

Abstract
The control of seed germination and seed dormancy are critical for the successful propagation of plant species, and are important agricultural traits. Seed germination is tightly controlled by the balance of gibberellin (GA) and abscisic acid (ABA), and is influenced by environmental factors. The COP9 Signalosome (CSN) is a conserved multi-subunit protein complex that is best known as a regulator of the Cullin-RING family of ubiquitin E3 ligases (CRLs). Multiple viable mutants of the CSN showed poor germination, except for csn5b-1. Detailed analyses showed that csn1-10 has a stronger seed dormancy, while csn5a-1 mutants exhibit retarded seed germination in addition to hyperdormancy. Both csn5a-1 and csn1-10 plants show defects in the timely removal of the germination inhibitors: RGL2, a repressor of GA signaling, and ABI5, an effector of ABA responses. We provide genetic evidence to demonstrate that the germination phenotype of csn1-10 is caused by over-accumulation of RGL2, a substrate of the SCF (CRL1) ubiquitin E3 ligase, while the csn5a-1 phenotype is caused by over-accumulation of RGL2 as well as ABI5. The genetic data are consistent with the hypothesis that CSN5A regulates ABI5 by a mechanism that may not involve CSN1. Transcriptome analyses suggest that CSN1 has a more prominent role than CSN5A during seed maturation, but CSN5A plays a more important role than CSN1 during seed germination, further supporting the functional distinction of these two CSN genes. Our study delineates the molecular targets of the CSN complex in seed germination, and reveals that CSN5 has additional functions in regulating ABI5, thus the ABA signaling pathway.
Author summary
The control of seed germination and seed dormancy are critical for successful propagation of plant species, and manipulation of these processes is important for agriculture. The COP9 Signalosome (CSN) is a multi-subunit protein complex that regulates proteasome-mediated protein degradation in part as a regulator of SCF ubiquitin E3 ligases. The CSN is important for timely germination of seeds, but its molecular targets in this process is unclear. In this study, we demonstrate that the CSN regulates protein stabilities of two different targets from two antagonistic hormonal pathways, RGL2 of the GA pathway and ABI5 of the ABA pathway. Our genetic and transcriptome analyses showed that, although csn1-10 and csn5a-1 exhibit similar defects in timely germination, the mechanisms of how the mutations affect seed germination differ. Since RGL2 is known to be targeted by SCF during germination, the defect in the timely degradation of RGL2 in csn1-10 and csn5a-1 is consistent with the role of CSN as a regulator of the SCF. In addition, we show that CSN5A, but not CSN1, has an additional function in regulating ABI5, a downstream inhibitor of germination.
Introduction
Seed germination launches the active growth phase of a plant, while seed dormancy prevents germination even under optimal growth conditions. The decision and the processes of seed germination are modulated by many factors but predominantly by gibberellin (GA) and abscisic acid (ABA), two phytohormones which act antagonistically on seed germination [1, 2]. ABA levels become elevated during seed maturation to establish and maintain seed dormancy, and its levels drop sharply upon imbibition of seeds. On the other hand, GA biosynthesis starts upon seed imbibition, and GA is necessary to release seed dormancy and stimulate germination [3]. In Arabidopsis, the GA biosynthetic mutant ga1-3 cannot germinate without an exogenous supply of GA, demonstrating the necessity of GA in seed germination [4, 5]. Various environmental factors such as light, moisture, temperature, and nutrients (e.g. nitrate) can affect germination both during seed maturation and during seed imbibition. Those environmental factors modulate germination in a large part through altering the levels of GA and ABA [6–8]. In the laboratory, seed dormancy is released by a period of dry storage (termed after-ripening) or by cold stratification.
The GA response pathway is negatively controlled by the DELLA proteins, consisting of five members in Arabidopsis: GA-INSENSITIVE (GAI), REPRESSOR OF ga1-3 (RGA), RGA-LIKE1 (RGL1), RGA-LIKE2 (RGL2), and RGA-LIKE3 (RGL3) [9]. In response to GA, the DELLA proteins are rapidly degraded by the ubiquitin-proteasome system via SCFSLY1/2, which results in GA-stimulated growth and development [10, 11]. Among the DELLA proteins, RGL2 plays a major role as a GA-regulated repressor in seed germination, as rgl2 can rescue the germination defect of ga1-3 in the absence of exogenous GA [12, 13]. In addition, RGA and GAI, together with PIL5/PIF1 regulate light-mediated control of seed germination [14, 15]. Under white light, RGL2 plays a predominant role in endosperm tissue, and it also has a central function in the crosstalk with ABA signaling during seed germination [16–18]. ABA induces a number of effectors, including the bZIP transcription factor ABA INSENSITIVE5 (ABI5). ABI5 accumulates during seed maturation and in dry seeds [19, 20]. During the normal course of seed germination, ABA and concomitantly ABI5 levels rapidly decline following imbibition and GA biosynthesis, enabling seed germination. ABI5 has been implicated as the final inhibitor of seed germination, possibly acting downstream of the GA repressor RGL2 [16, 17].
The COP9 signalosome (CSN) is a conserved heteromeric protein complex known to regulate the CULLIN-RING family of ubiquitin E3 ligases (CRLs), including the SCF sub-family of E3s [21]. Biochemically, CSN inhibits CRL E3 activity by removing the NEDD8 (RUB1) modification on the CULLIN subunit (a process known as de-neddylation or de-rubylation) [22, 23], and by direct interaction with the CRL core components [24–26]. However, genetic studies in several organisms including Arabidopsis have shown that CSN promotes the functions of the CRLs [22, 27]. In a number of cases, CSN activity has been shown to protect components of the CRL E3s against their autoubiquitination activity [28, 29]. Still, several studies also indicate that not all substrates of CRL and SCF E3s are regulated by CSN, since some of the SCF substrates display normal signal dependent degradation in CSN-deficient cells [30, 31]. Our understanding of the specific roles of the CSN in SCF-mediated substrate ubiquitination remains incomplete.
In Arabidopsis where the CSN was initially identified, complete loss of any one subunit destabilizes the entire complex [32]. As a result, all of the null csn mutants exhibit characteristic purple seeds (the fusca phenotype) and developmental arrest soon after germination [33, 34]. Since CSN5 and CSN6 are each encoded by two functionally redundant genes, CSN5A vs. CSN5B, and CSN6A vs. CSN6B, respectively, a null mutation in either of the CSN5 or CSN6 genes are viable, while knocking out both genes of either CSN5 (csn5a-1 5b-1) or CSN6 (csn6a-1 6b-1) lead to the lethal fusca phenotype, like that of other csn null mutants [35–37]. Studies using both lethal and weak mutants of the CSN have shown that the CSN is involved in multifaceted developmental processes and physiological responses. For examples, CSN works with SCFTIR1 in auxin responses, with SCFUFO in flower development, and with SCFCOI1 in JA responses. CSN also affects GA signaling, cell divisions, stem cell functions, root patterning, and defenses [22, 27, 38–42]. However, in many of these cases, the specific substrates of the SCF for the corresponding process have not been clearly identified.
It has been observed that lethal mutants of the CSN require extended cold stratification to germinate. The precise germination rates of the lethal mutants were difficult to measure, because the mutants can only be maintained as a heterozygous population. Weak mutants such as csn5a-2 also show defects in germination [39, 43]. However, the specific targets and the mechanisms underlying this phenotype remain obscure. In this study, we carried out a systematic study of the germination and dormancy phenotypes using viable csn mutants, including csn5a-1 and csn1-10. We demonstrate here that CSN regulates seed germination by modulating the levels of RGL2 and ABI5 in the GA and ABA pathways, respectively.
Results
Molecular characterization of viable mutants of the CSN
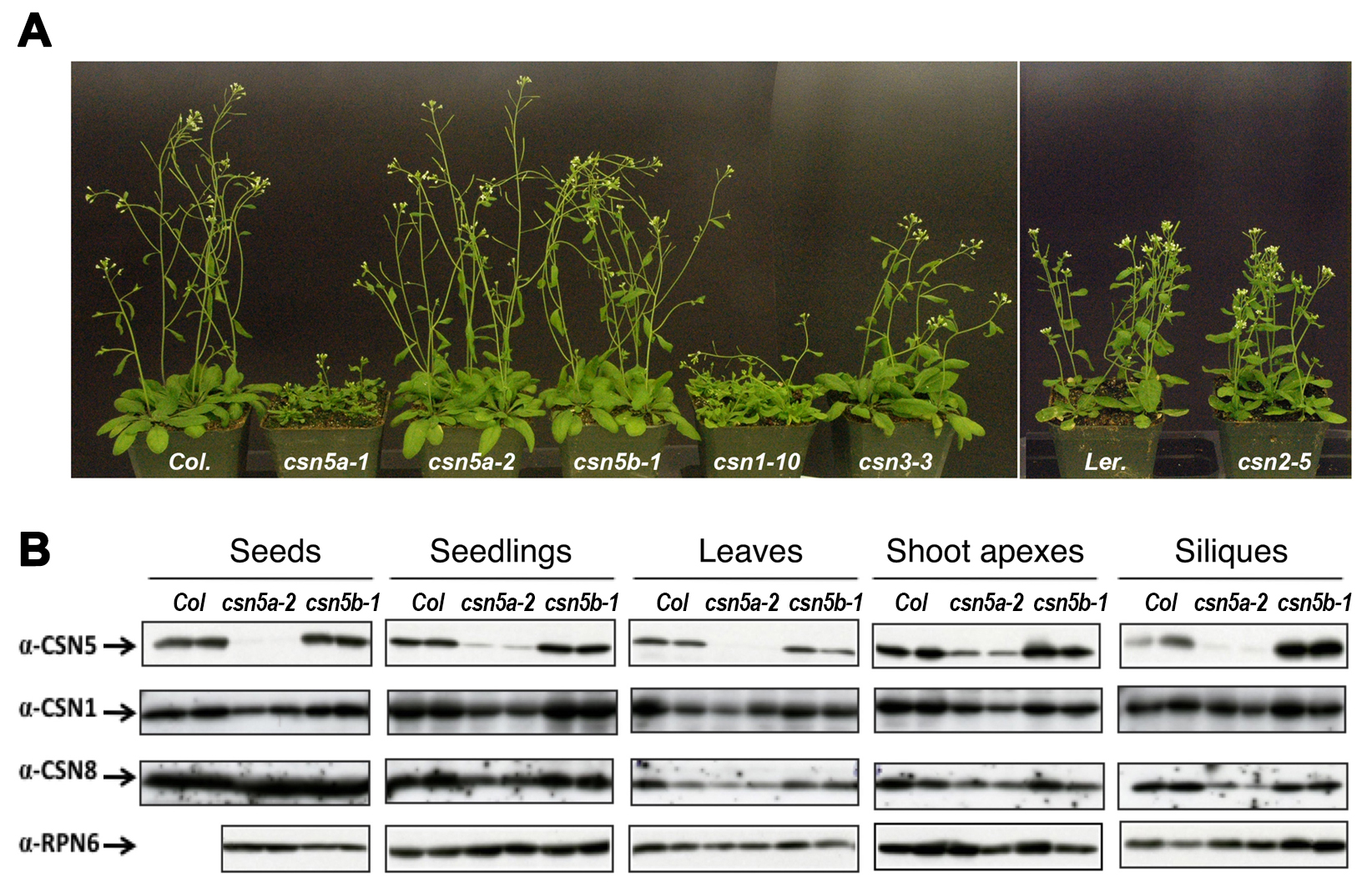
In recent years, the availability of viable and fertile mutants of Arabidopsis CSN, which can produce homozygous mutant seeds, has provided a feasible genetic material for a systematic analysis of the role of the CSN in seed germination. First, we conducted a basic molecular comparison of a number of viable csn mutants for the levels of CSN subunits in plants (Fig 1 and S1 Fig). csn5a-1 and csn5b-1 are null mutants of the respective genes, while csn5a-2 is a weak allele of CSN5A [37, 44]. The CSN5A gene is considered to predominate over CSN5B, based on multiple microarray datasets [36] as well as the observation that csn5a-1 mutants showed a more severe growth defect than those of csn5b-1 mutants [35, 37] (S1A Fig). The anti-CSN5A antibody could readily detect endogenous CSN5A, but not CSN5B (Fig 1A and S1B Fig), while the anti-CSN5B antibody could detect both CSN5A and CSN5B, and showed that endogenous CSN5B proteins migrate more slowly than CSN5A in SDS-PAGE (Fig 1B). It should be mentioned that under our growth conditions, csn5a-2 plants can tolerate a considerable reduction of CSN5A levels (S1B Fig), without showing obvious growth defects, although the same csn5a-2 allele was previously reported to show a noticeable growth defect [35, 37]. For the other viable mutants used, csn1-10, which contains a point mutation that reduces the level of CSN1 expression [45], exhibited a clear growth defect, while csn3-3 [46] and csn2-5 [Landsberg erecta (Ler) background] [47] appeared to be phenotypically similar to the respective wild type plants (S1A Fig).
Fig 1. Expression of CSN gene products and CUL1 modification status in various viable csn mutants.

(A) Apical apex tissues from indicated mutants were extracted and the proteins examined by immunoblotting using indicated antibodies. Lower levels of the CSN (CSN3 and CSN5A) and hyper-neddylation of CUL1 were found in csn5a-1, csn5a-2, csn1-10, and csn2-5. (B) Light-grown seedlings (4-day) of indicated genotypes were used for immunoblotting with anti-CSN5B. The antibody strongly reacted with CSN5B, but also cross-reacted with CSN5A. Endogenous CSN5B migrated slower than CSN5A on the SDS-PAGE. See also S1 Fig.
In Arabidopsis, CSN subunits are stabilized through assembly of the complex [32, 35, 48]. Immunoblot analyses showed that this group of viable csn mutants contained variably lowered protein levels of other CSN subunits (Fig 1 and S1 Fig). In particular, csn5a-1 and csn1-10 displayed the most noticeable reductions in the steady state level of CSN3 and CSN5 (Fig 1), suggesting that the level of the CSN complex was lower in these mutants. Consistent with previous reports, csn5a-1, csn5a-2, csn1-10, and csn2-5, but not csn5b-1 or csn3-3, caused hyperneddylation of CUL1, indicating that these mutants had lower CSN-mediated deneddylation activity (Fig 1). Therefore, csn5a-1, csn5a-2, csn1-10, and csn2-5 are definitively partial loss-of-function mutants of the CSN.
csn mutants exhibit hyperdormancy and delayed seed germination
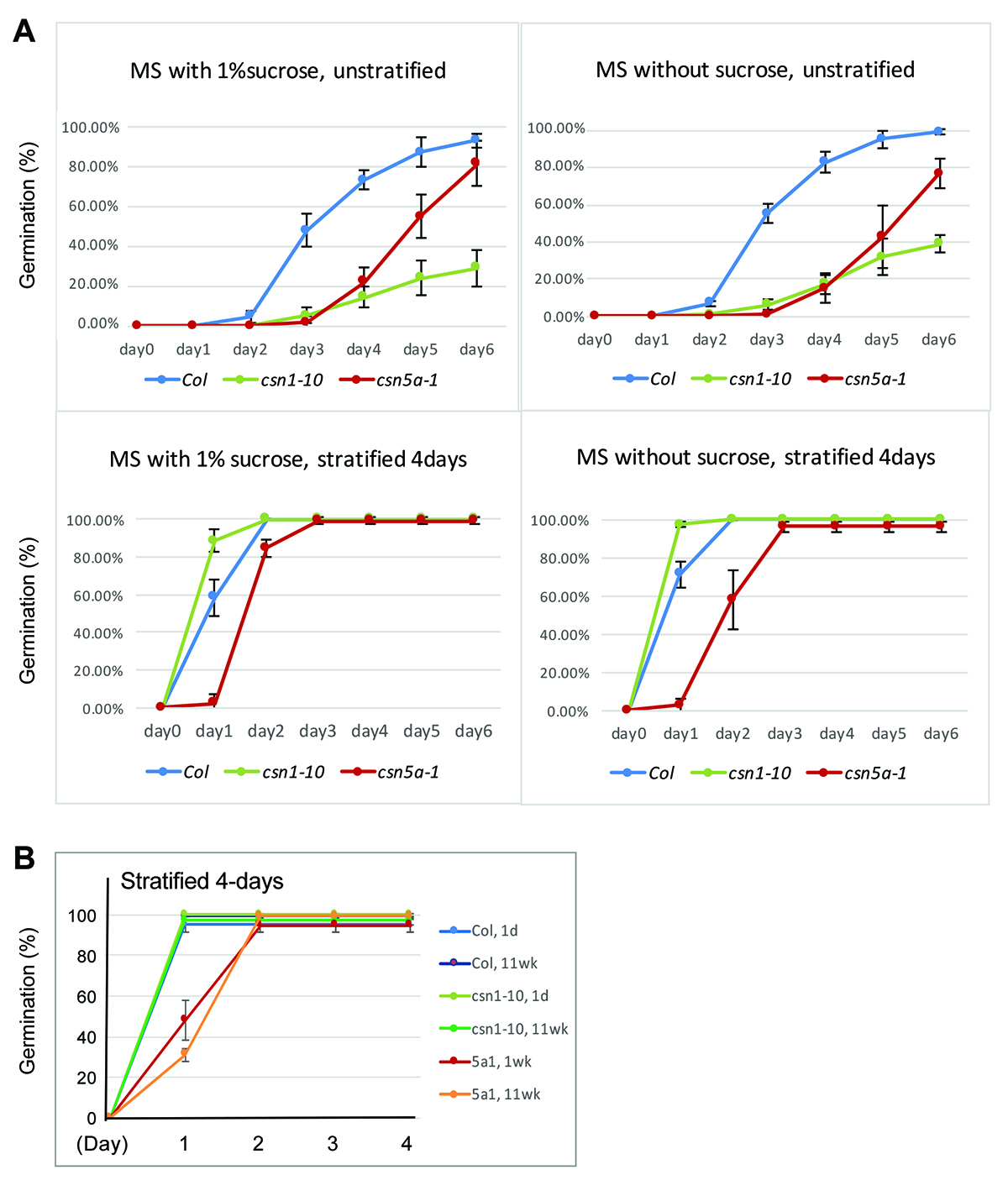
We examined germination of the csn mutant seeds with or without cold stratification. When Arabidopsis Columbia (Col) seeds were cold stratified at 4°C for 3 days or more, germination was accelerated by 12–24 hours over unstratified seeds under our growth and planting conditions (Fig 2A and 2B), indicating that those Col (wild type) seeds exhibited a weak dormancy response. Without cold stratification, csn1-10, csn3-3, 5a-1 and 5a-2 exhibited noticeably delayed and poor germination compared to Col. csn2-5 also exhibited a slight but consistently poor seed germination compared to the corresponding wild type (Ler). The germination defects could be nearly completely alleviated by cold stratification in csn1-10, csn3-3, and csn2-5 seeds (Fig 2A and 2B). However, in csn5a-1 and csn5a-2, cold stratification could significantly, but not completely, alleviate their germination defects (Fig 2B, S2 Fig). These observations suggest that most of the csn mutants had stronger seed dormancy. The exception was csn5b-1, which appeared to have a weaker dormancy than Col, as it germinated equally well regardless whether or not the seeds had been cold stratified. Since our medium contained 1% sucrose, we tested mutant germination on plates without sucrose, and found that csn1-10 and csn5a-1 showed similar germination phenotype regardless of whether sucrose was present or not (S2A Fig).
Fig 2. Seed germination phenotype of the csn mutants.

(A) Representative images showing seed germination status of indicated mutants. Seeds were cold stratified for 4 days (Cold Str) or had not stratified (Un-str). Photographs were taken at day 6. (B) Germination rates of unstratified (solid lines) or cold stratified (broken lines) seeds of the csn mutants. csn5a-1, csn5a-2, csn5b-1, csn1-10, csn3-3 were compared to Col wild type, while csn2-5 to Ler wild type. With the exception of csn5b-1, all other csn mutants tested exhibited slower and lower germination rates compared to Col, when not cold stratified. (C) Germination rates of Col, csn1-10 and csn5a-1 seeds that had been in dry storage for indicated period of time. csn1-10 and csn5a-1 seeds required extended after-ripening time than Col to release dormancy. csn5a-1 additionally showed delayed germination. (D) Germination rates of freshly collected, partially after-ripened (2 weeks), or fully after-ripened (11 weeks) seeds of Col, csn1-10, and csn5a-1. The germination deficiency of csn1-10 and csn5a-1 were most readily observed with the seeds of two weeks after seed-collection. csn5a-1 seeds exhibited a delay in germination even in fully after-ripened seeds. In all panels, error bars represent standard deviation from 4 repeats (n = 4). See also S2 and S3 Figs.
To further confirm the seed dormancy phenotype, we examined the germination rates in relation to different after-ripening storage ages of the seeds in csn1-10, csn5a-1, and the Col control. As shown in Fig 2C and 2D, freshly collected seeds (1 or 2 days after collection) of either Col or the mutants showed low and heterogeneous germination. While the Col seeds showed significant dormancy-release after two weeks, csn1-10 and csn5a-1 seeds continued to germinate slowly, consistent with the stronger seed dormancy of the mutants. After extended dry storage (11 weeks), csn1-10 mutants could catch up with Col in germination, but csn5a-1 still displayed a delay in germination compared to Col even in fully after-ripened seeds (1-year) (Fig 2C and 2D). This result, as well as the observation that cold stratification could not fully alleviate the germination defect of the csn5a mutants, suggest that while csn1-10 has a seed hyperdormancy phenotype, csn5a-1 shows a delay in germination in addition to hyperdormancy. As shown in Fig 2D, the germination phenotype of the mutants is best displayed in partially after-ripened seeds, typically between 1–4 weeks after seed collection, although the window of phenotypic alteration varies depending on different batches of seeds.
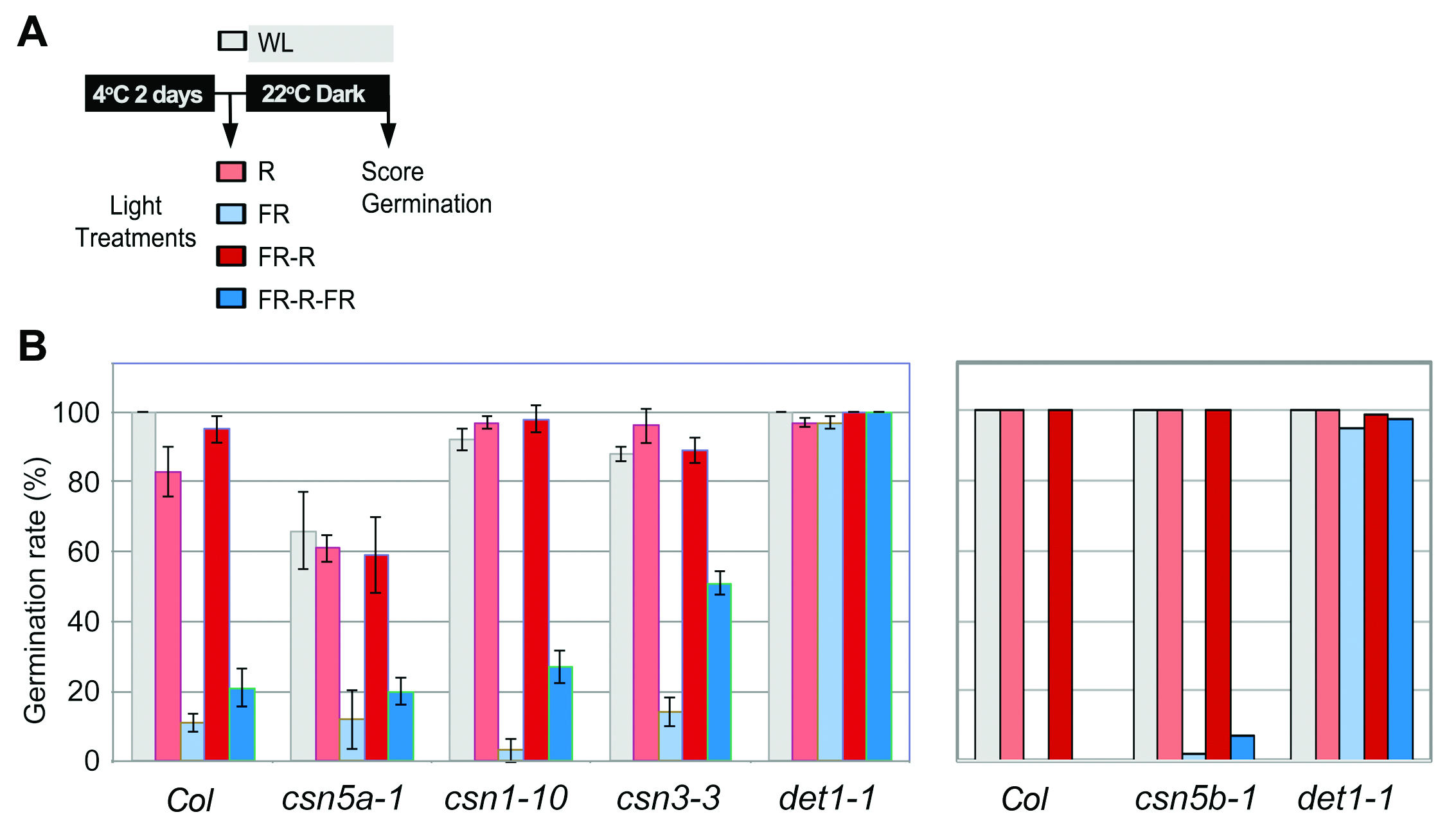
Since most of the csn mutants have been reported to have mild photomorphogenic phenotypes, we tested the response of the mutants to phytochrome B (phyB)-controlled seed germination. To distinguish the light response from other aspects of dormancy responses, cold stratified seeds were used. Seeds were then subjected to a 5-min pulse of red (R) light, far red (FR) light, or alternating R and FR treatments in a sequential order as indicated in S3 Fig. The det1-1 mutant, which lacks a phyB-controlled seed germination response and germinates regardless of light treatments [49], was used as a control. The data showed clearly that all of the tested csn mutants, including csn1-10, csn3-3, csn5a-1 and csn5b-1, displayed normal responses in phyB-mediated seed germination (S3 Fig). Thus, for the rest of the studies on the mechanisms of the CSN-regulated seed germination, we carried out the experiments under white light. When not specified, un-stratified seeds were used for the germination and biochemical analyses.
The germination phenotype of csn1-10 and csn5a-1 is seed-coat-dependent
The seed coat plays an important role in seed dormancy in many plant species including Arabidopsis [50]. The Arabidopsis seed coat consists of an outer layer of maternally derived material known as the testa. Underneath the testa is a single cell layer of endosperm that encloses the embryo. In the hyperdormant ecotypes Cvi and C24, or in non-germinating ga1-3 mutants, removal of the seed coat can break dormancy and allow the embryo to grow [51]. To determine whether the germination defects in csn1-10 and 5a-1 were imposed by the seed coat, we removed the seed coat by dissection, and observed the growth of the embryos with regard to radicle (embryonic root) growth and cotyledon greening and expansion.
Without the seed coat, both csn5a-1 (Fig 3A) and csn1-10 (Fig 3B) mutant embryos displayed radicle growth on day-2, and exhibited greening and cotyledon expansion on day-3, similar to the developmental time line of comparably treated Col embryos and seeds. Remarkably, the removal of the seed-coat even rescued the germination of csn1-1 (or fus6-1, cop11-1) (Fig 3C), the null mutant of csn1, which were otherwise extremely dormant such that mutant seeds rarely germinate without cold stratification. After germination, csn1-1 mutants arrested further development, and they died as purple seedlings (Fig 3C bottom right corner panel), similar to the final morphology of the seedlings germinated from intact seeds [34]. These results showed that csn1 and csn5a mutant embryos have the intrinsic capacity to initiate growth on a similar time scale to the Col embryos, and that the hyperdormancy phenotype of the mutant seeds is dependent on the seed coat. Seed coat-dependent inhibition of seed germination has been shown to be mediated at least in part by RGL2 in the endosperm, where it stimulates ABA synthesis, and ultimately ABI5 activity [16, 51].
Fig 3. The germination defects of csn1 and csn5a are seed-coat dependent.
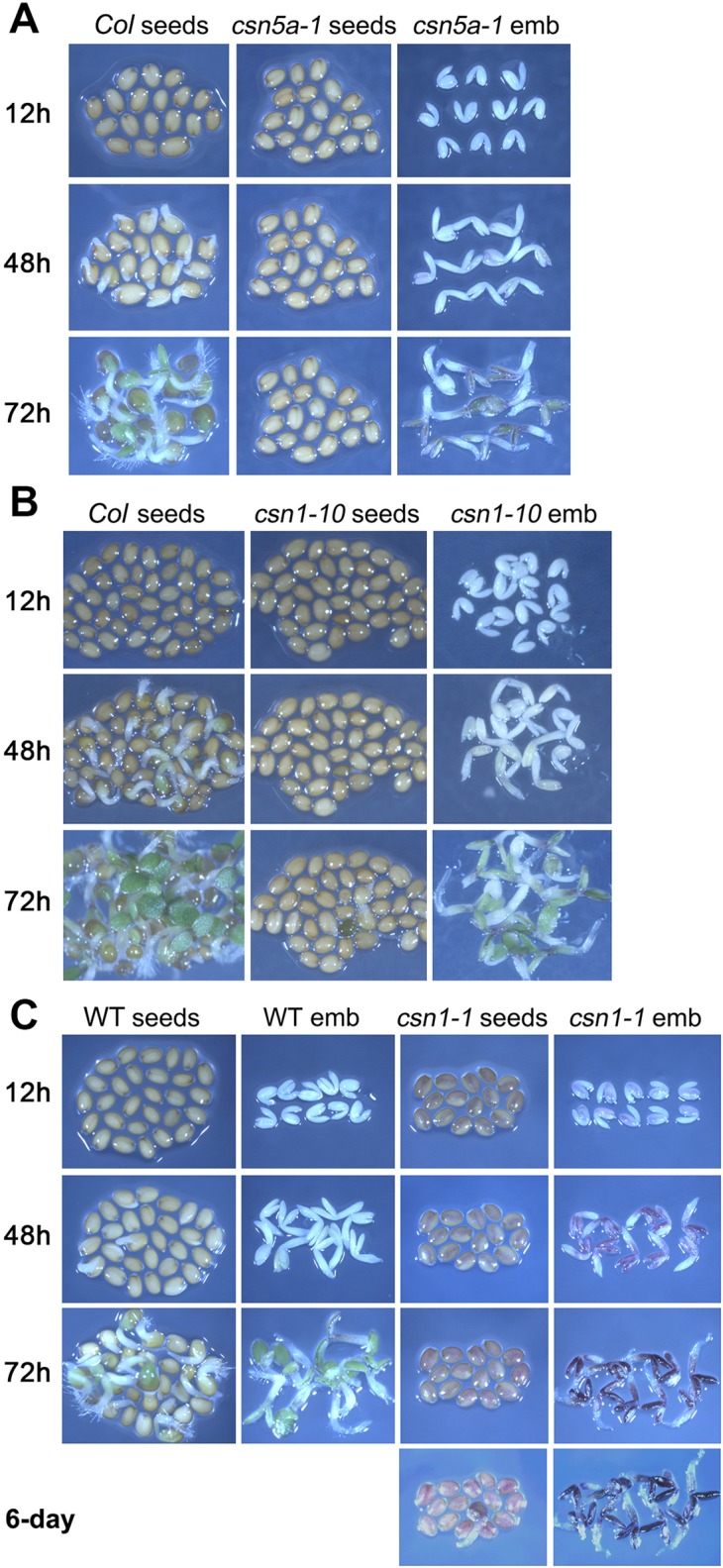
Intact seeds or embryos dissected from the same seed population were germinated on water-agar plates. (A) csn5a-1, or (B) csn1-10, with the corresponding wild type Col seeds as the controls. (C) csn1-1 (fus6-1) mutant seeds were picked out from the heterozygous (+/-) seed population based on the dark purple seed color, and normal colored seeds were used as the positive control (WT). While the seeds of the csn mutant could not germinate on time, their embryos showed normal progression of growth, with radicle elongation on day-2 and cotyledon greening and expansion on day-3. csn1-1 mutants arrested the growth after germination.
csn mutants are defective in timely removal of germination inhibitors RGL2 and ABI5
GA stimulate seed germination by removal of GA inhibitors known as DELLA proteins. Among the five DELLA proteins in Arabidopsis, RGA and GAI function mostly in the dark and in the embryos, while RGL2 is the main inhibitor of seed germination under light [12, 18]. RGL2 also plays a central role in seed coat-dependent inhibition of seed germination, which likely involves CSN. RGL2 is predominantly regulated at level of protein turnover via SCFSLY1 E3 ligase mediated protein degradation through the ubiquitin-proteasome system [10, 11]. We examined RGL2 protein levels in the csn mutants by anti-RGL2 immunoblotting in a time course analysis. Both csn1-10 (Fig 4A) and csn5a-1 (Fig 4B) displayed clear defects in timely degradation of RGL2. Abnormal accumulation of RGL2 is consistent with the idea that degradation of RGL2 via an SCF E3 ubiquitin ligase requires a fully active CSN complex.
Fig 4. csn1-10 and csn5a-1 showed defects in timely down-regulation of RGL2 and ABI5 proteins following seed imbibition.

(A and B) Time-course analysis of RGL2 levels by immunoblots showing the deficiency of csn1-10 (A) and csn5a-1 (B) in timely decline of RGL2 protein compared to wild type (Col). Anti-RPN6 blot or a background band were shown as the loading reference. (C) Time course analysis of ABI5 level by immunoblotting. csn1-10, csn5a-1, csn5a-2, but not csn5b-1, showed defect in timely decline of ABI5 protein following seed imbibition. Anti-histone H3 was used as an internal loading reference. The lower panel shows the densitometry quantification of ABI5 relative to the corresponding Histone H3 in each sample. Samples were collected at indicated time points in hours (hr) post imbibition, as described in Materials and Methods.
Another major inhibitor of seed germination is ABI5, which accumulates in dry seeds and in response to low GA or high ABA conditions. We next examined ABI5 protein levels by anti-ABI5 immunoblotting over the course of germination. While ABI5 proteins were depleted by day-3 in Col wild type and csn5b-1 after imbibition, they remained high in csn1-10 and csn5a-1 mutants, and showed slower kinetics of the decline in csn5a-2 (Fig 4C). These results implied that csn1 and csn5a mutants are defective in ABI5 downregulation following imbibition of the seeds. Taken together, csn1-10 and csn5a mutants exhibited defects in the timely removal of RGL2 and ABI5, two key inhibitors of seed germination in Arabidopsis, which correlated with the deficiency of the mutants in their timely germination.
RGL2 accumulation underlies the germination defect of csn1-10, but it is insufficient to account for the germination phenotype of csn5a-1
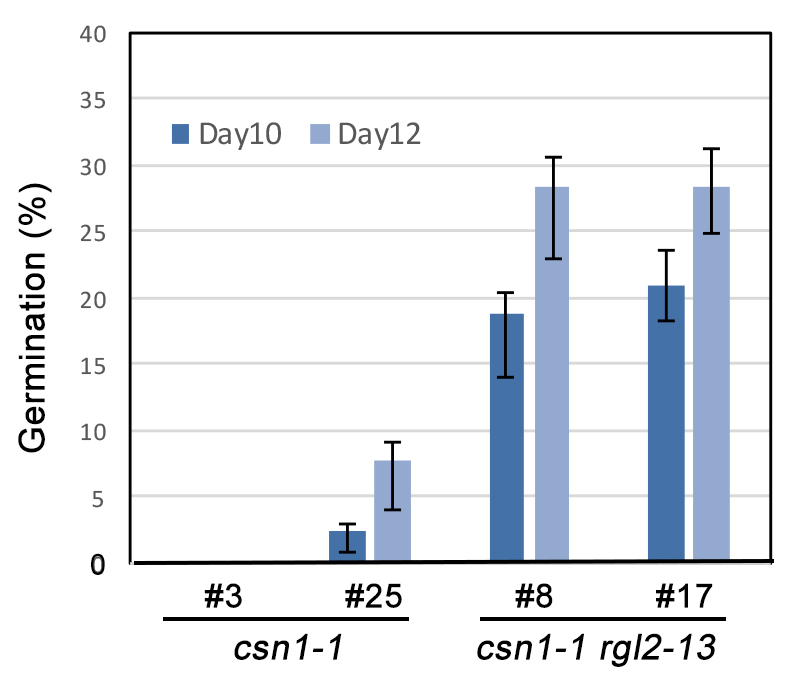
If accumulation of RGL2 is responsible for the germination phenotype of the csn mutants, genetic removal of the RGL2 gene should be able to rescue the defects of the csn mutants. To test this, we crossed the csn mutants with the null mutant of RGL2, rgl2-13 (Col) [13], and generated the csn1-10 rgl2-13 and csn5a-1 rgl2-13 double mutants. Germination tests showed that introduction of rgl2-13 into the csn1-10 mutant effectively rescued its germination phenotype (Fig 5A). Moreover, rgl2-13 could significantly alleviate the poor germination of the csn1-1 lethal allele, enabling the un-stratified csn1-1 rgl2-13 double mutant seeds to germinate, albeit at a slower rate (S4 Fig). In contrast to csn1, the germination defect of csn5a-1 could not be rescued by rgl2-13 (Fig 5B). The csn5a-1 rgl2-13 double mutant showed a poor germination profile that was similar to that of csn5a-1 (Fig 5B). These results suggest that, while over-accumulation of RGL2 is responsible for the seed hyperdormancy of csn1-10, additional components may be involved in the germination defects of csn5a-1.
Fig 5. csn1-10 and csn5a-1 show different genetic interactions with rgl2-13, and different responses to PAC.

(A and B) rgl2-13 rescued the germination deficiency of csn1-10 but not csn5a-1. (A) The csn1-10 rgl2-13 double mutants displayed normal germination rate as wild type Col and rgl2-13, in contrast to low germination of csn1-10. Double mutants db26 and db35 are two different lines of the csn1-10 rgl2-13. (B) Germination rate of csn5a-1 rgl2-13 double mutants displayed low germination rate similar to csn5a-1 single mutant, compared to wild type Col and rgl2-13. Double mutants db19 and db20 are two different lines of the csn5a-1 rgl2-13. (C) csn5a-1 was hypersensitive to PAC. Cold stratified seeds were germinated on regular GM plate, GM plate containing PAC (1 μM), or PAC (1 μM) plus GA3 (50 μM). Seed germination rates were shown in graphs. (D) rgl2-13 conferred resistance to PAC (1 μM) in germination of rgl2-13 and double mutant csn1-10 rgl2-13, but not double mutant csn5a-1 rgl2-13. (E) The csn5a-1 rgl2-13 double mutant showed abnormally prolonged accumulation of ABI5, similar to csn5a-1 single mutant. Samples were taken at indicated times after imbibition of the seeds, and proteins were probed with antibodies as indicated on the left. (A-D) Error bars represent standard deviation from 4 repeats (n = 4). See also S4 Fig.
The csn1-10 and csn5a-1 mutants also differ in their responses to low GA conditions induced by PAC (paclobutrazol), an inhibitor of GA biosynthesis. To test the sensitivity to PAC, cold stratified seeds were used to factor out the dormancy effect. Treatment with PAC inhibited germination of wild type seeds, and the inhibition could be reversed by simultaneously supplying GA3 (PAC + GA, Fig 5C). As previously reported [12, 13], rgl2-13 was resistant to PAC in germination assays. Both csn1-10 and csn5a-1 were sensitive to PAC as Col, and the germination of Col and csn1-10, but not of csn5a-1, could be significantly restored by addition of GA3 (Fig 5C), indicating that csn5a-1 was hypersensitive to PAC-induced low GA condition. We next examined the double mutants for their responses to PAC. As shown in Fig 5D, rgl2 and csn1 rgl2 mutants were able to germinate well in PAC, indicating that removal of RGL2 is sufficient to overcome the effect of PAC even when CSN1 is deficient. This result is in agreement with the notion that the main function of CSN1 in germination is to modulate RGL2 turnover. However, the rgl2 mutation had only a slight effect on overcoming PAC treatment in the background of csn5a-1, as the germination rates of csn5a rgl2 double mutants remained very low in PAC (Fig 5D), suggesting that removal of RGL2 was insufficient to allow germination in the absence of CSN5A. Thus, it appeared that an additional factor(s) apart from RGL2 was affected by csn5a-1, that prevented timely germination or for germinating under low GA conditions. Time-course immunoblotting showed that, as in the csn5a-1 single mutant, high levels of ABI5 still accumulated in csn5a-1 rgl2-13 on day-4 post imbibition (Fig 5E). This result pointed to ABI5 as a factor that is potentially regulated by CSN5A.
Loss of ABI5 can rescue the germination phenotype of both csn1-10 and csn5a
ABI5 is thought to act downstream of RGL2 to inhibit seed germination under unfavorable environments [16, 17, 52]. To address whether the characteristic over-accumulation of ABI5 protein in the csn mutants was responsible for their germination phenotypes, double mutants of abi5-4 with csn1-10 or csn5a were generated. Since the abi5-4 mutant is in the Wassilewskija (Ws) ecotype [20] while csn5a-1 and csn1-10 mutants are in the Col background, we used segregating sibling lines of different genotypes in all of the germination comparisons, including WT, csn1-10, and csn5a-1, abi5-4, and the double mutants (Fig 6), in which “WT”was a segregating sibling line with a wild type genotype. Clearly, abi5-4 rescued the germination phenotype of both csn1-10 and csn5a-1, as demonstrated by strongly improved germination in the respective double mutants, i.e. csn1-10 abi5-4 (Fig 6A), or csn5a-1 abi5-4 (Fig 6B) compared to csn1-10 or csn5a-1, respectively. In addition, the double mutant of abi5-1 csn5a-2 also rescued the slow germination of csn5a-2, a weaker mutant of csn5a (S5 Fig). The finding that abi5-4 can rescue the germination phenotype of csn1-10, or its over-accumulation of RGL2, is in agreement with the report that the abi5 mutant can suppress the RGL2-mediated block of germination such as in PAC treatment [16]. These data showed that ABI5 is epistatic to CSN1 in the same pathway during seed germination, and that accumulation of ABI5 is responsible for the germination defects of csn5a-1.
Fig 6. abi5-4 mutant rescued the germination of csn1-10 as well as csn5a-1.

(A and B) Germination of csn1-10 abi5-4 (A) or csn5a-1 abi5-4 (B) double mutants displayed normal rate, in contrast to slow germination of the corresponding single mutant csn1-10 and csn5a-1, respectively. All of the csn1-10 abi5-4 lines including double mutant db-5, wild type (WT), csn1-10, and abi5-4 are segregating sibling lines from the same genetic cross. Likewise, all of the csn5a-1 abi5-4 lines including the double mutant csn5a-1 abi5-4, wild type (WT), csn5a-1, and abi5-4 are segregating sibling lines from the same genetic cross. Error bars represent standard deviation from 4 repeats. (C and D) Anti-RGL2 immunoblots showing that timely removal of RGL2 was restored in csn1-10 abi5-4 double mutant (C) as well as in csn5a-1 abi5-4 double mutant (D). Samples of indicated lines were taken at indicated times after seed imbibition. See also S5 Fig.
Associated with the rescue in germination, time-course immunoblots showed, somewhat surprisingly, that timely removal of RGL2 has also been restored in the double mutants of csn1-10 abi5-4 as well as csn5a-1 abi5-4 following germination (Fig 6C and 6D). Although the mechanism is unclear as to how RGL2 turnover can be rescued in the absence of ABI5, these results are in agreement with previous reports that low ABI5 is associated with germination in low GA conditions (for example treatment of PAC) [16]. Together, our data further reinforce the notion that ABI5 functions downstream of RGL2 in seed germination, and that proper regulation of both germination inhibitors requires CSN5A.
Differential impacts of CSN1 and CSN5A on transcriptomes in dry seeds vs the imbibed seeds
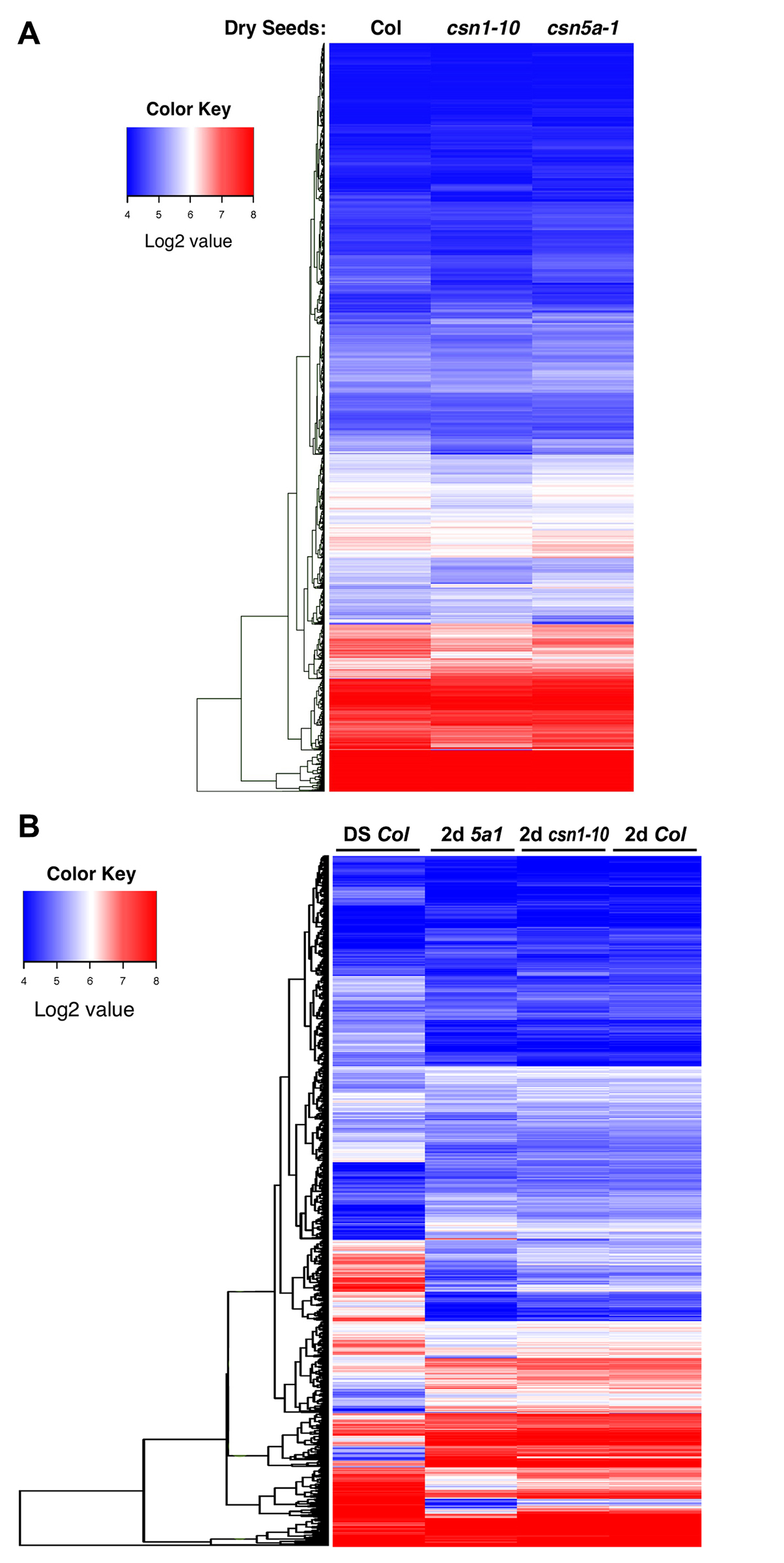
To understand the transcriptional changes in the csn mutants during germination, we conducted a transcriptome analysis on dry seeds and 2-day imbibed seeds of csn5a-1 (or 5a-1) and csn1-10 along with the Col control. We determined the number of genes whose expression was significantly changed (SSTF, statistically significant two-fold change) in csn1-10 or csn5a-1 compared to the Col controls in dry seeds (Fig 7A) or in 2-day imbibed seeds (Fig 7B). csn1-10 affected more genes (919 SSTF genes) than csn5a-1 (644 SSTF genes) in dry seeds, suggesting that CSN1 plays a greater role than CSN5A in seed maturation. However, upon seed imbibition for two days, we observed a robust expansion in the number of genes whose expression was affected by the csn5a-1 mutation (1502 SSTF genes) in comparison to those affected by csn1-10 (640 SSTF genes). This may indicate that CSN5A has more critical functions than CSN1 during germination and the active growth phase of Arabidopsis. This idea is also consistent with the expression value profiling analyses shown in heat-map plots (S6 Fig). In dry seeds, csn1-10 displayed greater abnormalities than csn5a-1 in overall expression profiles when compared to that of Col, but in 2-day imbibed seeds, the profile of csn5a-1 altered from that of Col more than csn1-10 did. Also, csn5a-1 exhibited an expression profile that appeared to have departed further from the dry seeds (S6 Fig). The top GO genes that are down-regulated in Col 2-day imbibed seeds compared to the dry seeds are genes responsive to various stimuli, and approximately the same GO catagory genes were further down-regulated in csn5a-1 at the 2-day timepoint (S1 Table). csn1-10, on the other hand, did not show strongly enriched GO groups at the 2-day timepoint according to the p-values, but in dry seeds, it showed strong misregulation of temperature responsive genes (S1 Table).
Fig 7. Effects of csn1-10 and csn5a-1 mutations on transcriptome and ABA sensitivity.

(A) The diagrams showing the number of SSTF (statistically significant two-fold change) genes in csn5a-1 and csn1-10 mutants compared to wild type Col in dry seeds, or after two-day imbibition. csn1-10 affected more genes than csn5a-1 in dry seeds, while csn5a-1 affected more genes than csn1-10 in 2-day imbibed seeds. The areas of the squares were drawn in proportion to the number of the genes in the indicated category. (B) Real-time qPCR analysis of representative ABA related genes on day2 and day3 imbibed seeds of Col, csn1-10, and csn5a-1. The expression level of each sample was normalized to IPA-like1 (AT1G17210). (C) Sensitivity to ABA-mediated inhibition of germination is enhanced in csn1-10, csn5a-1, and csn5a-1rgl2 double mutant. The csn5a-1abi5-4 double mutant showed resistance to ABA. Cold stratified seeds were used. Error bars represent standard deviation from 4 repeats. (D) ABA biosynthetic inhibitor norflurazon (5μM) rescued the low germination rates of both csn1-10 and csn5a-1, but did not rescue the delayed germination in csn5a-1. Error bars represent standard deviation from 4 repeats. (E) Yeast-two-hybrid sassy showing interactions of ABI5 with CSN5A and CSN5B. AD, activation-domain fusion vectors; BD, DNA-binding domain fusion vectors; EV, empty vector. See also S6 and S7 Figs.
Functions of CSN and CSN5A in ABA regulation of germination
The transcriptome results showed that, from dry seeds to the 2-day time-point, several GA biosynthetic genes were strongly induced in Col, and ABA biosynthetic genes appeared to be active in dry and 2-day imbibed seeds (S7 Fig). However, we were unable to find drastic differences in the mutants. We also examined a panel of genes that have been shown to regulate dormancy and seed germination, but the csn mutants exhibited a similar pattern of expression changes as Col (S7C Fig). Since the seed germination conditions used for transcriptome analysis were not identical to that used for the germination tests, the kinetics of germination could differ slightly. We thus conducted qRT-PCR analysis of several ABA-related genes using the material from our standard germination testing conditions. At day2 and day3 post-imbibition, the ABA biosynthetic gene AAO1 appeared to be expressed at higher levels in csn1-10, while the ABA catabolic gene CYP707A1, whose expression rose following seed imbibition, appeared to be lower in csn5a-1 (Fig 7B). The ABI5 transcript was significantly down-regulated from dry seeds after 2days imbibition in Col as well as in both csn mutants (S7C Fig). On day2 and day3 post seed imbibition, the levels of ABI5 were still moderately higher in both csn mutants compared to Col (Fig 7B), although not as substantial as the difference in its protein accumulation.
We tested the sensitivity of the mutants to exogenous ABA during germination. On ABA-containing plates, both csn5a1 and csn1-10 were more sensitive to ABA than Col (Fig 7C). In addition, the double mutant csn5a-1 rgl2-13 was hyper-sensitive to ABA, whereas csn5a-1 abi5-4 was hypo-sensitive to ABA. In a positive correlation, the mutants that were hypersensitive to ABA also showed abnormal accumulation of ABI5 (Fig 4 and Fig 5E).
We were interested in the causes of the sustained ABI5 accumulation, a key inhibitor that has prevented timely germination of both csn1-10 and csn5a-1 mutants. It has been shown that accumulation of RGL2 during germination can drive ABA biosynthesis, which would induce ABI5 expression and inhibit germination [16]. To investigate whether de novo ABA synthesis is required for the germination phenotypes of the csn mutants, we applied ABA biosynthetic inhibitor norflurazon to test its effect on germination. As shown in Fig 7D, norflurazon treatment nearly completely restored the germination of csn1-10. While it considerably alleviated the germination defects of csn5a-1, norflurazon treatment could not rescue the delayed germination of csn5a-1 (Fig 7D). This result shows that csn1-10-induced hyperdormancy requires ABA synthesis after seed imbibition, which suggest that the extended ABI5 accumulation in csn1-10 likely resulted from RGL2-induced ABA biosynthesis. By contrast, csn5a-1 appeared to cause a defect downstream of ABA synthesis, which would support a role of CSN5A in facilitating ABI5 protein degradation following seed imbibition.
How CSN5A regulates ABI5 protein degradation is unclear. To this end, we found that CSN5A, or CSN5B, can interact with ABI5 in a yeast-two-hybrid assay (Fig 7E). The precise mechanism by which CSN5A regulates ABI5 will be a challenge for future studies. Taken together, we propose a model illustrating the hierarchical functions of the CSN in seed germination (Fig 8). CSN1 and CSN5A, as parts of the CSN complex that regulates SCF ubiquitin E3s, are necessary for timely degradation of DELLA inhibitors, mainly RGL2. RGL2 inhibits germination by inducing ABA synthesis and promoting ABI5 expression, based on previous reports. CSN5A additionally plays a role in the timely removal of ABI5 proteins to facilitate seed germination.
Fig 8. CSN functions in the seed germination pathway.

A model indicating the targets of the CSN and CSN5A in the GA- and ABA- regulated seed germination pathway. The CSN complex (or CSN1) promotes germination by facilitating SCF-mediated degradation of RGL2 DELLA repressor. CSN5A, independently from the CSN complex, is involved in the turnover of ABI5, which acts downstream of RGL2. ABI5 appears to have a role in stabilizing RGL2, while RGL2 has been reported to activate ABI5 expression directly and by inducing ABA synthesis.
Discussion
The COP9 signalosome is known to be necessary for efficient seed germination, but the molecular targets and the mechanism of its action are unclear [34, 39, 43]. We demonstrate here that the role of the CSN in promoting seed germination is by regulating two key inhibitors of seed germination, the DELLA protein RGL2 and the ABA effector ABI5. csn1-10 and csn5a-1 mutants are both poor in germination, but they exhibit subtly different germination phenotypes, and the mechanisms by which CSN1 and CSN5A regulate germination are not identical. csn1-10 is hyper-dormant, while csn5a-1 exhibits both hyperdormancy and delayed germination. CSN1 primarily regulates RGL2 turnover, which in turn affects ABI5 as a result. CSN5A is necessary in timely removal of ABI5 by a mechanism that apparently does not involve CSN1 (or the CSN complex). Thus, our study has identified the specific targets, from two different hormonal pathways, that underlie the germination defects of the csn mutants, and as such has clarified a long-standing mystery.
CSN is required for timely degradation of RGL2 during seed germination
The CSN has been shown to regulate SCF family of ubiquitin E3 ligases. In fact, csn1-10 and csn3-3 were isolated as enhancers of tir1-1, a mutation of the auxin receptor that acts as the F-box component of SCFTIR1 [45, 46]. Thus, the CSN genetically promotes the function of SCFTIR1 in mediating auxin responses. Similarly, a csn1 hypomorphic line showed genetic interactions with F-box proteins UFO and COI1 overexpression lines, supporting the role of the CSN in facilitating the functions of the corresponding SCF E3s [27, 40]. Moreover, csn5a and several lethal csn mutants were shown to abnormally accumulate RGA, a DELLA protein and a substrate of SCFSLY1/2 that has an important role in seedling development [43]. In the same report, the germination defect of the csn mutants were described, but the molecular targets responsible for their germination phenotypes were not determined. In this study, we show that the inability to timely degrade RGL2, a key inhibitor of germination for dormancy response and a substrate of SCFSLY1/2, can fully account for the hyper-dormant phenotype of csn1-10. Considering what is known about the function of the CSN, we suggest that CSN1, or the CSN complex, most likely plays an important role in SCFSLY1/2-mediated RGL2 ubiquitination during the course of seed germination (Fig 8).
CSN5A regulates ABI5 downstream of CSN-facilitated degradation of RGL2 during germination
In Arabidopsis, complete loss of any one of the eight canonical subunits of the CSN leads to the destruction of the complex [32]. This explains why null mutants of any one of the CSN subunits all cause early seedling lethality, with a similar “fusca” phenotype. Probably as a consequence of this similarity, all CSN subunits have largely been indiscriminately viewed to have more or less same physiological functions in plants.
In a number of other species, differences in phenotypes and functions for different subunits of the CSN have been observed [53]. In fungi, deletion mutants of different CSN subunits cause distinct phenotypes [54, 55]. In mammalian cells, knock-down of CSN3 or CSN8 in cultured cells can accelerate cell proliferation [56, 57], whereas knock-down of CSN5 decreases cell proliferation and causes cell senescence [57, 58]. The human CSN5 has been extensively studied due to its association with many types of cancers [59]. Originally isolated in human cells as a coactivator of c-Jun, Jab1 (c-Jun activation domain-binding protein 1) [60], CSN5/JAB1 has been reported to bind to an array of transcription factors or other critical cellular regulators to stabilize the target protein or to facilitate its degradation. As an integral subunit of the CSN complex, CSN5 carries the catalytic center of the CSN deneddylase that is active only when assembled into the complex [61]. It remains unclear how the regulatory activities of CSN5 on its binding partners are related to the role of CSN5 in deneddylation of the CRL E3 ligases. Notably, in a highly conserved manner from yeast to plants and mammals, CSN5 has been found to exist both as part of the holo-CSN complex and in a free form unbound to the CSN [53]. Studies in mammalian systems have shown that CSN5 has functions independent of the CSN holocomplex [58]. Whether the Arabidopsis CSN5, like its mammalian counterpart, has functions apart from that of the CSN complex, has not been reported.
In Arabidopsis, csn1-10 and csn5a-1 both exhibit poor germination. However, careful analyses indicate that the two mutants differ in several aspects. First, csn1-10 shows deeper seed dormancy, such that its germination can be restored by cold stratification or extended after-ripening period, whereas csn5a-1 additionally exhibits a delayed germination that is resistant to these dormancy-breaking measures (Fig 2 and S2 Fig). Second, the csn1-10 germination phenotype can be suppressed by loss of RGL2, while csn5a-1 cannot (Fig 5A and 5B). Third, the responses to GA-synthesis inhibitor PAC and to ABA-synthesis inhibitor norflurazon are different. In particular, norflurazon completely rescues the germination of csn1-10, but cannot rescue the delayed germination of csn5a-1 (Fig 7D). We suggest that the different behaviors between the two mutants can be explained by csn5a-1-specific deficiency in protein degradation of ABI5, which occurs downstream of RGL2 in the germination pathway (Fig 8). PAC treatment has been shown to stabilize RGL2 and stimulate accumulation of ABI5 [16]. That csn5a-1 mutants failed to down-regulate ABI5 protein would render the mutant hypersensitive to PAC. As a consequence, even though rgl2 can confer resistance to PAC in wild type background, csn5a-1 rgl2 double mutant still accumulate ABI5 and consequently remain hypersensitive to PAC. The fact that rgl2 can fully rescue the germination of csn1-10 rgl2 suggests that CSN1 does not play a significant role in direct regulation of ABI5. The prolonged ABI5 accumulation in csn1-10 mutant is most likely caused by its extended accumulation of RGL2, which induce ABA biosynthesis and promote ABI5 expression.
The fourth difference between the csn1-10 and csn5a-1 is implied by their transcriptome profiles, which indicate that the two mutations have differential impacts on plants at different developmental stages. Relative to each other, csn1-10 strongly affected seed maturation, while csn5a-1 strongly affected seed germination and seedling establishment (Fig 7A).
We also found a peculiar behavior of csn5b-1, which has a weaker seed dormancy than Col (Fig 2), opposite to the phenotype of the rest of the csn mutants. Similar phenomena have been reported regarding the root phenotype. It was observed that csn5b-1 has more adventitious roots, opposite to that of other csn mutants which develop fewer (csn1-10 and csn3-3) or no (csn5a-1 and csn5a-2) adventitious roots [42]. It is possible that CSN5B has a different function from CSN5A, which results in the difference of the phenotype. Alternatively, it is possible that csn5b-1 only slightly reduces the activity of CSN to the extent that could not be definitively detected by immunoblotting. Since CSN’s de-neddylation activity biochemically inhibits SCF, the slight reduction of the CSN might increase the SCF activity without creating a hyperactive SCF that auto-ubiquitinates its own components, as in severe csn mutants. This might enhance the SCF activity in targeting RGL2, resulting in the reduced dormancy in csn5b-1. This idea is admittedly highly speculative.
CSN5 has unique functions in ABA signaling
With this study, we have for the first time revealed that CSN5A can directly or indirectly regulate protein stability of the b-ZIP transcription factor ABI5 (Fig 8), a key transcription factor that mediate the response to ABA, especially in seed germination [20, 62]. Elevated ABI5 may attribute to the hypersensitivity of the csn1-10, csn5a-1 and csn5a rgl2 in ABA-mediated inhibition of germination (Fig 7C). It has been reported that ABA receptor is targeted for degradation by the CRL4-CDDD E3 complex [63]. Given that CSN can regulate CRL4, it seems possible that CRL4’s activity in targeting the ABA receptor be compromised in the csn mutants, which might also contribute to the ABA hypersensitivity.
ABI5 transcripts are drastically down-regulated following seed imbibition in the csn mutants (S7C Fig), although the levels are still moderately higher than in the wild type at day2 and day3 after imbibition (Fig 7B). Nonetheless, the elevated level of ABI5 transcripts cannot fully account for the sustained high-level accumulation of its protein in the mutants (Fig 4). Moreover, inhibiting ABA biosynthesis, and thus ABA-induced ABI5 expression, cannot rescue the delayed germination of csn5a-1. These results suggest that the sustained ABI5 accumulation is probably caused by defective ABI5 degradation in csn5a-1, which results in a delay of the germination.
ABI5 protein stability has been shown to be regulated by several factors and ubiquitin E3 ligases, including KEEP ON GOING (KEG) [64], SALT- AND DROUGHT-INDUCIBLE RING FINGER 1 (SDIR1) [65], ABI FIVE BINDING PROTEIN (AFP) [66], CRL4ABD and CRL4DWA1/2 [67] [68]. In these studies, KEG and CRL4s ubiquitin E3 ligases are shown to target ABI5 in post-germination events. Our data suggest that CSN5A facilitates protein degradation of ABI5 during germination, but it is unclear which of those known ABI5 E3 ligases CSN5A works with. We also cannot exclude that there may be a different E3 yet to be identified that targets ABI5 specifically during germination. Furthermore, ABI5 protein undergoes various post-translational modifications including phosphorylation, SUMOylation and S-nitrosylation, all of which can modulate its protein stability [62, 69, 70]. It cannot be ruled out that CSN5A may modulate ABI5 protein stability by affecting those modifications on ABI5.
Our findings raise additional questions as to whether CSN5A may affect other aspects of ABI5 functions or other ABA responses. Regardless, the observations that ABI5 is specifically affected in csn5a but not in csn1 suggest that this function might represent the first CSN5-specific activity in plants. With this function of CSN5A, we can now add ABA responses to the repertoire of CSN/CSN5 regulated pathways.
Materials and methods
Plant material and growth conditions
The Arabidopsis thaliana mutants csn5a-1, csn5a-2, csn5b-1 [35], csn1-10 [45], csn3-3 [46], and rgl2-13 [13] are in the Col ecotype. csn2-5 [47] is in Langsberg eracte (Ler) background. The abi5-4 and csn1-1 (fus6-1) mutants are in Wassilewskija (Ws) ecotype. After the seeds were germinated and grown on GM plates (see below) for 6 to 9 days, the seedlings were transferred to soiled pots. The soil mix was composed of one part Vermiculite and one part Farfard#2 soil mix, which were soak in water with fertilizer (0.25 g/L water). The fertilizer is from Scotts (Peters Professional 20-10-20 #99250 from Scotts). Plants were grown in a walk-in growth room in 22°C long day light cycle.
Germination assays
When not specified, seeds between 1 week to 6 weeks were used and un-stratified in the germination tests or immunoblot analyses. For experiments requiring cold stratification to break the dormancy, such as testing sensitivity to PAC or ABA, seeds from 1 week to up to 6 month were used, and the seeds were cold stratified for 2–3 days regardless of the collecting time. Seeds to be used for direct comparisons were usually collected on the same day from the parental plants grown side-by-side under the same conditions. This was strictly the case for Col and csn1-10. However, since csn5a-1 plants is late–flowering, it was matched with later planted Col that had a same or close seed collection dates. Seeds were surface sterilized with a solution containing 30% bleach and 0.1% TritonX-100, washed. When not specified, seeds are saw on solid growth medium (GM) containing MS salt 4.4 g/L (M0404, Sigma), MES 0.5g/L (M5287, Sigma), 1% sucrose and 0.8% (w/v) Bacto-Agar. The plates were divided to 4 areas to aid in counting and scoring seed germination rates. To determine the germination rates, approximately 20–60 seeds in 4 repeats were examined daily under a Nikon/Zeiss dissecting microscope, and germination were scored based on radicle protrusion from endosperm. The germination rates were measured in DAI (days after imbibition). Seeds to be tested for germination were routinely carried out in pairs: one set of the seeds were un-stratified and the germination rates were presented unless otherwise noted; the other matching set of seeds were cold stratified (4°C) in the dark for 3–4 days, and their germination rates were recorded to make sure that the batch of the seeds were of high quality. Plates were incubated in a Percival growth chamber for 22°C with constant white light.
For PAC, GA, and ABA sensitivity test, cold stratified seeds were used. The medium was supplemented with PAC (paclobutrazol, sc-236284, Santa Cruz Biotechnology), GA3 (G7645 Sigma). For ABA sensitivity teste, solid GM medium described above without sucrose were used that contained ABA (A1049, Sigma-Aldrich) in indicated concentration. For testing of norflurazon (Sigma-Aldrich 34364) effect, unstratified seeds were used on no sucrose GM plates.
For light controlled seed germination experiment, seeds sow on GM plates were cold stratified for two days in the dark, then seeds were exposed to 5 min red light (13.4 watts.m-2) or far-red light (8.2 watts.m-2), or followed by another 5 min of light treatment as indicated. Plates containing the seeds were then wrapped in foil and kept in the dark at 22°C incubator for two days before counting for germination.
Seed coat dissection experiments
After sterilizing and washing, seeds were imbibed in water for 3 hours before dissection using a fine syringe needle according to a previously described procedure [51, 52]. Dissected embryos were placed on a water agar (1%) plate alone with corresponding seed controls, and were incubated in a 22°C growth chamber with constant white light. Embryos and seeds were examined through a Leica dissecting microscope and photographs were taken daily.
Protein extraction, antibodies and immunoblot analysis
For every 100 mg of fresh weight imbibed seeds or germinating seedlings, the sample was mixed with 180 microliter of chilled extraction buffer (50mM Tris-HCL, pH7.5, 150mM NaCl, 10mM MgCl2, 2.5mM EDTA, 1mM DTT, 0.1% Nonidet P-40, and freshly added 1mM protease inhibitors phenylmethylsulfonyl fluoride (PMSF) and 1x complete protease inhibitor cocktail (Roche Molecular Biochemicals) was added and mixture was homogenized. Then, 100 microliter of 4x sample buffer was added and the mixture was vortexed. Samples were then boiled for 10 min and span in a microfuge for 10 min. The supernatant was transferred to a new tube, from which samples were loaded onto SDS-PAGE for immunoblotting.
Antibodies used for this study include anti-CSN5A and anti-CSN3 [35], anti-CUL1 [48], anti-RPN6 [71], anti-RGL2 [72], and anti-ABI5 (Abcam, ab98831). The anti-CSN5B polyclonal antibodies were made by Beijing Protein Innovation Co., Ltd. (BPI). Briefly, an EcoRI/XhoI fragment containing the full-length CSN5B open reading frame was cloned into the EcoRI/XhoI sites of the pET-28a vector, so as to express 6×His-tagged CSN5B protein. The fusion protein was expressed in Escherichia coli, then purified and used as antigen to immunize rabbits for the production of polyclonal antiserum. Antigen affinity purified anti- CSN5B antibodies were used in immunoblots.
Transcriptome analyses by RNA-seq
Col-0, csn1-10, and csn5a-1 seeds were collected from plants grown side-by-side in the growth room. Seeds for Col-0 and csn1-10 were 1.5wks in storage, and csn5a-1 seeds were 4wk in storage. Approximately 100 microliter of settled seeds were used for RNA extraction for each sample. The 2-day imbibed seeds, prepared with the same volume of dry seeds as above, were sterilized, washed, and incubate on cell culture wells with 1ml of 0.5X liquid MS medium at 22°C under constant light for 2 days. Each sample points had three biological repeats. Seeds were centrifuged to remove the liquid, and were frozen in liquid nitrogen. The frozen seeds were ground to a fine powder using mortar and pestle in the presence of liquid nitrogen and small quantity of sterile quartz powder. RNA extraction was performed according to a published procedure [73].
High-throughput RNA-seq was carried out at Yale Center for Genome Analysis. The Single-End RNA-sequencing was carried out with Illumina Hi-seq 2000 platform (Genome Center, Yale West Campus). Specifically, libraries were analyzed with a Bioanalyzer 2100 instrument (Agilent, Santa Clara, CA), quantified by Qubit fluorometer (Life Technologies, Carlsbad, CA). The Arabidopsis thaliana genome obtained from TAIR10 (https://www.arabidopsis.org) was used as the genome reference. After adaptor trimming and contaminate sequence removing by fastqc (www.bioinformatics.babraham.ac.uk/projects/fastqc/) and fastx-toolkits (hannonlab.cshl.edu/fastx_toolkit/), Bowtie2 (http://bowtie-bio.sourceforge.net/bowtie2/index.shtml) was used for genome mapping and followed by the tophat (https://ccb.jhu.edu/software/tophat/index.shtml) transcript assembling. Gene differentially expression profiling was accomplished by cufflink and cuffdiff software package (cole-trapnell-lab.github.io/cufflinks/cuffdiff/) with default parameters and cutoffs, fold change cutoff was set to 2. The data set is accessible at NCBI GEO under accession number GSE106223.
qRT-PCR
Col, csn5a-1 and csn1-10 seeds (5-days in storage) were sterilized and sow on the solid medium plate as described above in Germination assay. Total RNAs were extracted from 2- or 3- day germinating Col, csn5a-1 and csn1-10 seeds following the procedures as described previously [73]. Then 1 μg of total RNA was used for reverse transcription reaction using SuperScript III Reverse Transcriptase (Invitrogen) and quantitative PCR (qPCR) reaction was performed in Bio-Rad CFX96 real-time system using iQ SYBR Green mix (Bio-Rad). Gene expression was normalized to IPA-like1 (AT1G17210).
Yeast-two-hybrid assay
Full-length cDNA coding region of CSN5A and CSN5B were each subcloned into the EcoRI/SalI and EcoRI/XhoI sites of pB42AD(AD) vector (Clontech). ABI5 was subcloned into the EcoRI/XhoI site of pLexA (BD) vector (Clontech). Yeast two-hybrid assay was performed according to the Matchmaker LexA Two-Hybrid System manual (Clontech, K1609-1). Briefly, all constructs were co-transformed into yeast strain EGY48 containing p8op-LacZ. Transformants were grown on SD/-His/-Trp/-Ura plates containing X-Gal for blue color development.
Supporting information
(DOCX)
(DOCX)
(DOCX)
(A) Adult phenotypes of 5-weeks csn mutant plants, compared to the wild type plants of their corresponding ecotype backgrounds. csn5a-1, csn5a-2, csn5b-1, csn1-10, csn3-3 are in Col background, while csn2-5 is in Ler background. (B) Expression of CSN subunits in various tissues of Arabidopsis in wild type and csn5 mutants. Tissues were collected from wild type (Col-0) or csn5a2 or csn5b1 mutants as indicated. Total proteins were analyzed by immunoblotting using antibodies against CSN5A, CSN1, and CSN8. Anti-RPN6 blots on the same samples were used as an internal reference. csn5b-1 mutation had no detectable effect on the level of CSN subunits in the tissues examined. csn5a-2 mutation resulted in notable reduction in CSN5A level, and had minor effect on CSN1 and CSN8 levels.
(JPG)
{kind=link}
(A) Col, csn1-10 and csn5a-1 seeds (4-day after collection) were tested for germination on solid growth medium containing 1% sucrose (left panels), or had no sucrose (right two panels). Seeds were not stratified (top panels) or cold stratified for 4 days (bottom panels). The slow germination phenotype was clearly displayed on both sucrose-containing or sucrose-less plates. (B) Col and mutant seeds of indicated storage age were cold stratified for 4 days before the germination test. csn5a-1 showed delayed germination even after cold stratification of fully after-ripened seeds.
(JPG)
{kind=link}
(A) The diagram illustrating the light treatment procedure used to test phyB-controlled seed germination. Seeds were cold stratified for 2 days prior to the light treatment. After light treatments, seeds were incubated in the dark at 22oC for two or three days. WL, constant white light; R, red light 5 min; FR, far-red light 5min; FR-R, far-red light 5min followed by red light 5 min; FR-R-FR, far-red light 5min followed by red light 5 min and followed by far-red light 5 min. (B) The germination rates at day-2 (or day-3 for csn5a-1) post light treatments are shown. The color code in (B) is identical to the colors indicating light treatment in (A). The csn mutants exhibited largely normal light responsive seed germination, while det1-1 (as a control) showed light-independent seed germination. Error bars represent standard deviation from 4 repeats.
(JPG)
{kind=link}
The graph shows the germination rates at day-10 and day-12 of two segregating sibling lines of csn1-1 and double mutant csn1-1 rgl2-13. Error bars represent standard deviation from 4 repeats.
(JPG)
{kind=link}
In the right panel, the csn5a-2 abi5-1 double mutants showed improved germination rates over csn5a-2. These mutant lines were segregating sibling lines from the same cross. Germination rate of one representative csn5a-2abi5-1 double and csn5a-2 single mutant lines were counted and graphed as shown on the left. Error bars represent standard deviation from 4 repeats.
(JPG)
{kind=link}
(A) Heatmap diagram of mRNA profiles in dry seeds of Col wild type, csn1-10, and csn5a-1. Genes with expression values from 13 and above were included. The color code is set from 13–300 in Log2 scale. Genes with expression level above 300 show the same color as those of 300 (red). The transcriptome profile of csn1-10 in dry seeds show greater dissimilarity to wild type than comparing csn5a-1 to wild type. (B) Heatmap plot showing gene expression profile in 2-day imbibed seeds of csn5a-1, csn1-10, and Col, compared to Col dry seeds. Genes with the expression value from 13 and above were included. The color code is set from 13 to 300 in Log2 scale. Genes with expression values above 300 are indicated with the same color as those of 300 (red). csn5a-1 displayed greater transcriptome changes than csn1-10 did at 2-day post imbibition.
(JPG)
{kind=link}
(A and B) Heatmap showing ABA-related genes (A) and GA related genes (B) in 2-day imbibed seeds of csn5a-1 (5a-1), csn1-10, and Col, compared to those of Col dry seeds. (C) A panel of known seed germination regulatory genes showed similar expression profiles in csn5a-1, csn1-10 and Col. These genes are: DAG1 (DOF AFFECTING GERMINATION 1 AT3G61850), DOG1 (DELAY OF GERMINATION 1, AT5G45830), SPY SPINDLY, AT3G11540), SPT SPATULA, AT4G36930), LDL1/2 (ARABIDOPSIS LYSINE-SPECIFIC HISTONE DEMETHYLAS AT1G62830/ AT3G13682), MFT (MOTHER OF FT AND TFL, AT1G18100), RGL2 (RGA-LIKE 2, AT3G03450), RDO5 (REDUCED DORMANCY 5, AT4G11040), ABI5 ABA INSENSITIVE 5, AT2G36270).
(JPG)
{kind=link}
Acknowledgments
We thank Dr. W. M. Gray for providing the csn1-10 and csn3-3 mutants, Dr. J. Stuttmann for csn2-5, and Dr. T. P. Sun for rgl2-13.
Data Availability
The data is accessible to the public at the NCBI site with the accession number GSE106223 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE106223).
Funding Statement
This work was supported in part by grants from National Institute of Health (R01GM047850) to NW and from United States – Israel Binational Science Foundation (Grant# 2013071) to NW, from the National Natural Science Foundation of China (31770321 and 31371221) to JL, and from Special Fund for Agro-Scientific Research in the Public Interest Grant(201303002), Ministry of Agriculture of China and The National Natural Science Foundation of China (31171564) to DK. DJ was supported by Fundamental Research Funds for the Central Universities from China (XDJK2017B001). MW was supported by China Scholarship Council and Jilin Business and Technology College. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Holdsworth MJ, Bentsink L, Soppe WJ. Molecular networks regulating Arabidopsis seed maturation, after-ripening, dormancy and germination. New Phytol. 2008;179(1):33–54. doi: 10.1111/j.1469-8137.2008.02437.x . [DOI] [PubMed] [Google Scholar]
- 2.Finch-Savage WE, Leubner-Metzger G. Seed dormancy and the control of germination. New Phytol. 2006;171(3):501–23. doi: 10.1111/j.1469-8137.2006.01787.x . [DOI] [PubMed] [Google Scholar]
- 3.Finkelstein R, Reeves W, Ariizumi T, Steber C. Molecular aspects of seed dormancy. Annu Rev Plant Biol. 2008;59:387–415. doi: 10.1146/annurev.arplant.59.032607.092740 . [DOI] [PubMed] [Google Scholar]
- 4.Koornneef M, van der Veen JH. Induction and analysis of gibberellin sensitive mutants in Arabidopsis thaliana (L.) heynh. Theor Appl Genet. 1980;58(6):257–63. doi: 10.1007/BF00265176 . [DOI] [PubMed] [Google Scholar]
- 5.Debeaujon I, Koornneef M. Gibberellin requirement for Arabidopsis seed germination is determined both by testa characteristics and embryonic abscisic acid. Plant Physiol. 2000;122(2):415–24. ; PubMed Central PMCID: PMCPMC58878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Seo M, Hanada A, Kuwahara A, Endo A, Okamoto M, Yamauchi Y, et al. Regulation of hormone metabolism in Arabidopsis seeds: phytochrome regulation of abscisic acid metabolism and abscisic acid regulation of gibberellin metabolism. Plant J. 2006;48(3):354–66. doi: 10.1111/j.1365-313X.2006.02881.x . [DOI] [PubMed] [Google Scholar]
- 7.Yamauchi Y, Ogawa M, Kuwahara A, Hanada A, Kamiya Y, Yamaguchi S. Activation of gibberellin biosynthesis and response pathways by low temperature during imbibition of Arabidopsis thaliana seeds. Plant Cell. 2004;16(2):367–78. doi: 10.1105/tpc.018143 ; PubMed Central PMCID: PMCPMC341910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Yan D, Easwaran V, Chau V, Okamoto M, Ierullo M, Kimura M, et al. NIN-like protein 8 is a master regulator of nitrate-promoted seed germination in Arabidopsis. Nat Commun. 2016;7:13179 doi: 10.1038/ncomms13179 ; PubMed Central PMCID: PMCPMC5064020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hauvermale AL, Ariizumi T, Steber CM. Gibberellin signaling: a theme and variations on DELLA repression. Plant Physiol. 2012;160(1):83–92. doi: 10.1104/pp.112.200956 ; PubMed Central PMCID: PMCPMC3440232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fu X, Richards DE, Fleck B, Xie D, Burton N, Harberd NP. The Arabidopsis mutant sleepy1gar2-1 protein promotes plant growth by increasing the affinity of the SCFSLY1 E3 ubiquitin ligase for DELLA protein substrates. Plant Cell. 2004;16(6):1406–18. doi: 10.1105/tpc.021386 ; PubMed Central PMCID: PMCPMC490035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Dill A, Thomas SG, Hu J, Steber CM, Sun TP. The Arabidopsis F-box protein SLEEPY1 targets gibberellin signaling repressors for gibberellin-induced degradation. Plant Cell. 2004;16(6):1392–405. doi: 10.1105/tpc.020958 ; PubMed Central PMCID: PMCPMC490034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lee S, Cheng H, King KE, Wang W, He Y, Hussain A, et al. Gibberellin regulates Arabidopsis seed germination via RGL2, a GAI/RGA-like gene whose expression is up-regulated following imbibition. Genes Dev. 2002;16(5):646–58. doi: 10.1101/gad.969002 ; PubMed Central PMCID: PMCPMC155355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Tyler L, Thomas SG, Hu J, Dill A, Alonso JM, Ecker JR, et al. Della proteins and gibberellin-regulated seed germination and floral development in Arabidopsis. Plant Physiol. 2004;135(2):1008–19. doi: 10.1104/pp.104.039578 ; PubMed Central PMCID: PMCPMC514135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Oh E, Kim J, Park E, Kim JI, Kang C, Choi G. PIL5, a phytochrome-interacting basic helix-loop-helix protein, is a key negative regulator of seed germination in Arabidopsis thaliana. Plant Cell. 2004;16(11):3045–58. doi: 10.1105/tpc.104.025163 ; PubMed Central PMCID: PMCPMC527197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Boccaccini A, Santopolo S, Capauto D, Lorrai R, Minutello E, Belcram K, et al. Independent and interactive effects of DOF affecting germination 1 (DAG1) and the Della proteins GA insensitive (GAI) and Repressor of ga1-3 (RGA) in embryo development and seed germination. BMC Plant Biol. 2014;14:200 doi: 10.1186/s12870-014-0200-z ; PubMed Central PMCID: PMCPMC4222566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Piskurewicz U, Jikumaru Y, Kinoshita N, Nambara E, Kamiya Y, Lopez-Molina L. The gibberellic acid signaling repressor RGL2 inhibits Arabidopsis seed germination by stimulating abscisic acid synthesis and ABI5 activity. Plant Cell. 2008;20(10):2729–45. doi: 10.1105/tpc.108.061515 ; PubMed Central PMCID: PMCPMC2590721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Liu X, Hu P, Huang M, Tang Y, Li Y, Li L, et al. The NF-YC-RGL2 module integrates GA and ABA signalling to regulate seed germination in Arabidopsis. Nat Commun. 2016;7:12768 doi: 10.1038/ncomms12768 ; PubMed Central PMCID: PMCPMC5027291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lee KP, Piskurewicz U, Tureckova V, Carat S, Chappuis R, Strnad M, et al. Spatially and genetically distinct control of seed germination by phytochromes A and B. Genes Dev. 2012;26(17):1984–96. doi: 10.1101/gad.194266.112 ; PubMed Central PMCID: PMCPMC3435500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Finkelstein RR, Lynch TJ. The Arabidopsis abscisic acid response gene ABI5 encodes a basic leucine zipper transcription factor. Plant Cell. 2000;12(4):599–609. ; PubMed Central PMCID: PMCPMC139856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lopez-Molina L, Mongrand S, Chua NH. A postgermination developmental arrest checkpoint is mediated by abscisic acid and requires the ABI5 transcription factor in Arabidopsis. Proc Natl Acad Sci U S A. 2001;98(8):4782–7. doi: 10.1073/pnas.081594298 ; PubMed Central PMCID: PMCPMC31911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wei N, Deng XW. The COP9 signalosome. Annu Rev Cell Dev Biol. 2003;19:261–86. doi: 10.1146/annurev.cellbio.19.111301.112449 . [DOI] [PubMed] [Google Scholar]
- 22.Schwechheimer C, Serino G, Callis J, Crosby WL, Lyapina S, Deshaies RJ, et al. Interactions of the COP9 signalosome with the E3 ubiquitin ligase SCFTIRI in mediating auxin response. Science. 2001;292(5520):1379–82. doi: 10.1126/science.1059776 . [DOI] [PubMed] [Google Scholar]
- 23.Lyapina S, Cope G, Shevchenko A, Serino G, Tsuge T, Zhou C, et al. Promotion of NEDD-CUL1 conjugate cleavage by COP9 signalosome. Science. 2001;292(5520):1382–5. doi: 10.1126/science.1059780 . [DOI] [PubMed] [Google Scholar]
- 24.Yang X, Menon S, Lykke-Andersen K, Tsuge T, Di X, Wang X, et al. The COP9 signalosome inhibits p27(kip1) degradation and impedes G1-S phase progression via deneddylation of SCF Cul1. Curr Biol. 2002;12(8):667–72. . [DOI] [PubMed] [Google Scholar]
- 25.Cavadini S, Fischer ES, Bunker RD, Potenza A, Lingaraju GM, Goldie KN, et al. Cullin-RING ubiquitin E3 ligase regulation by the COP9 signalosome. Nature. 2016;531(7596):598–603. doi: 10.1038/nature17416 . [DOI] [PubMed] [Google Scholar]
- 26.Emberley ED, Mosadeghi R, Deshaies RJ. Deconjugation of Nedd8 from Cul1 is directly regulated by Skp1-F-box and substrate, and the COP9 signalosome inhibits deneddylated SCF by a noncatalytic mechanism. J Biol Chem. 2012;287(35):29679–89. doi: 10.1074/jbc.M112.352484 ; PubMed Central PMCID: PMCPMC3436198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wang X, Feng S, Nakayama N, Crosby WL, Irish V, Deng XW, et al. The COP9 signalosome interacts with SCF UFO and participates in Arabidopsis flower development. Plant Cell. 2003;15(5):1071–82. doi: 10.1105/tpc.009936 ; PubMed Central PMCID: PMCPMC153717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wee S, Geyer RK, Toda T, Wolf DA. CSN facilitates Cullin-RING ubiquitin ligase function by counteracting autocatalytic adapter instability. Nat Cell Biol. 2005;7(4):387–91. doi: 10.1038/ncb1241 . [DOI] [PubMed] [Google Scholar]
- 29.Hetfeld BK, Helfrich A, Kapelari B, Scheel H, Hofmann K, Guterman A, et al. The zinc finger of the CSN-associated deubiquitinating enzyme USP15 is essential to rescue the E3 ligase Rbx1. Curr Biol. 2005;15(13):1217–21. doi: 10.1016/j.cub.2005.05.059 . [DOI] [PubMed] [Google Scholar]
- 30.Menon S, Chi H, Zhang H, Deng XW, Flavell RA, Wei N. COP9 signalosome subunit 8 is essential for peripheral T cell homeostasis and antigen receptor-induced entry into the cell cycle from quiescence. Nat Immunol. 2007;8(11):1236–45. doi: 10.1038/ni1514 . [DOI] [PubMed] [Google Scholar]
- 31.Schlierf A, Altmann E, Quancard J, Jefferson AB, Assenberg R, Renatus M, et al. Targeted inhibition of the COP9 signalosome for treatment of cancer. Nat Commun. 2016;7:13166 doi: 10.1038/ncomms13166 ; PubMed Central PMCID: PMCPMC5078989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Wei N, Chamovitz DA, Deng XW. Arabidopsis COP9 is a component of a novel signaling complex mediating light control of development. Cell. 1994;78(1):117–24. . [DOI] [PubMed] [Google Scholar]
- 33.Wei N, Deng XW. COP9: a new genetic locus involved in light-regulated development and gene expression in arabidopsis. Plant Cell. 1992;4(12):1507–18. doi: 10.1105/tpc.4.12.1507 ; PubMed Central PMCID: PMCPMC160237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Wei N, Kwok SF, von Arnim AG, Lee A, McNellis TW, Piekos B, et al. Arabidopsis COP8, COP10, and COP11 genes are involved in repression of photomorphogenic development in darkness. Plant Cell. 1994;6(5):629–43. doi: 10.1105/tpc.6.5.629 ; PubMed Central PMCID: PMCPMC160464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Gusmaroli G, Figueroa P, Serino G, Deng XW. Role of the MPN subunits in COP9 signalosome assembly and activity, and their regulatory interaction with Arabidopsis Cullin3-based E3 ligases. Plant Cell. 2007;19(2):564–81. Epub 2007/02/20. doi: 10.1105/tpc.106.047571 ; PubMed Central PMCID: PMC1867349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Jin D, Li B, Deng XW, Wei N. Plant COP9 signalosome subunit 5, CSN5. Plant Sci. 2014;224:54–61. doi: 10.1016/j.plantsci.2014.04.001 . [DOI] [PubMed] [Google Scholar]
- 37.Dohmann EMN, Kuhnle C, Schwechheimer C. Loss of the CONSTITUTIVE PHOTOMORPHOGENIC9 signalosome subunit 5 is sufficient to cause the cop/det/fus mutant phenotype in Arabidopsis. Plant Cell. 2005;17(7):1967–78. doi: 10.1105/tpc.105.032870. ISI:000230165400010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Franciosini A, Lombardi B, Iafrate S, Pecce V, Mele G, Lupacchini L, et al. The Arabidopsis COP9 SIGNALOSOME INTERACTING F-BOX KELCH 1 protein forms an SCF ubiquitin ligase and regulates hypocotyl elongation. Molecular plant. 2013;6(5):1616–29. doi: 10.1093/mp/sst045 . [DOI] [PubMed] [Google Scholar]
- 39.Franciosini A, Moubayidin L, Du K, Matari NH, Boccaccini A, Butera S, et al. The COP9 SIGNALOSOME Is Required for Postembryonic Meristem Maintenance in Arabidopsis thaliana. Molecular plant. 2015;8(11):1623–34. doi: 10.1016/j.molp.2015.08.003 . [DOI] [PubMed] [Google Scholar]
- 40.Feng S, Ma L, Wang X, Xie D, Dinesh-Kumar SP, Wei N, et al. The COP9 signalosome interacts physically with SCF COI1 and modulates jasmonate responses. Plant Cell. 2003;15(5):1083–94. doi: 10.1105/tpc.010207 ; PubMed Central PMCID: PMCPMC153718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Dohmann EM, Levesque MP, De Veylder L, Reichardt I, Jurgens G, Schmid M, et al. The Arabidopsis COP9 signalosome is essential for G2 phase progression and genomic stability. Development. 2008;135(11):2013–22. doi: 10.1242/dev.020743 . [DOI] [PubMed] [Google Scholar]
- 42.Pacurar DI, Pacurar ML, Lakehal A, Pacurar AM, Ranjan A, Bellini C. Erratum: The Arabidopsis Cop9 signalosome subunit 4 (CSN4) is involved in adventitious root formation. Scientific reports. 2017;7(1):6435 doi: 10.1038/s41598-017-04861-9 ; PubMed Central PMCID: PMCPMC5519674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Dohmann EM, Nill C, Schwechheimer C. DELLA proteins restrain germination and elongation growth in Arabidopsis thaliana COP9 signalosome mutants. Eur J Cell Biol. 2010;89(2–3):163–8. doi: 10.1016/j.ejcb.2009.12.001 . [DOI] [PubMed] [Google Scholar]
- 44.Gusmaroli G, Feng SH, Deng XW. The Arabidopsis CSN5A and CSN5B subunits are present in distinct COP9 signalosome complexes, and mutations in their JAMM domains exhibit differential dominant negative effects on development. Plant Cell. 2004;16(11):2984–3001. ISI:000225228500011. doi: 10.1105/tpc.104.025999 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zhang W, Ito H, Quint M, Huang H, Noel LD, Gray WM. Genetic analysis of CAND1-CUL1 interactions in Arabidopsis supports a role for CAND1-mediated cycling of the SCFTIR1 complex. Proc Natl Acad Sci U S A. 2008;105(24):8470–5. doi: 10.1073/pnas.0804144105 ; PubMed Central PMCID: PMCPMC2448860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Huang H, Quint M, Gray WM. The eta7/csn3-3 auxin response mutant of Arabidopsis defines a novel function for the CSN3 subunit of the COP9 signalosome. Plos One. 2013;8(6):e66578 doi: 10.1371/journal.pone.0066578 ; PubMed Central PMCID: PMCPMC3676356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Stuttmann J, Lechner E, Guerois R, Parker JE, Nussaume L, Genschik P, et al. COP9 signalosome- and 26S proteasome-dependent regulation of SCFTIR1 accumulation in Arabidopsis. J Biol Chem. 2009;284(12):7920–30. Epub 2009/01/17. doi: 10.1074/jbc.M809069200 ; PubMed Central PMCID: PMC2658085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Wang X, Kang D, Feng S, Serino G, Schwechheimer C, Wei N. CSN1 N-terminal-dependent activity is required for Arabidopsis development but not for Rub1/Nedd8 deconjugation of cullins: a structure-function study of CSN1 subunit of COP9 signalosome. Mol Biol Cell. 2002;13(2):646–55. doi: 10.1091/mbc.01-08-0427 ; PubMed Central PMCID: PMCPMC65656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Shi H, Wang X, Mo X, Tang C, Zhong S, Deng XW. Arabidopsis DET1 degrades HFR1 but stabilizes PIF1 to precisely regulate seed germination. Proc Natl Acad Sci U S A. 2015;112(12):3817–22. doi: 10.1073/pnas.1502405112 ; PubMed Central PMCID: PMCPMC4378405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Penfield S, Li Y, Gilday AD, Graham S, Graham IA. Arabidopsis ABA INSENSITIVE4 regulates lipid mobilization in the embryo and reveals repression of seed germination by the endosperm. Plant Cell. 2006;18(8):1887–99. doi: 10.1105/tpc.106.041277 ; PubMed Central PMCID: PMCPMC1533976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lee KP, Piskurewicz U, Tureckova V, Strnad M, Lopez-Molina L. A seed coat bedding assay shows that RGL2-dependent release of abscisic acid by the endosperm controls embryo growth in Arabidopsis dormant seeds. Proc Natl Acad Sci U S A. 2010;107(44):19108–13. doi: 10.1073/pnas.1012896107 ; PubMed Central PMCID: PMCPMC2973907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Lee KP, Lopez-Molina L. A seed coat bedding assay to genetically explore in vitro how the endosperm controls seed germination in Arabidopsis thaliana. Journal of visualized experiments: JoVE. 2013;(81):e50732 doi: 10.3791/50732 ; PubMed Central PMCID: PMCPMC3989494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Wei N, Serino G, Deng XW. The COP9 signalosome: more than a protease. Trends Biochem Sci. 2008;33(12):592–600. doi: 10.1016/j.tibs.2008.09.004. ISI:000261836700005. [DOI] [PubMed] [Google Scholar]
- 54.Mundt KE, Liu C, Carr AM. Deletion mutants in COP9/signalosome subunits in fission yeast Schizosaccharomyces pombe display distinct phenotypes. Mol Biol Cell. 2002;13(2):493–502. doi: 10.1091/mbc.01-10-0521. ISI:000174036700010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Wang J, Hu Q, Chen H, Zhou Z, Li W, Wang Y, et al. Role of individual subunits of the Neurospora crassa CSN complex in regulation of deneddylation and stability of cullin proteins. Plos Genet. 2010;6(12):e1001232 doi: 10.1371/journal.pgen.1001232 ; PubMed Central PMCID: PMCPMC2996332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Yoneda-Kato N, Tomoda K, Umehara M, Arata Y, Kato J. Myeloid leukemia factor 1 regulates p53 by suppressing COP1 via COP9 signalosome subunit 3. Embo J. 2005;24(9):1739–49. doi: 10.1038/sj.emboj.7600656. ISI:000228944200010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Liu C, Guo LQ, Menon S, Jin D, Pick E, Wang X, et al. COP9 signalosome subunit Csn8 is involved in maintaining proper duration of the G1 phase. J Biol Chem. 2013;288(28):20443–52. doi: 10.1074/jbc.M113.468959 ; PubMed Central PMCID: PMCPMC3711310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Yoshida A, Yoneda-Kato N, Kato JY. CSN5 specifically interacts with CDK2 and controls senescence in a cytoplasmic cyclin E-mediated manner. Scientific reports. 2013;3:1054 Epub 2013/01/15. doi: 10.1038/srep01054 ; PubMed Central PMCID: PMC3542532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Shackleford TJ, Claret FX. JAB1/CSN5: a new player in cell cycle control and cancer. Cell Div. 2010;5:26 Epub 2010/10/20. doi: 10.1186/1747-1028-5-26 ; PubMed Central PMCID: PMC2976740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Claret FX, Hibi M, Dhut S, Toda T, Karin M. A new group of conserved coactivators that increase the specificity of AP-1 transcription factors. Nature. 1996;383(6599):453–7. Epub 1996/10/03. doi: 10.1038/383453a0 . [DOI] [PubMed] [Google Scholar]
- 61.Cope GA, Suh GS, Aravind L, Schwarz SE, Zipursky SL, Koonin EV, et al. Role of predicted metalloprotease motif of Jab1/Csn5 in cleavage of Nedd8 from Cul1. Science. 2002;298(5593):608–11. Epub 2002/08/17. doi: 10.1126/science.1075901 . [DOI] [PubMed] [Google Scholar]
- 62.Yu F, Wu Y, Xie Q. Precise protein post-translational modifications modulate ABI5 activity. Trends Plant Sci. 2015;20(9):569–75. doi: 10.1016/j.tplants.2015.05.004 . [DOI] [PubMed] [Google Scholar]
- 63.Irigoyen ML, Iniesto E, Rodriguez L, Puga MI, Yanagawa Y, Pick E, et al. Targeted degradation of abscisic acid receptors is mediated by the ubiquitin ligase substrate adaptor DDA1 in Arabidopsis. Plant Cell. 2014;26(2):712–28. doi: 10.1105/tpc.113.122234 ; PubMed Central PMCID: PMCPMC3967035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Stone SL, Williams LA, Farmer LM, Vierstra RD, Callis J. KEEP ON GOING, a RING E3 ligase essential for Arabidopsis growth and development, is involved in abscisic acid signaling. Plant Cell. 2006;18(12):3415–28. doi: 10.1105/tpc.106.046532 ; PubMed Central PMCID: PMCPMC1785414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Zhang Y, Yang C, Li Y, Zheng N, Chen H, Zhao Q, et al. SDIR1 is a RING finger E3 ligase that positively regulates stress-responsive abscisic acid signaling in Arabidopsis. Plant Cell. 2007;19(6):1912–29. doi: 10.1105/tpc.106.048488 ; PubMed Central PMCID: PMCPMC1955734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Lopez-Molina L, Mongrand S, Kinoshita N, Chua NH. AFP is a novel negative regulator of ABA signaling that promotes ABI5 protein degradation. Genes Dev. 2003;17(3):410–8. doi: 10.1101/gad.1055803 ; PubMed Central PMCID: PMCPMC195991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Lee JH, Yoon HJ, Terzaghi W, Martinez C, Dai M, Li J, et al. DWA1 and DWA2, two Arabidopsis DWD protein components of CUL4-based E3 ligases, act together as negative regulators in ABA signal transduction. Plant Cell. 2010;22(6):1716–32. doi: 10.1105/tpc.109.073783 ; PubMed Central PMCID: PMCPMC2910972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Seo KI, Lee JH, Nezames CD, Zhong S, Song E, Byun MO, et al. ABD1 is an Arabidopsis DCAF substrate receptor for CUL4-DDB1-based E3 ligases that acts as a negative regulator of abscisic acid signaling. Plant Cell. 2014;26(2):695–711. doi: 10.1105/tpc.113.119974 ; PubMed Central PMCID: PMCPMC3967034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Albertos P, Romero-Puertas MC, Tatematsu K, Mateos I, Sanchez-Vicente I, Nambara E, et al. S-nitrosylation triggers ABI5 degradation to promote seed germination and seedling growth. Nat Commun. 2015;6:8669 doi: 10.1038/ncomms9669 ; PubMed Central PMCID: PMCPMC4639896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Skelly MJ, Frungillo L, Spoel SH. Transcriptional regulation by complex interplay between post-translational modifications. Curr Opin Plant Biol. 2016;33:126–32. doi: 10.1016/j.pbi.2016.07.004 . [DOI] [PubMed] [Google Scholar]
- 71.Kwok SF, Staub JM, Deng XW. Characterization of two subunits of Arabidopsis 19S proteasome regulatory complex and its possible interaction with the COP9 complex. J Mol Biol. 1999;285(1):85–95. doi: 10.1006/jmbi.1998.2315 . [DOI] [PubMed] [Google Scholar]
- 72.Feng S, Martinez C, Gusmaroli G, Wang Y, Zhou J, Wang F, et al. Coordinated regulation of Arabidopsis thaliana development by light and gibberellins. Nature. 2008;451(7177):475–9. doi: 10.1038/nature06448 ; PubMed Central PMCID: PMCPMC2562044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Vicient CM, Delseny M. Isolation of total RNA from Arabidopsis thaliana seeds. Anal Biochem. 1999;268(2):412–3. doi: 10.1006/abio.1998.3045 . [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
(DOCX)
(DOCX)
(A) Adult phenotypes of 5-weeks csn mutant plants, compared to the wild type plants of their corresponding ecotype backgrounds. csn5a-1, csn5a-2, csn5b-1, csn1-10, csn3-3 are in Col background, while csn2-5 is in Ler background. (B) Expression of CSN subunits in various tissues of Arabidopsis in wild type and csn5 mutants. Tissues were collected from wild type (Col-0) or csn5a2 or csn5b1 mutants as indicated. Total proteins were analyzed by immunoblotting using antibodies against CSN5A, CSN1, and CSN8. Anti-RPN6 blots on the same samples were used as an internal reference. csn5b-1 mutation had no detectable effect on the level of CSN subunits in the tissues examined. csn5a-2 mutation resulted in notable reduction in CSN5A level, and had minor effect on CSN1 and CSN8 levels.
(JPG)
(A) Col, csn1-10 and csn5a-1 seeds (4-day after collection) were tested for germination on solid growth medium containing 1% sucrose (left panels), or had no sucrose (right two panels). Seeds were not stratified (top panels) or cold stratified for 4 days (bottom panels). The slow germination phenotype was clearly displayed on both sucrose-containing or sucrose-less plates. (B) Col and mutant seeds of indicated storage age were cold stratified for 4 days before the germination test. csn5a-1 showed delayed germination even after cold stratification of fully after-ripened seeds.
(JPG)
(A) The diagram illustrating the light treatment procedure used to test phyB-controlled seed germination. Seeds were cold stratified for 2 days prior to the light treatment. After light treatments, seeds were incubated in the dark at 22oC for two or three days. WL, constant white light; R, red light 5 min; FR, far-red light 5min; FR-R, far-red light 5min followed by red light 5 min; FR-R-FR, far-red light 5min followed by red light 5 min and followed by far-red light 5 min. (B) The germination rates at day-2 (or day-3 for csn5a-1) post light treatments are shown. The color code in (B) is identical to the colors indicating light treatment in (A). The csn mutants exhibited largely normal light responsive seed germination, while det1-1 (as a control) showed light-independent seed germination. Error bars represent standard deviation from 4 repeats.
(JPG)
The graph shows the germination rates at day-10 and day-12 of two segregating sibling lines of csn1-1 and double mutant csn1-1 rgl2-13. Error bars represent standard deviation from 4 repeats.
(JPG)
In the right panel, the csn5a-2 abi5-1 double mutants showed improved germination rates over csn5a-2. These mutant lines were segregating sibling lines from the same cross. Germination rate of one representative csn5a-2abi5-1 double and csn5a-2 single mutant lines were counted and graphed as shown on the left. Error bars represent standard deviation from 4 repeats.
(JPG)
(A) Heatmap diagram of mRNA profiles in dry seeds of Col wild type, csn1-10, and csn5a-1. Genes with expression values from 13 and above were included. The color code is set from 13–300 in Log2 scale. Genes with expression level above 300 show the same color as those of 300 (red). The transcriptome profile of csn1-10 in dry seeds show greater dissimilarity to wild type than comparing csn5a-1 to wild type. (B) Heatmap plot showing gene expression profile in 2-day imbibed seeds of csn5a-1, csn1-10, and Col, compared to Col dry seeds. Genes with the expression value from 13 and above were included. The color code is set from 13 to 300 in Log2 scale. Genes with expression values above 300 are indicated with the same color as those of 300 (red). csn5a-1 displayed greater transcriptome changes than csn1-10 did at 2-day post imbibition.
(JPG)
(A and B) Heatmap showing ABA-related genes (A) and GA related genes (B) in 2-day imbibed seeds of csn5a-1 (5a-1), csn1-10, and Col, compared to those of Col dry seeds. (C) A panel of known seed germination regulatory genes showed similar expression profiles in csn5a-1, csn1-10 and Col. These genes are: DAG1 (DOF AFFECTING GERMINATION 1 AT3G61850), DOG1 (DELAY OF GERMINATION 1, AT5G45830), SPY SPINDLY, AT3G11540), SPT SPATULA, AT4G36930), LDL1/2 (ARABIDOPSIS LYSINE-SPECIFIC HISTONE DEMETHYLAS AT1G62830/ AT3G13682), MFT (MOTHER OF FT AND TFL, AT1G18100), RGL2 (RGA-LIKE 2, AT3G03450), RDO5 (REDUCED DORMANCY 5, AT4G11040), ABI5 ABA INSENSITIVE 5, AT2G36270).
(JPG)
Data Availability Statement
The data is accessible to the public at the NCBI site with the accession number GSE106223 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE106223).