

Abstract
This study sought to determine if the differences in serum estradiol we have previously observed to occur during the mid-follicular (MF) and mid-luteal (ML) phases of the female menstrual cycle could be attributed to estrogen-induced receptor activation and subsequent effects on myogenic-related genes which may otherwise impact muscle regeneration in response to eccentric exercise. Twenty-two physically-active females (20.9 ± 1.4 years, 63.5 ± 9.0 kg, 1.65 ± 0.08 m) underwent an eccentric exercise bout of the knee extensors during the MF and ML phases of their 28-day menstrual cycle. Prior to (PRE), at 6 (6HRPOST), and 24 (24HRPOST) hours post-exercise for each session, participants had muscle biopsies obtained. Skeletal muscle estradiol and estrogen receptor-α (ER-α) content and ER-DNA binding were determined with ELISA. Real-time PCR was used to assess ER-α, Myo-D, and cyclin D1 mRNA expression. Data were analyzed utilizing a 2 x 3 repeated measures univariate analyses of variance (ANOVA) for each criterion variable (p ≤ .05). Skeletal muscle estradiol levels were not significantly impacted by either menstrual phase (p > 0.05); however, both ER-α mRNA and protein were significantly increased during MF (p < 0.05). ER-DNA binding and Myo-D mRNA expression increased significantly in both menstrual phases in response to exercise but were not different from one another; however, cyclin D1 mRNA expression was significantly greater during MF. This study demonstrates that skeletal muscle ER-α activation in response to eccentric exercise up-regulates myogenic-related gene expression independent of serum estradiol levels occurring during the human menstrual cycle.
Key points.
Skeletal muscle estradiol levels do not differ during the MF and ML phase, despite differences in serum estradiol.
Skeletal muscle ER-α mRNA and protein expression was greater during MF, despite greater levels of serum estradiol during ML and no difference in muscle estradiol between MF and ML.
Skeletal muscle ER-DNA binding and Myo-D mRNA expression was not impacted by either menstrual phase but was elevated in response to eccentric exercise.
Skeletal muscle cyclin D1 mRNA expression was impacted by eccentric exercise and was greater for MF.
Key words: Estrogen, estrogen receptor, skeletal muscle, myogenesis
Introduction
17β-estradiol (estradiol) augments increases in satellite cell activation and myoblast proliferation in rodents following exercise-induced muscle damage (Thomas et al., 2010). In humans, the concentration of estradiol in circulation fluctuates during the menstrual cycle in a way that the levels are low during the early to midpoint (days 1-8) of the follicular phase and elevated during the midpoint of the luteal phase (days 18-24) (Longcope, 1986). Considering this, several studies have been conducted attempting to determine whether estradiol fluctuations during the menstrual cycle may influence muscle’s ability to undergo increases in strength and mass (Sakamaki-Sunaga et al. 2015; Sung et al., 2014) due to the impact satellite cell activation may have on muscle regeneration in response to resistance exercise.
We have previously shown that, during the mid-luteal (ML) phase of the menstrual cycle, females were less susceptible to the consequences of exercise-induced muscle damage and oxidative stress than males (Kerksick, et al., 2008). Carter et al. (2001) also concluded that muscle damage was attenuated in response to downhill running in women taking oral contraceptives and tested during the ML phase (high estrogen) when compared with eumenorrheic women not taking oral contraceptives and tested during the mid-follicular (MF) phase (low estrogen). We have also recently shown that females, who indeed did have lower serum estradiol levels during the MF than ML phase, experienced a greater amount of oxidative stress (McKinely-Barnard et al., 2017). As a result, based on the greater amount of muscle damage and oxidative stress that occurs during the follicular phase, it has been suggested that women’s muscles may have a better ability to respond to resistance training in the follicular phase compared to the luteal phase (Reis et al., 1995; Sung et al., 2014); although, the mechanisms by which this occurs are not completely understood.
Regarding estradiol’s impact on exercise-induced muscle damage, one possible mechanism is its association with the myogenic activation of satellite cells to assist with muscle repair and regeneration during recovery (Farup et al., 2014). Estradiol has been shown to influence satellite cell activation and proliferation in rats following eccentric exercise from downhill running (Enns and Tiidus, 2008). Furthermore, it has been shown that the ER, specifically ER-α, mediates estradiol-induced increases in rat skeletal muscle satellite cells post-exercise (Enns et al., 2008; Thomas et al., 2010). Estradiol functions by binding the ER, which is a 66 kDa nuclear regulatory protein that, when bound by estradiol, functions as a hormone-activated transcription factor. Activation of the ER is typically associated with estradiol-induced conformational changes in the structure of the ER (Klein-Hitpass et al., 1989; Kumar and Chambon, 1988). Extracellular estradiol readily diffuses across the sarcolemma and binds the ER, leading to dimerization of the ER and subsequent translocation to the nucleus. Upon translocation, the estradiol-ER complex binds with high affinity to a 13 bp palindromic sequence, known as the estrogen response element (ERE) (Beato et al., 1995). The enhancer-like ERE sequences are located upstream of the TATA promoter within the regulatory region of estradiol-responsive genes (Beato, 1991). After ERE binding, the estradiol-ER complex activates transcription of estradiol-responsive genes, such as myogenic differentiation factor D (Myo-D), thereby resulting in proliferation initiation (Weisz and Bresciani, 1988).
The Myo-D gene is a specific target for ER-induced expression due to the presence of three half-palindromic EREs approximately 650 bp of the 5’ upstream regulatory promoter region (Zingg et al., 1991). Because of ER-ERE binding, transcription of cis-linked target genes will be stimulated. In adult, fully-differentiated skeletal muscle, Myo-D is expressed at a level that is relatively low. However, in response to muscle damage the expression of Myo-D increases and is related to satellite cell activation and proliferation (Sakurai et al., 2013).
Cyclins function as positive regulatory subunits within a class of protein kinases known as cyclin-dependent kinases (CDKs), and have been shown to be important regulators of major cell cycle transitions. Specifically, cyclin D1 is a multifunctional G1-phase cyclin that activates ER-mediated transcription of estradiol-responsive genes. It has been shown that cyclin D1 activates ER-mediated transcription in an estradiol-independent manner. Cyclin D1 initiates transcription in the absence of estradiol; however, transcription is enhanced in the presence of estradiol (Zwijsen et al., 1997). This suggests that ER activation by cyclin D1 occurs independent of CDK binding and that the ER does not require occupancy of estradiol.
There appears to be a paucity of data in humans describing the role of estradiol and/or ER activation relative to the expression of genes targeted by ER signaling which are known to play a role in the myogenic activation of satellite cells. Furthermore, the role that exercise-induced muscle damage may play in regulating ER signaling relative to estradiol-responsive gene expression is also unknown. Therefore, this study builds upon the results we observed for serum estradiol and eccentric exercise from our previous study (McKinley-Barnard et al., 2017). In the present study, we used muscle samples that were obtained from our previous study to determine if the differences in estradiol we have previously observed to occur during the ML and MP phases of the female menstrual cycle (McKinley-Barnard et al., 2017) could be attributed to estradiol-induced ER activation and subsequent effects on myogenic-related genes which may impact muscle regeneration in response to eccentric exercise.
Methods
Participants
Twenty-two apparently healthy, physically-active, eumenorrheic females (20.9 ± 1.4 years, 63.5 ± 9.0 kg, 1.65 ± 0.08 m) who had not taken any form of hormonal contraception for one year prior to the study could participate. All eligible participants were informed of the risks and benefits of the study prior to signing the university-approved informed consent documents, and approval was granted by the Institutional Review Board of Human Subjects of Baylor University. Additionally, all experimental procedures involved in the study conformed to the ethical consideration of the Declaration of Helsinki.
Procedures
Participants calculated their menstrual cycle utilizing the calendar method to determine the length of their cycle, and subsequently their MF and ML phases. Participants completed two eccentric exercise protocols during each phase of the menstrual cycle (day 6 for MF and day 21 for ML).
Eccentric exercise protocol
The eccentric exercise protocol was performed unilaterally and isokinetically (Biodex System 3, Shirley, NY, USA) employing a previous protocol (Kerksick et al., 2013), and consisted of 10 sets of 10 repetitions with 3 minutes of rest between sets at an isokinetic speed of 30°/second. Participants were required to exert a minimum of 30% of their individual pre-determined maximal voluntary initiation of contraction (MVIC) for each repetition to ensure the appropriate effort was put forth for each bout. The eccentric bout had a torque threshold of 300 ft-lbs. set for each participant. The leg exercised for the first testing session during the MF phase was randomly selected. The subsequent testing session during the ML phase involved the opposing leg.
Muscle biopsies
Percutaneous muscle biopsies (15 to 20 mg) were obtained from the middle portion of the vastus lateralis muscle at the midpoint between the patella and the greater trochanter of the femur at a depth between 1 and 2 cm using a 14-gauge fine needle aspiration biopsy needle (Tru-Core I Biopsy Instrument, Medical Device Technologies, Gainesville, FL, USA). After the initial biopsy, the next biopsy attempts were made to extract tissue from approximately the same location as the initial biopsy by using the pre-biopsy markings and depth markings on the needle. After removal, adipose tissue was trimmed from the muscle specimens. Specimens were immediately frozen in liquid nitrogen and then stored at -80°C for later analysis. At each eccentric exercise testing session, muscle samples were obtained at PRE, 6HRPOST, and 24HRPOST.
Total RNA isolation from skeletal muscle samples
Approximately 10-15 mg of muscle tissue was used for biochemical analysis. Total cellular RNA was extracted from the homogenate of biopsy samples with a monophasic solution of phenol and guanidine isothiocyanate contained within the TRI-reagent (Sigma Chemical Co., St. Louis, MO). An aliquot of TRI-reagent was added to each tube, and then muscle samples were homogenized using a pestle. An aliquot of chloroform was added to each tube and shaken, then allowed to sit for 15 minutes. The clear aqueous phase was transferred into a new microfuge tube. Subsequently, an aliquot of 100% isopropanol was added to each tube and allowed to sit at room temperature for 5 to 10 minutes. Samples were then centrifuged at 12,000 x g at 4°C for 10 minutes, allowing for the formation of a RNA pellet. The supernatant was discarded, then an aliquot of 75% ethanol was added to wash the pellet. The samples were then centrifuged at 7500 x g at 4°C for 5 minutes, then the supernatant discarded. The washing procedure was repeated twice. The pellet was allowed to air dry for 5 minutes, then an aliquot of nuclease free water was added to the microfuge tube. Total RNA concentrations from each sample were determined spectrophotometrically with an optical density of 260 nm (OD260) and to verify RNA integrity and absence of RNA degradation, indicated by an OD260/OD280 ratio of approximately 2.0 (Ausubel et al., 1999). Our protocol produced an average (±SD) ratio of 1.93 (± 0.44) for all samples. The final concentration was adjusted to 200 ng•μl-1 by diluting the total RNA extracts into DEPC-treated nuclease-free H2O. The RNA samples were stored at -80°C until later analyses.
Reverse transcription and complementary DNA synthesis
A reverse transcription reaction mixture [200 ng of total cellular RNA, 5× reverse transcription buffer, a dNTP mixture containing dATP, dCTP, dGTP, and dTTP, MgCl2, RNase inhibitor, oligo(dT)15 primer, nuclease-free H2O, and 1 U•μl-1 MMLV reverse transcriptase enzyme (Bio-Rad, Hercules, CA)] were incubated at 42°C for 40 minutes, heated to 85°C for five minutes, and then quick-chilled on ice yielding the complementary DNA (cDNA) product. The standardized cDNA solutions were frozen at -80°C until real-time RT-PCR was performed.
Oligonucleotide primers for PCR
The mRNA sequences of human skeletal muscle ER-α (NM_001122742), Myo-D (X56677), cyclin-D1 (NM_001758), and GAPDH (NM_002046) published in the NCBI Entrez Nucleotide database (www.ncbi.nlm.nih.gov) were used to construct PCR primers using Beacon Designer software (Bio-Rad, Hercules, CA, USA), and then commercially synthesized (Integrated DNA Technologies, Coralville, IA). These primers (Table 1) amplify fragments of ER-α, Myo-D, cyclin-D1, and GAPDH of 142, 177, 108, and 155 bp, respectively.
Table 1.
Oligonucleotide primer sequences for real-time PCR.
| GAPDH | Forward | 5’-AAA GCC TGC CGG TGA CTA AC-3’ |
| Reverse | 5’-CGC CCA ATA CGA CCA AAT CAG A-3’ | |
| ER-α | Forward | 5’- AAT ATG CCC TTT TGC GATG -3’ |
| Reverse | 5’- ACA AAG CAA AGC TGC GAC AA -3’ | |
| Cyclin D1 | Forward | 5’-CAA CTT CCT GTC CTA CTA CC - 3’ |
| Reverse | 5’ -TCC TCC TCC TCC TCT TCC-3’ | |
| Myo-D | Forward | 5’-CGC CAC CGC CAG GAT ATG-3’ |
| Reverse | 5’ -GTC ATA GAA GTC CGT TGT G-3’ |
Real time-PCR
Aliquots of cDNA were added to each of the PCR reactions for ER-α, Myo-D, cyclin-D1, and GAPDH. Specifically, each PCR reaction contained the following mixtures: cDNA template along with 2× SYBR Green Supermix (Bio-Rad, Hercules, CA) [100 mM KCl mixture, 40 mM Tris-HCl, 0.4 mM of each dNTP, 50 U•ml-1 of iTaq DNA polymerase, 6.0 mM MgCl2, SYBR Green I, 20 nM flourescein], sense and anti-sense primers, and nuclease-free dH2O. Each PCR reaction was amplified with a thermal cycler (Bio Rad, Hercules, CA) and the amplification sequence involved a denaturation step at 95°C for 30 seconds, primer annealing at 55°C for 30 seconds, and extension at 72°C for 60 seconds. RT-PCR was performed over 40 cycles with emitted fluorescence from the SYBR green fluorophore being measured after each cycle. An emission of fluorescence occurs due to the integration of the SYBR green into the double-stranded cDNA produced during the PCR reaction. GAPDH was used as an external reference standard for detecting relative change in the quantity of target mRNA due to its consideration as a constitutively expressed housekeeping gene (Thellin et al., 1999). All CT values were assessed in the linear portion of amplification and a DNA melting curve analysis was performed after amplification to assure that the single gene products were amplified in absence of primer-dimers.
The specificity of the PCR was demonstrated with an absolute negative control reaction containing no cDNA template, and single gene products confirmed using DNA melt curve analysis. Based on our previous work (Kerksick et al., 2013), the expression of mRNA was determined from the post-exercise fold-change in gene expression analyzed using the Pfaffl method (Pfaffl, 2001) [i.e., E-ΔΔCT = E is 1.00 + (percent primer efficiency/100); ΔΔCT = (CT gene of interest - CT GAPDH) post-exercise - (CT gene of interest - CT GAPDH) pre-exercise]. A 0-fold change is a 100% down-regulation of the gene and a 1-fold change indicated no change in gene expression relative to baseline mRNA levels, whereas a 2-fold and 3-fold change in gene expression indicated a 100% and 200% increase in gene expression, respectively.
Skeletal muscle cellular and nuclear extraction and protein content
Portions of each muscle sample were homogenized using a cell extraction buffer (item # FNN0011, Life Technologies, Grand Island, NY, USA) and a tissue homogenizer. The cell extraction buffer was supplemented with phenylmethanesulphonyl fluoride (PMSF) (item # P7626, Sigma Chemical Company, St. Louis, MO, USA) and a protease inhibitor cocktail (item # P2714, Sigma Chemical Company, St. Louis, MO, USA) with broad specificity for the inhibition of serine, cysteine, and metallo-proteases. Remaining muscle samples were homogenized in nuclear extraction buffer (item # 40410, Active Motif, Carlsbad, CA). The nuclear extracts were supplemented with a phosphatase inhibitor buffer [34,35]. Total protein content for cellular- and nuclear-extracted samples were analyzed in duplicate and determined spectrophotometrically at a wavelength of 595 nm and using bovine serum albumin as the standard.
Skeletal muscle estradiol analysis
Estradiol was determined in each cellular extract by ELISA (Eagle Biosciences, Inc., Nashua, NH, USA) (Ekenros et al., 2017). The specificity of the kit for estradiol is 100% with the sensitivity estimated to be 8.68 pg/ml. All samples were analyzed in duplicate and in one run to avoid batch effects. The absorbances for each variableswere determined at a wavelength of 450 nm using a microplate reader (iMark, Bio-Rad, Hercules, CA) against a known standard curve, and final concentration expressed relative to total, cellular-extracted protein concentration. The overall intra-assay percent coefficient of variation was 7.4%.
Skeletal muscle ER-α protein analysis
In each PRE sample, the ER-α protein content was determined in nuclear-extracted muscle samples by a commercially-available nuclear receptor ELISA kit (Active Motif, Carlsbad, CA). The kit uses a specific polyclonal antibody which recognizes the N-terminal, ligand-independent, domain of the ER-α. The assay is 100% specific and has a sensitivity of 0.6 µg of ER in nuclear-extracted LNCaP cells. All samples were analyzed in duplicate and in one run to avoid batch effects. The absorbances for each variable were determined at a wavelength of 450 nm using a microplate reader (iMark, Bio-Rad, Hercules, CA) against a known standard curve consisting of ER-α peptide, and concentrations expressed relative to total, cellular-extracted protein concentration. The overall intra-assay percent coefficient of variation was 8.3%.
ER-DNA binding analysis
The extent of ER-DNA binding was determined in nuclear extracts by a commercially-available ELISA kit (Assay Bio Tech, Inc., Sunnyvale, CA, USA). This kit uses a consensus DNA oligonucleotide (AGAACA) representing the ERE to first bind the ER, after which a specific polyclonal antibody interacts with the N-terminal domain of the ER. The assay is 100% specific and has a sensitivity of 0.3 µg of AR-DNA binding in nuclear-extracted MCF7 cells. All samples were analyzed in duplicate and in one run to avoid batch effects, and were also run against nuclear-extracted MCF7 as a positive control. The absorbances were determined at a wavelength of 450 nm using a microplate reader (iMark, Bio-Rad, Hercules, CA) and expressed relative to total, nuclear-extracted protein concentration. The overall intra-assay percent coefficient of variation was 9.6%.
Statistical analyses
Statistical analyses were performed by utilizing separate 2 x 3 [Phase (MF or ML) x Time (PRE, 6HRPOST, and 24HRPOST)] factorial analyses of variance (ANOVA) with repeated measures. Significant interaction differences were determined by separate one-way ANOVA. Further analysis of the main effect for Time was performed involving the LSD Post Hoc Test. The index of effect size utilized was partial Eta squared (η2), which estimates the proportion of variance in the dependent variable that can be explained by the independent variable. Partial Eta squared effect sizes were determined to be: weak = 0.17, medium = 0.24, strong = 0.51, very strong = 0.70 (O’Connor et al., 2007). An a-priori power calculation from our previous study (McKinely-Barnard et al., 2017) demonstrated that 15 participants were adequate to detect a significant difference in the criterion variable of serum estradiol, given a type I error rate (α-level) of 0.05 and a power of 0.80. To determine if any significant differences in existed in mRNA expression, estradiol, and ER-DNA binding between legs at each PRE time point during the MF and ML phases, separate paired t-tests were performed. All statistical procedures were performed using SPSS 20.0 software and a probability level of p ≤ .05 was adopted throughout.
Results
Outcome Variable Differences Between Right and Left Legs at PRE
As can be seen in Table 2, for ER mRNA and protein, there was a significant difference between the right and left legs at the PRE time point. However, there were no significant differences detected between the right and left legs in for Myo-D and cyclin D1 mRNA expression, estradiol, and ER-DNA at the PRE time point (p > 0.05).
Table 2.
Means (±SD) for each variable at the PRE time point for the MF and ML phases.
| Variables | MF | ML |
|---|---|---|
| Estradiol | 15.36 (9.58) | 18.43 (13.81) |
| ER Protein | .063 (.011) | .038 (.020) † |
| ER-DNA Binding | .801 (.408) | .733 (.447) |
| ER mRNA | 1.36 (.021) | .871 (.081) # |
| Myo-D mRNA | .866 (.064) | .884 (.063) |
| Cyclin D1 mRNA | .846 (.083) | .882 (.052) |
ER mRNA and protein were significantly different between right and left legs at the PRE time point (p < 0.05). All other variables were not significantly different (p > 0.05).
† p ≤ 0.001
# p ≤ 0.003.
Skeletal muscle estradiol
As can be seen in Figure 1, skeletal muscle estradiol failed to demonstrate a significant cycle x time interaction (p = 0.669, effect size = 0.114) or main effect for cycle (p = 0.431, effect size = 0.123) and time (p = 0.399, effect size = 0.207).
Figure 1.
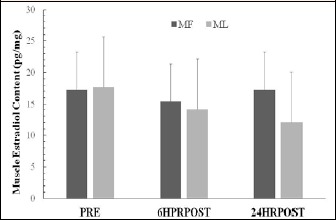
Skeletal muscle estradiol concentrations at pre-exercise (PRE) during the MF and ML menstrual phases. Data are presented as mean ± SD. No significant differences at any time point and between exercise sessions were detected (p > 0.05).
Skeletal muscle ER-α mRNA and protein expression
For ER-α expression at PRE, a significant main effect for cycle was observed for mRNA (p = 0.001, effect size = 1.00) and protein (p = 0.021, effect size = 0.985) showing both to be significantly greater at MF compared to ML (Figure 2).
Figure 2.
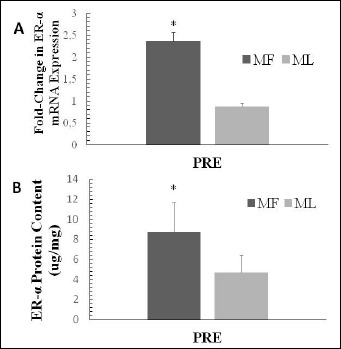
Skeletal muscle ER-α (A) mRNA and protein (B) expression at pre-exercise (PRE) during the MF and ML menstrual phases. Data are presented as mean ± SD. * indicates MF to be significantly greater than ML (p < 0.05).
ER-DNA binding
For ER-DNA binding, no significant cycle x time interaction (p = 0.924, effect size = 0.062) or main effect for cycle (p = 0.728, effect size = 0.064) was observed. However, a significant main effect for time was observed (p = 0.042, effect size = 0.610). Post-hoc analyses showed ER-DNA binding was greater at 6HRPSOT and 24HRPOST (Figure 3).
Figure 3.
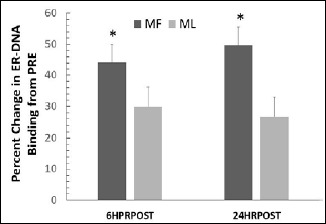
Skeletal muscle ER-DNA binding concentrations in response to eccentric exercise during the MF and ML menstrual phases. Data are presented as mean ± SD. * indicates ER-DNA binding was significantly greater in response to eccentric exercise (p < 0.05) but not impacted by either MF or ML (p > 0.05).
mRNA expression for gene markers of myogenic activation
For Myo-D, no significant cycle x time interaction (p = 0.853, effect size = 0.074) or main effect for cycle (p = 0.893, effect size = 0.086) was observed. However, a significant main effect for time existed (p = 0.003, effect size = 0.861). Post-hoc analyses showed that the up-regulation in Myo-D mRNA expression was significantly different at 6HRPSOT and 24HRPOST (Figure 4).
Figure 4.
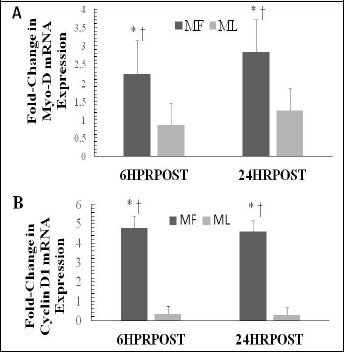
Skeletal muscle (A) Myo-D and (B) cyclin D1 mRNA expression in response to eccentric exercise during the MF and ML menstrual phases. Data are presented as mean ± SD and expressed as fold-change in expression. * indicates significantly greater expression in response to eccentric exercise (p < 0.05); † indicates significantly greater expression during MF (p < 0.05).
A significant cycle x time interaction existed for cyclin D1 (p = 0.014, effect size, 0.756). In addition, significant main effects were observed for cycle (p = 0.037, effect size = 0.551) and time (p = 0.026, effect size = 0.662). Post-hoc analyses showed that cyclin D1 mRNA expression was significantly greater for MF than ML at 6HRPOST and 24HRPOST (Figure 4).
Discussion
In our previous study (McKinley-Barnard et al., 2017), we showed the levels of serum estradiol to be significantly greater at ML than MF. However, in the present study we were interested to see if the differences in serum estradiol we previously observed had any impact on muscle estradiol, ER-α mRNA and protein expression, ER-DNA binding, and the subsequent effect on the expression of the ER target genes/markers of myogenic activity, Myo-D and cyclin D1. Based on the results presented herein, we have shown muscle estradiol levels to not be differentially affected by either the MF or ML phase, or by eccentric exercise. We originally assumed that the differences in serum estradiol levels between menstrual phases would possibly mirror those in muscle; however, muscle estradiol was found to be similar at baseline during each menstrual phase. We were unable to find published human data on muscle estradiol in females; however, data from rats showing no significant change in muscle estradiol levels following treadmill exercise at 30 m/min for 30 min (Aizawa et al., 2008) assist in corroborating our present results. Therefore, our data indicate muscle estradiol do not differ between MF and ML phases, and that elevated estradiol in circulation during ML does not differentially impact ER activation compared to MF.
Since a different leg was used for the eccentric exercise session and muscle biopsy at the MF and ML phase, we wanted to make sure our post-exercise results were not influenced simply by differences that may have existed between legs at the PRE (baseline) time point. With the exception of ER-α mRNA and protein, we found that no significant differences existed between legs at the PRE time point for all of our outcome variables. As a result, we are confident that our results are not influenced by any baseline differences due to the MF and ML phases. We were unable to find any supporting data from right and left legs in females during the MF and ML phases. However, our results are supported by those from a previous study involving older men which compared skeletal muscle gene expression between right and left legs using transcriptome expression from microarray and mRNA expression using real time PCR. Microarray analysis found that only 31 of 5499 genes (0.6%) and none of the mRNA species assessed were significantly different between legs (Tarnopolsky et al., 2007).
Even though our present results indicate no significant difference between menstrual phases in muscle estradiol before or after eccentric exercise, in baseline muscle samples we did show a significantly greater ER-α mRNA and protein expression at MF. This agrees with a previous study in humans showing a greater level of estradiol during the luteal phase compared to the follicular phase, yet a greater expression of ER-α mRNA and protein expression during the follicular phase (Ekenros et al., 2017). Aside from this study, we were unable to locate other published results on ER expression in skeletal muscle. Although, ER-α expression in the endometrium has been shown to increase during the follicular phase, to the highest levels of expression at ovulation, and then gradually decrease during the luteal phase (Giudice, 2006). Animal studies have also shown a significant elevation in ER-α in bovine cervix and vagina during the follicular phase (Sagsoz et al., 2010). The fact that serum estradiol levels are higher during the luteal phase than the follicular phase may be important because the much higher levels of progesterone could have a suppressing effect on estradiol function systemically. So, the higher levels of estradiol during the luteal phase are to offset the potential so-called “anti-estrogen” effect of the higher progesterone which occurs during this phase. However, our results suggest that the expression of ER-α mRNA and protein do not appear to be dependent on either serum or muscle estradiol levels. Furthermore, the reason for this occurrence is not well known.
Interestingly, we observed ER-DNA binding (indicative of ER activation) and Myo-D mRNA expression to be increased with eccentric exercise, but not impacted by either menstrual phase. However, cyclin D1 was impacted by eccentric exercise, but much more so during the MF phase which seems to suggest cyclin D1 gene expression may be preferentially affected by ER activation, rather than by serum and/or muscle estradiol levels. With the present study, we were interested in determining the role that serum estradiol and subsequent ER activation may play in increasing the expression of genes associated with the myogenic activation of satellite cells in response to eccentric exercise. Beyond its anti-oxidant capacity and ability to attenuate oxidative stress and muscle damage, estradiol may conceivably facilitate the repair of exercise-induced muscle damage through the activation of satellite cells. Although, in humans a non-steroidal anti-inflammatory drug (NSAID) which minimized the inflammation that accompanies muscle damage has been shown to attenuate the myogenic response to a single bout of eccentric exercise (Mikkelsen et al., 2009). This suggests that estradiol’s anti-oxidant capability to attenuate muscle damage may also hinder the myogenic response. However, estradiol has also been shown to induce satellite cell activation and proliferation following eccentric exercise in rodents through an ER-mediated mechanism (Enns and Tiidus, 2008), and one that appears to be specifically linked to the activation of ER-α (Enns et al., 2008; Thomas et al., 2010). Moreover, since we failed to observe a menstrual phase-induced increase in ER activation, the attenuation in exercised-induced muscle damage and oxidative stress that we (Kerksick et al., 2008; McKinley-Barnard et al., 2017) and others (Carter et al., 2001; Minahan et al., 2015) have previously shown to occur appears to be a ER-mediated process independent of serum estradiol (Enns et al., 2008).
The expression of cis-linked target genes will be up-regulated upon ER-ERE binding. Since the Myo-D gene is a specific target for ER-induced transcription (Zingg et al., 1991), this indicates that estradiol-bound and activated ER can specifically bind to EREs within the Myo-D gene, thereby up-regulating its expression. We showed that ER-DNA binding (indicative of estradiol-bound ER activation and subsequent ERE binding) was significantly increased in response to eccentric exercise, but was not dependent on either menstrual phase. Furthermore, since Myo-D is an ER target gene, it is not surprising that we also observed the same response with its expression, which is a similar response in Myo-D mRNA expression previously shown in rodents 24 hours following eccentric exercise (Sakurai et al.., 2013). Myo-D is a myogenic regulatory factor (MRF) that is expressed once satellite cells re-enter the cell cycle. Myo-D has been suggested to have a functional role during cell activation (Saboruin and Rudnicki, 2000). The mRNA expression of Myo-D serves as a marker of satellite cell proliferation (Sakurai et al., 2013). Under normal conditions, satellite cell differentiation following exercise-induced muscle injury is considered to be a cause of Myo-D mRNA expression observed at 24 hours post-exercise (Di Marco et al., 2005). Considering that ER-DNA binding results in transactivation, our data indicate that while Myo-D expression may be ER-dependent, it appears to be independent of serum estradiol levels as its expression in response to eccentric exercise was not impacted by either menstrual phase.
Along with the MRFs, the myogenic activation of satellite cells is also regulated by cyclin-dependent kinases (CDKs) which have a catalytic dependence upon the cyclin family of regulatory proteins. Cyclin D1 regulates progression through G1 and entry into the S phase of the cell cycle, and is responsible for its role in myoblast differentiation by activating CDK2 (Knight and Kothary, 2011). Cyclin D1 and estradiol are synergistic in stimulating ER-regulated gene transcription; therefore, cyclin D1 can apparently substitute for estradiol in activating the ER by two mechanisms. First is that, in the presence of estradiol, cyclin D1 is a strong enhancer of ERE-responsive gene transcription, but also activates the TATA box within the gene’s core promoter. As a result, the action of cyclin D1, while responsive to estradiol, is not promoter-specific (Zwijsen et al., 1997). Second is that activation of the ER by cyclin D1 is not due to a change in ER translocation to the nucleus, rather ER activation by cyclin D1 is associated with a direct physical interaction which is located within the E/F domain region of liganded, as well as unliganded, ER. Interestingly, we (Li et al., 2015) and others (Roberts et al., 2011) have shown the cyclin D1 gene to be responsive to resistance exercise; however, in the present study we showed that cyclin D1 mRNA expression was significantly increased in response to exercise that was greater during MF compared to ML. This suggests that ER activation by cyclin D1 does not require receptor binding of estradiol, and that low levels of estradiol during MF seems to preferentially increase the expression of the cyclin D1, perhaps to facilitate the increase in Myo-D gene expression we observed to be similar during ML.
Based on the ability of elevated circulating estradiol to minimize the severity of exercise-induce muscle damage, it is conceivable that skeletal muscle regeneration may be negatively impacted if exercise is performed during ML. However, our data indicate that ER activation and Myo-D mRNA expression was similarly impacted by eccentric exercise during MF and ML, independent of estradiol levels in circulation. Furthermore, cyclin D1 mRNA expression was increased by eccentric exercise to a much greater extent during MF and may play a role in ER activation during periods of lower circulating estradiol. An anabolic effect of estradiol on skeletal muscle in animals has been suggested (Enns and Tiidus, 2010; Skelton et al., 1999), and this could possibly be due to a cyclin D1-induced ER activation mechanism that results in subsequent increases in Myo-D mRNA expression. Therefore, in response to eccentric exercise, this may help explain the role estradiol seems to play in the myogenic activation of satellite cells as they assist with muscle repair and regeneration during recovery (Farup et al., 2014). While circulating estradiol may confer protective benefit to the sarcolemma against exercise and/or oxidatively-induced damage, our results imply that ER activation and subsequent expression of MyoD mRNA occurs independent of the levels of estradiol in circulation.
Conclusions
In conclusion, this study demonstrates that skeletal muscle ER-α activation in response to eccentric exercise up-regulates myogenic-related gene expression independent of serum estradiol levels occurring during the human menstrual cycle.
Acknowledgements
Funding for this study was obtained from an independent research grant awarded to Baylor University from Muscle Pharm, Inc. (Denver, CO, USA). Researchers involved in collecting data in this study have no financial or personal interest in the outcome of results or the sponsor. The experiments comply with the current laws of the country in which they were performed. The authors have no conflict of interest to declare.
Biographies
Mackenzie HAINES
Employment
An undergraduate student at Baylor University majoring in Exercise Physiology
Degree
BSc
Research interest
Exercise physiology and nutrition.
E-mail: mackenzie_haines@baylor.edu
Sarah MCKINLEY-BARNARD
Employment
Assistant Professor at the University of South Alabama.
Degree
PhD
Research interest
Exercise physiology and nutrition.
E-mail: sarahmckinley@southalabama.edu
Tom ANDRE
Employment
Assistant Professor at Clayton State University.
Degree
PhD
Research interest
Exercise physiology and nutrition.
E-mail: thomasandre@clayton.edu
Josh GANN
Employment
Assistant Professor at the University of Louisiana at Monroe.
Degree
PhD
Research interest
Exercise physiology.
E-mail: gann@ulm.edu
Darryn S. WILLOUGHBY
Employment
Professor of Exercise and Nutritional Biochemistry and Molecular Physiology in the Department of Health, Human Performance, & Recreation at Baylor University.
Degree
PhD
Research interest
The biochemical and molecular regulatory mechanisms regarding exercise performance and nutrition.
E-mail: darryn_willoughby@baylor.edu
References
- Aizawa K., Iemitsu M., Otsuki T., Maeda S., Miyauchi T., Mesaki N. (2008) Sex differences in steroidogenesis in skeletal muscle following a single bout of exercise in rats. Journal of Applied Physiology 104, 67-74. [DOI] [PubMed] [Google Scholar]
- Ausubel F.M., Brent R., Kingston R.E., Morre D.D., Seidman J.G., Smith J.A., Struhl K. (1999) Short Protocols in Molecular Biology. 4th edition John Wiley & Sons; A3-10. [Google Scholar]
- Beato M. (1991) Transcriptional control by nuclear receptors. FASEB Journal 5, 2044-2051. [DOI] [PubMed] [Google Scholar]
- Beato M., Herrlich P., Schütz G. (1995) Steroid hormone receptors: many actors in search of a plot. Cell 83, 851-857. [DOI] [PubMed] [Google Scholar]
- Carter A., Dobridge J., Hackney A.C. (2001) Influence of estrogen on markers of muscle tissue damage following eccentric exercise. Fiziologii Cheloveka 27, 133-137. [PubMed] [Google Scholar]
- Di Marco S., Mazroui R., Dallaire P., Chittur S., Tenenbaum S.A., Radzioch D., Marette A., Gallouzi I.E. (2005) NF-kappa B-mediated MyoD decay during muscle wasting requires nitric oxide synthase mRNA stabilization, HuR protein, and nitric oxide release. Molecular and Cellular Biology 25, 6533-6545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ekenros L., Papoutsi A., Friden C., Dahlman Wright K., Linden Hirschgerg A. (2017) Expression of sex steroid hormone receptors in human skeletal muscle during the menstrual cycle. Acta Physiologica 219, 486-493. [DOI] [PubMed] [Google Scholar]
- Enns D.L., Tiidus P. (2008) Estrogen influenes satellite cell activation and proliferation following downhill running in rats. Journal of Applied Physiology 104, 347-353. [DOI] [PubMed] [Google Scholar]
- Enns D.L., Tiidus P.M. (2010) The influence of estrogen on skeletal muscle: sex matters. Sports Medicine 40, 41-58. [DOI] [PubMed] [Google Scholar]
- Farup J., Rahbek S.K., Riis S., Vendelbo M.H., Paoli F., Vissing K. (2014) Influence of exercise contraction mode and protein supplementation on human skeletal muscle satellite cell content and muscle fiber growth. Journal of Applied Physiology 117, 898-809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giudice L.C. (2006) Endometrium in PCOS: implantation and predisposition to endocrine CA. Best Practice in Research and Clinical Endocrinology Metabolism 20, 383-89. [DOI] [PubMed] [Google Scholar]
- Kerksick C.M., Roberts M.D., Dalbo V.J., Kreider R.K., Willoughby D.S. (2013) Changes in skeletal muscle proteolytic gene expression expression after prophylactic supplementation with EGCG and NAC and eccentric damage. Food and Chemical Toxicology 61, 47-52. [DOI] [PubMed] [Google Scholar]
- Kerksick. C., Taylor L., Harvey A., Willoughby D. (2008) Gender-related differences in muscle injury, oxidative stress, and apoptosis. Medicine and Science in Sports and Exercise 40, 1772-80. [DOI] [PubMed] [Google Scholar]
- Klein-Hitpass L., Tsai S.Y., Greene G.L., Clark J.H., Tsai M.J., O’Malley B.W. (1989) Specific binding of estrogen receptor to the estrogen response element. Molecular and Cellular Biology 9, 43-49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knight J.D., Kothary R. (2011) The myogenic kinome: protein kinases critical to mammalian skeletal myogenesis. Skeletal Muscle 1, 1-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar V., Chambon P. (1988) The estrogen receptor binds tightly to its responsive element as a ligand-induced homodimer. Cell 55, 145-156. [DOI] [PubMed] [Google Scholar]
- Li R., Ferreira M.P., Cooke M.B., La Bounty P., Campbell B., Greenwood M., Willoughby D.S., Kreider R.B. (2015) Co-ingestion of carbohydrate with branched-chain amino acids of L-leucine does not preferentially increase serum IGF-1 and expression of myogenic-related genes in response to a single bout of resistance exercise. Amino Acids 47, 1203-1213. [DOI] [PubMed] [Google Scholar]
- Longcope C. (1986) Adrenal and gonadal androgen secretion in normal females. Clinical Endocrinology and Metabolism 15, 213-228. [DOI] [PubMed] [Google Scholar]
- Mackey A.L., Kjaer M., Dandanell S., Mikkelsen K.H., Holm L., Dossing S., Kadi F, Koskinen S.O., Jensen C.H., Schroder H.D., Langberg H. (2007) The influence of anti-inflammatory medication on exercise-induced myogenic precursor cell responses in humans. Journal of Applied Physiology 103, 425-431. [DOI] [PubMed] [Google Scholar]
- McKinley-Barnard S.K., Andre T.L., Gann J.J., Hwang P.S., Willoughby D.S. (2017) The effectiveness of fish oil supplementation in attenuating exercise-induced muscle damage in females during mid-follicular and mid-luteal menstrual phases. Journal of Strength and Conditioning Research (Epub ahead of print) Sep 11. [DOI] [PubMed] [Google Scholar]
- Mikkelsen U.R., Langberg H., Helmark I.C., Skovgaard D., Andersen L.L., Kjaer M., Mackey A.L. (2009) Local NSAID infusion inhibits satellite cell proliferation in human skeletal muscle after eccentric exercise. Journal of Applied Physiology 107, 1600-1611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minahan C., Joyce S., Bulme A., Cronin N., Sabapathy S. (2015) The influence of estradiol on muscle damage and leg strength after intense eccentric exercise. European Journal of Applied Physiology 115, 1493-1500. [DOI] [PubMed] [Google Scholar]
- O’Connor K., Stip E., Pelissier M.C., Aardema F., Guay S., Gaudette G., Van Haaster I., Robillard S., Grenier S., Careau Y., Doucet P., Leblanc. V. (2007) Treating delusional disorder: a comparison of cognitive-behavioural therapy and attention placebo control. Canadian Journal of Psychiatry Reviews 52, 182-190. [DOI] [PubMed] [Google Scholar]
- Pfaffl M.W. (2001) A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Research 29, e45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reis E, Frick U., Schmidtbleicher D. (1995) Frequency variation of strength training sessions triggered by the phases of the menstrual cycle. International Journal of Sports Medicine 16, 545-550. [DOI] [PubMed] [Google Scholar]
- Roberts M.D., Dalbo V.J., Sunderland K., Poole C., Hassell S.E., Kerksick C.M. (2011). Myogenic mRNA markers in young and old human skeletal muscle prior to and following sequential exercise bouts. Applied Physiology Nutrition and Metabolism 36, 96-106. [DOI] [PubMed] [Google Scholar]
- Sabourin L.A., Rudnicki M.A. (2000) The molecular regulation of myogenesis. Clinical Genetics 57, 16-25. [DOI] [PubMed] [Google Scholar]
- Sagsoz H., Akabalik M.E., Saruhan B.G., Ketani M.A. (2010) Localization of estrogen receptor α and progesterone receptor B in bovine cervix and vagina during the follicular and luteal phases of the sexual cycle. Biotechniques in Histochemistry 86, 262-271. [DOI] [PubMed] [Google Scholar]
- Sakamaki-Sunaga M., Min S., Kamemoto K., Okamoto T. (2016) Effects of menstrual phase-dependent resistance training frequency on muscular hypertrophy and strength. Journal of Strength and Conditioning Research 30, 1727-34. [DOI] [PubMed] [Google Scholar]
- Sakurai T., Kashimura O., Kano Y., Ohno H., Ji L.L., Izawa T., Best T.M. (2013). Role of nitric oxide in muscle regeneration following eccentric muscle contractions in rat skeletal muscle. Journal of Physiological Science 63, 263-70 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skelton D.A., Phillips S.K., Bruce S.A., Naylor C.H., Woledge R.C. (1999) Hormone replacement therapy increases isometric muscle strength of adductor pollicis in post-menopausal women. Clinical Science (London) 96, 357-364. [PubMed] [Google Scholar]
- Sakamaki-Sunaga M., Min S., Kamemoto K., Okamoto T. (2015) Effects of menstrual phase-dependent resistance training frequency on muscular hypertrophy and strength. Journal of Strength and Conditioning Research 30, 1727-34. [DOI] [PubMed] [Google Scholar]
- Sung E., Han A., Hinrichs T., Vorgerd M., Manchado C., Platen P. (2014) Effects of follicular versus luteal phase-based strength training in young women. SpringerPlus 3, 668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tarnopolsky M., Phillips S., Parise G., Varbanov J., DeMuth J., Stevens P., Qu A., Wang F., Isfort R. (2007). Gene expression, fiber type, and strength are similar between left and right legs in older adults. Journal of Gerontology 62A, 1088-1095. [DOI] [PubMed] [Google Scholar]
- Thellin O., Zorzi W., Lakaye B., De Borman B., Coumans B., Hennen G., Grisar T., Igout A., Heinen E. (1999) Housekeeping genes as internal standards: use and limits. Journal of Biotechnology 75, 291-295. [DOI] [PubMed] [Google Scholar]
- Thomas A., Bunyan K., Tiidus P.M. (2010) Oestrogen receptor-alpha activation augments post-exercise myoblast proliferation. Acta Physiologica 198, 81-89. [DOI] [PubMed] [Google Scholar]
- Weisz A., Bresciani F. (1988) Estrogen induces expression of c-fos and c-myc protooncogenes in rat uterus. Molecular Endocrinology 2, 816-824. [DOI] [PubMed] [Google Scholar]
- Wilborn C.D., Taylor L.W., Greenwood M., Kreider R.B., Willoughby D.S. (2009) Effects of different intensities of resistance exercise on regulators of myogenesis. Journal of Strength and Conditioning Research 23, 2179-2187. [DOI] [PubMed] [Google Scholar]
- Willoughby D.S., Nelson M.J. (2002) Myosin heavy-chain mRNA expression after a single session of heavy-resistance exercise. Medicine and Science in Sports and Exercise 34, 1262-1269. [DOI] [PubMed] [Google Scholar]
- Zingg J.M., Alva G.P., Jost J.P. (1991) Characterization of a genomic clone covering the structural mouse MyoD1 gene and its promoter region. Nucleic Acids Research 19, 6433-6439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zwijsen R.M., Wientjens E., Klompmaker R., van der Sman J., Bernards R., Michalides R. (1997) CDK-independent activation of estrogen recptor by cyclin D1. Cell 88, 405-415. [DOI] [PubMed] [Google Scholar]