

Abstract
The effectiveness of physical exercise on fat loss and improvement of aerobic capacity varies considerably between individuals. A strong linkage exists between common allelic variants of the adrenergic receptor genes and weight gain, as well as changes in body composition. Therefore we aimed to check if body composition and metabolic variables were modulated by the ADRB2 (Gly16Arg and Glu27Gln), ADRB3 (Trp64Arg) and ADRA2A (rs553668 G/A) gene polymorphisms in 163 Polish sedentary women (age 19-24; body mass index (BMI) 21.7 ± 0.2 kg·m-2) involved in a 12-week aerobic training program. Only 74.8% of participants lost fat mass. On average, participants lost 5.8 (10.4)% of their relative fat mass with training (range: +28.3 to -63.6%). The improvement of VO2max was significantly greater in women who could lose their fat mass compared to women who were unsuccessful in fat loss (4.5 (5.6)% vs. 1.5 (3.8)%; p = 0.0045). The carriers of a low number (0-3) of obesity-related risk alleles (ADRB2 Gly16, ADRB2 Glu27, ADRA2A rs553668 G) were more successful in fat mass loss compared to the carriers of a high number (5-6) of risk alleles (7.7 (9.8) vs 4.0 (9.4)%, p = 0.0362). The presented results support the assumption that variation within adrenergic receptor genes contributes to interindividual changes of body composition in response to physical exercise.
Key points.
There is a wide range of individual variability in the change of relative fat mass and BMI in response to a 12-week aerobic training program.
The efficiency of fat loss was inversely correlated with the improvement of VO2max in response to a 12-week aerobic training.
The carriers of a low number of obesity-related risk alleles were more successful in fat mass loss compared to the carriers of a high number of risk alleles.
Key words: ADRB2, ADRB3, ADRA2A, polygenic, obesity, fat, HDL
Introduction
Unhealthy lifestyle habits like lack of physical activity and excessive energy intake may result in overweight and obesity (Rank et al., 2012). The latter is one of the major and growing health problems of the XXI century. The presence of the elevated adipose tissue (increased adiposity) increases the likelihood of various medical conditions, such as hypertension, coronary heart disease, type 2 diabetes mellitus, and certain types of cancer (Masuo and Lambert, 2011). Thus, it is at least partly preventable by developing healthy diet and regular physical exercises that, in consequence, could help participants stay at a healthy weight (Greenway, 2015). However, a wide range of inter-individual variability in weight gain and changes in body composition induced by physical exercises and diets is seen in human populations which indicates the role of non-environmental factors such as genetic modifiers (Bouchard, 2008; Eynon et al., 2013; Garenc et al., 2003; Leońska-Duniec et al., 2016; Masuo et al., 2001; Wolfarth et al., 2005).
Various epidemiological and clinical studies indicate that strong linkage exists between common allelic variants of the adrenergic receptor genes and weight gain as well as changes in body composition (Bea et al., 2010; Masuo et al., 2005a; Phares et al., 2004; Szendrei et al., 2016). These adrenergic receptors (ADRs) encoded by the ADRA (α-adrenergic receptors - inhibitory) and ADRB (β-adrenergic receptors - stimulatory) genes are part of the sympathetic nervous system and exert their actions via coupling with the catecholamines. Because catecholamines are important regulators of lipolysis and energy expenditure during both energy restriction as well as exercise, it is clearly understandable that sympathetic nerve activation may play a role in modifying weight gain and changes in body composition. Reduced energy expenditure and lowered resting metabolic rate are predictive of overweight and obesity (Ahles and Engelhardt, 2014; O’Dell et al., 2015).
Recent studies have shown that allelic variation in the ADRs family exists, with the single nucleotide polymorphisms (SNPs) as the most common genetic polymorphisms (Green et al., 1993; Ikegami et al., 1996). Genetic diversity of the ADRs influence receptor expression, activity, and agonist regulation, in consequence contribute to the variable changes in body composition as well as weight gain and obesity (Hellstrom et al., 1999; Large et al., 1997). Within the β-adrenergic receptor family genes, the ADRB2 and the ADRB3 are of particular interest. β2-adrenergic receptors (β2-ADRs) encoded by the ADRB2 gene are the dominant lipolytic receptors in white adipose tissue and skeletal muscle (Enoksson et al., 2000; Hagstrom-Toft et al., 1998). β2-ADRs are also expressed throughout the smooth muscles of the cardiovascular and respiratory tracts and in the heart. Therefore, they play a pivotal role in the metabolic and musculoskeletal systems, promoting gluconeogenesis and glycogenolysis in the liver and skeletal muscles. They also influene insulin secretion and regulate energy expenditure through lipid mobilization from white adipose tissue (Brodde, 2008; Sarpeshkar and Bentley, 2010).
Gly16Arg (rs1042713,46G>A, G285A) and Glu27Gln (rs1042714, 79G>C, G318C) are the most common investigated polymorphisms of the ADRB2 gene (Gjesing et al., 2009; Masuo and Lambert, 2011; Meirhaeghe et al., 2000; Petrone et al., 2006; Szendrei et al., 2016). Studies of agonist stimulation in cultured cells revealed that neither Gly16Arg nor Glu27Gln affected the function of the β2-ADRs in terms of ligand binding or adenylyl cyclase activity. However, transfected cells expressing the Gly16 variant of the receptor were shown to have greater reduction in numbers or undergo significantly enhanced agonist-promoted downregulation when compared to Arg16. In contrast to Gly16, the Glu27 receptor form appears to be resistant to downregulation when compared to Gln27 variant (Green et al., 1994; 1995).
Numerous studies have investigated the impact of these polymorphic variants on changes in body composition, weight gain and obesity, as well as physical activity and athletic performance and conflicting results have been obtained (Ahmetov et al., 2016; Leońska-Duniec et al., 2016). In some studies it was found, that subjects carrying the Gly16 or Glu27 alone or both had increased risk of obesity (Gonzalez Sanchez et al., 2003; Lange et al., 2005; Large et al., 1997; Masuo et al., 2005a; Kawaguchi et al., 2006). Specifically, it was observed that Glu27 polymorphism interacts with physical activity influencing obesity risk among female subjects (Corbalan et al., 2002). Some research groups on the contrary, reported that the Gln27 is the risk allele (Meirhaeghe et al., 2000; Pereira et al., 2003). However, others found no relationship between Gly16Arg and Gln27Glu polymorphisms and obesity-related phenotypes (Bea et al., 2010; Echwald et al., 1998; Gjesing et al., 2009; Kortner et al., 1999; Rosado et al., 2015).
The β3-adrenergic receptors (β3-ADRs) that are encoded in human by ADRB3 gene are mainly expressed in adipose tissue and differ from the β2-ADRs in terms of a lower affinity for catecholamines, and resistance to desensitisation and downregulation (Masuo and Lambert, 2011). These differences lead to the different effects of catecholamine on β3-ADRs - they exert their effects mainly by lipolysis in white adipose tissue and thermogenesis in brown adipose tissue (Collins et al., 2004; Hoffstedt et al., 1999; Kirstein and Insel, 2004). It seems that decreased function of β3-ADRs in white adipose tissue could slow the mobilization of lipids from the white adipose tissue and, in consequence cause the retention of lipids in adipocytes. It could also affect thermogenesisin brown adipose tissue influencing body weight in humans. Within the ADRB3 gene the Trp64Arg (rs4994, T387C) polymorphism exists in human population. It was shown that adipose cells carrying at least one mutated Arg64 allele exhibit 2/3-fold reduced ability to produce cyclic adenosine monophosphate (cAMP) and lypolitic glycerol when compared to Trp64 homozygotes (Ahles and Engelhardt, 2014; Pietri-Rouxel et al., 1997; Umekawa et al., 1999). These results suggest that the Arg64 allele carriers have less ability to stimulate adenylyl cyclase, and in consequence lipolytic activity through the β3-adrenergic might be suppressed. Many studies have confirmed increased BMI (average 0.28 kg/m2) and body fat in carriers of the Arg64 allele (Ahles and Engelhardt, 2014; Leońska-Duniec et al., 2016). Polymorphic variants of the Trp64Arg are associated in many studies with abdominal obesity, weight gain, difficulty in losing weight, and lower resting metabolic rate as well as changes in body weight in response to exercise pointing at the Arg64 allele as risk allele or showed that the Trp64 allele is protective against obesity (Clement et al., 1995; Walston et al., 1995; Widen et al., 1995; Yoshida et al., 1995; Kim-Motoyama et al., 1997, Ukkola et al., 2000; Corella et al., 2001). In some cases no association was observed between the Trp64Arg polymorphism and obesity-phenotypes (Gagnon et al., 1996; Bea et al., 2010) or even contradictory results have been obtained (Phares et al., 2004).
As opposed to ADRB genes, there is less information about relationship between polymorphisms in ADRA genes and changes in body composition as well as weight gain and obesity-related phenotypes. However there are evidences that polymorphisms of the ADRA2B and the ADRA2A genes could be involved (Bea et al., 2010; Phares et al., 2004). Specifically, it was observed that the A allele of the G1780A (rs553668) polymorphism localized in the 3’UTR of the ADRA2A gene is associated with obesity and type 2 diabetes, as well as body mass index (BMI) and percentage of body fat (Lima et al., 2007; Langberg et al., 2013).
Considering the aforementioned facts we have decided to check if body mass and body composition, as well as metabolic variables observed in physically active participants will be modulated by the ADRB2, ADRB3 and ADRA2A gene polymorphisms. To test this hypothesis, we have performed a genetic association study that aimed to detect a correlation between the Gly16Arg and Glu27Gln of the ADRB2 gene, Trp64Arg of the ADRB3 gene as well as G1780A of the ADRA2A gene polymorphisms and selected body composition measurements as well as obesity-related metabolic traits in response to a 12-week aerobic training program.
Methods
Ethics statement
All the procedures followed in the study were approved by the Ethics Committee of the Regional Medical Chamber in Szczecin (Approval number 09/KB/IV/2011) and were conducted ethically according to the principles of the World Medical Association Declaration of Helsinki and ethical standards in sport and exercise science research. The experimental procedures were conducted in accordance with the set of guiding principles for reporting the results of genetic association studies defined by the Strengthening the Reporting of Genetic Association studies (STREGA) Statement (Little et al., 2009).
Participants
One hundred and sixty three Polish European Caucasian women mean age 21 ± 1 years (range 19–24) met the inclusion criteria and were included in the study. None of these individuals had engaged in regular physical activity in the previous 6 months. They had no history of any metabolic, cardiovascular diseases or previous musculoskeletal injuries. Participants were nonsmokers and refrained from taking any medications or supplements known to affect metabolism. Prior to the start of the training phase, participants were included into a dietary program and on the basis of individual dietary plan they were asked to keep a balanced diet, of approximately 2000 kcal a day. The participants were asked to keep a food diary every day. Weekly consultations were held on which the quality and quantity of meals were analyzed and, if necessary, minor adjustments were made.
Physical exercise training protocol
The training stage was preceded by a week-long familiarization stage, when the examined women exercised 3 times a week for 30 min, at an intensity of about 50% of their maximum heart rate (HRmax) (Zarębska et al., 2016). After the week-long familiarization stage, the main training started. Each training unit consisted of a warm-up routine (10 min), the main aerobic routine (43 min), and cool-down phase (stretching and breathing exercise for 7 min). The main aerobic routine was a combination of 2 alternating styles – low and high impact. Music of variable rhythm intensity (tempo) was incorporated into both styles. A 12-week program of low-high impact aerobics was divided as follows: (i) 3 weeks (9 training units), 60 min each, at about 50%–60% of HRmax, tempo 135–140 beats per minute (BPM), (ii) 3 weeks (9 training units), 60 min each, at 60%–70% of HRmax, tempo 140–152 BPM, (iii) 3 weeks (9 training units), 60 min with the intensity of 65%−75% of HRmax, tempo 145–158 BPM, and (iv) 3 weeks (9 training units), 60 min with an intensity of 65%−80% of HRmax, tempo 145–160 BPM. All 36 training units were administered and supervised by the same instructor.
Body composition measurements
All participants were measured for selected body mass and body composition variables before and after the completion of a 12-week training period. Body composition tests took place after fasting for at least 8 hours. Body mass and body composition were assessed with the bioimpedance method using a Tanita TBF 300M electronic scale (Arlington Heights, Illinois, USA). Body mass and body composition measurements taken with the use of the Tanita electronic scale are as follows: total body mass (kg), fat free mass (FFM, kg), fat mass (kg), fat mass percentage (FM, %), BMI (kg/m2), tissue impedance (Ohm), total body water (TBW, kg) and basal metabolic rate (BMR, kJ or kcal).
Biochemical and hematological analyses
Fasting blood samples were obtained in the morning from the elbow vein. Blood samples from each participant were collected in 2 tubes. Biochemical and hematological analyses were performed before the start of the aerobic fitness training programme and repeated at the 12th week of this training programme (after the 36th training unit). The analyses were performed immediately after the blood collection. Complete blood count, including white blood cells (WBC), red blood cells (RBC), hemoglobin (HGB), hematocrit (HTC), mean corpuscular volume (MCV), mean corpuscular hemoglobin (MCH), mean corpuscular hemoglobin concentration (MCHC), and total platelet level (PLT) were obtained using Sysmex K-4500 Haematology Analyzer (TOA SYSMEX, Kobe, Japan). All biochemical analyses were conducted using Random Access Automatic Biochemical Analyzer for Clinical Chemistry and Turbidimetry A15 (BIO- SYSTEMS S.A., Barcelona, Spain). Blood plasma was used to determine lipid profile: triglycerides (Tg), cholesterol (Chol), high-density lipoprotein (HDL) and low-density lipoprotein (LDL) concentrations. Plasma Tg and Chol concentrations were determined using diagnostic colorimetric enzymatic method according to the manufacturer’s protocol (BioMaxima S.A., Lublin, Poland). HDL plasma concentration was determined using human anti-ß-lipoprotein antibody and colorimetric enzymatic method according to the manufacturer’s protocol (BioMaxima S.A.). Plasma concentrations of LDL were determined using a direct method according to the manufacturer’s protocol (PZ Cormay S.A., Lomianki, Poland). All analysis procedures were verified with the use of multiparameteric control serum (BIOLABO S.A.S, Maizy, France), as well as control serum of normal level (BioNormL) and high level (BioPathL) lipid profiles (BioMaxima S.A.).
VO2max measurement
Subjects performed a continuous graded exercise test on an electronically braked cycle ergometer (VIAsprint™ 150P Bicycle, CareFusion Germany GmbH, Hoechberg, Germany) with an automatically calibrated volume sensor and a breath-by-breath gas analyzer (Oxycon Pro, Erich JAEGER GmbH, Hoechberg, Germany) to determine their maximal oxygen uptake (VO2max) before and after the completion of a 12-week training period. The device was calibrated in accordance with the manufacturer’s instructions. The test began by 5 min continuous pedaling, with a frequency of 60 revolutions per minute (RPM) and a relative load of 1.2 W/kg. After this phase, the workload was systematically increased by 15 watts every minute until voluntary exhaustion. The effort was interrupted when pedaling frequency declined by 10%, that is, when the pedalling frequency fell below 54 RPM. All of the participants reached RER greater than 1.0. The highest value of oxygen uptake was considered to be VO2max.
Genetic analyses
The buccal cells donated by the subjects were collected in Resuspension Solution (GenElute Mammalian Genomic DNA Miniprep Kit; Sigma, Steinheim, Germany) with the use of sterile foam-tipped applicators (Puritan, Guilford, Maine, USA). DNA was extracted from the buccal cells using a GenElute Mammalian Genomic DNA Miniprep Kit (Sigma) according to the manufacturer’s protocol. Obtained concentration of genomic DNA was 30–50 ng/1 μl. All samples were genotyped in duplicate using allelic discrimination assays with TaqMan® probes (Applied Biosystems, Carlsbad, California, USA) on CFX96 Touch™ Real-Time PCR Detection System (Bio-Rad, Hercules, California, USA). Freshly purified/sterile water was used as a negative control for PCR. To discriminate the ADRB2 Gly16Arg and Glu27Gln, ADRB3 Trp64Arg as well as ADRA2A G1780A alleles, TaqMan® Pre-Designed SNP Genotyping Assays were used (assay IDs: C___2084764_20, C___2084765_20, C___2215549_20 and C____996424_20, respectively), including appropriate primers and fluorescently labelled (FAM and VIC) MGB™ probes to detect the alleles.
Statistical analyses
Hardy-Weinberg equilibrium was tested by comparing the observed genotype frequencies with the expected ones using the Chi-square test with one degree of freedom in Microsoft EXCEL. Training responses were expressed as percentage change from baseline. The percentage changes were compared across genotypes using either parametric (t-test or one-way ANOVA) or non-parametric (Mann-Whitney or Kruskal-Wallis) tests for normally and non-normally distributed data, respectively. Spearman’s (non-parametric) correlations were used to assess the relationships between different phenotypes. Differences in phenotypes between groups were analysed using unpaired t tests. Power was calculated using non-central chi-square distribution (pchisq function in R, https://cran-r.projec.org) assuming alpha 0.05 and a variance explained of 0.01, 0.05, and 0.10 by additive effects at the marker of interest. The power for a stated sample size (n =163) was 24.8%, 81.5% and 98.1% for variances of 0.01, 0.05 and 0.10, respectively, Normality of the distribution was evaluated using the Kolmogorov-Smirnov test. The association of ADRB2 haplotypes with training responses was analysed using haplo.stats package for R. The regression of percentage change of body composition parameters, lipids and glucose on ADRB2 haplotypes was conducted using haplo.glm function assuming the additive model and minimum haplotype frequency of 5%.
Power for a stated sample size (n = 163) was calculated for a given set of haplotypes, their population frequencies and a specified genetic effect size (additive model of haplotype effects) in terms of a regression model R squared value (a haplo.power.qt function of the haplo.stats package). The power for the R squared values 1%, 5% and 10% was 18.9%, 74.1% and 97.3%, respectively. Linear regression coefficients corresponding to R squared 1%, 5% and 10% were 0.14, 0.32, 0.45, respectively
Gene-gene interactions among ADRB2, ADRB3 and ADRA2A polymorphisms were analysed using non-parametric model-free method of reducing genotype combinations called multifactor dimensionality reduction (MDR) using he MDR software package (version 3.0.2, http://sourceforge.net) (Gui et al., 2013; Ritchie et al., 2001). The 10-fold cross-validation training scores, cross-validation testing scores (the t-test statistic for the unequal variance computed by comparing phenotype between high- and low-level genotypes for the ten pooled testing sets), as well as cross validation consistency (CVC, the number of times the same model was chosen in the training set) were determined. P values for interactions were calculated using permutation tests. Single-locus analyses were carried out in STATISTICA data analysis software system, version 12 (StatSoft, Inc. 2014, www.statsoft.com).
Results
Individual variability in the change of relative fat mass and BMI are shown in Figures 1-2. Only 74.8% of participants could lose their relative fat mass in response to a 12-week aerobic training program, which was not dependent on their initial relative fat mass (p = 0.744) or BMI (p = 0.988). On average, participants lost 5.8 (10.4)% of their relative fat mass with training (range: +28.3 to -63.6%). The improvement of VO2max was significantly greater in women who could lose their fat mass compared to women who were unsuccessful in fat loss (4.5 (5.6)% vs. 1.5 (3.8)%; p = 0.0045). The efficiency of fat loss was inversely correlated with the improvement of VO2max in response to a 12-week aerobic training (r = – 0.37; p < 0.0001).
Figure 1.
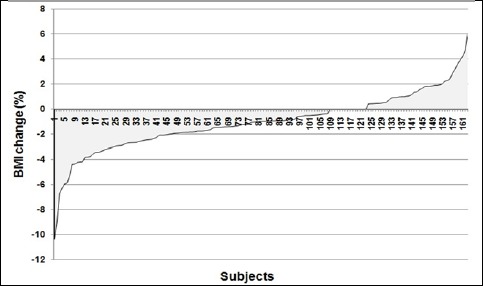
Individual variability in the BMI change (%) in response to a 12-week aerobic training of 163 women.
Figure 2.
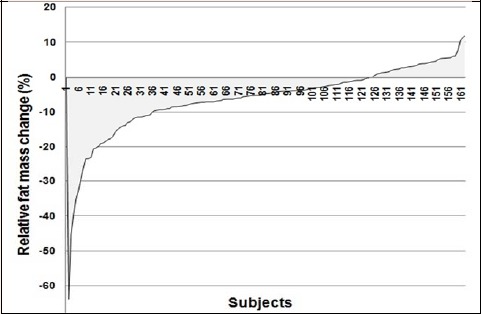
Individual variability in the relative fat mass change (%) in response to a 12-week aerobic training of 163 women.
All investigated polymorphisms conformed to Hardy-Weinberg expectations (W2 = 0.92, p = 0.337; 02 = 2.36, p = 0.124; 02 = 2.31, p = 0.129; 02 = 0.17, p = 0.680, for Gly16Arg, Glu27Gln of the ADRB2, Trp64Arg of the ADRB3 and G1780A of the ADRA2A, respectively. Owing to the low number of the ADRB3 Arg64/Arg64 (n = 3) and ADRA2A AA (n = 2) homozygotes, they were pooled together with corresponding heterozygous genotypes. There were no differences in percentage changes across genotypes for any of the analysed polymorphisms (Tables 1-4). However, polygenic analysis has shown that the carriers of a low number (0-3) of obesity-related risk alleles (ADRB2 Gly16, ADRB2 Glu27, ADRA2A rs553668 G) were more successful in fat mass loss compared to the carriers of a high number (5-6) of risk alleles (7.7 (9.8)% vs 4.0 (9.4)%, p = 0.0362).
Table 1.
The ADRB2 Gly16Arg genotypes and response to training (% change from baseline).
| Variables | Gly16/Gly16 (n=73) | Gly16/Arg16 (n=68) | Arg16/Arg16 (n=22) | p |
|---|---|---|---|---|
| body mass (kg) | -0.86±2.52 | -1.42±2.51 | -1.67±2.16 | 0.259 |
| BMI | -0.54±2.25 | -1.40±2.42 | -1.48±1.74 | 0.051 |
| BMR | -0.33 (-0.82, 0.19) | -0.43 (-0.82, 0.0) | -0.55 (-0.79, -0.13) | 0.214† |
| %FM | -3.80 (-7.14, 2.73) | -5.45 (-9.53, -1.77) | -6.94 (-10.98, -0.29) | 0.088† |
| FFM | 0.63 (-0.65, 1.36) | 1.07 (-0.33, 2.29) | 1.27 (-0.70, 2.06) | 0.524† |
| TBW | 0.61 (-0.58, 1.67) | 0.77 (-0.92, 2.35) | 1.51 (-0.63, 2.24) | 0.813† |
| TC | 0.10±12.30 | -0.64±11.67 | 0.54±14.67 | 0.904 |
| TGL | 10.06±40.55 | 8.25±35.40 | 20.73±40.06 | 0.410 |
| HDL | -4.58±17.06 | -5.40±18.41 | -5.29±15.92 | 0.959 |
| LDL | 4.89±20.89 | 4.06±20.54 | 3.20±22.59 | 0.938 |
| glucose | -4.55 (-11.25, 2.86) | -2.74 (-9.97, 4.98) | -0.69 (-6.10, 2.50) | 0.512† |
Mean ± SD or median with interquartile range (in brackets),
† Kruskal-Wallis test; BMI – body mass index; BMR – basal metabolic rate; %FM – fat mass percentage; FFM – fat free mass; TBW – total body water; TC – total cholesterol; TGL – triglycerides; HDL – high density lipoprotein, LDL –low density lipoprotein
Table 2.
The ADRB2 Glu27Gln genotypes and response to training (% change from baseline).
| Variables | Glu27/Glu27 (n=38) | Glu27/Gln27 (n=71) | Gln27/Gln27 (n=54) | p |
|---|---|---|---|---|
| body mass (kg) | -0.98±2.85 | -1.10±2.38 | -1.50±2.32 | 0.545 |
| BMI | -0.59±2.32 | -1.03±2.44 | -1.33±2.05 | 0.310 |
| BMR | -0.24 (-0.99, 0.20) | -0.42 (-0.83, 0.07) | -0.47 (-0.79, -0.12) | 0.450† |
| %FM | -3.31 (-7.04, 0.98) | -4.33 (-8.33, -0.82) | -6.09 (-10.71, -0.29) | 0.412† |
| FFM | 0.77 (-0.42, 1.93) | 0.64 (-0.88, 1.92) | 1.15 (-0.69, 2.18) | 0.955† |
| TBW | 0.60 (-0.31, 1.73) | 0.67 (-0.90, 2.06) | 1.19 (-0.63, 2.29) | 0.967† |
| TC | -1.35±11.02 | 1.94±12.49 | -2.05±12.75 | 0.159 |
| TGL | 3.69±35.67 | 14.07±40.31 | 11.34±37.67 | 0.404 |
| HDL | -4.11±16.60 | -6.35±19.13 | -3.90±15.65 | 0.693 |
| LDL | 3.71±18.83 | 7.48±21.30 | 0.59±21.33 | 0.184 |
| glucose | -5.26 (-11.27, 3.95) | -2.70 (-9.64, 4.35) | -2.62 (-9.41, 3.23) | 0.787† |
Mean ± SD or median with interquartile range (in brackets),
† Kruskal-Wallis test; BMI – body mass index; BMR – basal metabolic rate; %FM – fat mass percentage; FFM – fat free mass; TBW – total body water; TC – total cholesterol; TGL – triglycerides; HDL – high density lipoprotein, LDL –low density lipoprotein
Table 3.
The ADRB3 Trp64Arg genotypes and response to training (% change from baseline).
| Variables | Trp64/Trp64 (n=136) | Arg64/Arg64+Trp64/Arg64 (n=27) | p |
|---|---|---|---|
| body mass (kg) | -1.14±2.45 | -1.56±2.62 | 0.416 |
| BMI | -0.99±2.35 | -1.23±2.02 | 0.620 |
| BMR | -0.42 (-0.83, 0.04) | -0.39 (-0.80, 0.23) | 0.551† |
| %FM | -4.34 (-9.03, 0.0) | -5.08 (-7.04, -0.82) | 0.986† |
| FFM | 0.65 (-0.67, 2.13) | 1.08 (-0.44, 1.71) | 0.915† |
| TBW | 0.74 (-0.63, 2.32) | 1.11 (-0.60, 1.67) | 0.787† |
| TC | 0.08±12.51 | -1.28±11.44 | 0.603 |
| TGL | 11.46±39.36 | 7.14±33.44 | 0.594 |
| HDL | -6.10±17.58 | 0.44±15.61 | 0.074 |
| LDL | 5.13±21.10 | 0.21±19.40 | 0.263 |
| glucose | -2.84 (-9.61, 3.82) | -3.95 (-11.25, 4.29) | 0.844† |
Mean ± SD or median with interquartile range (in brackets),
† Mann-Whitney test ; BMI – body mass index; BMR – basal metabolic rate; %FM – fat mass percentage; FFM – fat free mass; TBW – total body water; TC – total cholesterol; TGL – triglycerides; HDL – high density lipoprotein, LDL –low density lipoprotein
Table 4.
The ADRA2A G1780A genotypes and response to training (% change from baseline).
| Variables | GG (n=124) | AA+AG (n=39) | p |
|---|---|---|---|
| body mass (kg) | -1.18±2.45 | -1.30±2.59 | 0.796 |
| BMI | -1.10±2.40 | -0.80±1.92 | 0.470 |
| BMR | -0.40 (-0.84, 0.08) | -0.48 (-0.79, 0.0) | 0.782† |
| %FM | -4.44 (-8.65, -0.15) | -5.11 (-10.98, 0.0) | 0.472† |
| FFM | 0.64 (-0.57, 1.92) | 1.08 (-0.93, 2.40) | 0.547† |
| TBW | 0.85 (-0.59, 2.11) | 0.81 (-1.24, 2.29) | 0.849† |
| TC | -0.51±12.10 | 0.99±13.08 | 0.509 |
| TGL | 11.08±38.44 | 9.67±38.68 | 0.841 |
| HDL | -6.47±16.42 | -0.39±19.71 | 0.057 |
| LDL | 4.33±20.64 | 4.27±21.79 | 0.987 |
| glucose | -2.74 (-9.50, 4.32 ) | -3.95 (-11.59, 2.38) | 0.610† |
Mean ± SD or median with interquartile range (in brackets),
† Mann-Whitney test ; BMI – body mass index; BMR – basal metabolic rate; %FM – fat mass percentage; FFM – fat free mass; TBW – total body water; TC – total cholesterol; TGL – triglycerides; HDL – high density lipoprotein, LDL –low density lipoprotein
Three haplotypes (Gly16;Glu27, 45.1%; Arg16; Gln27, 34.4%; Gly16; Gln27, 20.6%) were reconstructed. The most prevalent ADRB2 haplotype was Gly16;Glu27 (45.1%) and it was used as a reference in haplotype-based association analysis. The Arg16; Gln27 haplotype was associated with significantly smaller percentage change of BMI (-0.57 per copy of haplotype, p = 0.036) compared with reference Gly16;Glu27 haplotype (Table 5). Additionally, a two-way ADRB3 x ADRA2A interaction was detected for HDL percentage change (cross-validation consistency 10/10, cross-validation testing score 3.67, p = 0.018, Table 6). The double heterozygotes (ADRB3 Trp64Arg64/ADRA2A AG) and double homozygotes (ADRB3 Trp64Trp64/ADRA2A GG) exhibited a decrease while single heterozygotes (ADRB3 Trp64Arg64/ADRA2A GG and ADRB3 Trp64Trp64/ADRA2A AG) showed an increase in HDL serum concentration in response to training (Figure 3).
Table 5.
Regression of percentage change of body composition parameters, lipids and glucose on ADRB2 haplotypes.
| Haplotype/intercept | Intercept | [Arg16; Gln27] | [Gly16; Gln27] |
|---|---|---|---|
| body mass (kg) | -0.93 (-2.67) ** | -0.43 (-1.49) | 0.05 (0.13) |
| BMI | -0.65 (-2.04) * | -0.57 (-2.11) * | 0.03 (0.08) |
| BMR | -0.28 (-1.15) | -0.36 (-1.71 | -0.16 (-0.60) |
| %FM | -4.70 (-3.23) *** | -1.86 (-1.51) | 0.37 (0.24) |
| FFM | 1.0 (2.50) * | 0.12 (0.35) | -0.23 (-0.53) |
| TBW | 1.01 (2.45) * | 0.001 (0.005) | -0.04 (-0.10 |
| TC | 0.58 (0.36) | -0.33 (-0.23) | -1.22 (-0.66) |
| TGL | 7.18 (1.33) | 3.92 (0.86) | 2.12 (0.37) |
| HDL | -5.39 (-2.21) * | -0.18 (-0.09) | 1.22 (0.47) |
| LDL | 6.54 (2.24) * | -1.53 (-0.62) | -2.85 (-0.91) |
| glucose | -2.06 (-1.03) * | 1.19 (0.70) | -2.34 (-1.09) |
Regression coefficients and t statistic (in brackets); minimum frequency for a haplotype to be included 5%; the most common haplotype [Gly16;Glu27] (45.1%) was the reference haplotype; BMI – body mass index; BMR – basal metabolic rate; %FM – fat mass percentage; FFM – fat free mass; TBW – total body water; TC – total cholesterol; TGL – triglycerides; HDL – high density lipoprotein, LDL –low density lipoprotein.
* p < O.O5
** p < 0.01
*** p < 0.001.
Table 6.
Analysis of the two-way and three-way interactions between ADRB2, ADRB3 and ADRA2A genes using quantitative multifactor dimensionality reduction for body composition parameters, lipids and glucose.
| Variables | Best model* | Cross-validation training score | Cross-validation testing score | CVC* | p† |
|---|---|---|---|---|---|
| body mass (kg) | Gly16Arg | 1.57 | 0.61 | 9/10 | 0.557 |
| BMI (kg/m2) | Gly16Arg | 2.35 | 1.06 | 9/10 | 0.435 |
| %FM (%) | Gly16Arg,G1780A | 2.45 | -0.75 | 6/10 | 0.875 |
| FFM (kg) | G1780A | 1.18 | -2.29 | 6/10 | 0.995 |
| TBW (kg) | Gly16Arg, Glu27Gln,Trp64Arg | 2.20 | -1.86 | 6/10 | 0.986 |
| TC (mg/dL) | Glu27Gln | 1.81 | 1.91 | 10/10 | 0.213 |
| TGL (mg/dL) | Glu27Gln | 2.11 | -1.56 | 6/10 | 0.969 |
| HDL (mg/dL) | Trp64Arg, G1780A | 3.69 | 3.67 | 10/10 | 0.018 |
| LDL (mg/dL) | Glu27Gln | 1.76 | 0.55 | 7/10 | 0.557 |
| Glucose (mg/dL) | Gly16Arg,Glu27Gln, Trp64Arg | 2.45 | -1.25 | 9/10 | 0.960 |
*ablethe best gene-gene interaction model was determined using cross-validation consistency (CVC) and cross-validation testing score
† permuted p value (1000 permutations)
Figure 3.
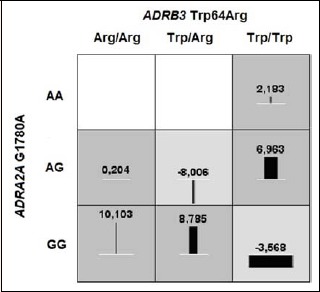
A two-way interaction between ADRB3 Trp64Arg and ADRA2AG1780A for HDL change.
Discussion
Our genetic association study was designed to test whether variation in the ADRB2, ADRB3 and ADRA2A genes can modulate changes in selected body mass, body composition and metabolic variables following 12 weeks of supervised aerobic exercise training in women. Despite the fact that there were no differences in percentage changes across genotypes for any of the analysed polymorphisms of the ADRB2 (Gly16Arg, Glu27Gln), ADRB3 (Trp64Arg) and ADRA2A (G1780A), the polygenic analysis has shown that the carriers of a low number (0-3) of obesity-related risk alleles (ADRB2 Gly16, ADRB2 Glu27, ADRA2A rs553668 G) were more successful in fat mass loss compared to the carriers of a high number (5-6) of risk alleles (7.7 (9.8)% vs 4.0 (9.4)%, p = 0.0362). Moreover, ADRB3xADRA2A interaction was detected for HDL percentage change. Therefore the presented results support the assumption that, i) genetic variation contributes to interindividual changes of selected body mass, body composition and metabolic variables in response to physical exercise as well as that ii) complex interaction of multiple genetic polymorphisms rather than an individual effect of a single polymorphic site have an influence on individual variation in responsiveness to exercise training (Jensen et al., 2009; Rankinen and Bouchard, 2012).
The ADRs gene family members have been extensively studied in the obesity field because of their participation in the regulation of energy expenditure (Marti et al., 2008; Ochoa et al., 2004). Particularly, the role of the lipolytic receptors genes, ADRB2, with its Gly16Arg and Glu27Gln polymorphisms, alone or in haplotype combination, in weight gain, obesity and changes in body composition have been investigated by many scientists. It has been shown that the Gly16 allele may influence the propensity to higher BMI, because the Gly16 allele is associated with lower receptor density, and in consequence reduced efficiency, when compared to Arg16 allele (Chou et al., 2012). A higher frequency of the Gly16 allele in men resistant to weight loss and those who regained body weight after successful initial weight loss at 6 months was noticed in a study of overweight men who participated in a 24-month weight loss programme consisting of a low-calorie diet and everyday aerobic exercise (Masuo et al., 2005b). Numerous studies have also focused on the second polymorphic site in the ADRB2 gene. Some studies showed that the Glu27 allele may limit ADRB2 downregulation and thus affect body weight (Kawaguchi et al., 2006; Lange et al., 2005). Corbalan et al. (2002) reported that women who were more active during their free time and were carriers of the Glu27 allele had higher body weight compared to non-carriers, suggesting that these women may be more resistant to losing weight.
In contrast, the study by Phares et al. (2004) and Szendrei et al. (2016) showed that Glu27 carriers had a tendency for a greater loss of percent total body fat, greater weight and BMI reductions compared with noncarriers. What is more, the study of Bea et al. (2010) showed gene x exercise interactions for ADRB2 Glu27Gln on change in lean soft tissue (LST). There was a significant LST gain with exercise of the Glu27 allele carriers compared to loss among controls and no intervention effect of the Glu27 allele noncarriers (Bea et al., 2010).
In our study, we have observed only a tendency of association of Gly16Arg and Glu27Gln alone with changes of selected body mass and body composition variables. However, we found that the ADRB2 risk alleles (Gly16 and Glu27) in combination with another risk allele of the ADRA2A gene (rs553668 G) were associated with significantly smaller change of fat mass following 12 weeks of supervised aerobic exercise training in women. The results of Jensen et al. (2009) were focused on analyzing the haplotype structure of the ADRB2 gene in Danish Caucasian subjects and association with BMI. The investigation clearly suggested that when multiple SNPs from a single gene were analyzed, unique interactions in specific haplotype pairs rather than individual SNPs may affect BMI.
Because the main role of catecholamines in human fat cells depends on the balance between lipolytic ADRB and antilipolytic ADRA receptors activities, gene-gene interactions in genes involved in the reciprocal regulation of lipolysis is inevitable (Phares et al., 2004). Indeed, such polygenic interactions have been spotted by some researchers. It has been reported that interactive effect of the ADRA2B Glu12/Glu9 and the ADRB3 Trp64Arg polymorphisms on obesity-related phenotypes in healthy white women exist (Dionne et al., 2001). It is worth noting that when the Glu12/Glu9 ADRA2B polymorphism did not associate with obesity-related phenotypes alone, subjects that carried the Arg64 ADRB3 and Glu9 ADRA2B variants had 9.3 kg greater fat mass and 4.8% greater percent body fat compared with subjects carrying only the Arg64 ADRB3 variant. Phares et al. (2004) found that the combined effects of the Glu12/Glu9, ADRA2B, Trp64Arg ADRB3, and Gln27Glu ADRB2 gene polymorphisms and their gene-gene interactions contribute significantly to explaining interindividual variability in body fat responses to exercise training. However, ADRA2B and ADRB3 interaction was the most significant source of variation for change in total body fat, trunk fat, and fat mass.
In the current study, we also observed interaction between ADRs genes; specifically, ADRB3xADRA2A interaction was detected for HDL percentage change. Our study showed that only single heterozygotes (ADRB3 Trp64Arg64/ADRA2A GG and ADRB3 Trp64Trp64/ADRA2A AG) had an increase in HDL serum concentration in response to training; the double heterozygotes (ADRB3 Trp64Arg64/ADRA2A AG) and double homozygotes (ADRB3 Trp64Trp64/ADRA2A GG) exhibited a decrease in HDL serum concentration after completion of 12 weeks of supervised aerobic exercise training.
It is widely accepted that regular aerobic exercise increases HDL-Chol. It was showed that beneficial adaptations in lipoprotein profile is achieved with moderate training intensities below the anaerobic threshold and training above the anaerobic threshold has no or even negative effects on blood lipoprotein profiles (Aellen et al., 1993; Drygas et al., 2000). Meta-anlysis by Kodama et al. (2007) showed that the minimum aerobic exercise volume for an increase in HDL level exist - minimal weekly exercise volume for HDL level increase was 900 kcal of energy expenditure or 120 minutes of exercise per week (Kodama et al., 2007). The intensity of our 12-week exercise program gradually increased from 50-60% HRmax to 65%−80% of HRmax in the last 3 weeks of the training programme and the weekly exercise volume was 180 minutes. Each training unit consisted of a warm-up, the main aerobic routine which was a combination of 2 alternating styles – low and high impact, and cool-down phase. Despite that, low-high impact aerobics in general refer to cardio with moderate training intensities zones, one may speculate that low- and high-impact workouts are at least partly similar to interval training with the high-intensity periods that are typically at or close to anaerobic exercise, while the recovery periods involve activity of lower intensity.
With respect to our findings, there is a reason for us to hypothesise that the subjects with combination of ADRB 3Arg64 / ADRA2A G and ADRB3 Trp64 / ADRA2A A rather than ADRB3 Trp64 / ADRA2A G respond better to our physical exercise program in terms of larger increase in HDL. It seems that duration, intensity as well as exercise frequency of our physical exercise programme were appropriate stimuli for ADRB3 Arg64 / ADRA2A G and ADRB3 Trp64 / ADRA2A A carriers to increase the HDL level. On the other hand, the ADRB3 Trp64 / ADRA2A G rather than ADRB3 Arg64 / ADRA2A A carriers respond with lowered HDL serum concentration in response to our 12 weeks of supervised aerobic exercise training. Therefore, there is a reason to hypothesise that low- and high- aerobic exercise training is not suitable for ADRB3 Trp64 / ADRA2A G carriers in terms of lower HDL levels, and the observed effect can be explained by an increase in energy consumption and achieving an ‘energy expenditure threshold’ during physical effort (Gibala and McGee, 2008; Kostrzewa-Nowak et al., 2015). It is also, higly probable, that other, rhythmic and repeated, aerobic exercises with moderate training intensities below the anaerobic threshold such as bicycling, jogging, or swimming would be more appropriate for this group of participants.
Conclusion
In summary, our findings suggest that the carriers of a low number of obesity-related risk alleles were more successful in fat mass loss compared to the carriers of a high number of risk alleles, as well as ADRB3xADRA2A gene interaction modifies the effects of aerobic exercise training in women on HDL levels. However, when we consider all aforementioned facts, the impact of genetic markers on determination of obesity-related traits is still unclear. Thus, the true level and the nature of the genotype x physical activity interactions in the field of obesity-related traits deserves to be further investigated. One of the possibile ways is using a composite score of genetic markers that have been identified in GWAS as an obesity risk SNPs in the gene × physical activity interaction analyses (Li et al., 2010). However, only a comprehensive understanding of the underlying genetic and epigenetic mechanisms will enable us to uncover the “missing heritability” of the obesity-related traits (Herrera et al., 2011, Rankinen and Bouchard, 2012).
Acknowledgements
The study was supported by National Science Centre (grant no. 2012/07/B/NZ7/01155). The authors declare that they have no conflicts of interest regarding the publication of this paper. Current experiment complied with the current laws of the Poland.
Biographies
Agata LEOŃSKA-DUNIEC
Employment
Faculty of Tourism and Recreation, Gdansk University of Physical Education and Sport, Gdansk, Poland
Degree
PhD, MD
Research interests
Genetics, Genomics
E-mail: leonska.duniec@gmail.com
Zbigniew JASTRZĘBSKI
Employment
Faculty of Tourism and Recreation, Gdansk University of Physical Education and Sport, Gdansk, Poland
Degree
PhD, MD
Research interests
Sport Exercise, Physiology, Genetics
E-mail: zb.jastrzebski@op.pl
Aleksandra JAŻDZEWSKA
Employment
Faculty of Tourism and Recreation, Gdansk University of Physical Education and Sport, Gdansk, Poland
Degree
PhD, MD
Research interests
Sport Exercise, Physiology, Genetics
E-mail: olazarebska@o2.pl
Waldemar MOSKA
Employment
Faculty of Tourism and Recreation, Gdansk University of Physical Education and Sport, Gdansk, Poland
Degree
PhD, MD
Research interests
Sport Exercise, Sport Managements
E-mail: waldemarmoska@wp.pl
Ewelina LILIŃSKA-KUKLIK
Employment
Faculty of Physical Education, Gdansk University of Physical Education and Sport, Gdansk, Poland
Degree
PhD, MD
Research interests
Traumatology, Physical Exercise, Physiology
E-mail: e.lulinska-kuklik@osw2wejherowo.pl
Marek SAWCZUK
Employment
Faculty of Tourism and Recreation, Gdansk University of Physical Education and Sport, Gdansk, Poland
Degree
PhD, MD
Research interests
Genetics, Genomics
E-mail: sawczuk_marek@wp.pl
Svetlana I. GUBAYDULLINA
Employment
Sport Technology Research Centre, Volga Region State Academy of Physical Culture, Sport and Tourism, Kazan, Russia
Degree
MS
Research interests
Anthropology, nutrition, genetics
E-mail: gubajdullina_svetlana@mail.ru
Alsu T. SHAKIROVA
Employment
Department of Propaedeutics of Childhood Diseases, Kazan State Medical University, Kazan, Russia
Degree
PhD, MD
Research interests
Nutrition, Genetics
E-mail: a.t.shakirova@mail.ru
Paweł CIĘSZCZYK
Employment
Faculty of Physical Education, Gdansk University of Physical Education and Sport, Gdansk, Poland
Degree
PhD, MD
Research interests
Genetics, Genomics
E-mail: cieszczyk@poczta.onet.pl
Adam MASZCZYK
Employment
Department of Theory and Practice of Sport; Academy of Physical Education in Katowice; Poland.
Degree
PhD, MD
Research interests
Statistics, Physical Exercise, Physiology
E-mail: a.maszczyk@awf.katowice.pl
Ildus I. AHMETOV
Employment
Head of the Laboratory of Molecular Genetics, Kazan State Medical University, Kazan, Russia.
Degree
Dr. Med. Sci., PhD, MD
Research interests
Exercise and sports genomics
E-mail: genoterra@mail.ru
References
- Aellen R., Hollmann W., Boutellier U. (1993) Effects of aerobic and anaerobic training on plasma lipoproteins. International Journal of Sports Medicine 14(7), 396-400. [DOI] [PubMed] [Google Scholar]
- Ahles A., Engelhardt S. (2014) Polymorphic variants of adrenoceptors: pharmacology, physiology, and role in disease. Pharmacological Reviews 66(3), 598-637. [DOI] [PubMed] [Google Scholar]
- Ahmetov I.I., Egorova E.S., Gabdrakhmanova L.J., Fedotovskaya ON. (2016) Genes and Athletic Performance: An Update. Medicine and Sport Science 61, 41-54. [DOI] [PubMed] [Google Scholar]
- Bea J.W., Lohman T.G., Cussler E.C., Going S.B., Thompson P.A. (2010) Lifestyle modifies the relationship between body composition and adrenergic receptor genetic polymorphisms, ADRB2, ADRB3 and ADRA2B: a secondary analysis of a randomized controlled trial of physical activity among postmenopausal women. Behavioral Genetics 40(5), 649-659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouchard C. (2008) Gene–environment interactions in the etiology of obesity: defining the fundamentals. Obesity (Silver Spring) 16(Suppl 3), S5-S10. [DOI] [PubMed] [Google Scholar]
- Brodde O.E. (2008) Beta1- and beta2-adrenoceptor polymorphisms and cardiovascular diseases. Fundamental and Clinical Pharmacology 22(2), 107-125. [DOI] [PubMed] [Google Scholar]
- Chou Y.C., Tsai C.N., Lee Y.S., Pei J.S. (2012) Association of adrenergic receptor gene polymorphisms with adolescent obesity in Taiwan. Pediatrics International 54(1), 111-116. [DOI] [PubMed] [Google Scholar]
- Clément K., Vaisse C., Manning B.S., Basdevant A., Guy-Grand B., Ruiz J., Silver K.D., Shuldiner A.R., Froguel P., Strosberg A.D. (1995) Genetic variation in the b3-adrenergic receptor and an increased capacity to gain weight in patients with morbid obesity. New England Journal of Medicine 333(6), 352-354. [DOI] [PubMed] [Google Scholar]
- Collins S., Cao W., Robidoux J. (2004) Learning new tricks from old dogs: beta-adrenergic receptors teach new lessons on firing up adipose tissue metabolism. Molecular Endocrinology 18(9), 2123-2131. [DOI] [PubMed] [Google Scholar]
- Corbalán M.S., Marti A., Forga L., Martínez-González M.A., Martínez J.A. (2002) The 27Glu polymorphism of the beta2-adrenergic receptor gene interacts with physical activity influencing obesity risk among female subjects. Clinical Genetics 61(4), 305-307. [DOI] [PubMed] [Google Scholar]
- Corella D., Guillen M., Portoles O., Sorli J.V., Alonso V., Folch J., Sáiz C. (2001) Gender specific associations of the Trp64Arg mutation in the beta3-adrenergic receptor gene with obesity-related phenotypes in a Mediterranean population: interaction with a common lipoprotein lipase gene variation. Journal of Internal Medicine 250(4), 348-360. [DOI] [PubMed] [Google Scholar]
- Dionne I.J., Turner A.N., Tchernof A., Pollin T.I., Avrithi D., Gray D., Shuldiner A.R., Poehlman E.T. (2001) Identification of an interactive effect of beta3- and alpha2b-adrenoceptor gene polymorphisms on fat mass in Caucasian women. Diabetes 50(1), 91-95. [DOI] [PubMed] [Google Scholar]
- Drygas W., Kostka T., Jegier A., Kuński H. (2000) Long-term effects of different physical activity levels on coronary heart disease risk factors in middle-aged men. International Journal of Sports Medicine 21(4), 235-241. [DOI] [PubMed] [Google Scholar]
- Echwald S.M., Sorensen T.I., Tybjaerg-Hansen A., Andersen T., Pedersen O. (1998) Gln27Glu variant of the human beta2-adrenoreceptor gene is not associated with early-onset obesity in Danish men. Diabetes 47(10), 1657-1658. [DOI] [PubMed] [Google Scholar]
- Enoksson S., Talbot M., Rife F., Tamborlane W.V., Sherwin R.S., Caprio S. (2000) Impaired in vivo stimulation of lipolysis in adipose tissue by selective β2-adrenergic agonist in obese adolescent girls. Diabetes 49(12), 2149-2153. [DOI] [PubMed] [Google Scholar]
- Eynon N., Nasibulina E.S., Banting L.K., Cieszczyk P., Maciejewska-Karlowska A., Sawczuk M., Bondareva E.A., Shagimardanova R.R., Raz M., Sharon Y., Williams A.G., Ahmetov I.I., Lucia A., Birk R. (2013) The FTO A/T polymorphism and elite athletic performance: a study involving three groups of European athletes. PLoS One 8(4), e60570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gagnon J., Mauriege P., Roy S., Sjostrom D., Chagnon Y.C., Dionne F.T., Oppert J.M., Pérusse L., Sjöström L., Bouchard C. (1996) The Trp64Arg mutation of the beta3 adrenergic receptor gene has no effect on obesity phenotypes in the Quebec Family Study and Swedish Obese Subjects cohorts. Journal of Clinical Investigation 98(9), 2086-2093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garenc C., Pérusse L., Chagnon Y. C., Rankinen T., Gagnon J., Borecki I. B., Leon A. S., Skinner J. S., Wilmore J. H., Rao D.C., Bouchard C., HERITAGE Family Study (2003) Effects of β2-Adrenergic Receptor Gene Variants on Adiposity: The HERITAGE Family Study. Obesity Research 11, 612–618. [DOI] [PubMed] [Google Scholar]
- Gibala M.J., McGee S.L. (2008) Metabolic adaptations to short-term high-intensity interval training: a little pain for a lot of gain? Exercise and Sport Sciences Reviews 36(2), 58-63. [DOI] [PubMed] [Google Scholar]
- Gjesing A.P., Sparsø T., Borch-Johnsen K., Jørgensen T., Pedersen O., Hansen T., Olsen N.V. (2009) No consistent effect of ADRB2 haplotypes on obesity, hypertension and quantitative traits of body fatness and blood pressure among 6,514 adult Danes. PLoS One 4(9), e7206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- González Sánchez J.L., Proenza A.M., Martínez Larrad M.T., Ramis J.M., Fernández Pérez C., Palou A., Serrano Ríos M. (2003) The glutamine 27 glutamic acid polymorphism of the beta2-adrenoceptor gene is associated with abdominal obesity and greater risk of impaired glucose tolerance in men but not in women: a population-based study in Spain. Clinical Endocrinology (Oxford) 59(4), 476-481. [DOI] [PubMed] [Google Scholar]
- Green S.A., Cole G., Jacinto M., Innis M., Liggett S.B. (1993) A polymorphism of the human beta 2-adrenergic receptor within the fourth transmembrane domain alters ligand binding and functional properties of the receptor. Journal of Biological Chemistry 268(31), 23116-23121. [PubMed] [Google Scholar]
- Green S.A., Turki J., Bejarano P., Hall I.P., Liggett S.B. (1995) Influence of beta 2-adrenergic receptor genotypes on signal transduction in human airway smooth muscle cells. Am. J. Respir. Molecular and Cellular Biology 13(1), 25-33. [DOI] [PubMed] [Google Scholar]
- Green S.A., Turki J., Innis M., Liggett S.B. (1994) Aminoterminal polymorphisms of the human β2-adrenergic receptor impart distinct agonist promoted regulatory properties. Biochemistry 33(32), 9414-9419. [DOI] [PubMed] [Google Scholar]
- Greenway F.L. (2015) Physiological adaptations to weight loss and factors favouring weight regain. International Journal of Obesity (London) 39(8), 1188-1196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gui J., Moore J.H., Williams S.M., Andrews P., Hillege H.L., van der Harst P., Navis G., Van Gilst W.H., Asselbergs F.W., Gilbert-Diamond D. (2013) A Simple and Computationally Efficient Approach to Multifactor Dimensionality Reduction Analysis of Gene-Gene Interactions for Quantitative Traits. PLoS One 8(6), e66545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagstrom-Toft E., Enoksson S., Moberg E., Bolinder J., andArner P. (1998) β-adrenergic regulation of lipolysis and blood flow in human skeletal muscle in vivo. American Journal of Physiology 275(6 Pt 1), E909-916. [DOI] [PubMed] [Google Scholar]
- Hellstrom L., Large V., Reynisdottir S., Wahrenberg H., Arner P. (1999) The different effects of a Gln27Glu β2-adrenoceptor gene polymorphism on obesity in males and in females. Journal of Internal Medicine 245(3), 253-259. [DOI] [PubMed] [Google Scholar]
- Herrera B.M., Keildson S., Lindgren C.M. (2011) Genetics and epigenetics of obesity. Maturitas 69(1), 41-49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffstedt J., Poirier O., Thorne A., Lonnqvist F., Herrmann S.M., Cambien F., Arner P. (1999) Polymorphism of the human beta3-adrenoceptor gene forms a well-conserved haplotype that is associated with moderate obesity and altered receptor function. Diabetes 48(1), 203-205. [DOI] [PubMed] [Google Scholar]
- Ikegami H., Yamato E., Fujisawa T., Hamada Y., Fujioka Y., Rakugi H., Higaki J., Murakami H., Shimamoto K., Ogihara T. (1996) Analysis of candidate genes for insulin resistance in essential hypertension. Hypertension Research 19(Suppl 1), S31-34. [DOI] [PubMed] [Google Scholar]
- Jensen M.K., Nielsen M., Koefoed P., Nielsen H.B., Ullum H., Haastrup E., Romner B., Moltke F.B., Olsen N.V. (2009) Haplotype structure of the beta2-adrenergic receptor gene in 814 Danish Caucasian subjects and association with body mass index. Scandinavian Journal of Clinical and Laboratory Investigation 69(7), 801-808. [DOI] [PubMed] [Google Scholar]
- Kawaguchi H., Masuo K., Katsuya T., Sugimoto K., Rakugi H., Ogihara T., Tuck M.L. (2006) β2- and β3- adrenoceptor polymorphisms relate to subsequent weight gain and blood pressure elevation in obese normotensive individuals. Hypertension Research 29(12), 951-959. [DOI] [PubMed] [Google Scholar]
- Kim-Motoyama H., Yasuda K., Yamaguchi T., Yamada N., Katakura T., Shuldiner A.R., Akanuma Y., Ohashi Y., Yazaki Y., Kadowaki T. (1997) A mutation of the beta-3-adrenergic receptor is associated with visceral obesity but decreased serum triglyceride. Diabetologia 40, 469-472. [DOI] [PubMed] [Google Scholar]
- Kirstein S.L., Insel P.A. (2004) Autonomic nervous system pharmacogenomics: a progress report. Pharmacology Reviews 56(1), 31-52. [DOI] [PubMed] [Google Scholar]
- Kodama S., Tanaka S., Saito K., Shu M., Sone Y., Onitake F., Suzuki E., Shimano H., Yamamoto S., Kondo K., Ohashi Y., Yamada N., Sone H. (2007) Effect of aerobic exercise training on serum levels of high-density lipoprotein cholesterol. Archives of Internal Medicine 167(10), 999-1008. [DOI] [PubMed] [Google Scholar]
- Kortner B., Wolf A., Wendt D., Beisiegel U., Evans D. (1999) Lack of association between a human beta-2 adrenoceptor gene polymorphism (gln27glu) and morbid obesity. International Journal of Obesity and Related Metabolic Disorders 23(10), 1099–1100. [DOI] [PubMed] [Google Scholar]
- Kostrzewa-Nowak D., Nowak R., Jastrzębski Z., Zarębska A., Bichowska M., Drobnik-Kozakiewicz I., Radzimiński Ł., Leońska-Duniec A., Ficek K., Cięszczyk P. (2015) Effect of 12-week-long aerobic training programme on body composition, aerobic capacity, complete blood count and blood lipid profile among young women. Biochemia Medica 25(1), 103-113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Långberg E.C., Seed Ahmed M., Efendic S., Gu H.F., Östenson C.G. (2013) Genetic association of adrenergic receptor alpha 2A with obesity and type 2 diabetes. Obesity (Silver Spring) 21(8), 1720-1725. [DOI] [PubMed] [Google Scholar]
- Lange L.A., Norris J.M., Langefeld C.D., Nicklas B.J., Wagenknecht L.E., Saad M.F., Bowden D.W. (2005) Association of adipose tissue deposition and beta-2 adrenergic receptor variants: the IRAS family study. International Journal of Obesity (London) 29(5), 449-457. [DOI] [PubMed] [Google Scholar]
- Large V., Hellström L., Reynisdottir S., Lönnqvist F., Eriksson P., Lannfelt L., Arner P. (1997) Human beta-2 adrenoceptor gene polymorphisms are highly frequent in obesity and associate with altered adipocyte beta-2 adrenoceptor function. Journal of Clinical Investigation 100(12), 3005-3013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leońska-Duniec A., Ahmetov I.I., Zmijewski P. (2016) Genetic variants influencing effectiveness of exercise training programmes in obesity - an overview of human studies. Biology of Sport 33(3), 207-214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S., Zhao J.H., Luan J., Ekelund U., Luben R.N., Khaw K.T., Wareham N.J., Loos R.J. (2010) Physical activity attenuates the genetic predisposition to obesity in 20,000 men and women from EPIC-Norfolk prospective population study. PLoS Medicine 7(8), e1000332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lima J.J., Feng H., Duckworth L., Wang J., Sylvester J.E., Kissoon N., Garg H. (2007) Association analyses of adrenergic receptor polymorphisms with obesity and metabolic alterations. Metabolism 56, 757-765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Little J., Higgins J.P., Ioannidis J.P., Moher D., Gagnon F., von Elm E., Khoury M.J., Cohen B., Davey-Smith G., Grimshaw J., Scheet P., Gwinn M., Williamson R.E., Zou G.Y., Hutchings K., Johnson C.Y., Tait V., Wiens M., Golding J., van Duijn C., McLaughlin J., Paterson A., Wells G., Fortier I., Freedman M., Zecevic M., King R., Infante-Rivard C., Stewart A., Birkett N. (2009) Strengthening the reporting of genetic association studies (STREGA), an extension of the STROBE Statement. Human Genetics 125(2), 131-151. [DOI] [PubMed] [Google Scholar]
- Marti A., Martinez-González M.A., Martinez J.A. (2008) Interaction between genes and lifestyle factors on obesity. Proceedings of the Nutrition Society 67(1), 1-8. [DOI] [PubMed] [Google Scholar]
- Masuo K., Lambert G.W. (2011) Relationships of adrenoceptor polymorphisms with obesity. Journal of Obesity 2011, 609485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masuo K., Katsuya T., Fu Y., Rakugi H., Ogihara T., Tuck M.L. (2005a) β2- and β3-adrenergic receptor polymorphisms are related to the onset of weight gain and blood pressure elevation over 5 years. Circulation 111(25), 3429-3434. [DOI] [PubMed] [Google Scholar]
- Masuo K., Katsuya T., Kawaguchi H., Fu Y., Rakugi H., Ogihara T., Tuck M.L. (2005b) Rebound weight gain as associated with high plasma norepinephrine levels that are mediated through polymorphisms in the beta2-adrenoceptor. American Jornal of Hypertension 18(11), 1508-1516. [DOI] [PubMed] [Google Scholar]
- Masuo K., Mikami H., Ogihara T., Tuck M.L. (2001) Familial obesity, sympathetic activation and blood pressure level. Blood Pressure 10(4), 199-204. [DOI] [PubMed] [Google Scholar]
- Meirhaeghe A., Helbecque N., Cottel D., Amouyel P. (2000) Impact of polymorphisms of the human β2-adrenoceptor gene on obesity in a French population. International journal of obesity and related metabolic disorders 24(3), 382-387. [DOI] [PubMed] [Google Scholar]
- Ochoa M.C., Marti A., Martinez J.A. (2004) Obesity studies in candidate genes. Medicina Clinica 122(14), 542-551. [DOI] [PubMed] [Google Scholar]
- O’Dell T.J., Connor S.A., Guglietta R., Nguyen PV. (2015) β-Adrenergic receptor signaling and modulation of long-term potentiation in the mammalian hippocampus. Learning & Memory 22(9), 461-471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pereira A.C., Floriano M.S., Mota G.F., Cunha R.S., Herkenhoff F.L., Mill J.G., Krieger J.E. (2003) Beta2 adrenoceptor functional gene variants, obesity and blood pressure level interactions in the general population. Hypertension 42(4), 685-692. [DOI] [PubMed] [Google Scholar]
- Petrone A., Zavarella S., Iacobellis G., Zampetti S., Vania A., Di Pietro S., Galgani A., Leonetti F., Di Mario U., Buzzetti R. (2006) Association of β2 adrenergic receptor polymorphisms and related haplotypes with triglyceride and LDL-cholesterol levels. Eur. Journal of Human Genetics 14(1), 94-100. [DOI] [PubMed] [Google Scholar]
- Phares D.A., Halverstadt A.A., Shuldiner A.R., Ferrell R.E., Douglass L.W., Ryan A.S., Goldberg A.P., Hagberg J.M. (2004) Association between body fat response to exercise training and multilocus ADR genotypes. Obesity Research 12(5), 807-815. [DOI] [PubMed] [Google Scholar]
- Piétri-Rouxel F., St John Manning B., Gros J., Strosberg A.D. (1997) The biochemical effect of the naturally occurring Trp64-->Arg mutation on human beta3-adrenoceptor activity. Europena Journal of Biochemistry 247(3), 1174-1179. [DOI] [PubMed] [Google Scholar]
- Rank M., Siegrist M., Wilks D.C., Haller B., Wolfarth B., Langhof H., Halle M. (2012) Long-term effects of an inpatient weight-loss program in obese children and the role of genetic predisposition-rationale and design of the LOGIC-trial. BMC Pediatrics 12, 30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rankinen T., Bouchard C. (2012) Gene-exercise interactions. Progress in Molecular Biology and Translational Science 108, 447-460. [DOI] [PubMed] [Google Scholar]
- Ritchie M.D., Hahn L.W., Roodi N., Bailey L.R., Dupont W.D., Parl F.F., Moore J.H. (2001) Multifactor-dimensionality reduction reveals high-order interactions among estrogen-metabolism genes in sporadic breast cancer. American Journal of Human Genetics 69, 138-147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosado E.L., Bressan J., Martinez J.A. (2015) Environmental factors and beta2-adrenergic receptor polymorphism: influence on the energy expenditure and nutritional status of obese women. Lipids 50(5), 459-467. [DOI] [PubMed] [Google Scholar]
- Sarpeshkar V., Bentley D. J. (2010) Adrenergic-beta(2) receptor polymorphism and athletic performance. Journal of Human Genetics 55(8), 479-485. [DOI] [PubMed] [Google Scholar]
- Szendrei B., González-Lamuño D., Amigo T., Wang G., Pitsiladis Y., Benito P.J., Gomez-Candela C., Calderón F.J., Cupeiro R., PRONAF Study Group (2016) Influence of ADRB2 Gln27Glu and ADRB3 Trp64Arg polymorphisms on body weight and body composition changes after a controlled weight-loss intervention. Applied Physiology, Nutrition, and Metabolism 41(3), 307-314. [DOI] [PubMed] [Google Scholar]
- Ukkola O., Rankinen T., Weisnagel S.J., Sun G., Pérusse L., Chagnon Y.C., Després J.P., Bouchard C. (2000) Interactions among the alpha2-, beta2- and beta3-adrenergic receptor genes and obesity-related phenotypes in the Quebec Family Study. Metabolism 49(8), 1063-1070. [DOI] [PubMed] [Google Scholar]
- Umekawa T., Yoshida T., Sakane N., Kogure A., Kondo M., Honjyo H. (1999) Trp64Arg mutation of beta3-adrenoceptor gene deteriorates lipolysis induced by beta3-adrenoceptor agonist in human omental adipocytes. Diabetes 48(1), 117-120. [DOI] [PubMed] [Google Scholar]
- Walston J., Silver K., Bogardus C., Knowler W.C., Celi F.S., Austin S., Manning B., Strosberg A.D., Stern M.P., Raben N., Sorkin J.D., Roth J., Shuldiner A.R. (1995) Time of onset of non-insulin- dependent diabetes mellitus and genetic variation in the b3-adrenergic receptor gene. New England Journal of Medicine 333(6), 343-347. [DOI] [PubMed] [Google Scholar]
- Widén E., Lehto M., Kanninen T., Walston J., Shuldiner A.R., Groop L.C. (1995) Association of a polymorphism in the b3-adrenergic receptor gene with features of the insulin resistance syndrome in Finns. New England Journal of Medicine 333(6), 348-351. [DOI] [PubMed] [Google Scholar]
- Wolfarth B., Bray M.S., Hagberg J.M., Perusse L., Rauramaa R., Rivera M.A., Roth S.M., Rankinen T., Bouchard C. (2005) The human gene map for performance and health-related fitness phenotypes: the 2004 update. Medicine and Science in Sports and Exercise 37(6), 881-903. [PubMed] [Google Scholar]
- Yoshida T., Sakane N., Umekawa T., Sakai M., Takahashi T., Kondo M. (1995) Mutation of b3-adrenergic-receptor gene and response to treatment of obesity. Lancet 46(8987), 1433-1434. [DOI] [PubMed] [Google Scholar]
- Zarębska A., Jastrzębski Z., Moska W., Leońska-Duniec A., Kaczmarczyk M., Sawczuk M., Maciejewska-Skrendo A., Żmijewski P., Ficek K., Trybek G., Lulińska-Kuklik E., Semenova E.A., Ahmetov I.I., Cięszczyk P. (2016) The AGT Gene M235T Polymorphism and Response of Power-Related Variables to Aerobic Training. Journal of Sports Science and Medicine 15(4), 616-624. [PMC free article] [PubMed] [Google Scholar]