google Scholar]
- Court J, Clementi F. Distribution of nicotinic subtypes in human brain. Alzheimer Dis Assoc Disord. 1995;9(Suppl 2):6–14. doi: 10.1097/00002093-199501002-00003. [DOI] [PubMed] [Google Scholar]
- Court JA, et al. Nicotine binding in human striatum: elevation in schizophrenia and reductions in dementia with Lewy bodies, Parkinson’s disease and Alzheimer’s disease and in relation to neuroleptic medication. Neuroscience. 2000;98:79–87. doi: 10.1016/s0306-4522(00)00071-3. [DOI] [PubMed] [Google Scholar]
- Cragg SJ, Baufreton J, Xue Y, Bolam JP, Bevan MD. Synaptic release of dopamine in the subthalamic nucleus. Eur J Neurosci. 2004;20:1788–1802. doi: 10.1111/j.1460-9568.2004.03629.x. [DOI] [PubMed] [Google Scholar]
- Crittenden JR, et al. Striosome-dendron bouquets highlight a unique striatonigral circuit targeting dopamine-containing neurons. Proc Natl Acad Sci U S A. 2016;113:11318–11323. doi: 10.1073/pnas.1613337113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cummings JL. Depression and Parkinson’s disease: a review. Am J Psychiatry. 1992;149:443–454. doi: 10.1176/ajp.149.4.443. [DOI] [PubMed] [Google Scholar]
- D’Amato RJ, Lipman ZP, Snyder SH. Selectivity of the parkinsonian neurotoxin MPTP: toxic metabolite MPP+ binds to neuromelanin. Science. 1986;231:987–989. doi: 10.1126/science.3080808. [DOI] [PubMed] [Google Scholar]
- D’Amato RJ, et al. Aminergic systems in Alzheimer’s disease and Parkinson’s disease. Ann Neurol. 1987;22:229–236. doi: 10.1002/ana.410220207. [DOI] [PubMed] [Google Scholar]
- Damier P, Hirsch EC, Agid Y, Graybiel AM. The substantia nigra of the human brain. I. Nigrosomes and the nigral matrix, a compartmental organization based on calbindin D(28K) immunohistochemistry. Brain. 1999a;122(Pt 8):1421–1436. doi: 10.1093/brain/122.8.1421. [DOI] [PubMed] [Google Scholar]
- Damier P, Hirsch EC, Agid Y, Graybiel AM. The substantia nigra of the human brain. I. Nigrosomes and the nigral matrix, a compartmental organization based on calbindin D(28K) immunohistochemistry. Brain: a journal of neurology. 1999b;122(Pt 8):1421–1436. doi: 10.1093/brain/122.8.1421. [DOI] [PubMed] [Google Scholar]
- Damier P, Hirsch EC, Agid Y, Graybiel AM. The substantia nigra of the human brain. II. Patterns of loss of dopamine-containing neurons in Parkinson’s disease. Brain. 1999c;122(Pt 8):1437–1448. doi: 10.1093/brain/122.8.1437. [DOI] [PubMed] [Google Scholar]
- Damier P, Hirsch EC, Agid Y, Graybiel AM. The substantia nigra of the human brain. II. Patterns of loss of dopamine-containing neurons in Parkinson’s disease. Brain: a journal of neurology. 1999d;122(Pt 8):1437–1448. doi: 10.1093/brain/122.8.1437. [DOI] [PubMed] [Google Scholar]
- Datta S, Spoley EE, Mavanji VK, Patterson EH. A novel role of pedunculopontine tegmental kainate receptors: a mechanism of rapid eye movement sleep generation in the rat. Neuroscience. 2002;114:157–164. doi: 10.1016/s0306-4522(02)00250-6. [DOI] [PubMed] [Google Scholar]
- Dauer W, Przedborski S. Parkinson’s disease: mechanisms and models. Neuron. 2003;39:889–909. doi: 10.1016/s0896-6273(03)00568-3. [DOI] [PubMed] [Google Scholar]
- Davis MR, et al. Initial human PET imaging studies with the dopamine transporter ligand 18F-FECNT. J Nucl Med. 2003;44:855–861. [PubMed] [Google Scholar]
- Decamp E, Schneider JS. Attention and executive function deficits in chronic low-dose MPTP-treated non-human primates. Eur J Neurosci. 2004;20:1371–1378. doi: 10.1111/j.1460-9568.2004.03586.x. [DOI] [PubMed] [Google Scholar]
- Decamp E, Schneider JS. Effects of nicotinic therapies on attention and executive functions in chronic low-dose MPTP-treated monkeys. Eur J Neurosci. 2006;24:2098–2104. doi: 10.1111/j.1460-9568.2006.05077.x. [DOI] [PubMed] [Google Scholar]
- Del Tredici K, Braak H. Dysfunction of the locus coeruleus-norepinephrine system and related circuitry in Parkinson’s disease-related dementia. J Neurol Neurosurg Psychiatry. 2013;84:774–783. doi: 10.1136/jnnp-2011-301817. [DOI] [PubMed] [Google Scholar]
- Del Tredici K, Rub U, De Vos RA, Bohl JR, Braak H. Where does parkinson disease pathology begin in the brain? Journal of neuropathology and experimental neurology. 2002;61:413–426. doi: 10.1093/jnen/61.5.413. [DOI] [PubMed] [Google Scholar]
- Delwaide PJ. Parkinsonian rigidity. Funct Neurol. 2001;16:147–156. [PubMed] [Google Scholar]
- Deutch AY, Goldstein M, Baldino F, Jr, Roth RH. Telencephalic projections of the A8 dopamine cell group. Ann N Y Acad Sci. 1988;537:27–50. doi: 10.1111/j.1749-6632.1988.tb42095.x. [DOI] [PubMed] [Google Scholar]
- Devine MJ, Gwinn K, Singleton A, Hardy J. Parkinson’s disease and alpha-synuclein expression. Mov Disord. 2011;26:2160–2168. doi: 10.1002/mds.23948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devoto P, Flore G, Saba P, Cadeddu R, Gessa GL. Disulfiram stimulates dopamine release from noradrenergic terminals and potentiates cocaine-induced dopamine release in the prefrontal cortex. Psychopharmacology (Berl) 2012;219:1153–1164. doi: 10.1007/s00213-011-2447-5. [DOI] [PubMed] [Google Scholar]
- Di Monte DA, McCormack A, Petzinger G, Janson AM, Quik M, Langston WJ. Relationship among nigrostriatal denervation, parkinsonism, and dyskinesias in the MPTP primate model. Mov Disord. 2000;15:459–466. doi: 10.1002/1531-8257(200005)15:3<459::AID-MDS1006>3.0.CO;2-3. [DOI] [PubMed] [Google Scholar]
- Dogali M, et al. Anatomic and physiological considerations in pallidotomy for Parkinson’s disease. Stereotact Funct Neurosurg. 1994;62:53–60. doi: 10.1159/000098597. [DOI] [PubMed] [Google Scholar]
- Dopeso-Reyes IG, et al. Calbindin content and differential vulnerability of midbrain efferent dopaminergic neurons in macaques. Front Neuroanat. 2014;8:146. doi: 10.3389/fnana.2014.00146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doty RL, Risser JM. Influence of the D-2 dopamine receptor agonist quinpirole on the odor detection performance of rats before and after spiperone administration. Psychopharmacology (Berl) 1989;98:310–315. doi: 10.1007/BF00451680. [DOI] [PubMed] [Google Scholar]
- Doty RL, Stern MB, Pfeiffer C, Gollomp SM, Hurtig HI. Bilateral olfactory dysfunction in early stage treated and untreated idiopathic Parkinson’s disease. J Neurol Neurosurg Psychiatry. 1992;55:138–142. doi: 10.1136/jnnp.55.2.138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dreher JC, Burnod Y. An integrative theory of the phasic and tonic modes of dopamine modulation in the prefrontal cortex. Neural Netw. 2002;15:583–602. doi: 10.1016/s0893-6080(02)00051-5. [DOI] [PubMed] [Google Scholar]
- Drouot X, et al. Functional recovery in a primate model of Parkinson’s disease following motor cortex stimulation. Neuron. 2004;44:769–778. doi: 10.1016/j.neuron.2004.11.023. [DOI] [PubMed] [Google Scholar]
- Duchamp-Viret P, Coronas V, Delaleu JC, Moyse E, Duchamp A. Dopaminergic modulation of mitral cell activity in the frog olfactory bulb: a combined radioligand binding-electrophysiological study. Neuroscience. 1997;79:203–216. doi: 10.1016/s0306-4522(96)00646-x. [DOI] [PubMed] [Google Scholar]
- Dunnett SB, Lelos M. Behavioral analysis of motor and non-motor symptoms in rodent models of Parkinson’s disease. Prog Brain Res. 2010;184:35–51. doi: 10.1016/S0079-6123(10)84003-8. [DOI] [PubMed] [Google Scholar]
- Dunning CJ, George S, Brundin P. What’s to like about the prion-like hypothesis for the spreading of aggregated alpha-synuclein in Parkinson disease? Prion. 2013;7:92–97. doi: 10.4161/pri.23806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunning CJ, Reyes JF, Steiner JA, Brundin P. Can Parkinson’s disease pathology be propagated from one neuron to another? Prog Neurobiol. 2012;97:205–219. doi: 10.1016/j.pneurobio.2011.11.003. [DOI] [PubMed] [Google Scholar]
- Duty S, Jenner P. Animal models of Parkinson’s disease: a source of novel treatments and clues to the cause of the disease. Br J Pharmacol. 2011;164:1357–1391. doi: 10.1111/j.1476-5381.2011.01426.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eblen F, Graybiel AM. Highly restricted origin of prefrontal cortical inputs to striosomes in the macaque monkey. J Neurosci. 1995;15:5999–6013. doi: 10.1523/JNEUROSCI.15-09-05999.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eckert T, et al. Abnormal metabolic networks in atypical parkinsonism. Mov Disord. 2008;23:727–733. doi: 10.1002/mds.21933. [DOI] [PubMed] [Google Scholar]
- Eidelberg D, et al. The metabolic topography of parkinsonism. J Cereb Blood Flow Metab. 1994;14:783–801. doi: 10.1038/jcbfm.1994.99. [DOI] [PubMed] [Google Scholar]
- Elsworth JD, Deutch AY, Redmond DE, Jr, Taylor JR, Sladek JR, Jr, Roth RH. Symptomatic and asymptomatic 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-treated primates: biochemical changes in striatal regions. Neuroscience. 1989;33:323–331. doi: 10.1016/0306-4522(89)90212-1. [DOI] [PubMed] [Google Scholar]
- Elsworth JD, Taylor JR, Sladek JR, Jr, Collier TJ, Redmond DE, Jr, Roth RH. Striatal dopaminergic correlates of stable parkinsonism and degree of recovery in old-world primates one year after MPTP treatment. Neuroscience. 2000;95:399–408. doi: 10.1016/s0306-4522(99)00437-6. [DOI] [PubMed] [Google Scholar]
- Emborg ME. Nonhuman primate models of Parkinson’s disease. ILAR J. 2007;48:339–355. doi: 10.1093/ilar.48.4.339. [DOI] [PubMed] [Google Scholar]
- Erickson SL, Sesack SR, Lewis DA. Dopamine innervation of monkey entorhinal cortex: postsynaptic targets of tyrosine hydroxylase-immunoreactive terminals. Synapse. 2000;36:47–56. doi: 10.1002/(SICI)1098-2396(200004)36:1<47::AID-SYN5>3.0.CO;2-O. [DOI] [PubMed] [Google Scholar]
- Eslamboli A, et al. Long-term consequences of human alpha-synuclein overexpression in the primate ventral midbrain. Brain. 2007;130:799–815. doi: 10.1093/brain/awl382. [DOI] [PubMed] [Google Scholar]
- Fallon SJ, Smulders K, Esselink RA, van de Warrenburg BP, Bloem BR, Cools R. Differential optimal dopamine levels for set-shifting and working memory in Parkinson’s disease. Neuropsychologia. 2015;77:42–51. doi: 10.1016/j.neuropsychologia.2015.07.031. [DOI] [PubMed] [Google Scholar]
- Fearnley JM, Lees AJ. Ageing and Parkinson’s disease: substantia nigra regional selectivity. Brain. 1991;114(Pt 5):2283–2301. doi: 10.1093/brain/114.5.2283. [DOI] [PubMed] [Google Scholar]
- Fernagut PO, et al. Dopamine transporter binding is unaffected by L-DOPA administration in normal and MPTP-treated monkeys. PLoS One. 2010;5:e14053. doi: 10.1371/journal.pone.0014053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrante RJ, Schulz JB, Kowall NW, Beal MF. Systemic administration of rotenone produces selective damage in the striatum and globus pallidus, but not in the substantia nigra. Brain research. 1997;753:157–162. doi: 10.1016/s0006-8993(97)00008-5. [DOI] [PubMed] [Google Scholar]
- Fifel K, Piggins H, Deboer T. Modeling sleep alterations in Parkinson’s disease: How close are we to valid translational animal models? Sleep Med Rev. 2016;25:95–111. doi: 10.1016/j.smrv.2015.02.005. [DOI] [PubMed] [Google Scholar]
- Filion M, Tremblay L, Bedard PJ. Effects of dopamine agonists on the spontaneous activity of globus pallidus neurons in monkeys with MPTP-induced parkinsonism. Brain Res. 1991;547:152–161. [PubMed] [Google Scholar]
- Flores G, Liang JJ, Sierra A, Martinez-Fong D, Quirion R, Aceves J, Srivastava LK. Expression of dopamine receptors in the subthalamic nucleus of the rat: characterization using reverse transcriptase-polymerase chain reaction and autoradiography. Neuroscience. 1999;91:549–556. doi: 10.1016/s0306-4522(98)00633-2. [DOI] [PubMed] [Google Scholar]
- Forgacs PB, Bodis-Wollner I. Nicotinic receptors and cognition in Parkinson’s Disease: the importance of neuronal synchrony. J Neural Transm (Vienna) 2004;111:1317–1331. doi: 10.1007/s00702-004-0169-0. [DOI] [PubMed] [Google Scholar]
- Fornai F, et al. Parkinson-like syndrome induced by continuous MPTP infusion: convergent roles of the ubiquitin-proteasome system and alpha-synuclein. Proc Natl Acad Sci U S A. 2005;102:3413–3418. doi: 10.1073/pnas.0409713102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forno LS, DeLanney LE, Irwin I, Langston JW. Similarities and differences between MPTP-induced parkinsonsim and Parkinson’s disease. Neuropathologic considerations Adv Neurol. 1993;60:600–608. [PubMed] [Google Scholar]
- Forno LS, DeLanney LE, Irwin I, Langston JW. Ultrastructure of eosinophilic inclusion bodies in the amygdala-parahippocampal region of aged squirrel monkeys treated with 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine, a dopaminergic neurotoxin. Neurosci Lett. 1995;184:44–47. doi: 10.1016/0304-3940(94)11164-e. [DOI] [PubMed] [Google Scholar]
- Forno LS, Langston JW, DeLanney LE, Irwin I, Ricaurte GA. Locus ceruleus lesions and eosinophilic inclusions in MPTP-treated monkeys. Ann Neurol. 1986;20:449–455. doi: 10.1002/ana.410200403. [DOI] [PubMed] [Google Scholar]
- Fox SH, Brotchie JM. The MPTP-lesioned non-human primate models of Parkinson’s disease. Past, present, and future. Prog Brain Res. 2010;184:133–157. doi: 10.1016/S0079-6123(10)84007-5. [DOI] [PubMed] [Google Scholar]
- Francois C, Savy C, Jan C, Tande D, Hirsch EC, Yelnik J. Dopaminergic innervation of the subthalamic nucleus in the normal state, in MPTP-treated monkeys, and in Parkinson’s disease patients. J Comp Neurol. 2000;425:121–129. doi: 10.1002/1096-9861(20000911)425:1<121::aid-cne10>3.0.co;2-g. [DOI] [PubMed] [Google Scholar]
- Freeman A, et al. Nigrostriatal collaterals to thalamus degenerate in parkinsonian animal models. Ann Neurol. 2001;50:321–329. doi: 10.1002/ana.1119. [DOI] [PubMed] [Google Scholar]
- Friedman A, et al. A Corticostriatal Path Targeting Striosomes Controls Decision-Making under Conflict. Cell. 2015;161:1320–1333. doi: 10.1016/j.cell.2015.04.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujiyama F, Sohn J, Nakano T, Furuta T, Nakamura KC, Matsuda W, Kaneko T. Exclusive and common targets of neostriatofugal projections of rat striosome neurons: a single neuron-tracing study using a viral vector. Eur J Neurosci. 2011;33:668–677. doi: 10.1111/j.1460-9568.2010.07564.x. [DOI] [PubMed] [Google Scholar]
- Gai WP, Halliday GM, Blumbergs PC, Geffen LB, Blessing WW. Substance P-containing neurons in the mesopontine tegmentum are severely affected in Parkinson’s disease. Brain. 1991;114(Pt 5):2253–2267. doi: 10.1093/brain/114.5.2253. [DOI] [PubMed] [Google Scholar]
- Galvan A, Hu X, Rommelfanger KS, Pare JF, Khan ZU, Smith Y, Wichmann T. Localization and function of dopamine receptors in the subthalamic nucleus of normal and parkinsonian monkeys. J Neurophysiol. 2014;112:467–479. doi: 10.1152/jn.00849.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galvan A, Smith Y. The primate thalamostriatal systems: Anatomical organization, functional roles and possible involvement in Parkinson’s disease. Basal Ganglia. 2011;1:179–189. doi: 10.1016/j.baga.2011.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia-Cabezas MA, Martinez-Sanchez P, Sanchez-Gonzalez MA, Garzon M, Cavada C. Dopamine innervation in the thalamus: monkey versus rat. Cereb Cortex. 2009;19:424–434. doi: 10.1093/cercor/bhn093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia-Cabezas MA, Rico B, Sanchez-Gonzalez MA, Cavada C. Distribution of the dopamine innervation in the macaque and human thalamus. Neuroimage. 2007;34:965–984. doi: 10.1016/j.neuroimage.2006.07.032. [DOI] [PubMed] [Google Scholar]
- Garvey J, et al. Administration of MPTP to the common marmoset does not alter cortical cholinergic function. Mov Disord. 1986;1:129–134. doi: 10.1002/mds.870010207. [DOI] [PubMed] [Google Scholar]
- Gaspar P, Berger B, Febvret A, Vigny A, Henry JP. Catecholamine innervation of the human cerebral cortex as revealed by comparative immunohistochemistry of tyrosine hydroxylase and dopamine-beta-hydroxylase. J Comp Neurol. 1989;279:249–271. doi: 10.1002/cne.902790208. [DOI] [PubMed] [Google Scholar]
- Gaspar P, Duyckaerts C, Alvarez C, Javoy-Agid F, Berger B. Alterations of dopaminergic and noradrenergic innervations in motor cortex in Parkinson’s disease. Ann Neurol. 1991;30:365–374. doi: 10.1002/ana.410300308. [DOI] [PubMed] [Google Scholar]
- Gaspar P, Febvret A, Colombo J. Serotonergic sprouting in primate MTP-induced hemiparkinsonism. Exp Brain Res. 1993;96:100–106. doi: 10.1007/BF00230443. [DOI] [PubMed] [Google Scholar]
- Gaspar P, Stepniewska I, Kaas JH. Topography and collateralization of the dopaminergic projections to motor and lateral prefrontal cortex in owl monkeys. J Comp Neurol. 1992;325:1–21. doi: 10.1002/cne.903250102. [DOI] [PubMed] [Google Scholar]
- Gauthier J, Parent M, Levesque M, Parent A. The axonal arborization of single nigrostriatal neurons in rats. Brain Res. 1999;834:228–232. doi: 10.1016/s0006-8993(99)01573-5. [DOI] [PubMed] [Google Scholar]
- Gerfen CR. The neostriatal mosaic: compartmentalization of corticostriatal input and striatonigral output systems. Nature. 1984;311:461–464. doi: 10.1038/311461a0. [DOI] [PubMed] [Google Scholar]
- Gerfen CR, Herkenham M, Thibault J. The neostriatal mosaic: II. Patch- and matrix-directed mesostriatal dopaminergic and non-dopaminergic systems. J Neurosci. 1987;7:3915–3934. doi: 10.1523/JNEUROSCI.07-12-03915.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- German DC, Manaye KF, Sonsalla PK, Brooks BA. Midbrain dopaminergic cell loss in Parkinson’s disease and MPTP-induced parkinsonism: sparing of calbindin-D28k-containing cells. Ann N Y Acad Sci. 1992a;648:42–62. doi: 10.1111/j.1749-6632.1992.tb24523.x. [DOI] [PubMed] [Google Scholar]
- German DC, et al. Disease-specific patterns of locus coeruleus cell loss. Annals of neurology. 1992b;32:667–676. doi: 10.1002/ana.410320510. [DOI] [PubMed] [Google Scholar]
- Gibb WR, Terruli M, Lees AJ, Jenner P, Marsden CD. The evolution and distribution of morphological changes in the nervous system of the common marmoset following the acute administration of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Mov Disord. 1989;4:53–74. doi: 10.1002/mds.870040109. [DOI] [PubMed] [Google Scholar]
- Gilman S, et al. REM sleep behavior disorder is related to striatal monoaminergic deficit in MSA. Neurology. 2003;61:29–34. doi: 10.1212/01.wnl.0000073745.68744.94. [DOI] [PubMed] [Google Scholar]
- Gimenez-Amaya JM, Graybiel AM. Modular organization of projection neurons in the matrix compartment of the primate striatum. J Neurosci. 1991;11:779–791. doi: 10.1523/JNEUROSCI.11-03-00779.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giovanni A, Sieber BA, Heikkila RE, Sonsalla PK. Studies on species sensitivity to the dopaminergic neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Part 1: Systemic administration. J Pharmacol Exp Ther. 1994a;270:1000–1007. [PubMed] [Google Scholar]
- Giovanni A, Sonsalla PK, Heikkila RE. Studies on species sensitivity to the dopaminergic neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Part 2: Central administration of 1-methyl-4-phenylpyridinium. J Pharmacol Exp Ther. 1994b;270:1008–1014. [PubMed] [Google Scholar]
- Goldman-Rakic PS. The cortical dopamine system: role in memory and cognition. Adv Pharmacol. 1998;42:707–711. doi: 10.1016/s1054-3589(08)60846-7. [DOI] [PubMed] [Google Scholar]
- Goldman-Rakic PS, Brown RM. Regional changes of monoamines in cerebral cortex and subcortical structures of aging rhesus monkeys. Neuroscience. 1981;6:177–187. doi: 10.1016/0306-4522(81)90053-1. [DOI] [PubMed] [Google Scholar]
- Gonzales KK, Smith Y. Cholinergic interneurons in the dorsal and ventral striatum: anatomical and functional considerations in normal and diseased conditions. Ann N Y Acad Sci. 2015;1349:1–45. doi: 10.1111/nyas.12762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graybiel AM, Ragsdale CW., Jr Histochemically distinct compartments in the striatum of human, monkeys, and cat demonstrated by acetylthiocholinesterase staining. Proc Natl Acad Sci U S A. 1978;75:5723–5726. doi: 10.1073/pnas.75.11.5723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graybiel AM, Ragsdale CW, Jr, Yoneoka ES, Elde RP. An immunohistochemical study of enkephalins and other neuropeptides in the striatum of the cat with evidence that the opiate peptides are arranged to form mosaic patterns in register with the striosomal compartments visible by acetylcholinesterase staining. Neuroscience. 1981;6:377–397. doi: 10.1016/0306-4522(81)90131-7. [DOI] [PubMed] [Google Scholar]
- Grimbergen YA, Munneke M, Bloem BR. Falls in Parkinson’s disease. Curr Opin Neurol. 2004;17:405–415. doi: 10.1097/01.wco.0000137530.68867.93. [DOI] [PubMed] [Google Scholar]
- Gut NK, Winn P. The pedunculopontine tegmental nucleus-A functional hypothesis from the comparative literature. Mov Disord. 2016;31:615–624. doi: 10.1002/mds.26556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guttman M, et al. Brain serotonin transporter binding in non-depressed patients with Parkinson’s disease. Eur J Neurol. 2007;14:523–528. doi: 10.1111/j.1468-1331.2007.01727.x. [DOI] [PubMed] [Google Scholar]
- Guttman M, Fibiger HC, Jakubovic A, Calne DB. Intracarotid 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine administration: biochemical and behavioral observations in a primate model of hemiparkinsonism. J Neurochem. 1990;54:1329–1334. doi: 10.1111/j.1471-4159.1990.tb01966.x. [DOI] [PubMed] [Google Scholar]
- Hadipour-Niktarash A, Rommelfanger KS, Masilamoni GJ, Smith Y, Wichmann T. Extrastriatal D2-like receptors modulate basal ganglia pathways in normal and Parkinsonian monkeys. J Neurophysiol. 2012;107:1500–1512. doi: 10.1152/jn.00348.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halliday GM, Blumbergs PC, Cotton RG, Blessing WW, Geffen LB. Loss of brainstem serotonin- and substance P-containing neurons in Parkinson’s disease. Brain Res. 1990;510:104–107. doi: 10.1016/0006-8993(90)90733-r. [DOI] [PubMed] [Google Scholar]
- Hantraye P, et al. Dopamine fiber detection by [11C]-CFT and PET in a primate model of parkinsonism. Neuroreport. 1992;3:265–268. doi: 10.1097/00001756-199203000-00013. [DOI] [PubMed] [Google Scholar]
- Hantraye P, Varastet M, Peschanski M, Riche D, Cesaro P, Willer JC, Maziere M. Stable parkinsonian syndrome and uneven loss of striatal dopamine fibres following chronic MPTP administration in baboons. Neuroscience. 1993;53:169–178. doi: 10.1016/0306-4522(93)90295-q. [DOI] [PubMed] [Google Scholar]
- Hassani OK, Francois C, Yelnik J, Feger J. Evidence for a dopaminergic innervation of the subthalamic nucleus in the rat. Brain Res. 1997;749:88–94. doi: 10.1016/s0006-8993(96)01167-5. [DOI] [PubMed] [Google Scholar]
- Hawkes CH, Shephard BC, Daniel SE. Olfactory dysfunction in Parkinson’s disease. J Neurol Neurosurg Psychiatry. 1997;62:436–446. doi: 10.1136/jnnp.62.5.436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heckers S, Geula C, Mesulam MM. Cholinergic innervation of the human thalamus: dual origin and differential nuclear distribution. J Comp Neurol. 1992;325:68–82. doi: 10.1002/cne.903250107. [DOI] [PubMed] [Google Scholar]
- Hedreen JC. Tyrosine hydroxylase-immunoreactive elements in the human globus pallidus and subthalamic nucleus. J Comp Neurol. 1999;409:400–410. doi: 10.1002/(sici)1096-9861(19990705)409:3<400::aid-cne5>3.0.co;2-4. [DOI] [PubMed] [Google Scholar]
- Heikkila RE, Nicklas WJ, Vyas I, Duvoisin RC. Dopaminergic toxicity of rotenone and the 1-methyl-4-phenylpyridinium ion after their stereotaxic administration to rats: implication for the mechanism of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine toxicity. Neuroscience letters. 1985;62:389–394. doi: 10.1016/0304-3940(85)90580-4. [DOI] [PubMed] [Google Scholar]
- Henderson JM, Carpenter K, Cartwright H, Halliday GM. Degeneration of the centre median-parafascicular complex in Parkinson’s disease. Ann Neurol. 2000a;47:345–352. [PubMed] [Google Scholar]
- Henderson JM, Carpenter K, Cartwright H, Halliday GM. Loss of thalamic intralaminar nuclei in progressive supranuclear palsy and Parkinson’s disease: clinical and therapeutic implications. Brain. 2000b;123(Pt 7):1410–1421. doi: 10.1093/brain/123.7.1410. [DOI] [PubMed] [Google Scholar]
- Herkenham M, Little MD, Bankiewicz K, Yang SC, Markey SP, Johannessen JN. Selective retention of MPP+ within the monoaminergic systems of the primate brain following MPTP administration: an in vivo autoradiographic study. Neuroscience. 1991;40:133–158. doi: 10.1016/0306-4522(91)90180-v. [DOI] [PubMed] [Google Scholar]
- Herkenham M, Pert CB. Mosaic distribution of opiate receptors, parafascicular projections and acetylcholinesterase in rat striatum. Nature. 1981;291:415–418. doi: 10.1038/291415a0. [DOI] [PubMed] [Google Scholar]
- Herrero MT, Hirsch EC, Javoy-Agid F, Obeso JA, Agid Y. Differential vulnerability to 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine of dopaminergic and cholinergic neurons in the monkey mesopontine tegmentum. Brain Res. 1993a;624:281–285. doi: 10.1016/0006-8993(93)90088-5. [DOI] [PubMed] [Google Scholar]
- Herrero MT, et al. Does neuromelanin contribute to the vulnerability of catecholaminergic neurons in monkeys intoxicated with MPTP? Neuroscience. 1993b;56:499–511. doi: 10.1016/0306-4522(93)90349-k. [DOI] [PubMed] [Google Scholar]
- Hirsch EC, Graybiel AM, Duyckaerts C, Javoy-Agid F. Neuronal loss in the pedunculopontine tegmental nucleus in Parkinson disease and in progressive supranuclear palsy. Proc Natl Acad Sci U S A. 1987;84:5976–5980. doi: 10.1073/pnas.84.16.5976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoglinger GU, et al. Chronic systemic complex I inhibition induces a hypokinetic multisystem degeneration in rats. Journal of neurochemistry. 2003a;84:491–502. doi: 10.1046/j.1471-4159.2003.01533.x. [DOI] [PubMed] [Google Scholar]
- Hoglinger GU, et al. Chronic systemic complex I inhibition induces a hypokinetic multisystem degeneration in rats. J Neurochem. 2003b;84:491–502. doi: 10.1046/j.1471-4159.2003.01533.x. [DOI] [PubMed] [Google Scholar]
- Hornykiewicz O. Brain monoamines and parkinsonism. Natl Inst Drug Abuse Res Monogr Ser. 1975:13–21. doi: 10.1037/e472122004-001. [DOI] [PubMed] [Google Scholar]
- Hornykiewicz O. Biochemical aspects of Parkinson’s disease. Neurology. 1998;51:S2–9. doi: 10.1212/wnl.51.2_suppl_2.s2. [DOI] [PubMed] [Google Scholar]
- Hornykiewicz O, Kish SJ. Biochemical pathophysiology of Parkinson’s disease. Adv Neurol. 1987;45:19–34. [PubMed] [Google Scholar]
- Hosp JA, Pekanovic A, Rioult-Pedotti MS, Luft AR. Dopaminergic projections from midbrain to primary motor cortex mediate motor skill learning. J Neurosci. 2011;31:2481–2487. doi: 10.1523/JNEUROSCI.5411-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsia AY, Vincent JD, Lledo PM. Dopamine depresses synaptic inputs into the olfactory bulb. J Neurophysiol. 1999;82:1082–1085. doi: 10.1152/jn.1999.82.2.1082. [DOI] [PubMed] [Google Scholar]
- Huang C, et al. Changes in network activity with the progression of Parkinson’s disease. Brain. 2007;130:1834–1846. doi: 10.1093/brain/awm086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huisman E, Uylings HB, Hoogland PV. A 100% increase of dopaminergic cells in the olfactory bulb may explain hyposmia in Parkinson’s disease. Mov Disord. 2004;19:687–692. doi: 10.1002/mds.10713. [DOI] [PubMed] [Google Scholar]
- Iacopino A, Christakos S, German D, Sonsalla PK, Altar CA. Calbindin-D28K-containing neurons in animal models of neurodegeneration: possible protection from excitotoxicity. Brain Res Mol Brain Res. 1992a;13:251–261. doi: 10.1016/0169-328x(92)90033-8. [DOI] [PubMed] [Google Scholar]
- Iacopino AM, Christakos S, Modi P, Altar CA. Nerve growth factor increases calcium binding protein (calbindin-D28K) in rat olfactory bulb. Brain Res. 1992b;578:305–310. doi: 10.1016/0006-8993(92)90262-8. [DOI] [PubMed] [Google Scholar]
- Iravani MM, Syed E, Jackson MJ, Johnston LC, Smith LA, Jenner P. A modified MPTP treatment regime produces reproducible partial nigrostriatal lesions in common marmosets. The European journal of neuroscience. 2005;21:841–854. doi: 10.1111/j.1460-9568.2005.03915.x. [DOI] [PubMed] [Google Scholar]
- Irizarry MC, Growdon W, Gomez-Isla T, Newell K, George JM, Clayton DF, Hyman BT. Nigral and cortical Lewy bodies and dystrophic nigral neurites in Parkinson’s disease and cortical Lewy body disease contain alpha-synuclein immunoreactivity. J Neuropathol Exp Neurol. 1998;57:334–337. doi: 10.1097/00005072-199804000-00005. [DOI] [PubMed] [Google Scholar]
- Jackson-Lewis V, Blesa J, Przedborski S. Animal models of Parkinson’s disease Parkinsonism. Relat Disord. 2012;18(Suppl 1)(11):S183–185. 70057–8. doi: 10.1016/S1353-8020. [DOI] [PubMed] [Google Scholar]
- Jagmag SA, Tripathi N, Shukla SD, Maiti S, Khurana S. Evaluation of Models of Parkinson’s Disease. Front Neurosci. 2015;9:503. doi: 10.3389/fnins.2015.00503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jan C, Pessiglione M, Tremblay L, Tande D, Hirsch EC, Francois C. Quantitative analysis of dopaminergic loss in relation to functional territories in MPTP-treated monkeys. Eur J Neurosci. 2003;18:2082–2086. doi: 10.1046/j.1460-9568.2003.02946.x. [DOI] [PubMed] [Google Scholar]
- Jellinger K. The pedunculopontine nucleus in Parkinson’s disease, progressive supranuclear palsy and Alzheimer’s disease. J Neurol Neurosurg Psychiatry. 1988;51:540–543. doi: 10.1136/jnnp.51.4.540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jenner P. Pharmacology of dopamine agonists in the treatment of Parkinson’s disease. Neurology. 2002;58:S1–8. doi: 10.1212/wnl.58.suppl_1.s1. [DOI] [PubMed] [Google Scholar]
- Jenner P, Marsden CD. The actions of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine in animals as a model of Parkinson’s disease. J Neural Transm Suppl. 1986;20:11–39. [PubMed] [Google Scholar]
- Jimenez-Castellanos J, Graybiel AM. Subdivisions of the dopamine-containing A8-A9-A10 complex identified by their differential mesostriatal innervation of striosomes and extrastriosomal matrix. Neuroscience. 1987;23:223–242. doi: 10.1016/0306-4522(87)90285-5. [DOI] [PubMed] [Google Scholar]
- Joel D, Weiner I. The connections of the dopaminergic system with the striatum in rats and primates: an analysis with respect to the functional and compartmental organization of the striatum. Neuroscience. 2000;96:451–474. doi: 10.1016/s0306-4522(99)00575-8. [DOI] [PubMed] [Google Scholar]
- Johannessen JN. A model of chronic neurotoxicity: long-term retention of the neurotoxin 1-methyl-4-phenylpyridinium (MPP+) within catecholaminergic neurons. Neurotoxicology. 1991;12:285–302. [PubMed] [Google Scholar]
- Johannessen JN, Chiueh CC, Burns RS, Markey SP. Differences in the metabolism of MPTP in the rodent and primate parallel differences in sensitivity to its neurotoxic effects. Life Sci. 1985;36:219–224. doi: 10.1016/0024-3205(85)90062-1. [DOI] [PubMed] [Google Scholar]
- Johnson ME, Bobrovskaya L. An update on the rotenone models of Parkinson’s disease: their ability to reproduce the features of clinical disease and model gene-environment interactions. Neurotoxicology. 2015;46:101–116. doi: 10.1016/j.neuro.2014.12.002. [DOI] [PubMed] [Google Scholar]
- Johnston TM, Fox SH. Symptomatic Models of Parkinson’s Disease and L-DOPA-Induced Dyskinesia in Non-human Primates. Curr Top Behav Neurosci. 2015;22:221–235. doi: 10.1007/7854_2014_352. [DOI] [PubMed] [Google Scholar]
- Jouvet M. The role of monoamines and acetylcholine-containing neurons in the regulation of the sleep-waking cycle. Ergeb Physiol. 1972;64:166–307. doi: 10.1007/3-540-05462-6_2. [DOI] [PubMed] [Google Scholar]
- Kaasinen V, Nurmi E, Bruck A, Eskola O, Bergman J, Solin O, Rinne JO. Increased frontal [(18)F]fluorodopa uptake in early Parkinson’s disease: sex differences in the prefrontal cortex. Brain. 2001;124:1125–1130. doi: 10.1093/brain/124.6.1125. [DOI] [PubMed] [Google Scholar]
- Karachi C, et al. Cholinergic mesencephalic neurons are involved in gait and postural disorders in Parkinson disease. J Clin Invest. 2010;120:2745–2754. doi: 10.1172/JCI42642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kharkwal G, et al. Parkinsonism Driven by Antipsychotics Originates from Dopaminergic Control of Striatal Cholinergic Interneurons. Neuron. 2016;91:67–78. doi: 10.1016/j.neuron.2016.06.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kilbourn MR, Sherman P, Abbott LC. Reduced MPTP neurotoxicity in striatum of the mutant mouse tottering. Synapse. 1998;30:205–210. doi: 10.1002/(SICI)1098-2396(199810)30:2<205::AID-SYN10>3.0.CO;2-0. doi:10.1002/(SICI)1098-2396(199810)30:2<205::AID-SYN10>3.0.CO;2-0. [DOI] [PubMed] [Google Scholar]
- Kish SJ, Shannak K, Hornykiewicz O. Uneven pattern of dopamine loss in the striatum of patients with idiopathic Parkinson’s disease. Pathophysiologic and clinical implications. N Engl J Med. 1988;318:876–880. doi: 10.1056/NEJM198804073181402. [DOI] [PubMed] [Google Scholar]
- Kish SJ, Tong J, Hornykiewicz O, Rajput A, Chang LJ, Guttman M, Furukawa Y. Preferential loss of serotonin markers in caudate versus putamen in Parkinson’s disease. Brain. 2008;131:120–131. doi: 10.1093/brain/awm239. [DOI] [PubMed] [Google Scholar]
- Kliem MA, Maidment NT, Ackerson LC, Chen S, Smith Y, Wichmann T. Activation of nigral and pallidal dopamine D1-like receptors modulates basal ganglia outflow in monkeys. J Neurophysiol. 2007;98:1489–1500. doi: 10.1152/jn.00171.2007. [DOI] [PubMed] [Google Scholar]
- Kliem MA, Pare JF, Khan ZU, Wichmann T, Smith Y. Ultrastructural localization and function of dopamine D1-like receptors in the substantia nigra pars reticulata and the internal segment of the globus pallidus of parkinsonian monkeys. Eur J Neurosci. 2010;31:836–851. doi: 10.1111/j.1460-9568.2010.07109.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- JH Ko, et al. Prefrontal dopaminergic receptor abnormalities and executive functions in Parkinson’s disease. Hum Brain Mapp. 2013;34:1591–1604. doi: 10.1002/hbm.22006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ko WK, Camus SM, Li Q, Yang J, McGuire S, Pioli EY, Bezard E. An evaluation of istradefylline treatment on Parkinsonian motor and cognitive deficits in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-treated macaque models. Neuropharmacology. 2016;110:48–58. doi: 10.1016/j.neuropharm.2016.07.012. [DOI] [PubMed] [Google Scholar]
- Koprich JB, Johnston TH, Reyes G, Omana V, Brotchie JM. Towards a Non-Human Primate Model of Alpha-Synucleinopathy for Development of Therapeutics for Parkinson’s Disease: Optimization of AAV1/2 Delivery Parameters to Drive Sustained Expression of Alpha Synuclein and Dopaminergic Degeneration in Macaque. PLoS One. 2016;11:e0167235. doi: 10.1371/journal.pone.0167235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Korczyn AD. Dementia in Parkinson’s disease. J Neurol. 2001;248(Suppl 3):III1–4. doi: 10.1007/pl00022916. [DOI] [PubMed] [Google Scholar]
- Koster NL, Norman AB, Richtand NM, Nickell WT, Puche AC, Pixley SK, Shipley MT. Olfactory receptor neurons express D2 dopamine receptors. J Comp Neurol. 1999;411:666–673. doi: 10.1002/(sici)1096-9861(19990906)411:4<666::aid-cne10>3.0.co;2-s. [DOI] [PubMed] [Google Scholar]
- Kostic VS, Djuricic BM, Covickovic-Sternic N, Bumbasirevic L, Nikolic M, Mrsulja BB. Depression and Parkinson’s disease: possible role of serotonergic mechanisms. J Neurol. 1987;234:94–96. doi: 10.1007/BF00314109. [DOI] [PubMed] [Google Scholar]
- Kumakura Y, et al. Elevated [(18)F]FDOPA utilization in the periaqueductal gray and medial nucleus accumbens of patients with early Parkinson’s disease. Neuroimage. 2010;49:2933–2939. doi: 10.1016/j.neuroimage.2009.11.035. [DOI] [PubMed] [Google Scholar]
- Lange KW, Wells FR, Jenner P, Marsden CD. Altered muscarinic and nicotinic receptor densities in cortical and subcortical brain regions in Parkinson’s disease. J Neurochem. 1993;60:197–203. doi: 10.1111/j.1471-4159.1993.tb05838.x. [DOI] [PubMed] [Google Scholar]
- Langer LF, Graybiel AM. Distinct nigrostriatal projection systems innervate striosomes and matrix in the primate striatum. Brain Res. 1989;498:344–350. doi: 10.1016/0006-8993(89)91114-1. [DOI] [PubMed] [Google Scholar]
- Langston JW. The Parkinson’s complex: parkinsonism is just the tip of the iceberg. Annals of neurology. 2006;59:591–596. doi: 10.1002/ana.20834. [DOI] [PubMed] [Google Scholar]
- Langston JW, Ballard P, Tetrud JW, Irwin I. Chronic Parkinsonism in humans due to a product of meperidine-analog synthesis. Science. 1983;219:979–980. doi: 10.1126/science.6823561. [DOI] [PubMed] [Google Scholar]
- Langston JW, Ballard PA., Jr Parkinson’s disease in a chemist working with 1-methyl-4-phenyl-1,2,5,6-tetrahydropyridine. N Engl J Med. 1983;309:310. doi: 10.1056/nejm198308043090511. [DOI] [PubMed] [Google Scholar]
- Langston JW, Forno LS, Tetrud J, Reeves AG, Kaplan JA, Karluk D. Evidence of active nerve cell degeneration in the substantia nigra of humans years after 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine exposure. Ann Neurol. 1999;46:598–605. doi: 10.1002/1531-8249(199910)46:4<598::aid-ana7>3.0.co;2-f. [DOI] [PubMed] [Google Scholar]
- Lavoie B, Parent A. Dopaminergic neurons expressing calbindin in normal and parkinsonian monkeys. Neuroreport. 1991;2:601–604. doi: 10.1097/00001756-199110000-00012. [DOI] [PubMed] [Google Scholar]
- Lavoie B, Smith Y, Parent A. Dopaminergic innervation of the basal ganglia in the squirrel monkey as revealed by tyrosine hydroxylase immunohistochemistry. J Comp Neurol. 1989;289:36–52. doi: 10.1002/cne.902890104. [DOI] [PubMed] [Google Scholar]
- Leblois A, Boraud T, Meissner W, Bergman H, Hansel D. Competition between feedback loops underlies normal and pathological dynamics in the basal ganglia. J Neurosci. 2006;26:3567–3583. doi: 10.1523/JNEUROSCI.5050-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee JY, et al. Extrastriatal dopaminergic changes in Parkinson’s disease patients with impulse control disorders. J Neurol Neurosurg Psychiatry. 2014;85:23–30. doi: 10.1136/jnnp-2013-305549. [DOI] [PubMed] [Google Scholar]
- Lees AJ, Smith E. Cognitive deficits in the early stages of Parkinson’s disease. Brain. 1983;106(Pt 2):257–270. doi: 10.1093/brain/106.2.257. [DOI] [PubMed] [Google Scholar]
- Lemke MR. Depressive symptoms in Parkinson’s disease. Eur J Neurol. 2008;15(Suppl 1):21–25. doi: 10.1111/j.1468-1331.2008.02058.x. [DOI] [PubMed] [Google Scholar]
- Lenka A, Hegde S, Jhunjhunwala KR, Pal PK. Interactions of visual hallucinations, rapid eye movement sleep behavior disorder and cognitive impairment in Parkinson’s disease: A review Parkinsonism. Relat Disord. 2016;22:1–8. doi: 10.1016/j.parkreldis.2015.11.018. [DOI] [PubMed] [Google Scholar]
- Lewis DA, Campbell MJ, Foote SL, Goldstein M, Morrison JH. The distribution of tyrosine hydroxylase-immunoreactive fibers in primate neocortex is widespread but regionally specific. J Neurosci. 1987;7:279–290. doi: 10.1523/JNEUROSCI.07-01-00279.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis DA, Melchitzky DS, Haycock JW. Expression and distribution of two isoforms of tyrosine hydroxylase in macaque monkey brain. Brain Res. 1994;656:1–13. doi: 10.1016/0006-8993(94)91360-9. [DOI] [PubMed] [Google Scholar]
- Lewis DA, Melchitzky DS, Sesack SR, Whitehead RE, Auh S, Sampson A. Dopamine transporter immunoreactivity in monkey cerebral cortex: regional, laminar, and ultrastructural localization. J Comp Neurol. 2001;432:119–136. doi: 10.1002/cne.1092. [DOI] [PubMed] [Google Scholar]
- Liang CL, Sinton CM, German DC. Midbrain dopaminergic neurons in the mouse: co-localization with Calbindin-D28K and calretinin. Neuroscience. 1996a;75:523–533. doi: 10.1016/0306-4522(96)00228-x. [DOI] [PubMed] [Google Scholar]
- Liang CL, Sinton CM, Sonsalla PK, German DC. Midbrain dopaminergic neurons in the mouse that contain calbindin-D28k exhibit reduced vulnerability to MPTP-induced neurodegeneration. Neurodegeneration. 1996b;5:313–318. doi: 10.1006/neur.1996.0042. [DOI] [PubMed] [Google Scholar]
- Lin X, et al. Proteomic profiling in MPTP monkey model for early Parkinson disease biomarker discovery. Biochim Biophys Acta. 2015;1854:779–787. doi: 10.1016/j.bbapap.2015.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindgren HS, Dunnett SB. Cognitive dysfunction and depression in Parkinson’s disease: what can be learned from rodent models? Eur J Neurosci. 2012;35:1894–1907. doi: 10.1111/j.1460-9568.2012.08162.x. [DOI] [PubMed] [Google Scholar]
- Low K, Aebischer P. Use of viral vectors to create animal models for Parkinson’s disease. Neurobiol Dis. 2012;48:189–201. doi: 10.1016/j.nbd.2011.12.038. [DOI] [PubMed] [Google Scholar]
- Lu L. Regional vulnerability of mesencephalic dopaminergic neurons prone to degenerate in Parkinson’s disease: a post-mortem study in human control subjects. Neurobiol Dis. 2006;23:409–421. doi: 10.1016/j.nbd.2006.04.002. [DOI] [PubMed] [Google Scholar]
- Luft AR, Schwarz S. Dopaminergic signals in primary motor cortex. Int J Dev Neurosci. 2009;27:415–421. doi: 10.1016/j.ijdevneu.2009.05.004. doi:S0736-5748(09)00067-7[pii] 10.1016/j.ijdevneu.2009.05.004. [DOI] [PubMed] [Google Scholar]
- Marmion DJ, Kordower JH. alpha-Synuclein nonhuman primate models of Parkinson’s disease. J Neural Transm (Vienna) 2017 doi: 10.1007/s00702-017-1720-0. [DOI] [PubMed] [Google Scholar]
- Masilamoni G, Smith Y. Chronic MPTP treatment induces extrastriatal monoaminergic denervation and α-synuclein aggregates in a rhesus monkey model of Parkinson’s disease. Society of Neuroscience. 2011 [Google Scholar]
- Masilamoni G, et al. (18)F-FECNT: validation as PET dopamine transporter ligand in parkinsonism. Experimental neurology. 2010;226:265–273. doi: 10.1016/j.expneurol.2010.08.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masilamoni G, Weinkle A, Bogenpohl J, Groover O, Wichmann T, Y S. A nonhuman primate model of Parkinson’s disease associated with cortical and subcortical dopaminergic, noradrenergic and serotonergic neuronal degeneration. Mov Disord. 2011a;26(Suppl. 2):S23–S24. abstract. [Google Scholar]
- Masilamoni GJ, et al. Metabotropic glutamate receptor 5 antagonist protects dopaminergic and noradrenergic neurons from degeneration in MPTP-treated monkeys. Brain. 2011b;134:2057–2073. doi: 10.1093/brain/awr137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masilamoni GJ, Groover O, Smith Y. Reduced noradrenergic innervation of ventral midbrain dopaminergic cell groups and the subthalamic nucleus in MPTP-treated parkinsonian monkeys. Neurobiol Dis. 2016;100:9–18. doi: 10.1016/j.nbd.2016.12.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCallum SE, Parameswaran N, Perez XA, Bao S, McIntosh JM, Grady SR, Quik M. Compensation in pre-synaptic dopaminergic function following nigrostriatal damage in primates. J Neurochem. 2006;96:960–972. doi: 10.1111/j.1471-4159.2005.03610.x. [DOI] [PubMed] [Google Scholar]
- McCance-Katz EF, Marek KL, Price LH. Serotonergic dysfunction in depression associated with Parkinson’s disease. Neurology. 1992;42:1813–1814. doi: 10.1212/wnl.42.9.1813. [DOI] [PubMed] [Google Scholar]
- McCormack AL, Mak SK, Shenasa M, Langston WJ, Forno LS, Di Monte DA. Pathologic modifications of alpha-synuclein in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-treated squirrel monkeys. J Neuropathol Exp Neurol. 2008;67:793–802. doi: 10.1097/NEN.0b013e318180f0bd. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McNaught KS, Perl DP, Brownell AL, Olanow CW. Systemic exposure to proteasome inhibitors causes a progressive model of Parkinson’s disease. Ann Neurol. 2004;56:149–162. doi: 10.1002/ana.20186. [DOI] [PubMed] [Google Scholar]
- Meissner W, Prunier C, Guilloteau D, Chalon S, Gross CE, Bezard E. Time-course of nigrostriatal degeneration in a progressive MPTP-lesioned macaque model of Parkinson’s disease. Mol Neurobiol. 2003;28:209–218. doi: 10.1385/MN:28:3:209. [DOI] [PubMed] [Google Scholar]
- Mejias-Aponte CA, Drouin C, Aston-Jones G. Adrenergic and noradrenergic innervation of the midbrain ventral tegmental area and retrorubral field: prominent inputs from medullary homeostatic centers. J Neurosci. 2009;29:3613–3626. doi: 10.1523/JNEUROSCI.4632-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menegas W, et al. Dopamine neurons projecting to the posterior striatum form an anatomically distinct subclass. Elife. 2015;4:e10032. doi: 10.7554/eLife.10032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mesholam RI, Moberg PJ, Mahr RN, Doty RL. Olfaction in neurodegenerative disease: a meta-analysis of olfactory functioning in Alzheimer’s and Parkinson’s diseases. Arch Neurol. 1998;55:84–90. doi: 10.1001/archneur.55.1.84. [DOI] [PubMed] [Google Scholar]
- Mihatsch W, Russ H, Gerlach M, Riederer P, Przuntek H. Treatment with antioxidants does not prevent loss of dopamine in the striatum of MPTP-treated common marmosets: preliminary observations. J Neural Transm Park Dis Dement Sect. 1991;3:73–78. doi: 10.1007/BF02251138. [DOI] [PubMed] [Google Scholar]
- Miller GW, Erickson JD, Perez JT, Penland SN, Mash DC, Rye DB, Levey AI. Immunochemical analysis of vesicular monoamine transporter (VMAT2) protein in Parkinson’s disease. Exp Neurol. 1999a;156:138–148. doi: 10.1006/exnr.1998.7008. [DOI] [PubMed] [Google Scholar]
- Miller GW, Gainetdinov RR, Levey AI, Caron MG. Dopamine transporters and neuronal injury. Trends Pharmacol Sci. 1999b;20:424–429. doi: 10.1016/s0165-6147(99)01379-6. [DOI] [PubMed] [Google Scholar]
- Miyasaki JM, et al. Practice Parameter: evaluation and treatment of depression, psychosis, and dementia in Parkinson disease (an evidence-based review): report of the Quality Standards Subcommittee of the American Academy of Neurology. Neurology. 2006;66:996–1002. doi: 10.1212/01.wnl.0000215428.46057.3d. [DOI] [PubMed] [Google Scholar]
- Moore RY, Whone AL, Brooks DJ. Extrastriatal monoamine neuron function in Parkinson’s disease: an 18F-dopa PET study. Neurobiol Dis. 2008;29:381–390. doi: 10.1016/j.nbd.2007.09.004. [DOI] [PubMed] [Google Scholar]
- Moratalla R, Quinn B, DeLanney LE, Irwin I, Langston JW, Graybiel AM. Differential vulnerability of primate caudate-putamen and striosome-matrix dopamine systems to the neurotoxic effects of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Proceedings of the National Academy of Sciences of the United States of America. 1992;89:3859–3863. doi: 10.1073/pnas.89.9.3859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morin N, Jourdain VA, Di Paolo T. Modeling dyskinesia in animal models of Parkinson disease. Exp Neurol. 2014;256:105–116. doi: 10.1016/j.expneurol.2013.01.024. [DOI] [PubMed] [Google Scholar]
- Morissette M, Di Paolo T. Non-human primate models of PD to test novel therapies. J Neural Transm (Vienna) 2017 doi: 10.1007/s00702-017-1722-y. [DOI] [PubMed] [Google Scholar]
- Mounayar S, et al. A new model to study compensatory mechanisms in MPTP-treated monkeys exhibiting recovery. Brain: a journal of neurology. 2007;130:2898–2914. doi: 10.1093/brain/awm208. [DOI] [PubMed] [Google Scholar]
- Mundinano IC, et al. Increased dopaminergic cells and protein aggregates in the olfactory bulb of patients with neurodegenerative disorders. Acta Neuropathol. 2011;122:61–74. doi: 10.1007/s00401-011-0830-2. [DOI] [PubMed] [Google Scholar]
- Mundinano IC, Hernandez M, Dicaudo C, Ordonez C, Marcilla I, Tunon MT, Luquin MR. Reduced cholinergic olfactory centrifugal inputs in patients with neurodegenerative disorders and MPTP-treated monkeys. Acta Neuropathol. 2013;126:411–425. doi: 10.1007/s00401-013-1144-3. [DOI] [PubMed] [Google Scholar]
- Munro-Davies LE, Winter J, Aziz TZ, Stein JF. The role of the pedunculopontine region in basal-ganglia mechanisms of akinesia. Exp Brain Res. 1999;129:511–517. doi: 10.1007/s002210050921. [DOI] [PubMed] [Google Scholar]
- Nakano I, Hirano A. Parkinson’s disease: neuron loss in the nucleus basalis without concomitant Alzheimer’s disease. Ann Neurol. 1984;15:415–418. doi: 10.1002/ana.410150503. [DOI] [PubMed] [Google Scholar]
- Nandi D, Aziz TZ, Giladi N, Winter J, Stein JF. Reversal of akinesia in experimental parkinsonism by GABA antagonist microinjections in the pedunculopontine nucleus. Brain. 2002;125:2418–2430. doi: 10.1093/brain/awf259. [DOI] [PubMed] [Google Scholar]
- Naneix F, Marchand AR, Di Scala G, Pape JR, Coutureau E. A role for medial prefrontal dopaminergic innervation in instrumental conditioning. J Neurosci. 2009;29:6599–6606. doi: 10.1523/JNEUROSCI.1234-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Narayanan NS, Rodnitzky RL, Uc EY. Prefrontal dopamine signaling and cognitive symptoms of Parkinson’s disease. Rev Neurosci. 2013;24:267–278. doi: 10.1515/revneuro-2013-0004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nayyar T, et al. Cortical serotonin and norepinephrine denervation in parkinsonism: preferential loss of the beaded serotonin innervation. Eur J Neurosci. 2009;30:207–216. doi: 10.1111/j.1460-9568.2009.06806.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niu Y, et al. Early Parkinson’s disease symptoms in alpha-synuclein transgenic monkeys. Hum Mol Genet. 2015;24:2308–2317. doi: 10.1093/hmg/ddu748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nobin A, Bjorklund A. Topography of the monoamine neuron systems in the human brain as revealed in fetuses. Acta Physiol Scand Suppl. 1973;388:1–40. [PubMed] [Google Scholar]
- Ohama E, Ikuta F. Parkinson’s disease: distribution of Lewy bodies and monoamine neuron system. Acta Neuropathol. 1976;34:311–319. doi: 10.1007/BF00696560. [DOI] [PubMed] [Google Scholar]
- Oke AF, Adams RN. Elevated thalamic dopamine: possible link to sensory dysfunctions in schizophrenia. Schizophr Bull. 1987;13:589–604. doi: 10.1093/schbul/13.4.589. [DOI] [PubMed] [Google Scholar]
- Oliveras-Salva M, et al. rAAV2/7 vector-mediated overexpression of alpha-synuclein in mouse substantia nigra induces protein aggregation and progressive dose-dependent neurodegeneration. Mol Neurodegener. 2013;8:44. doi: 10.1186/1750-1326-8-44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papadopoulos GC, Parnavelas JG. Distribution and synaptic organization of dopaminergic axons in the lateral geniculate nucleus of the rat. J Comp Neurol. 1990;294:356–361. doi: 10.1002/cne.902940305. [DOI] [PubMed] [Google Scholar]
- Parent A, Cossette M. Extrastriatal dopamine and Parkinson’s disease. Adv Neurol. 2001;86:45–54. [PubMed] [Google Scholar]
- Parent A, Smith Y. Differential dopaminergic innervation of the two pallidal segments in the squirrel monkey (Saimiri sciureus) Brain Res. 1987;426:397–400. doi: 10.1016/0006-8993(87)90896-1. [DOI] [PubMed] [Google Scholar]
- Parent A, Smith Y, Filion M, Dumas J. Distinct afferents to internal and external pallidal segments in the squirrel monkey. Neurosci Lett. 1989;96:140–144. doi: 10.1016/0304-3940(89)90047-5. [DOI] [PubMed] [Google Scholar]
- Parry TJ, Eberle-Wang K, Lucki I, Chesselet MF. Dopaminergic stimulation of subthalamic nucleus elicits oral dyskinesia in rats. Exp Neurol. 1994;128:181–190. doi: 10.1006/exnr.1994.1126. [DOI] [PubMed] [Google Scholar]
- Patt S, Gerhard L. A Golgi study of human locus coeruleus in normal brains and in Parkinson’s disease. Neuropathol Appl Neurobiol. 1993;19:519–523. doi: 10.1111/j.1365-2990.1993.tb00480.x. [DOI] [PubMed] [Google Scholar]
- Pavese N, et al. In vivo assessment of brain monoamine systems in parkin gene carriers: a PET study. Exp Neurol. 2010;222:120–124. doi: 10.1016/j.expneurol.2009.12.021. [DOI] [PubMed] [Google Scholar]
- Pavese N, Rivero-Bosch M, Lewis SJ, Whone AL, Brooks DJ. Progression of monoaminergic dysfunction in Parkinson’s disease: a longitudinal 18F-dopa PET study. Neuroimage. 2011;56:1463–1468. doi: 10.1016/j.neuroimage.2011.03.012. [DOI] [PubMed] [Google Scholar]
- Pearce RK, Jackson M, Smith L, Jenner P, Marsden CD. Chronic L-DOPA administration induces dyskinesias in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-treated common marmoset (Callithrix Jacchus) Mov Disord. 1995;10:731–740. doi: 10.1002/mds.870100606. [DOI] [PubMed] [Google Scholar]
- Perez-Otano I, Herrero MT, Oset C, De Ceballos ML, Luquin MR, Obeso JA, Del Rio J. Extensive loss of brain dopamine and serotonin induced by chronic administration of MPTP in the marmoset. Brain Res. 1991;567:127–132. doi: 10.1016/0006-8993(91)91444-6. [DOI] [PubMed] [Google Scholar]
- Perez-Otano I, Oset C, Luquin MR, Herrero MT, Obeso JA, Del Rio J. MPTP-induced parkinsonism in primates: pattern of striatal dopamine loss following acute and chronic administration. Neurosci Lett. 1994a;175:121–125. doi: 10.1016/0304-3940(94)91094-4. [DOI] [PubMed] [Google Scholar]
- Perez-Otano I, Oset C, Luquin MR, Herrero MT, Obeso JA, Del Rio J. MPTP-induced parkinsonism in primates: pattern of striatal dopamine loss following acute and chronic administration. Neuroscience letters. 1994b;175:121–125. doi: 10.1016/0304-3940(94)91094-4. [DOI] [PubMed] [Google Scholar]
- Perry EK, et al. Cholinergic correlates of cognitive impairment in Parkinson’s disease: comparisons with Alzheimer’s disease. J Neurol Neurosurg Psychiatry. 1985;48:413–421. doi: 10.1136/jnnp.48.5.413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pessiglione M, Guehl D, Jan C, Francois C, Hirsch EC, Feger J, Tremblay L. Disruption of self-organized actions in monkeys with progressive MPTP-induced parkinsonism: II. Effects of reward preference. Eur J Neurosci. 2004;19:437–446. doi: 10.1111/j.0953-816x.2003.03089.x. [DOI] [PubMed] [Google Scholar]
- Petrides M, Pandya DN. Dorsolateral prefrontal cortex: comparative cytoarchitectonic analysis in the human and the macaque brain and corticocortical connection patterns. Eur J Neurosci. 1999;11:1011–1036. doi: 10.1046/j.1460-9568.1999.00518.x. [DOI] [PubMed] [Google Scholar]
- Petryszyn S, Di Paolo T, Parent A, Parent M. The number of striatal cholinergic interneurons expressing calretinin is increased in parkinsonian monkeys. Neurobiol Dis. 2016;95:46–53. doi: 10.1016/j.nbd.2016.07.002. [DOI] [PubMed] [Google Scholar]
- Pifl C, Bertel O, Schingnitz G, Hornykiewicz O. Extrastriatal dopamine in symptomatic and asymptomatic rhesus monkeys treated with 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) Neurochem Int. 1990;17:263–270. doi: 10.1016/0197-0186(90)90148-m. [DOI] [PubMed] [Google Scholar]
- Pifl C, Hornykiewicz O, Giros B, Caron MG. Catecholamine transporters and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine neurotoxicity: studies comparing the cloned human noradrenaline and human dopamine transporter. J Pharmacol Exp Ther. 1996;277:1437–1443. [PubMed] [Google Scholar]
- Pifl C, Reither H, Del Rey NL, Cavada C, Obeso JA, Blesa J. Early Paradoxical Increase of Dopamine: A Neurochemical Study of Olfactory Bulb in Asymptomatic and Symptomatic MPTP Treated Monkeys. Front Neuroanat. 2017;11:46. doi: 10.3389/fnana.2017.00046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pifl C, Schingnitz G, Hornykiewicz O. The neurotoxin MPTP does not reproduce in the rhesus monkey the interregional pattern of striatal dopamine loss typical of human idiopathic Parkinson’s disease. Neurosci Lett. 1988a;92:228–233. doi: 10.1016/0304-3940(88)90066-3. [DOI] [PubMed] [Google Scholar]
- Pifl C, Schingnitz G, Hornykiewicz O. The neurotoxin MPTP does not reproduce in the rhesus monkey the interregional pattern of striatal dopamine loss typical of human idiopathic Parkinson’s disease. Neuroscience letters. 1988b;92:228–233. doi: 10.1016/0304-3940(88)90066-3. [DOI] [PubMed] [Google Scholar]
- Pifl C, Schingnitz G, Hornykiewicz O. Effect of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine on the regional distribution of brain monoamines in the rhesus monkey. Neuroscience. 1991;44:591–605. doi: 10.1016/0306-4522(91)90080-8. [DOI] [PubMed] [Google Scholar]
- Pillon B, Dubois B, Cusimano G, Bonnet AM, Lhermitte F, Agid Y. Does cognitive impairment in Parkinson’s disease result from non-dopaminergic lesions? J Neurol Neurosurg Psychiatry. 1989;52:201–206. doi: 10.1136/jnnp.52.2.201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Politis M, Wu K, Loane C, Kiferle L, Molloy S, Brooks DJ, Piccini P. Staging of serotonergic dysfunction in Parkinson’s disease: an in vivo 11C-DASB PET study. Neurobiol Dis. 2010;40:216–221. doi: 10.1016/j.nbd.2010.05.028. [DOI] [PubMed] [Google Scholar]
- Porras G, Li Q, Bezard E. Modeling Parkinson’s disease in primates: The MPTP model. Cold Spring Harb Perspect Med. 2012;2:a009308. doi: 10.1101/cshperspect.a009308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Potts LF, Wu H, Singh A, Marcilla I, Luquin MR, Papa SM. Modeling Parkinson’s disease in monkeys for translational studies, a critical analysis. Exp Neurol. 2014;256:133–143. doi: 10.1016/j.expneurol.2013.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prensa L, Cossette M, Parent A. Dopaminergic innervation of human basal ganglia. J Chem Neuroanat. 2000;20:207–213. doi: 10.1016/s0891-0618(00)00099-5. [DOI] [PubMed] [Google Scholar]
- Preuss TM. Do rats have prefrontal cortex? The rose-woolsey-akert program reconsidered. J Cogn Neurosci. 1995;7:1–24. doi: 10.1162/jocn.1995.7.1.1. [DOI] [PubMed] [Google Scholar]
- Rajput AH, Sitte HH, Rajput A, Fenton ME, Pifl C, Hornykiewicz O. Globus pallidus dopamine and Parkinson motor subtypes: clinical and brain biochemical correlation. Neurology. 2008;70:1403–1410. doi: 10.1212/01.wnl.0000285082.18969.3a. [DOI] [PubMed] [Google Scholar]
- Rakshi JS, et al. Frontal, midbrain and striatal dopaminergic function in early and advanced Parkinson’s disease A 3D [(18)F]dopa-PET study. Brain. 1999;122(Pt 9):1637–1650. doi: 10.1093/brain/122.9.1637. [DOI] [PubMed] [Google Scholar]
- Rappold PM, Tieu K. Astrocytes and therapeutics for Parkinson’s disease. Neurotherapeutics. 2010;7:413–423. doi: 10.1016/j.nurt.2010.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Recasens A, et al. Lewy body extracts from Parkinson disease brains trigger alpha-synuclein pathology and neurodegeneration in mice and monkeys. Ann Neurol. 2014;75:351–362. doi: 10.1002/ana.24066. [DOI] [PubMed] [Google Scholar]
- Reichmann H, Brandt MD, Klingelhoefer L. The nonmotor features of Parkinson’s disease: pathophysiology and management advances. Curr Opin Neurol. 2016;29:467–473. doi: 10.1097/WCO.0000000000000348. [DOI] [PubMed] [Google Scholar]
- Rey NL, Wesson DW, Brundin P. The olfactory bulb as the entry site for prion-like propagation in neurodegenerative diseases. Neurobiol Dis. 2016 doi: 10.1016/j.nbd.2016.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roeltgen DP, Schneider JS. Chronic low-dose MPTP in nonhuman primates: a possible model for attention deficit disorder. J Child Neurol. 1991;6(Suppl):S82–89. doi: 10.1177/0883073891006001s10. [DOI] [PubMed] [Google Scholar]
- Roeltgen DP, Schneider JS. Task persistence and learning ability in normal and chronic low dose MPTP-treated monkeys. Behav Brain Res. 1994;60:115–124. doi: 10.1016/0166-4328(94)90138-4. [DOI] [PubMed] [Google Scholar]
- Rogers JD, Brogan D, Mirra SS. The nucleus basalis of Meynert in neurological disease: a quantitative morphological study. Ann Neurol. 1985;17:163–170. doi: 10.1002/ana.410170210. [DOI] [PubMed] [Google Scholar]
- Rommelfanger KS, Weinshenker D, Miller GW. Reduced MPTP toxicity in noradrenaline transporter knockout mice. J Neurochem. 2004;91:1116–1124. doi: 10.1111/j.1471-4159.2004.02785.x. [DOI] [PubMed] [Google Scholar]
- Rommelfanger KS, Wichmann T. Extrastriatal dopaminergic circuits of the Basal Ganglia. Front Neuroanat. 2010;4:139. doi: 10.3389/fnana.2010.00139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russ H, Mihatsch W, Gerlach M, Riederer P, Przuntek H. Neurochemical and behavioural features induced by chronic low dose treatment with 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) in the common marmoset: implications for Parkinson’s disease? Neurosci Lett. 1991;123:115–118. doi: 10.1016/0304-3940(91)90171-o. [DOI] [PubMed] [Google Scholar]
- Sanchez-Gonzalez MA, Garcia-Cabezas MA, Rico B, Cavada C. The primate thalamus is a key target for brain dopamine. J Neurosci. 2005;25:6076–6083. doi: 10.1523/JNEUROSCI.0968-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanghera MK, Manaye K, McMahon A, Sonsalla PK, German DC. Dopamine transporter mRNA levels are high in midbrain neurons vulnerable to MPTP. Neuroreport. 1997;8:3327–3331. doi: 10.1097/00001756-199710200-00027. [DOI] [PubMed] [Google Scholar]
- Scatton B, Javoy-Agid F, Rouquier L, Dubois B, Agid Y. Reduction of cortical dopamine, noradrenaline, serotonin and their metabolites in Parkinson’s disease. Brain Res. 1983;275:321–328. doi: 10.1016/0006-8993(83)90993-9. [DOI] [PubMed] [Google Scholar]
- Schneider JS. Modeling Cognitive Deficits Associated with Parkinsonism in the Chronic-Low-Dose MPTP-Treated Monkey. In: Levin ED, Buccafusco JJ, editors. Animal Models of Cognitive Impairment Frontiers in Neuroscience. Boca Raton (FL): 2006. [PubMed] [Google Scholar]
- Schneider JS, Kovelowski CJ., 2nd Chronic exposure to low doses of MPTP. I. Cognitive deficits in motor asymptomatic monkeys. Brain Res. 1990;519:122–128. doi: 10.1016/0006-8993(90)90069-n. [DOI] [PubMed] [Google Scholar]
- Schneider JS, Tinker JP, Van Velson M, Menzaghi F, Lloyd GK. Nicotinic acetylcholine receptor agonist SIB-1508Y improves cognitive functioning in chronic low-dose MPTP-treated monkeys. J Pharmacol Exp Ther. 1999;290:731–739. [PubMed] [Google Scholar]
- Sesack SR, Bressler CN, Lewis DA. Ultrastructural associations between dopamine terminals and local circuit neurons in the monkey prefrontal cortex: a study of calretinin-immunoreactive cells. Neurosci Lett. 1995;200:9–12. doi: 10.1016/0304-3940(95)12076-g. [DOI] [PubMed] [Google Scholar]
- Shimozawa A, et al. Propagation of pathological alpha-synuclein in marmoset brain. Acta Neuropathol Commun. 2017;5:12. doi: 10.1186/s40478-017-0413-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slovin H, Abeles M, Vaadia E, Haalman I, Prut Y, Bergman H. Frontal cognitive impairments and saccadic deficits in low-dose MPTP-treated monkeys. J Neurophysiol. 1999;81:858–874. doi: 10.1152/jn.1999.81.2.858. [DOI] [PubMed] [Google Scholar]
- Smith Y, et al. The thalamostriatal system in normal and diseased states. Front Syst Neurosci. 2014;8:5. doi: 10.3389/fnsys.2014.00005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith Y, Kieval JZ. Anatomy of the dopamine system in the basal ganglia. Trends Neurosci. 2000;23:S28–33. doi: 10.1016/s1471-1931(00)00023-9. [DOI] [PubMed] [Google Scholar]
- Smith Y, Raju D, Nanda B, Pare JF, Galvan A, Wichmann T. The thalamostriatal systems: anatomical and functional organization in normal and parkinsonian states. Brain Res Bull. 2009;78:60–68. doi: 10.1016/j.brainresbull.2008.08.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith Y, Raju DV, Pare JF, Sidibe M. The thalamostriatal system: a highly specific network of the basal ganglia circuitry. Trends Neurosci. 2004;27:520–527. doi: 10.1016/j.tins.2004.07.004. [DOI] [PubMed] [Google Scholar]
- Smith Y, Surmeier DJ, Redgrave P, Kimura M. Thalamic contributions to Basal Ganglia-related behavioral switching and reinforcement. J Neurosci. 2011;31:16102–16106. doi: 10.1523/JNEUROSCI.4634-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith Y, Villalba R. Striatal and extrastriatal dopamine in the basal ganglia: an overview of its anatomical organization in normal and Parkinsonian brains. Mov Disord. 2008;23(Suppl 3):S534–547. doi: 10.1002/mds.22027. [DOI] [PubMed] [Google Scholar]
- Solari N, Bonito-Oliva A, Fisone G, Brambilla R. Understanding cognitive deficits in Parkinson’s disease: lessons from preclinical animal models. Learn Mem. 2013;20:592–600. doi: 10.1101/lm.032029.113. [DOI] [PubMed] [Google Scholar]
- Spillantini MG, Schmidt ML, Lee VM, Trojanowski JQ, Jakes R, Goedert M. Alpha-synuclein in Lewy bodies. Nature. 1997;388:839–840. doi: 10.1038/42166. [DOI] [PubMed] [Google Scholar]
- Steriade M. Sleep, epilepsy and thalamic reticular inhibitory neurons. Trends Neurosci. 2005;28:317–324. doi: 10.1016/j.tins.2005.03.007. [DOI] [PubMed] [Google Scholar]
- Sterpenich V, et al. The locus ceruleus is involved in the successful retrieval of emotional memories in humans. J Neurosci. 2006;26:7416–7423. doi: 10.1523/JNEUROSCI.1001-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Surmeier DJ, Obeso JA, Halliday GM. Selective neuronal vulnerability in Parkinson disease. Nat Rev Neurosci. 2017;18:101–113. doi: 10.1038/nrn.2016.178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sutoo D, Akiyama K, Yabe K. Quantitative imaging of tyrosine hydroxylase and calmodulin in the human brain. J Neurosci Res. 2001;63:369–376. doi: 10.1002/1097-4547(20010301)63:5<369::AID-JNR1031>3.0.CO;2-3. doi: 10.1002/1097-4547(20010301)63:5<369::AID-JNR1031>3.0.CO;2-3. [DOI] [PubMed] [Google Scholar]
- Tagliavini F, Pilleri G, Bouras C, Constantinidis J. The basal nucleus of Meynert in idiopathic Parkinson’s disease. Acta Neurol Scand. 1984;70:20–28. doi: 10.1111/j.1600-0404.1984.tb00798.x. [DOI] [PubMed] [Google Scholar]
- Takakusaki K, Saitoh K, Harada H, Okumura T, Sakamoto T. Evidence for a role of basal ganglia in the regulation of rapid eye movement sleep by electrical and chemical stimulation for the pedunculopontine tegmental nucleus and the substantia nigra pars reticulata in decerebrate cats. Neuroscience. 2004;124:207–220. doi: 10.1016/j.neuroscience.2003.10.028. [DOI] [PubMed] [Google Scholar]
- Tanimura A, Pancani T, Lim SAO, Tubert C, Melendez AE, Shen W, Surmeier DJ. Striatal cholinergic interneurons and Parkinson’s disease. Eur J Neurosci. 2017 doi: 10.1111/ejn.13638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor JR, Elsworth JD, Roth RH, Sladek JR, Jr, Redmond DE., Jr Cognitive and motor deficits in the acquisition of an object retrieval/detour task in MPTP-treated monkeys. Brain. 1990;113(Pt 3):617–637. doi: 10.1093/brain/113.3.617. [DOI] [PubMed] [Google Scholar]
- Taylor TN, et al. Nonmotor symptoms of Parkinson’s disease revealed in an animal model with reduced monoamine storage capacity. J Neurosci. 2009;29:8103–8113. doi: 10.1523/JNEUROSCI.1495-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor TN, Greene JG, Miller GW. Behavioral phenotyping of mouse models of Parkinson’s disease. Behav Brain Res. 2010;211:1–10. doi: 10.1016/j.bbr.2010.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thevathasan W, et al. The impact of low-frequency stimulation of the pedunculopontine nucleus region on reaction time in parkinsonism. J Neurol Neurosurg Psychiatry. 2010;81:1099–1104. doi: 10.1136/jnnp.2009.189324. [DOI] [PubMed] [Google Scholar]
- Thompson JL, et al. Prefrontal dopamine D1 receptors and working memory in schizotypal personality disorder: a PET study with [(1)(1)C]NNC112. Psychopharmacology (Berl) 2014;231:4231–4240. doi: 10.1007/s00213-014-3566-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tolwani RJ, Jakowec MW, Petzinger GM, Green S, Waggie K. Experimental models of Parkinson’s disease: insights from many models. Lab Anim Sci. 1999;49:363–371. [PubMed] [Google Scholar]
- Unguez GA, Schneider JS. Dopaminergic dorsal raphe neurons in cats and monkeys are sensitive to the toxic effects of MPTP. Neurosci Lett. 1988;94:218–223. doi: 10.1016/0304-3940(88)90298-4. [DOI] [PubMed] [Google Scholar]
- Varastet M, Riche D, Maziere M, Hantraye P. Chronic MPTP treatment reproduces in baboons the differential vulnerability of mesencephalic dopaminergic neurons observed in Parkinson’s disease. Neuroscience. 1994;63:47–56. doi: 10.1016/0306-4522(94)90006-x. [DOI] [PubMed] [Google Scholar]
- Venator DK, Lewis DA, Finlay JM. Effects of partial dopamine loss in the medial prefrontal cortex on local baseline and stress-evoked extracellular dopamine concentrations. Neuroscience. 1999;93:497–505. doi: 10.1016/s0306-4522(99)00131-1. [DOI] [PubMed] [Google Scholar]
- Vezoli J, et al. Early presymptomatic and long-term changes of rest activity cycles and cognitive behavior in a MPTP-monkey model of Parkinson’s disease. PLoS One. 2011;6:e23952. doi: 10.1371/journal.pone.0023952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Villalba R, Smith Y. Significant increase in the density of striatal cholinergic interneurons in the caudate nucleus of MPTPtreated parkinsonian monkeys. Society of Neuroscience. 2017 [Google Scholar]
- Villalba RM, Smith Y. Differential structural plasticity of corticostriatal and thalamostriatal axo-spinous synapses in MPTP-treated Parkinsonian monkeys. J Comp Neurol. 2011;519:989–1005. doi: 10.1002/cne.22563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Villalba RM, Wichmann T, Smith Y. Neuronal loss in the caudal intralaminar thalamic nuclei in a primate model of Parkinson’s disease. Brain Struct Funct. 2014;219:381–394. doi: 10.1007/s00429-013-0507-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vingerhoets FJ, Snow BJ, Tetrud JW, Langston JW, Schulzer M, Calne DB. Positron emission tomographic evidence for progression of human MPTP-induced dopaminergic lesions. Ann Neurol. 1994;36:765–770. doi: 10.1002/ana.410360513. [DOI] [PubMed] [Google Scholar]
- Voorn P, Kalsbeek A, Jorritsma-Byham B, Groenewegen HJ. The pre- and postnatal development of the dopaminergic cell groups in the ventral mesencephalon and the dopaminergic innervation of the striatum of the rat. Neuroscience. 1988;25:857–887. doi: 10.1016/0306-4522(88)90041-3. [DOI] [PubMed] [Google Scholar]
- Waters CM, Hunt SP, Jenner P, Marsden CD. An immunohistochemical study of the acute and long-term effects of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine in the marmoset. Neuroscience. 1987;23:1025–1039. doi: 10.1016/0306-4522(87)90178-3. [DOI] [PubMed] [Google Scholar]
- Whitehouse PJ, Hedreen JC, White CL, 3rd, Price DL. Basal forebrain neurons in the dementia of Parkinson disease. Ann Neurol. 1983;13:243–248. doi: 10.1002/ana.410130304. [DOI] [PubMed] [Google Scholar]
- Whone AL, Moore RY, Piccini PP, Brooks DJ. Plasticity of the nigropallidal pathway in Parkinson’s disease. Ann Neurol. 2003;53:206–213. doi: 10.1002/ana.10427. [DOI] [PubMed] [Google Scholar]
- Wichmann T, DeLong MR. Pathophysiology of Parkinson’s disease: the MPTP primate model of the human disorder. Ann N Y Acad Sci. 2003;991:199–213. doi: 10.1111/j.1749-6632.2003.tb07477.x. [DOI] [PubMed] [Google Scholar]
- Williams SM, Goldman-Rakic PS. Widespread origin of the primate mesofrontal dopamine system. Cereb Cortex. 1998a;8:321–345. doi: 10.1093/cercor/8.4.321. [DOI] [PubMed] [Google Scholar]
- Williams SM, Goldman-Rakic PS. Widespread origin of the primate mesofrontal dopamine system. Cerebr Cortex. 1998b;8:321–345. doi: 10.1093/cercor/8.4.321. [DOI] [PubMed] [Google Scholar]
- Wilson DA, Sullivan RM. The D2 antagonist spiperone mimics the effects of olfactory deprivation on mitral/tufted cell odor response patterns. J Neurosci. 1995;15:5574–5581. doi: 10.1523/JNEUROSCI.15-08-05574.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winterer G, Weinberger DR. Genes, dopamine and cortical signal-to-noise ratio in schizophrenia. Trends Neurosci. 2004;27:683–690. doi: 10.1016/j.tins.2004.08.002. [DOI] [PubMed] [Google Scholar]
- Yang W, et al. Mutant alpha-synuclein causes age-dependent neuropathology in monkey brain. J Neurosci. 2015;35:8345–8358. doi: 10.1523/JNEUROSCI.0772-15.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan HH, Chen RJ, Zhu YH, Peng CL, Zhu XR. The neuroprotective effect of overexpression of calbindin-D(28k) in an animal model of Parkinson’s disease. Mol Neurobiol. 2013;47:117–122. doi: 10.1007/s12035-012-8332-3. [DOI] [PubMed] [Google Scholar]
- Yue Z. Genetic mouse models for understanding LRRK2 biology, pathology and pre-clinical application. Parkinsonism Relat Disord. 2012;18(Suppl 1):S180–182. doi: 10.1016/S1353-8020(11)70056-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zarow C, Lyness SA, Mortimer JA, Chui HC. Neuronal loss is greater in the locus coeruleus than nucleus basalis and substantia nigra in Alzheimer and Parkinson diseases. Arch Neurol. 2003a;60:337–341. doi: 10.1001/archneur.60.3.337. [DOI] [PubMed] [Google Scholar]
- Zarow C, Lyness SA, Mortimer JA, Chui HC. Neuronal loss is greater in the locus coeruleus than nucleus basalis and substantia nigra in Alzheimer and Parkinson diseases. Archives of neurology. 2003b;60:337–341. doi: 10.1001/archneur.60.3.337. [DOI] [PubMed] [Google Scholar]
- Zeng BY, Iravani MM, Jackson MJ, Rose S, Parent A, Jenner P. Morphological changes in serotoninergic neurites in the striatum and globus pallidus in levodopa primed MPTP treated common marmosets with dyskinesia. Neurobiol Dis. 2010;40:599–607. doi: 10.1016/j.nbd.2010.08.004. [DOI] [PubMed] [Google Scholar]
- Zesiewicz TA, Baker MJ, Wahba M, Hauser RA. Autonomic Nervous System Dysfunction in Parkinson’s Disease. Curr Treat Options Neurol. 2003;5:149–160. doi: 10.1007/s11940-003-0005-0. [DOI] [PubMed] [Google Scholar]
- Zesiewicz TA, Hauser RA. Depression in Parkinson’s disease. Curr Psychiatry Rep. 2002;4:69–73. doi: 10.1007/s11920-002-0016-7. [DOI] [PubMed] [Google Scholar]
- Zilles K, et al. Structural asymmetries in the human forebrain and the forebrain of non-human primates and rats. Neurosci Biobehav Rev. 1996;20:593–605. doi: 10.1016/0149-7634(95)00072-0. [DOI] [PubMed] [Google Scholar]
- Zweig RM, Jankel WR, Hedreen JC, Mayeux R, Price DL. The pedunculopontine nucleus in Parkinson’s disease. Ann Neurol. 1989;26:41–46. doi: 10.1002/ana.410260106. [DOI] [PubMed] [Google Scholar]