

Abstract
This is the first genetic anthropology study on Arabs in MENA (Middle East and North Africa) region. The present meta-analysis included 100 populations from 36 Arab and non-Arab communities, comprising 16,006 individuals, and evaluates the genetic profile of Arabs using HLA class I (A, B) and class II (DRB1, DQB1) genes. A total of 56 Arab populations comprising 10,283 individuals were selected from several databases, and were compared with 44 Mediterranean, Asian, and sub-Saharan populations. The most frequent alleles in Arabs are A*01, A*02, B*35, B*51, DRB1*03:01, DRB1*07:01, DQB1*02:01, and DQB1*03:01, while DRB1*03:01-DQB1*02:01 and DRB1*07:01-DQB1*02:02 are the most frequent class II haplotypes. Dendrograms, correspondence analyses, genetic distances, and haplotype analysis indicate that Arabs could be stratified into four groups. The first consists of North Africans (Algerians, Tunisians, Moroccans, and Libyans), and the first Arabian Peninsula cluster (Saudis, Kuwaitis, and Yemenis), who appear to be related to Western Mediterraneans, including Iberians; this might be explained for a massive migration into these areas when Sahara underwent a relatively rapid desiccation, starting about 10,000 years BC. The second includes Levantine Arabs (Palestinians, Jordanians, Lebanese, and Syrians), along with Iraqi and Egyptians, who are related to Eastern Mediterraneans. The third comprises Sudanese and Comorians, who tend to cluster with Sub-Saharans. The fourth comprises the second Arabian Peninsula cluster, made up of Omanis, Emiratis, and Bahrainis. It is noteworthy that the two large minorities (Berbers and Kurds) are indigenous (autochthonous), and are not genetically different from “host” and neighboring populations. In conclusion, this study confirmed high genetic heterogeneity among present-day Arabs, and especially those of the Arabian Peninsula.
Introduction
The human leukocyte antigens (HLA) system plays a key role in self-nonself recognition, and is divided into class I (HLA-A, -B, and -C) and class II (HLA-DP, -DQ, and -DR) loci, and comprises 220 genes in a 3.6 Mb region found on the short arm of chromosome 6. HLA system is highly polymorphic, and in excess of 17,000 alleles were detected. For example, there are 4,828 B, 3,968 A, and 3,579 C class I alleles, compared with 2,103 DRB1, and 1,142 DQB1 class II alleles. Several HLA alleles were associated with various auto-immune and infectious diseases [1]. HLA class I and class II loci are characterized by high (80–90%) heterozygosity, and thus constitute reliable genetic markers for phylogenetic study, and thus are useful for anthropological studies.
Population studies confirmed varied frequencies of HLA alleles and haplotypes according to ethnicity and geographic origin. Given the codominant nature of the expression of HLA markers, this enables distinguishing between heterozygotes from homozygotes, hence allowing assignment of genotypes and allele frequencies [2]. Linkage disequilibrium (LD) analysis between HLA alleles identified the number of generations in-between two closely related populations from the time of their separation. Diversity in haplotype distribution, allele frequency, and LD analysis reflect the extent of variation between closely related populations. Allele frequency-based genetic distance analysis allows for construction of phylogenetic tree (Dendrograms), so as to infer relative estimate of the time that elapsed since the populations existed as single cohesive units [3–6].
Arabs are a major panethnic group, and their union, Arab League, is a cultural and ethnic union of 22 member states. As of 2013, nationals of the Arab League countries are 357 millions, who populate an area of 13 million km2, straddling Africa and Asia [7]. Ethnic, religious, and linguistic diversity (triple heterogeneity) characterize Arabs. Most Arabs follow Islam, and Christianity is the second largest religion, with over 15 million Christians. There are also smaller but significant religious minorities (as Druze, Jews), and a number of non-Arab ethnic minorities (as Berbers, Kurds) [7, 8].
The history of Arabs extends from circa 1200 BC when Southern Arabian Peninsula was ruled by three successive civilizations: Mineans, who established their capital Karna (1200–650 BC), Sabeans in Marib (1000 BC—570 AD), and the Himyarite (2nd-6th centuries AD) in Dhafar (Oman) [9–11]. These civilizations were built by authentic Yemeni tribes. The kingdom of Kinda was established in Central Arabia in 4th-early 6th century AD, while Dilmun civilization was founded in Eastern Arabia. In 3rd century AD, East African Kingdom of Aksum extended into Yemen and Western Saudi Arabia [12]. In addition, the Lakhmids (Yemeni origin), established a dynasty which ruled part of present-day Iraq and Syria in 300–602 AD [10, 13, 14]. The Arab Christian Ghassanids (220–638 AD), originating from Southern Arabia, migrated in 3rd century to Jordan, where they established their kingdom that extended from Syria to Yathrib (Saudi Arabia)[12.13]. Islam was introduced in 610 AD to Arabian Peninsula. Shortly thereafter, Arabian tribes were united as a single Islamic state in the Arabian Peninsula, which was spear-headed by the Islamic prophet Muhammad. This Islamic state progressively grew in area, and in types and numbers of populations, and extended from Andalusia (Spain) to the west, to Indus in the east [14].
Subsequent spread of Islam involved swift invasion of Persia (637-651AD), Iraq, Levant, and Egypt (639 AD), which extended into North Africa (640–709), and to Spain, Portugal, and France (Poitiers) in 8th century AD. Eastwards, Arab expansion to Central Asia, Bukhara (Uzbekistan), Afghanistan (637–709), and the Indus border (664–712) followed. Northwards, Arab invaders were in contact with the Byzantine Empire, and the Caspian and Caucasus to the north [15, 16]. With the Islamic expansion from 7th century, social and political groups were gradually Arabized. The spreading of Arab-Muslim culture was at the expense of local languages (as Berber, Kurdish), especially in Middle East and North Africa, resulting in the Arabized population speaking variants of Arabic, mixed with original languages (dialect). The extent of gene Arab exchange with these autochthonous groups is undetermined but is thought to be lower than religious/cultural influence.
Given the large number of conquests, Arabs were in contact with different ethnicities residing on a vast area stretching from Mauritania (West Africa) to the western China border (East Asia). This suggests that cultural and perhaps genetic relationships were established with these ethnic groups. This work aims to study the HLA distribution in North African and Oriental Arab populations, and compare them to neighboring populations (Sub-Saharans Africans, Europeans, and Asians).
Populations and methods
Search strategy
Datasets of HLA allele frequencies were collected from a systematic review performed per Preferred Reporting Items for Systematic Reviews and Meta-analyses (PRISMA) criteria [Only the criteria from 1–10, 17, and 26 are applicable to this type of study (S1 Checklist)] [17]. PubMed, ScienceDirect, AlleleFrequencies.net, and ResearchGate databases were searched for all papers on HLA polymorphism, and HLA disease associations in Arabs. This systematic literature search covering published papers up to May 31, 2017 was conducted by two investigators (H.A and H.L); the search terms used were: ‘HLA Arabs’, or ‘Human Leukocyte Antigen Arabs’. A search per country followed: ‘HLA Tunisians’, ‘HLA Saudis’, and so on. This was repeated for remaining countries, which resulted in excess of 50 keywords used. A database from International Histocompatibility Workshops was also used. Some authors were also contacted by e-mail, or through ResearchGate, requesting information and missing data. While most datasets were taken from studies with an explicit anthropological focus, control groups from case-control disease studies were also used. There was no language restriction used for this search.
Inclusion and exclusion criteria
All included studies met the following criteria. HLA allele frequencies must be obtained by molecular typing, and that subjects should be typed for at least one of the following: HLA-A, HLA-B, HLA-DRB1, and HLA-DQB1. Publications were excluded in case of serological data; sample size less than 35 individuals, typed individuals (or controls) were either related and not randomly selected, presentation of duplicate data sets. Studies were also excluded if they presented incomplete/partial allele frequencies, or there were significant ambiguities in the typing.
Data extraction
Studies were independently selected by two authors (H.A and H.L). An external referee was invited in case of disagreements not resolved by both reviewers. Data extracted from selected papers included publication year, study type (anthropology, association), sample size, HLA-A, -B, -C, -DRB1, and -DQB1 allele frequencies, haplotype frequencies, region, country, and typed loci.
Statistical analysis
A three-dimensional correspondence analysis and bi-dimensional representation were performed using VISTA V5.02 software [18]. Phylogenetic trees were constructed based on allele frequencies using the Neighbor-Joining (NJ) method [19], and standard genetic distances (SGD) [20], using DISPAN software containing GNKDST and TREEVIEW software [21, 22].
Results
Study flow
The use of more than fifty key words allowed identification of 5,456 papers and HLA datasets, of which 315 were deemed relevant to the study. Of these, 42 articles and 11 HLA datasets containing information on 56 Arab populations, and meeting the study criteria, were included. The study flow is illustrated in Fig 1. In addition, 20 articles and 18 HLA datasets which meet the criteria of this study, containing complete information on 44 other populations were selected, but without going through systematic review. The populations used in the comparison were chosen mainly from neighboring Arab countries. This study relied on a database consisting of 100 populations (of which data of 11 populations were extracted from association studies) from 36 countries Arab and worldwide countries, and belonging to Asia, Europe, and Africa. The distribution of populations by region is illustrated in Fig 2A. These populations represent allele frequency data for 16,006 individuals (160.06 individuals/population), and from 63 references.
Fig 1. Flow diagram of the study selection process.
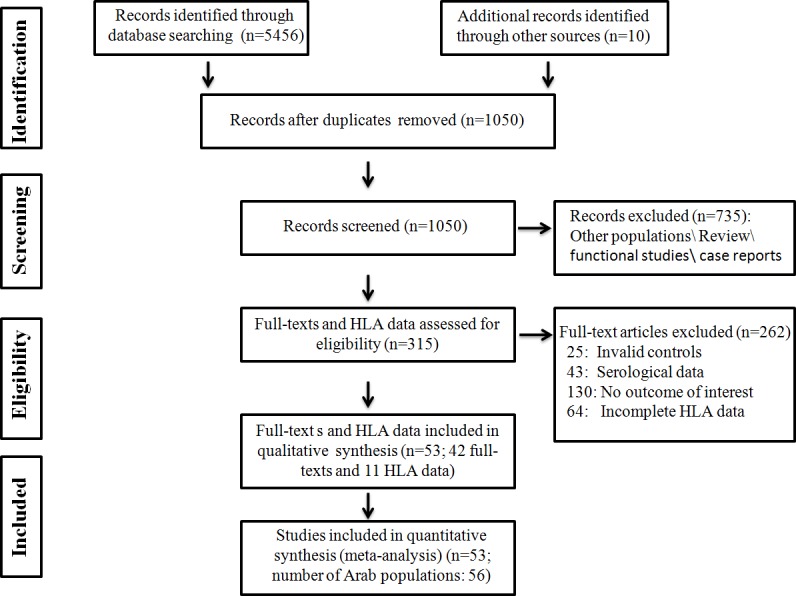
Fig 2. The distribution of studied populations by region (A) and country (B).
Selected populations
Arab populations
The 42 articles and 11 HLA datasets (http://www.allelefrequencies.net) selected provided information on 56 populations (Table 1), comprising 10,283 individuals [23–67]. The 56 different ethnic and religious populations were selected from 18 Arab countries. There were no reliable HLA data for the remaining countries (Somalia, Djibouti, Mauritania, and Qatar) (Fig 2B). The studied populations are divided into 29 African (26 North Africans and 3 Sub-Saharans), and 27 Asian populations (13 Levantines, and 14 Arabian Peninsula). With the exception of 8 populations [28, 38, 47, 48, 50, 52, 53, 55], where HLA data were extracted from association studies, the 50-remaining studies were extracted from anthropological ones.
Table 1. List of Arab populations used in the present work.
| No | Populations | Symbols | Size | References | No | Populations | Symbols | Size | References |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Algiers | Alg | 102 | [67] | 29 | Comorians | Com | 117 | [43] |
| 2 | Algerians-B | Alg-B | 97 | [23] | 30 | Jordanians | Jor | 146 | [31] |
| 3 | Algerians-A | Alg-A | 132 | [48] | 31 | Jordanians-A | Jor-A | 1254 | [46] |
| 4 | Algerians-Oran | Ora | 100 | [23] | 32 | Syrians | Syr | 200 | [47] |
| 5 | Gabesians | Gab | 77 | [59] | 33 | Syrians-A | Syr-A | 225 | [58] |
| 6 | Gabesians-A | Gab-A | 96 | [40] | 34 | Lebanese | Leb | 95 | [35] |
| 7 | Ghannouchians | Gha | 82 | [33] | 35 | Lebanese-A | Leb-A | 1123 | [45] |
| 8 | Berbers-Jerba | Ber-J | 55 | [40] | 36 | Lebanese-B | Leb-B | 191 | [44] |
| 9 | Berbers-Matmata | Ber-M | 81 | [40] | 37 | Lebanese-Armen | Leb-Ar | 368 | [27] |
| 10 | Berbers-Zrawa | Ber-Z | 70 | [24] | 38 | Lebanese-KZ | Leb-Kz | 93 | [39] |
| 11 | Tunisians | Tun | 376 | [61] | 39 | Lebanese-NS | Leb-Ns | 59 | [39] |
| 12 | Tunisians-A | Tun-A | 80 | [60] | 40 | Lebanese-Yohmor | Leb-Y | 75 | [39] |
| 13 | Tunisians-B | Tun-B | 101 | [34] | 41 | Palestinians | Pal | 165 | [29] |
| 14 | Tunisians-C | Tun-C | 100 | [63] | 42 | Palestinians-A | Pal-A | 109 | [36] |
| 15 | Tunisians-M | Tun-M | 123 | [26] | 43 | Saudis | Sau | 105 | [28] |
| 16 | Southern Tunisians | Tun-S | 250 | [62] | 44 | Saudis-A | Sau-A | 213 | [23] |
| 17 | Libyans | Lib | 118 | [32] | 45 | Saudis-B | Sau-B | 158 | [49] |
| 18 | Libyans-Jews | Lib-J | 119 | [36] | 46 | Saudis-C | Sau-C | 499 | [23] |
| 19 | Berbers-Metelsa | Ber-Me | 99 | [64] | 47 | Saudis-D | Sau-D | 383 | [50] |
| 20 | Moroccans | Mor | 96 | [25] | 48 | Omanis-A | Oma-A | 259 | [30] [51] |
| 21 | Moroccans-A | Mor-A | 110 | [42] | 49 | Kuwaitis | Kuw | 212 | [52] |
| 22 | Moroccans-Agadir | Mor-Ag | 98 | [37] | 50 | Kuwaitis-A | Kuw-A | 114 | [53] |
| 23 | Moroccans-Chaouya | Mor-Ch | 98 | [65] | 51 | Bahrainis | Bah | 72 | [35] |
| 24 | Moroccans-Jews | Mor-J | 94 | [66] | 52 | Emiratis | Emi | 373 | [23] |
| 25 | Egyptians | Egy | 101 | [39] | 53 | Iraq kurds | Ira-K | 209 | [54] |
| 26 | Egyptians-A | Egy-A | 121 | [38] | 54 | Yemenite-Jews | Yem-J | 76 | [36] |
| 27 | Sudanese | Sud | 200 | [23] | 55 | Yemen-sana'a | Yem | 50 | [55] |
| 28 | Sudanese-Nuba | Sud-N | 46 | [23] | 56 | Omanis | Oma | 118 | [56] [57] |
Neighboring populations
Forty-four worldwide populations [23, 34, 39, 66, 68–85] comprising 5,723 individuals, were selected from 18 countries in three continents, using the same criteria previously described (Table 2). These comprised 22 European, 11 non-Arab Asian, and 11 Sub-Saharan African populations. Of the 11 Asian populations, there were two Arab minorities living in Iran (Khuzestan and Famoori).
Table 2. Worldwide populations included in the meta-analysis.
| No | Populations | Symbols | Size | References | No | Populations | Symbols | Size | References |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Spaniards | Spa | 176 | [41] | 23 | Mossi | Mos | 42 | [39] |
| 2 | Portuguese | Por | 118 | [39] | 24 | Mandenka | Mad | 200 | [39] |
| 3 | Murcians | Mur | 173 | [80] | 25 | Amhara | Amh | 98 | [39] |
| 4 | Italians | Ita | 284 | [68] | 26 | Bubi | Bub | 101 | [39] |
| 5 | Basques-A | Bas-A | 82 | [41] | 27 | Congolese | Con | 85 | [72] |
| 6 | Basques-Arratia | Bas-Ar | 83 | [77] | 28 | Fulani | Ful | 38 | [39] |
| 7 | Basques-B | Bas-B | 99 | [70] | 29 | Gabonese | Gab | 167 | [85] |
| 8 | French | Fre | 179 | [68] | 30 | Nigerians | Nig | 258 | [23] |
| 9 | French-Rennes | Fre-R | 200 | [34] | 31 | Oromo | Oro | 83 | [39] |
| 10 | Balearic | Bal | 90 | [71] | 32 | Rimaibe | Rim | 39 | [39] |
| 11 | Corsica | Cor | 100 | [71] | 33 | Senegalese | Sen | 177 | [39] |
| 12 | Sardinians | Sar | 91 | [68] | 34 | Famoori Arabs | Fam | 84 | [73] |
| 13 | Ashkenazi-Jews | Ash-J | 132 | [66] | 35 | India-Northeast | Ind-N | 188 | [83] |
| 14 | Greeks-A | Gre-A | 96 | [39] | 36 | Indians-Delhi | Ind-D | 112 | [84] |
| 15 | Greeks-B | Gre-B | 101 | [39] | 37 | Iranian-Jews | Ira-J | 91 | [73] |
| 16 | Greeks-C | Gre-C | 98 | [39] | 38 | Iranians | Ira | 120 | [74] |
| 17 | Greeks-D | Gre-D | 242 | [23] | 39 | Iranians-A | Ira-A | 100 | [75] |
| 18 | Macedonians | Mac | 172 | [78] | 40 | Iranians-Azeri | Ira-Az | 100 | [81] |
| 19 | Turks | Tur | 250 | [23] | 41 | Iranians-Kurd | Ira-k | 100 | [81] |
| 20 | Turks-A | Tur-A | 228 | [79] | 42 | Khuzestani Arabs | Khu | 50 | [73] |
| 21 | Albanians | Alb | 160 | [76] | 43 | Pakistanis-Pathan | Pak-P | 100 | [82] |
| 22 | Cretans | Cre | 135 | [69] | 44 | Pakistanis-Sindh | Pak-S | 101 | [82] |
Data of only three populations [74, 75, 84] were extracted from association studies. These populations were typed for at least HLA-A, -B, -DRB1, or DQB1.
HLA allele frequencies features of Arab populations
Table 3 shows the most frequent HLA-A and -B alleles in Arab populations. A*02 was the most prevalent allele, and its frequency exceeded 25% in some populations, such as Saudis (30.4%) [23], Tunisian Berbers of Zrawa (29.3%) [24], Moroccans (26.2%) [25], and Sudanese (25.9%) [23]. A*01, *03, *24, *30, and *68 alleles were also common in most Arab populations. For example, the highest frequency of A*01 was seen in Tunisians (15%) [26] and Moroccans (14.8%) [25], while A*03 was prevalent among Iraqi Kurds (15.1%) [23], and A*30 was prevalent among Sudanese (17.6%) [23]. In addition, A*24 was common among Lebanese-Armenians (17.3%) [27], while A*68 was prevalent in Saudis (10.5%) [28]. In contrast, A*25, *28, *34, *36, *43, *66, *69, *74, and *80 are rare among Arabs. It is noteworthy that A*34, described as rare allele among Arabs, is found at a high frequency (22.2%) in Tunisian Berbers from Zrawa [24], the highest reported for any population worldwide.
Table 3. Most frequent HLA-A* and–B* alleles in Arab populations.
| HLA-A | A*01 | A*02 | A*03 | A*24 | A*30 | A*68 | ||||||
| Population | % | Population | % | Population | % | Population | % | Population | % | Population | % | |
| Tun-M | 15.0 | Sau-D | 30.4 | Ira-k | 15.1 | Leb-Ar | 17.3 | Sud | 17.6 | Sau | 10.5 | |
| Mor | 14.8 | Ber-Z | 29.3 | Leb-Ar | 14.0 | Gha | 15.2 | Mor-C | 13.0 | Tun-M | 09.4 | |
| Jor-A | 14.7 | Mor | 26.2 | Pal | 10.7 | Ira-k | 13.9 | Tun-A | 11.8 | Mor | 09.3 | |
| Ira-k | 13.2 | sud | 25.9 | Lib | 10.3 | Sau-B | 13.3 | Jor | 11.5 | Alg-K | 08.6 | |
| Pal | 12.5 | Emi | 25.2 | Mor-A | 10.0 | Jor-A | 10.7 | Alg-K | 10.2 | sud | 08.5 | |
| Leb-A | 12.2 | Oma | 24.9 | Alg-K | 09.3 | Pal | 10.1 | Sau-B | 10.2 | Emi | 08.4 | |
| Sau-A | 12.2 | Alg | 24.6 | Jor-A | 09.1 | Alg | 09.4 | Pal | 08.4 | Lib | 08.2 | |
| Alg | 11.9 | Lib | 23.5 | Emi | 09.1 | Lib | 09.3 | Oma-A | 07.5 | Jor | 07.6 | |
| Lib | 11.5 | Jor-A | 22.0 | Sau-A | 08.9 | Mor | 07.3 | Leb-A | 06.7 | Oma-A | 07.1 | |
| Oma | 07.2 | pal | 20.5 | Gab | 07.7 | Oma | 06.3 | Lib | 06.4 | Leb-A | 05.1 | |
| Sud | 06.5 | Leb-A | 18.7 | Sud | 07.1 | Sud | 06.1 | Emi | 05.0 | Ira-k | 03.8 | |
| Emi | 06.2 | Ira-k | 17.0 | Oma | 06.4 | Emi | 05.2 | Ira-k | 03.8 | Pal | 03.6 | |
| HLA-B | B*07 | B*08 | B*35 | B*44 | B*50 | B*51 | ||||||
| Population | % | Population | % | Population | % | Population | % | Population | % | Population | % | |
| Jor | 27.1 | Oma | 11.0 | Pal | 20.3 | Ber-Z | 32.8 | Sau-D | 18.8 | Sau-C | 19.3 | |
| Sau-A | 11.7 | sau-B | 10.1 | Leb-Ar | 19.8 | Ira-k | 10.3 | Lib | 16.1 | Oma | 17.5 | |
| Mor | 09.0 | Emi | 08.6 | Ira-k | 15.6 | Mor-C | 10.2 | Ber-Z | 15.7 | Emi | 156 | |
| Lib | 07.7 | Gha | 08.5 | Oma-A | 15.3 | pal | 09.6 | Tun-S | 14.2 | Ira-K | 15.6 | |
| Tun-A | 07.5 | Ira-k | 07.2 | Jor-A | 14.9 | Alg | 08.8 | Mor-C | 12.5 | Gha | 12.2 | |
| Alg-k | 07.1 | Lib | 06.4 | Emir | 11.1 | Leb-Ar | 08.4 | Emi | 09.4 | Leb-Ar | 12.1 | |
| Leb-Ar | 04.5 | Mor-C | 06.2 | Alg | 10.3 | Lib | 07.6 | Jor-A | 06.4 | Lib | 11.1 | |
| Ira-k | 04.1 | Jor | 04.7 | Lib | 10.1 | Jor-A | 05.6 | Pal | 05.8 | Jor-A | 10.3 | |
| Oma-A | 03.1 | Sud | 04.0 | Tun-M | 09.8 | Sau-D | 03.5 | Leb-Ar | 05.2 | Sud | 07.8 | |
| Sud | 02.8 | Alg | 03.5 | Sau | 08.6 | Sud | 02.3 | Alg | 05.1 | Mor | 07.4 | |
| Emi | 02.4 | Leb-Ar | 03.0 | Mor-C | 06.9 | Emi | 02.3 | Oma-A | 04.2 | Pal | 06.4 | |
| Pal | 01.8 | Pal | 02.7 | sud | 06.1 | Oma-A | 02.1 | Sud | 02.5 | Alg-k | 04.7 | |
Only one population per country is illustrated; the frequencies are ranked from highest to lowest for each allele; to identify the population and country see Table 1
Results of HLA-B locus are presented in Table 3. B*35 was the most frequent B* allele in Palestinians (20.3%) [29] and Lebanese-Armenians (19.8%) [27]. B*35 was found at varied frequencies in Iraqi Kurds (15.6%) [23], Omanis (15.3%) [30], Jordanians (14.9%) [31], and Arab Emirati (11.1%) [23] populations. B*51 was the second most frequent allele, and high frequencies were recorded for Saudis (19.3%) [23], Omanis (17.5%) [30], and Arab Emirati (15.6%) [23] populations. B*50 was also a frequent B* allele in most Arabs, including Saudis (18.8%) [23], and Libyans (16.1%) [31], along with B*08, and B*44 among the Tunisian Berbers of Zrawa (32.8%) [24], the latter being the highest frequency worldwide. Similarly, the frequency of B*27 is the highest among Jordanians (27.1%) [31]. In contrast, B*37, *42, *46, *47, *48, *54, *59, *67, and *78 alleles are extremely rare or virtually in all Arab populations.
The most common DRB1 and DQB1 alleles among Arabs are shown in Table 4. DRB1*07:01 was the most frequent allele among Tunisians from Ghannouch (28.6%) [33], Jordanians (26.9%) [31], and Saudis (26.6%) [23], while Egyptians (8.3%) and Sudanese had the lowest frequencies of DRB1*07:01. DRB1*03:01 was the second most frequent DRB1* allele in some Arabs, such as Tunisians of Tunis (21.9%) [34] and Moroccans of Metelsa (20.2%) [23], but rare in Jordanians (2.4%) [31]. DRB1*11:01 was also frequent among some Arabs, such as Lebanese (36.8%) [35], but rare among Saudis (4.8%) and Moroccans of Chayoua (2.5%) [23]. Furthermore, DRB1*13:01, *13:02, and *15:01 alleles are relatively frequent among Arabs. High frequency of DRB1*13:01 were recorded for Sudanese (23.3%), while DRB1*13:02 was virtually absent in Bahraini [35] and Sudanese [23]. All DRB1*09, *12, and *14 subtypes are extremely rare among Arabs. In addition, DRB1*16 subtypes are rare in all Arab populations except for Bahrain, where DRB1*16:01 is found at a high frequency (13.9%) [35].Haut du formulaire.
Table 4. Most frequent HLA-DRB1* and–DQB1* alleles in Arab populations.
| HLA-DRB1* | 03:01 | 07:01 | 11:01 | 13:01 | 13:02 | 15:01 | ||||||
| Population | % | Population | % | Population | % | Population | % | Population | % | Population | % | |
| Tun-B | 21.9 | Gha | 28.6 | Leb | 36.8 | Sud | 23.3 | Mor-Me | 11.1 | Alg-B | 13.4 | |
| Mor-Me | 20.2 | Jor | 26.9 | Bah | 16.0 | Sau-A | 10.6 | Lib | 09.3 | Mor-c | 12.6 | |
| Sau-B | 16.5 | Sau-B | 26.6 | Egy-A | 13.2 | Ber-M | 08.0 | Sau-A | 08.9 | Ber-Z | 11.4 | |
| Ora | 15.1 | Yem-J | 22.1 | Gab-A | 11.2 | Leb-B | 06.8 | Egy-A | 07.4 | Jor | 09.0 | |
| Bah | 13.9 | Mor-Ag | 20.5 | Pal | 10.0 | Alg-B | 05.6 | Tun-C | 06.7 | Sau-A | 08.9 | |
| Sud | 13.8 | Lib-Y | 19.6 | Ora | 08.6 | Lib | 05.5 | Leb-N | 05.0 | Bah | 07.6 | |
| Lib | 13.6 | Lib | 17.0 | Sud | 08.3 | Yem-J | 05.4 | Ora | 04.5 | Leb | 04.7 | |
| Yem-J | 12.0 | Alg-B | 15.9 | Jor | 08.3 | Egy-A | 04.6 | Yem-J | 04.0 | Lib | 04.2 | |
| Leb-B | 09.6 | Pal | 12.7 | Lib | 05.1 | Mor-Me | 03.5 | Pal | 03.9 | Pal | 03.6 | |
| Pal | 07.6 | Bah | 09.0 | Sau-A | 04.8 | Jor | 02.1 | Jor | 00.3 | Sud | 03.3 | |
| Egy-A | 07.0 | Egy-A | 08.3 | Yem-J | 03.4 | Bah | 02.1 | Sud | 00.0 | Egy-A | 02.5 | |
| Jor | 02.4 | Sud | 07.8 | Mor-C | 02.5 | Pal | 00.9 | Bah | 00.0 | Yem-J | 02.0 | |
| HLA-DQB1* | 02:0X | 03:01 | 03:02 | 05:01 | 06:02 | 06:03 | ||||||
| Population | % | Population | % | Population | % | Population | % | Population | % | Population | % | |
| Gha | 40.1 | Leb-NS | 45.0 | Gha | 20.7 | Bah | 29.2 | Mor-C | 12.9 | Egy-A | 10.2 | |
| Yem-J | 39.1 | Ora | 35.1 | Jor | 17.8 | Ber-J | 22.7 | Alg | 12.8 | Jor | 08.3 | |
| Mor-Ag | 37.8 | Lib-J | 29.6 | Pal | 17.6 | Leb | 20.5 | Egy-A | 12.7 | Ber-J | 07.8 | |
| Sau-B | 37.3 | Ber-J | 27.4 | Leb | 16.8 | Alg | 13.9 | Tun-A | 12.6 | Lib-J | 07.4 | |
| Jor | 35.9 | Pal | 26.7 | Yem-J | 14.2 | Mor-C | 12.3 | Jor | 10.7 | Yem-J | 06.1 | |
| Lib-J | 33.3 | Yem-J | 19.1 | Lib-J | 13.0 | Pal | 11.8 | Sau-B | 05.1 | Ora | 04.3 | |
| Bah | 25.7 | Bah | 16.0 | Alg | 12.3 | Sau-B | 10.1 | Pal | 04.2 | Sau-B | 04.1 | |
| ora | 24.5 | Mor-C | 15.4 | Mor-C | 12.3 | Jor | 09.3 | Leb-Y | 03.7 | Leb-Y | 03.3 | |
| Pal | 20.9 | Egy | 11.9 | Bah | 09.7 | Egy-A | 08.5 | Yem-J | 02.0 | Mor-C | 01.8 | |
| Leb-Y | 20.0 | Jor | 10.0 | Sau-B | 08.9 | Yem-J | 06.1 | Lib-J | 00.8 | Pal | 01.2 | |
Only one population per country is illustrated; the frequencies are ranked from highest to lowest for each allele; to identify the population and country see Table 1
DQB1*02:0X and *03:01 alleles are the most frequent DQB1* in Arabs. The highest frequencies of DQB1*02:0X were reported for Tunisians (Ghannouch; 40.01%) [33], Yemenites-Jews (39.1%) [36], Moroccans (Agadir-Souss; 37.8%) [37] and Saudis (37.3%) [23], while the lowest frequency was found in Egyptians (6%) [38]. On the other hand, DQB1*03:01 is very common among Lebanese (45%) [39] and Algerians (Oran; 35.1%) [23], but not Saudis (7.6%) [23]. DQB1*03:02 and *05:01 are also frequent in most Arabs, such as Tunisians (Ghannouch; 20.7%) [33], Jordanians (17.8%) [31], Palestinians (17.6%) [29] and Lebanese (16.8%) [35]. DQB1*05:01 is frequent among Bahrainis (29.2%) [35], Tunisians (Berbers of Jerba; 22.7%) [40], and Lebanese (20.5%) [35]. Among DQB1*06 subtypes, DQB1*06:02 and *06:03 were the most frequent in most Arab populations, but absent in Bahrainis where DQB1*06:01 is very frequent (13.20%) [35]. Furthermore, all DQB1*04 subtypes are rare among Arabs, particularly DQB1*04:01 which is virtually absent, except in Egyptians (10.17%) [38]. The most common DQB1*04 subtype in Arabs is DQB1*04:02.
Allelic comparison between Tunisians and other populations
Allelic comparisons were done at Neighbor-Joining, correspondence analysis, and standard genetic distances. Analyses were performed with Class I and Class II markers, and at generic and high-resolution levels to make the most of available data, and seeing that some of the populations included in these comparisons lack high-resolution data.
Neighbor-joining dendrograms
Comparison at the generic level was made using genetic distances based on DRB1* and DQB1* allelic frequencies. Four groups can be interpreted from Fig 3. The first group comprises North African Arabs (Tunisians, Algerians, Moroccans, Libyans), Western Mediterranean Europeans (Iberians, French), Arabian Peninsula Arabs (Saudis, Kuwaitis, Yemenis), and Arab minority of Iran (Khuzestani). The second group is formed by Eastern Mediterranean Europeans (Greeks, Cretans, Albanians, Turks, Macedonians), Italians, Levant Arabs (Palestinians, Lebanese, Syrians), Iraqi-Kurds, Tunisian Berbers (Djerba), and Iranians. The third group comprises Sub-Saharan Africans (Fulani, Mossi, Rimaibe, Bubi, Mandenka, and Senegalese). Omanis, Bahrainis, Egyptians, and Sudanese form a heterogeneous group containing Asians and Sub-Saharan Africans. Similar results but with notable differences, were observed in dendrograms built with standards genetic distances (SGD) based on generic DRB1(S1 Fig) and generic B loci (S2 Fig).
Fig 3. Neighbor-Joining dendrograms, based on Standard genetic distances (SGD), showing relatedness between Arabs and other populations using generic HLA-DRB1* and -DQB1* allele frequencies data.

Populations’ data were taken from references detailed in Tables 1 and 2. Bootstrap values from 1.000 replicates are shown.
Correspondence analysis
High-resolution DRB1 correspondence analysis (Fig 4) demonstrated the clustering of the studied populations into three groups. The first containing North Africans (Tunisians, Algerians, Moroccans, and Libyans), Iberians (Basques, Spaniards, Portuguese, Murcians), French, Saudis, Yeminis-Jews, and Khuzestani Arabs. The second contains Eastern Mediterraneans (Greeks, Cretans, Lebanese, Palestinians, and Macedonians), Berbers of Djerba, Italians, Iraqi-Kurds, Iranians, Egyptians, Ashkenazi-Jews, and Moroccan-Jews. The last cluster consists of Sub-Saharan populations. It should be noted that Jordanians, Bahrainis, and Sudanese were outside these main groups. Similarly, correspondence analysis using class I (A and B) identified three main clusters (Fig 5). The first cluster contained all Sub-Saharan Africans along with Sudanese. The second cluster contains Eastern Mediterranean populations (Albanians, Greeks, Cretans, Lebanese, Palestinians, and Macedonians), Italians, Iraqi-Kurds, Ashkenazi-Jews, and Jordanians-A. The last cluster includes North Africans (Tunisians, Algerians, Moroccans, and Libyans), Iberians (Basques, Spaniards), French, and Saudis.
Fig 4. Correspondence analysis (bi-dimensional representation), based on the standard genetic distances, showing the relationship between Arabs and other populations according to high resolution HLA-DRB1* allele frequencies data.
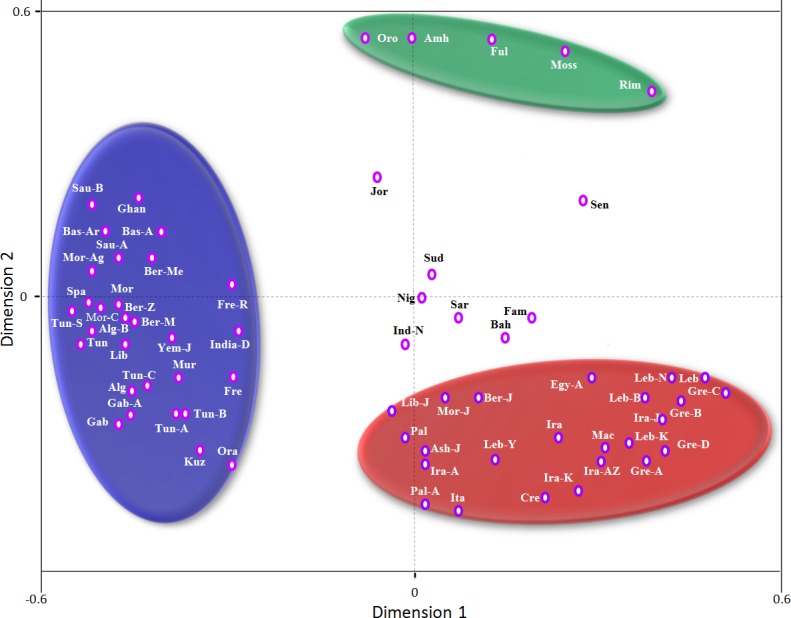
Only individuals with defined DRB1* subtypes are considered. Populations data were taken from references detailed in Tables 1 and 2.
Fig 5. Correspondence analysis (bi-dimensional representation), based on the standard genetic distances, showing a global view of the relationship among Arabs and other populations according to generic HLA*-A and–B* allele frequencies data.

Populations data were taken from references detailed in Tables 1 and 2.
Correspondence analysis based on generic DRB1 data, and using only Arab populations shows that Arabs can cluster into four groups (Fig 6). The first contains the North Africans (Tunisians, Algerians, Moroccans, and Libyans), Saudis, Yemenis, Kuwaitis, and Khuzestanis (Iranian Arabs). The second cluster includes the Arabs of Levant (Palestinians, Jordanians, Lebanese, Syrians), Egyptians, Iraqi Kurds, and Moroccans Jews. The third group consists of Bahrainis, Omanis, Emiratis and Famoori (Iranian Arab). The fourth is composed of Sudanese, Sudanese from Nuba, and Comorians.
Fig 6. Correspondence analysis (bi-dimensional representation), based on the standard genetic distances, showing the relationship between different Arab populations according to generic HLA-DRB1* allele frequencies data.

Populations data were taken from references detailed in Tables 1 and 2.
Genetic distances
Table 5 illustrates standard genetic distances (SGD) between Arabs and other populations, using generic DRB1* allele frequencies. North Africans and Iberians are the closest to Saudis. Moroccans (Agadir, 0.0024), Basques-Ar (0.0057), and Tunisians-S.
Table 5. The closest populations to Arabs using standard genetic distances (SGD) based on HLA-DRB1* alleles.
| Saudis-B | Emiratis | Omanis-A | Sudanese | ||||
| Population | SGD | Population | SGD | Population | SGD | Population | SGD |
| Moroccans-Ag | 0.0024 | Omanis-A | 0.0411 | Emirates | 0.0411 | Nigerians | 0.0497 |
| Basques-Ar | 0.0057 | Bahrain | 0.0429 | Sardinians | 0.0939 | Egyptians-A | 0.0556 |
| Tunisians-S | 0.0124 | Sardinians | 0.0593 | Bahrain | 0.1327 | Congolese | 0.0594 |
| Saudis-C | 0.0160 | Kuwaitis | 0.0688 | Kuwait | 0.2014 | Egyptians | 0.0620 |
| Ghanouchians | 0.0203 | Tunisians-B | 0.1169 | Famoori Arabs | 0.2377 | Mandenka | 0.0908 |
| Saudis | 0.0258 | Khuzestanis | 0.1213 | Macedonians | 0.2461 | Moroccans | 0.0984 |
| Tunisians | 0.0272 | Tunisians-A | 0.1276 | Tunisians-B | 0.3071 | Senegalese | 0.1044 |
| Kuwaitis-A | 0.0312 | Algerians-Oran | 0.1371 | Khuzestanis | 0.3192 | Bubi | 0.1078 |
| Khuzestanis | 0.0349 | Algerians-A | 0.1407 | Greeks-B | 0.3197 | Palestinians-A | 0.1111 |
| Spaniards | 0.0354 | Algerians-B | 0.1612 | Tunisians-A | 0.3261 | Pakistanis-S | 0.1122 |
| Saudis-D | 0.0374 | Algiers | 0.1639 | Kuwaitis-A | 0.3544 | Tunisians-A | 0.1133 |
| Gabesians | 0.0377 | Saudis-C | 0.1746 | Algerians-Oran | 0.3600 | Libyans | 0.1197 |
| Gabesians-A | 0.0394 | Macedonians | 0.1756 | Algerians-A | 0.3639 | Sudanese-Nuba | 0.1234 |
| Jordanians | 0.0428 | Gabesians | 0.1820 | Greeks-D | 0.3657 | Algerians-B | 0.1315 |
| Algerians-B | 0.0433 | Saudis-D | 0.1820 | Algerians-B | 0.3867 | Berbers-Matmata | 0.1317 |
| Basques-B | 0.0449 | Moroccans-Agadir | 0.1830 | Greeks-C | 0.3927 | Algerians-A | 0.1407 |
| Saudis-A | 0.0450 | Kuwaitis-A | 0.1837 | Turks | 0.3944 | Berbers-Zrawa | 0.1409 |
| Algerians-A | 0.0497 | Famoori Arabs | 0.1894 | Saudis-C | 0.3984 | Gabesians | 0.1413 |
| Tunisians-C | 0.0533 | Moroccans-A | 0.1900 | Algiers | 0.4027 | Jordanians-A | 0.1434 |
| Yemenite-J | 0.0536 | Gabesians-A | 0.1908 | Albanians | 0.4034 | Gabesians-A | 0.1442 |
| Khuzestanis | Tunisians | Syrians-A | Comorians | ||||
| Population | SGD | Population | SGD | Population | SGD | Population | SGD |
| Gabesians | -0.0086 | Gabesians | -0.0139 | Cretans | -0.0001 | Congolese | 0.0519 |
| Orans | -0.0074 | Gabesians-A | -0.0081 | Lebanese-Ar | 0.0050 | Nigerians | 0.0828 |
| Gabesians-A | -0.0025 | Moroccans-Agadir | -0.0080 | Syrians | 0.0076 | Greeks-A | 0.0836 |
| Algerians-A | -0.0015 | Southern Tunisians | -0.0062 | Iranians-Kurd | 0.0100 | Gabonese | 0.0904 |
| Moroccans-Ag | 0.0106 | Algerians-A | -0.0055 | Lebanese-A | 0.0149 | Iranians-A | 0.0947 |
| Tunisians-S | 0.0140 | Moroccans-A | 0.0010 | Lebanese-Y | 0.0151 | Egyptians-A | 0.1090 |
| Tunisians | 0.0161 | Algerians-B | 0.0019 | Iranians | 0.0159 | Iranians | 0.1184 |
| Tunisians-C | 0.0195 | Berbers-Zrawa | 0.0027 | Lebanese-B | 0.0161 | Italians | 0.1222 |
| Yemenite-J | 0.0217 | Libyans | 0.0028 | Iranians-Azeri | 0.0185 | Iranians-Azeri | 0.1394 |
| Tunisians-M | 0.0225 | Algerians-Oran | 0.0033 | Turks | 0.0192 | Iranians-Kurd | 0.1418 |
| Saudis-C | 0.0231 | Tunisians-M | 0.0038 | Iraq kurdistan | 0.0198 | Albanians | 0.1426 |
| Spaniards | 0.0291 | Saudis-C | 0.0061 | Ashkenazi-Jews | 0.0222 | Turks | 0.1428 |
| Saudis | 0.0324 | Tunisians-C | 0.0083 | Iranians-A | 0.0223 | Syrians | 0.1470 |
| Saudis-B | 0.0349 | Algiers | 0.0103 | Palestinians-A | 0.0228 | Cretans | 0.1483 |
| Algerians-B | 0.0353 | Berbers-Matmata | 0.0106 | Italians | 0.0241 | Egyptians | 0.1483 |
| Tunisians-B | 0.0422 | Moroccans-Chaouya | 0.0111 | Turks-A | 0.0288 | Greeks-C | 0.1487 |
| Indians-Delhi | 0.0454 | Spaniards | 0.0126 | Lebanese | 0.0320 | Palestinians-A | 0.1559 |
| Algiers | 0.0461 | Moroccans | 0.0144 | Jordanians-A | 0.0355 | Iraq Kurdistan | 0.1564 |
| Basques-Ar | 0.0471 | Saudis-D | 0.0159 | Lebanese-KZ | 0.0368 | Greeks-D | 0.1594 |
| Libyans | 0.0485 | Khuzestani Arabs | 0.0161 | Greeks-A | 0.0407 | Syrians-A | 0.1617 |
(0.0124) had the closest genetic distances from Saudis, while Emiratis were closely related to Omanis (0.0411), Bahrainis (0.0429), Sardinians (0.0593), and Kuwaitis (0.0688). On the other hand, Sudanese are related to Sub-Saharans, including Nigerians (0.0497), Congolese (0.0594), and Egyptians (0.556).
Syrians are genetically close to Eastern Mediterranean, as Cretans (-0.0001) and Lebanese Armenians (0.0050), while Tunisians are closed to Western Mediterraneans as North Africans and Iberians, and Saudis. The populations most related to Tunisians are the other Tunisian populations (Gabesians, -0.0139), Moroccans (Agadir; -0.0080), and Algerians (-0.0055). Sub-Saharans such as Congolese (0.0519) and Nigerians (0.0828), and Greeks (0.0836) showed the closest genetic distances to Comorians. It is noteworthy that Arab minority in Khuzestan (Iran) displayed close relatedness with North Africans [as Gabesians from Tunisia (-0.0086) and Orans from Algeria], and Saudis (0.0231).
HLA Class I and Class II haplotype
HLA-A-B haplotypes
HLA A-B haplotypic data are extremely rare in Arabs. The most frequent A-B haplotypes in Arabs are shown in Table 6. A*02:01-B*50:01 (9.0%) and A*02:01-B*44:02/03 (7.5%) were the haplotypes with the highest frequencies in Berbers of Zrawa. Diversity in A-B haplotype frequencies are found among Arabs, hence demonstrating comparable frequencies of A-B haplotype in Arab populations, which did not exceed 5.3% in Gabesians (Tunisia). For example, while A*34:02-B*08:01 and A*29:01-B*45:01 characterize Tunisians, A*01-B*57(02.9%), A*30-B*18 (01.50%), and A*33:01-B*14:01 (02.50%) characterize Algerians. Several haplotypes identified in Arabs were also seen in other Mediterraneans. For example, A*32:01-B*40:02 was seen in Greeks (2%) [39] and Spaniards (0.5%) [41], while A*02:01-B*50:01 was seen in Italians (2%) [68], Portuguese (3%) [39], and Moroccan Jews (3%) [66]. A*24:02-B*08:01 (4.75%) and A*30:02-B*53:01 (3.48%) were only identified in Saudis.
Table 6. Most frequent (%) HLA Class I (A-B) two-locus haplotypes with significant linkage disequilibrium (P<0.05) in Arabs.
| A-B haplotype | Tun | Saudi-B | Alg | Mor-Ch | Mor-a | Ber-Z | Lib | Gab |
|---|---|---|---|---|---|---|---|---|
| 01:01–50:01 | - | - | - | 04.10 | - | - | - | - |
| 01–57 | - | - | 02.90 | - | - | - | - | - |
| 02:01–07:02 | - | - | - | - | - | - | 02.97 | - |
| 02:01–44:02/03 | 03.86 | - | - | 02.10a | 02.95c | 07.50b | - | 05.26 |
| 02:01–50:01 | 03.30 | - | - | - | 01.99d | 09.01 | - | - |
| 02:01–51:01 | - | 04.66 | - | 03.40 | 01.62f | - | - | - |
| 23:01–50:01 | - | 04.90 | - | - | - | - | 02.97 | - |
| 24:02–08:01 | - | 04.75 | - | - | - | - | - | - |
| 29:01–45:01 | 01.79 | - | - | - | - | - | - | 02.10 |
| 29:02–44:03 | - | - | - | 02.70 | - | - | - | - |
| 30–18 | - | - | 01.50 | - | 02.60 | 03.00 | - | - |
| 30:02–53:01 | - | 03.48 | - | - | - | - | - | - |
| 32:01–40:02 | 00.80 | - | - | - | - | 05.66 | - | - |
| 33:01–14:01 | - | - | 02.50 | - | 01.86e | 01.41 | - | - |
| 34:02–08:01 | 02.12 | - | - | - | - | 06.11 | - | 02.10 |
a02:01–44:02.
b02:01–44.
c02-44.
d02-50.
e33-14.
f02-51.
HLA-DRB1-DQB1 haplotypes
The most frequent DRB1-DQB1 haplotypes with significant LD in Arabs are listed in Table 7. In general, class II haplotype frequencies are markedly higher than those of class I haplotypes. DRB1*03:01-DQB1*02:01 haplotype was the most frequent DRB1-DQB1 haplotype in Arabs (Table 7), and its frequency ranging from 3.2% in Lebanese to 16.60% in Tunisians. DRB1*03:01-DQB1*02:01 is a common class II haplotype in the Mediterranean basin, and is frequent among Basques (17.5%) [41], Moroccans (17.3%) [25], Algerians (11.3%) [67], and Cretans (7.4%) [69]. In addition, DRB1*07:01-DQB1*02:02 is also frequent in Arabs, such as Moroccans (16.70%), and is reportedly common in Spaniards (17.3%) [41], and Moroccans (12.6%) [25], but rare in Southern Tunisians (2.10%) (Gabesians). In addition, DRB1*07:01-DQB1*02:01 is also a common DRB1-DQB1 haplotype, and its frequency exceeds 4% in several Arab populations.
Table 7. Most frequent (%) HLA Class II (DRB1-DQB1) two-locus haplotypes with significant linkage disequilibrium (P<0.05) in Arabs.
| HLA-DRB1-DQB1 | Tun | Sau-B | Mor-Ch | Bah |
Leb |
Alg | Lib-J | Yem-J | Ber-Z | Ber-J |
|---|---|---|---|---|---|---|---|---|---|---|
| 01:02–05:01 | 02.40 | 02.85 | - | - | - | 08.00 | 02.10 | 0.70 | 09.85 | 04.50 |
| 07:01–02:02 | 14.80 | 12.32 | 16.70 | - | - | - | 24.70a | 22.10a | 16.03 | - |
| 03:01–02:01 | 16.60 | 13.56 | 12.30 | 12.02 | 03.21 | 11.30 | 05.60a | 12.00a | 11.26 | - |
| 10:01–05:01 | 03.80 | 03.80 | - | 01.35 | 04.90 | 00.30 | 00.80 | 04.00 | 01.41 | 03.30 |
| 07:01–02:01 | - | - | - | 09.38 | 04.20 | 09.90 | - | - | - | 11.00 |
| 15:01–06:02 | 07.80 | 03.80 | 08.90 | - | - | 09.90 | - | - | 11.26 | 02.00 |
| 04:02–03:02 | 02.60 | - | 06.20 | - | - | 04.20 | 03.00 | 07.50 | 05.15 | - |
| 13:01–06:03 | 02.40 | - | - | - | - | 03.30 | 07.70 | 05.40 | 05.63 | 01.80 |
| 16:01–05:01 | - | - | - | 13.18 | 03.79 | - | - | - | - | - |
| 04:01–03:02 | - | - | - | 02.78 | 14.16 | - | - | - | - | - |
| 11:01–03:01 | 07.20b | 02.22 | - | 11.98 | 31.42 | 04.70 | 09.30 | 03.40 | 07.00b | 03.20 |
aDQB1*02
b11:01/04-03:01
In addition, DRB1*16:01-DQB1*05:01 and DRB1*04:01-DQB1*03:02, rare in neighboring populations and Mediterraneans, were identified only in Lebanese and Bahraini Arabs. The high frequency of DRB1*11:01-DQB1*03:01 haplotype (31.42%) among Lebanese is noteworthy, since it is the highest in all populations studied, but rare in Saudi (2.2%). Furthermore, DRB1*11:01/04-DQB1*03:01, identified in Arabs, is also frequent in Cretans (18.5%) [69] and Basques (3.1%)[41], while DRB1*01:02-DQB1*05:01 was seen in Spaniards (6.30%) [41]. Varied frequency of DRB1*13:01-DQB1*06:03 was also reported for Spaniards (13.23%) [86], Cretans (3.3%) [69], and Germans (10.8%) [87]. Likewise, DRB1*15:01-DQB1*06:02 was observed in Cretans (2.6%) [65], German population (25.2%) [87], and Southern Ireland (14.90%) [23].
HLA class I and class II extended haplotypes
Table 8 shows the most frequent extended haplotypes in Arab populations, and their likely origins. The systematic review did not reveal haplotypes shared by Arab populations because of partial presentation of haplotypic data, disparity in the level of typing resolution, variability of the studied loci, and lack of data. In addition, Arab populations share their frequent extended haplotypes with several European, especially Mediterranean, and Asian populations (Table 8). Furthermore, the possible origins of the most frequent extended haplotypes among Arabs are mainly European, Asian or Autochthonous.
Table 8. The most frequent (%) HLA extended haplotypes in Arabs.
| HLA Extended haplotypes | Arab Populations [references] | Possible origin |
|---|---|---|
| A*02:01-B*50:01-DRB1*07:01-DQB1*02:02a | Southern Tunisians (3.2%)[62], Berbers of Zrawa (8.12%) [24] | Euro-Asiatic |
| A*02:01–B*44– DRB1*04:02–DQB1*03:02b | Berbers of Zrawa (6.5%)[24] Tunisians (0.6%) [61] | Western European |
| A*24:02-B*08:01-C*07:02-DRB1*03:01c | Saudis (3.16%) [49] | Euro-Asiatic |
| A*23:01-B*50:01-C*06:02-DRB1*07:01 | Saudis (3.16%) [49] | Autochthonous |
| A*33-C*8-B*14-DRB1*01:02-DQA1*01:01-DQB1*05:01d | Algerians (1.5%) [88] | Mediterranean |
| A*30-C*5-B*18-DRB1*03:01-DQA1*05:01-DQB1*02:01e | Algerians (1.5%) [88] | Iberian-paleo-North African |
| A*02:01-C*06:02-B*50:01-DRB1*07:01-DQA1*02:01-DQB1*02:02f | Moroccans (2.9%) [65] | Euro-Asiatic |
| A*01:01-C*06:02-B*50:01-DRB1*03:01-DQA1*05:01-DQB1*02:01g | Moroccans (2.9%) [65] | Mediterranean |
| A*30-B*07-DRB1*03-DQA1*05:01-DQB1*02:01h | Jordanians (1.38%) [31] | Euro-Asiatic |
| A*1-B*8-DRB1*03-DQA1*05:01-DQB1*02:01i | Jordanians (1.03%) [31] | Pan-European |
| A*02:01-B*50:01-DRB1*07:01j | Libyans (4.24%) [32] Tunisians (1.8%) [60], and Ghannouch (2.5%) [33]. | North African |
| A*11:01-B*52:01-DRB1*15:02k | Libyans (2.54%) [32]; Yemen Jews (0.93%) [23] | Mediterranean |
| A*69-B*49-DRB1*04:03-DQB1*03:02 | Palestinians (2.4%) [29] | Autochthonous |
| A*24-B*18-DRB1*11:04-DQB1*03:01l | Palestinians (1.8%) [29] | Central-South-Eurasian |
a present in Spaniards (1.2%) [41], Turks (1.3%) [79], Italians (0.5%) [68], and Moroccan Jews (2%) [66].
b also found in British (2.6%), Cornish (7.9%), Danes (2%) [39], Italians (0.9%) [68], Spaniards (0.6%) [41], Spanish Basques (1.9%), Pasiegos (3.3%), Cabuemigos (2.2%) [77], and Portuguese (3.1%) [39].
c present at low frequencies in the Euro-Asian minorities of Germany [23].
d found in Armenians (0.031), Sardinians (0.027), French (0.014), Greeks (0.011), and Italians (0.007) [68].
e also found in Sardinians (11.4%), and French-Basques (4.7%) [68].
g found in Spaniards, Italians, and north Africans [65].
h present in Cornish (0.084), British (3.3%), and Danes (3.8%) [68].
i present in Basques (5%), Spaniards (3.4%) [41], Macedonians (4.9%) [78], Yugoslavians (7.7%), British (2.9%), and Germans (4.8%) [68].
j found in Poland Jews (1.15%); Ashkenazi Jews (0.92%) [23].
k present in Ashkenazi Jews (1.05%) [23].
l found in Armenians (2.1%) and Italians (0.7%) [23].
Discussion
This meta-analysis is the first genetic anthropology study in MENA region, and included 100 populations from 36 Arab and neighbouring countries, and comprising in excess of 16,000 individuals. A main outcome of the study is the lack of striking differences in the distribution of HLA alleles and haplotypes between North Africans and Arabian Peninsula populations. On the contrary, key differences were noted between Levant Arabs (Lebanese, Palestinians, Syrians), and other Arab populations, highlighted by high frequencies of A*24, B*35, DRB1*11:01, DQB1*03:01, and DRB1*11:01-DQB1*03:01 haplotype in Levantine Arabs compared to other Arab populations. Class I haplotype frequencies are lower than Class II haplotypes, because of weak LD between A and B loci, due to long physical distance between them, compared to DRB1 and DQB1 loci. The identification of shared haplotypes between Arabs and other Mediterranean and Asian populations is attributed to the higher admixture of Mediterraneans and Asians in Arab populations.
Iberians, North Africans, and Arabian Peninsula inhabitants
The relatedness between North Africans and Iberians was previously discussed [29, 59–62, 69, 78, 79, 86, 88]. Using correspondence analysis, NJ trees and genetic distances, our results show that North Africans are genetically close to Iberians, which is supported by historical events. First, this relatedness is attributed to the Berber migration from the African Sahara northwards in 10000–4000 BC, because of hyper-arid conditions [69]. It may also be explained by the similar history between Iberians and North Africans, both of whom were invaded by Phoenicians, Romans, Germans, Muslim Arabs [89]; the respective invading armies had a mixed genetic complexity; indeed, most of them were mercenaries recruited in recent conquests like in the case of Phoenicians [90] and Muslim who invaded Iberia had troops that were mostly Berbers. The invasion of Iberia by Muslims in the 8th century AD may have had a role in the relatedness between North Africans and Iberians for two reasons: first, most Muslim invaders recruits were North African Berbers, and the second is explained by the 8 centuries period of settlement of the Muslims in Iberia, although more ancient and continuous gene exchange since prehistoric times between Iberia and North Africa may have been induced the main exchange [86]; massive mixed marriages and breeding across religious Iberian groups under Muslim rule is not documented.
The analyses performed showed that current North Africans are closely related to Tunisian (Zrawa and Matmata) and Moroccan (Sousse-Agadir and Eljadida) Berbers, suggesting that North Africans have a genetic Berber profile. On the contrary, North Africans displayed a greater distance from the Arabs of Levant (Palestinians, Syrians, Lebanese, and Jordanians), indicating low genetic contribution of Phoenician and Levant Arab invasion of North Africa. These observations based on HLA markers prompted the conclusion that all Berbers of North Africa constitute a homogeneous genetic unit, except for small isolates, such as the Berbers of Djerba, who display a Berber genetic profile.
Saudi populations used in this study originated from Eastern Saudi Arabia, especially from Riyadh province. There is no reliable HLA data on Eastern Saudi Arabia that shed light on pre-Islamic history; some ancient people may have originated from old Persians, but quantification is difficult and undetermined [91]. The genetic heterogeneity between Eastern and Western Saudi Arabia is very possible, and should be taken into account in further interpretation. All analyses performed here, using HLA-A,-B, -DRB1, and DQB1 markers support the notion that Saudis along with the Kuwaitis and Yemenis are closely related to North Africans.
The most plausible explanation for West Arabia and Yemen clustering with Iberian/North Africans is a possible important massive migration that occurred when Sahara underwent desiccation in all directions [92, 93]. Cultural and language relatedness of many Mediterranean languages, including old Iberian and Basque [92], with Berber language are concordant with our genetic findings and Saharan origin hypothesis; also a part of Arabian Peninsula inhabitants (including Yemen) may had been reached by Saharan people. In fact, Malika Hachid who has been studying Saharan and North African Archaeology, culture and rock painting/writing of prehistoric Sahara, even suggests that first known writing alphabet was originated in Sahara. Proto-Berber writing rock characters have been used (very similar to present day used Berber scripts). This Proto-Berber language could have appeared 5,000 years BC [94, 95].
Explanation to HLA Kuwait genetic similarity to this group seems more difficult to achieve but interaction between Arabian Peninsula and Mesopotamia through this strategic Kuwait area is documented since 6,500 years BC (Ubard Period) [96].
Arabs of Levant
Using genetic distances, correspondence analysis and NJ trees, we showed earlier [61, 62] and in this study that Palestinians, Syrians, Lebanese and Jordanians are closely related to each other and to Eastern Mediterranean Europeans (Turks, Cretans, Greeks), Egyptians and Iranians, and confirmed by HLA class I (A, B) and class II markers (DRB1 and DQB1) analysis. However, Levant Arabs are distant from North African Arabs (Tunisians, Algerians, Moroccans and Libyans) and Iberians (Basques, Spaniards). The strong relatedness between Levant Arab populations is explained by their common ancestry, the ancient Canaanites, who came either from Africa or Arabian Peninsula via Egypt in 3300 BC [97], and settled in Levant lowlands after collapse of Ghassulian civilization in 3800–3350 BC [98]. The relatedness is also attributed to the close geographical proximity, which constituted one territory before 19th century British and French colonization.
The close relatedness of Levant Arabs to Egyptians, as confirmed genetic distances using HLA markers, may be due to three reasons. First, Egypt is a neighbor to Levant Arab countries, and historically part of the Levant. Second, the Egyptians invaded the Levant several times throughout history; the most significant was 1468 BC invasion, where they settled for 12 centuries [99]. Third, the Canaanites, the likely ancestors of Levant Arabs, may have originated from Africa through Egypt, where they settled for a long period, suggesting likely admixture between Canaanites and Egyptians.
Historically, Levant is a wider region that included countries along the Eastern Mediterranean with its islands, and extended from Greece to Cyrenaica [100]. Broadly, Levant was historically characterized by high migratory flow between its sub-regions in all directions. For example, present-day Levant comprising Palestine, Lebanon, Syria, and Jordan has undergone successive invasions by populations originating from the great Levant, including Egyptians (1468 BC), Horites, Amorites, Hitites (Turks), Greeks (1200 BC), Assyrians (1090 BC) [99], and more recently the Ottomans. This has favored admixture, reduced distances and homogenized Great Levant populations, thus explaining the close relatedness of Levant Arabs to Eastern Mediterranean populations. On the other hand, Levant Arabs are distant from Saudis, Kuwaitis, and Yeminis, an indication that the contribution of the Arabian Peninsula populations to Levantine gene pool is low, probably due to the absence of the demographic aspect of 7th century invasion.
Sudanese and Comorians
Sudanese are close to sub-Saharan Africans (Nigerians, Congolese, and Senegalese), and North Africans, in particular Egyptians, suggesting that the genetic profile of Sudanese is the admixture between North Africans (especially Egyptians) and sub-Saharan Africans throughout history. The close relatedness of Sudanese to sub-Saharan Africans suggests a reduced genetic effect of Arabs on Sudanese. Also, the Comorians (Comoros islands officially joined League of Arab Countries in 1993) are close to sub-Saharan Africans (Congolese, Nigerians, and Gabonese) [43], Egyptians, Iranians, and Eastern Mediterranean. This suggests high admixture between populations belonging to three continents in the Comoro Islands, and can be explained by their geographical position as a corridor for international trade.
Bahrainis, Emiratis, and Omanis
Bahrainis, Emiratis, and Omanis are geographically similar populations, which explains their genetic relationship as demonstrated in this study. These three populations tend to form a heterogeneous group with Pakistanis, Indians, Iranian Arabs (Famoori), Sardinians (the later probably close to Iberians/North Africans but behaving as out layer group in analyses because of they are a genetic island isolate), Egyptians, and some sub-Saharan Africans, such as Congolese. These populations appear close to certain Eastern Mediterranean populations including Greeks, Macedonians, and those further, in particular North Africans, hence explaining their intermediate grouping, and distinction from two main clusters. Collectively, this suggests high admixture in these populations brought about by their commercially important position. Sardinia is a relative genetic isolate “founded” by Iberian Norax/Nora (first documented Sardinian capital close to Cagliari) and Iberians/North Africans may be genetically related to Sardinians (A*30-B*18-Cw*5 basic HLA haplotype is very high in Sardinia, Iberia, and North Africa) [93].
Minorities of Arab World
Ethnic minorities
The Kurds and Berbers are the two major ethnic minorities in Arab world. Berbers are indigenous North African ethnic group found over a vast area stretching from Atlantic Ocean to Siwa Oasis in Egypt, and from Mediterranean Sea to Niger River. Berbers number about 20 million people, and constitute 40–45% of Moroccans, 20–25% of Algerians, and 2–7% in both Libya and Tunisia. The Kurds live in the northern regions of Iraq (15–20%) and Syria (10%). They constitute an Indo-European ethnic group, and speak Kurdish. Less important minorities include Armenians, Nubians, Assyrians, and Turkmen [99].
Berbers populations used in this work are closely linked to each other, as well as to present-day North Africans, and to Western Mediterranean populations, especially Iberians. Indeed, the Moroccan Berbers are not genetically different from the current Moroccans, nor those of neighboring populations, like Algerians and Tunisians. This also applies to Tunisian Berbers, except those of the island of Djerba, who appear to be related to Eastern Mediterranean populations, including Levant Arabs. This suggests that North African Berbers are in perfect harmony with their environments, and that differences between them are cultural rather than genetic due to 7th century Arabization of the region.
Clustering and genetic distances analyses demonstrated that Iraqi and Iranian Kurds are not genetically different from Iranians or neighboring populations, including Levant Arab, and are close to Turks and other Eastern Mediterranean populations. This suggests that Kurds originate from the region, and are in genetic harmony with neighboring populations, despite the clear cultural differences. This suggests that Kurds, Syrians, Jordanians, Palestinians, Iraqis, Lebanese, and Iranians probably share the same genetic profile, with few differences. Accordingly, our findings confirm the results of an earlier study of Arnaiz-Villena on Iraqi Kurds [54].
Religious minorities
Sunni Muslims constitute the majority (80%) of Arab populations, followed by Shi'a Muslims (10%) who are present in parts of Iraq, Lebanon, Saudi Arabia, Kuwait, Yemen, and Bahrain. Non-Muslims make up about 10% of all Arabs, and Christianity (6%) is the second largest religion among Arabs, with about 20 million Christians living in Lebanon, Egypt, Iraq, Syria, and Jordan. Other minor religions (4%) such as Judaism, Druze and others are practiced on a much smaller scale [99].
HLA data on Sunni and Shiite Arabs are not available, same as comparison of Muslims to Christians. The only available data are those concerning Arab Jews. In this study, data are available for three Jewish populations, including two from North Africa (Moroccan and Libyan Jews) and one from the Arabian Peninsula (Yemenite Jews). While genetic distances separating these three groups of Jews are small (S1 Table), genetic heterogeneity between these Jewish populations was noted. For example, Yemenite Jews are related to Western Mediterranean populations, including North Africans and Iberians, while Libyan Jews are related to Eastern Mediterraneans, including Levantine Arabs. The relatedness of Moroccan Jews depends to other communities on the studied HLA loci; they associate with Eastern Mediterraneans using DRB1, but group with Eastern Mediterraneans when the other markers are used.
Conclusion
This study supports the notion that Arabs are divided into four groups. The first consisting of North Africans (Algerians, Tunisians, Moroccans, and Libyans), Saudis, Kuwaitis, and Yemenis, with relatedness to Western Mediterraneans, including Iberians. The second includes Levantine Arabs (Palestinians, Jordanians, Lebanese, and Syrians), Iraqi, and Egyptians, who appear to be related to the Eastern Mediterranean and Iranians, who in turn belonged to 'Great Levant' historically described. The third consists of Sudanese and Comorians who associate with Sub-Saharan Africans. Finally, the fourth group of Arabs comprises Omanis, Emiratis, and Bahrainis. This group associates with heterogeneous populations (Mediterranean, Asian and sub-Saharan). Lastly, the two main indigenous minorities, Berbers and Kurds, are not genetically different from the ‘host’ and neighboring populations.
Supporting information
(DOC)
Populations’ data were taken from references detailed in Tables 1 and 2. Bootstrap values from 1.000 replicates are shown.
(TIF)
Populations’ data were taken from references detailed in Tables 1 and 2. Bootstrap values from 1.000 replicates are shown.
(TIF)
(DOC)
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
The authors received no specific funding for this work.
References
- 1.HLA allele database: http://hla.alleles.org (last accessed on September 17, 2017)
- 2.Hudson RR. Analysis of population subdivision in Handbook of statistical genetics, MBD. Balding MBD and Cannings C. (Eds). pp. 309–324. John Wiley & Sons; Chichester, UK, 2001 [Google Scholar]
- 3.Takezaki N, Nei M. Empirical tests of the reliability of phylogenetic trees constructed with microsatellite DNA. Genetics. 2008; 178(1): 385–92. doi: 10.1534/genetics.107.081505 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Nei M. Phylogenetic analysis in molecular evolutionary genetics. Annual Review of Genetics. 1996; 30: 371–403. doi: 10.1146/annurev.genet.30.1.371 [DOI] [PubMed] [Google Scholar]
- 5.Tamura K, Nei M, Kumar S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proceedings of the National Academy of Sciences USA. 2004; 101(30): 11030–5. doi: 10.1073/pnas.0404206101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Excoffier L, Slatkin M. Maximum-likelihood estimation of molecular haplotype frequencies in a diploid population. Molecular Biology and Evolution. 1995; 12(5): 921–7. doi: 10.1093/oxfordjournals.molbev.a040269 [DOI] [PubMed] [Google Scholar]
- 7.The World Factbook: https://www.cia.gov/library/publications/the-world-factbook
- 8.Bengio O, Ben-Dor G. Minorities and the State in the Arab World. Lynne Rienner Publishers, 1999. –224 pages [Google Scholar]
- 9.Encyclopædia Britannica, Himyar: https://www.britannica.com/topic/Himyar
- 10.Korotayev A. Ancient Yemen. Oxford: Oxford University Press, 1995. [Google Scholar]
- 11.Korotayev A. Pre-Islamic Yemen. Wiesbaden: Harrassowitz Verlag, 1996. [Google Scholar]
- 12.Munro-Hay, Stuart C. Aksum: An African Civilization of Late Antiquity 1991. Edinburgh: Edinburgh University Press, 1991. [Google Scholar]
- 13.Robin CJ. Arabia and Ethiopia, 'in Johnson Scott (ed.) The Oxford Handbook of Late Antiquity, Oxford University Press; 2012. pp. 247–333, p.279. [Google Scholar]
- 14.Hoyland R. Arabia and the Arabs: From the Bronze Age to the Coming of Islam, Routledge, 2001, p.51. [Google Scholar]
- 15.Encyclopædia wikipedia: https://en.wikipedia.org/wiki/History_of_Islam
- 16.Hourani A. A History of the Arab Peoples. Harvard University Press; 2002; pp. 15–19. ISBN 9780674010178. [Google Scholar]
- 17.Moher D, Liberati A, Tetzlaff J, Altman DG, and PRISMA Group, “Reprint—preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement”. Physical Therapy. 2009; 89(9): 873–80. https://doi.org/10.1093/ptj/89.9.873 [PubMed] [Google Scholar]
- 18.Young FW, Bann CM. A visual statistics system In Stine RA, Fox J, eds. Statistical computing environments for social researches. New York: Sage publications; 1996; 207–36. [Google Scholar]
- 19.Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular Biology and Evolution. 1987; 4(4): 406–425. doi: 10.1093/oxfordjournals.molbev.a040454 [DOI] [PubMed] [Google Scholar]
- 20.Nei M. Genetic distances between populations. The American Naturalist. 1972; 106:283 http://jstor.org/stable/2459777 [Google Scholar]
- 21.Nei M. Analysis of gene diversity in subdivided populations. Proceedings of the National Academy of Sciences USA. 1973; 70(12): 3321–3. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Nei M, Tajima F, Tateno Y. Accuracy of estimated phylogenetic trees from molecular data. II. Gene frequency data.Journal of Molecular Evolution. 1983; 19(2): 153–70. https://doi.org/10.1007/BF02300753 [DOI] [PubMed] [Google Scholar]
- 23.Database of allele frequencies: http://www.allelefrequencies.net, 2017
- 24.Hajjej A, Sellami MH, Kaabi H, Hajjej G, El-Gaaied A, Boukef K, et al. HLA class I and class II polymorphisms in Tunisian Berbers. Annals of Human Biology. 2011; 38 (2): 156–64. doi: 10.3109/03014460.2010.504195 [DOI] [PubMed] [Google Scholar]
- 25.Gomez-Casado E, del Moral P, Martinez-Laso J, García-Gómez A, Allende L, Silvera-Redondo C, et al. HLA gene in Arabic-Speaking Moroccans: close relatedness to Berbers and Iberians. Tissue Antigens. 2000; 55(3): 239–49. https://doi.org/10.1034/j.1399-0039.2000.550307.x [DOI] [PubMed] [Google Scholar]
- 26.Mahfoudh N, Ayadi I, Kamoun A, Ammar R, Mallek B, Maalej L, et al. Analysis of HLA-A, -B, -C, -DR, -DQ polymorphisms in the South Tunisian population and a comparison with other populations. Annals of Human Biology. 2013; 40(1): 41–7. doi: 10.3109/03014460.2012.734334 [DOI] [PubMed] [Google Scholar]
- 27.Matevosyan L, Chattopadhyay S, Madelian V, Avagyan S, Nazaretyan M, Hyussian A, et al. HLA-A, HLA-B, and HLA-DRB1 allele distribution in a large Armenian population sample. Tissue Antigens. 2011; 78(1): 21–30. doi: 10.1111/j.1399-0039.2011.01668.x [DOI] [PubMed] [Google Scholar]
- 28.Hamdi NM, Al-Hababi FH, Eid AE. HLA class I and class II associations with ESRD in Saudi Arabian population. PLoS One. 2014. November 7; 9(11): e111403 doi: 10.1371/journal.pone.0111403 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Arnaiz-Villena A, Elaiwa N, Silvera C, Rostom A, Moscoso J, Gómez-Casado E, et al. The origin of Palestinians and their genetic relatedness with other Mediterranean populations. Retraction in: Suciu-Foca N, Lewis R. Human Immunology. 2001; 62(9): 889–900. (Accessed on https://commons.wikimedia.org/wiki/File:Palestinians_hla.pdf) [DOI] [PubMed] [Google Scholar]
- 30.Albalushi KR, Sellami MH, Alriyami H, varghese M, Boukef MK, Hmida S. The Investigation of the Evolutionary History of the Omani Population by Analysis of HLA Class I Polymorphism. Anthropologist. 2014; 18(1): 205–210 [Google Scholar]
- 31.Sánchez-Velasco P, Karadsheh NS, García-Martín A, Ruíz de Alegría C, Leyva-Cobián F. Molecular analysis of HLA allelic frequencies and haplotypes in Jordanians and comparison with other related populations. Human Immunology. 2001; 62(9): 901–9. https://doi.org/10.1016/S0198-8859(01)00289-0. . [DOI] [PubMed] [Google Scholar]
- 32.Galgani A, Mancino G, Martínez-Labarga C, Cicconi R, Mattei M, Amicosante M, et al. HLA-A, -B and -DRB1 allele frequencies in Cyrenaica population (Libya) and genetic relationships with other populations. Hum Immunol. 2013; 74(1): 52–9. doi: 10.1016/j.humimm.2012.10.001 [DOI] [PubMed] [Google Scholar]
- 33.Hajjej A, Hmida S, Kaabi H, Dridi A, Jridi A, El Gaaled A, et al. HLA genes in Southern Tunisians (Ghannouch area) and their relationship with other Mediterraneans. European Journal Medical Genetics. 2006; 49(1): 43–56. doi: 10.1016/j.ejmg.2005.01.001 [DOI] [PubMed] [Google Scholar]
- 34.Hmida S, Gauthier A, Dridi A, Quillivic F, Genetet B, Boukef K, et al. HLA class II gene polymorphism in Tunisians. Tissue Antigens. 1995; 45(1): 63–8. https://doi.org/10.1111/j.1399-0039.1995.tb02416.x [DOI] [PubMed] [Google Scholar]
- 35.Almawi WY, Busson M, Tamim H, Al-Harbi EM, Finan RR, Wakim-Ghorayeb SF, et al. HLA class II profile and distribution of HLA-DRB1 and HLA-DQB1 alleles and haplotypes among Lebanese and Bahraini Arabs. Clinical and Diagnostic Laboratory Immunology. 2004; 11(4): 770–4. doi: 10.1128/CDLI.11.4.770-774.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Amar A, Kwon OJ, Motro U, Witt CS, Bonne-Tamir B, Gabison R, et al. Molecular analysis of HLA class II polymorphisms among different ethnic groups in Israel. Human Immunology. 1999; 60(8): 723–30. https://doi.org/10.1016/S0198-8859(99)00043-9 [DOI] [PubMed] [Google Scholar]
- 37.Izaabel H, Garchon HJ, Caillat-Zucman S, Beaurain G, Akhayat O, Bach JF, et al. HLA class II DNA polymorphism in a Moroccan population from the Souss, Agadir area. Tissue Antigens. 1998; 51(1): 106–10. https://doi.org/10.1111/j.1399-0039.1998.tb02954.x [DOI] [PubMed] [Google Scholar]
- 38.Al-Tonbary Y, Abdel-Razek N, Zaghloul H, Metwaly S, El-Deek B, El-Shawaf R. HLA class II polymorphism in Egyptian children with lymphomas. Hematology. 2004; 9(2): 139–45. doi: 10.1080/1024533042000205487 [DOI] [PubMed] [Google Scholar]
- 39.Clayton J, Lonjou C. Allele and Haplotype frequencies for HLA loci in various ethnic groups In Charron D, ed. Genetic diversity of HLA. Functional and medical implications. Vol 1 Paris: EDK; 1997; 665–820. [Google Scholar]
- 40.Abdennaji Guenounou B, Loueslati BY, Buhler S, Hmida S, Ennafaa H, Khodjet-Elkhil H, et al. HLA class II genetic diversity in Southern Tunisia and the Mediterranean area. International Journal Immunogenetics. 2006; 33(2): 93–103. doi: 10.1111/j.1744-313X.2006.00577.x [DOI] [PubMed] [Google Scholar]
- 41.Martinez-Laso J, De Juan D, Martinez-Quiles N, Gomez-Casado E, Cuadrado E, Arnaiz-Villena A. The contribution of the HLA-A, -B, -C and -DR, -DQ DNA typing to the study of the origins of Spaniards and Basques. Tissue Antigens. 1995; 45(4): 237–45. https://doi.org/10.1111/j.1399-0039.1995.tb02446.x . [DOI] [PubMed] [Google Scholar]
- 42.Brick C, Bennani N, Atouf O, Essakalli M. HLA-A, -B, -DR and -DQ allele and haplotype frequencies in the Moroccan population: a general population study. Transfusion Clinique et Biologique. 2006; 13(6): 346–52. doi: 10.1016/j.tracli.2006.12.003 [DOI] [PubMed] [Google Scholar]
- 43.Gibert M, Touinssi M, Reviron D, Mercier P, Boëtsch G, Chiaroni J. HLA-DRB1 frequencies of the Comorian population and their genetic affinities with Sub-Saharan African and Indian Oceanian populations. Annals of Human Biology. 2006; 33(3): 265–78. doi: 10.1080/03014460600578599 [DOI] [PubMed] [Google Scholar]
- 44.Samaha H, Rahal EA, Abou-Jaoude M, Younes M, Dacchache J, Hakime N. HLA class II allele frequencies in the Lebanese population. Molecular Immunology. 2003; 39(17–18): 1079–81. https://doi.org/10.1016/S0161-5890(03)00073-7 [DOI] [PubMed] [Google Scholar]
- 45.Khansa S, Hoteit R, Shammaa D, Khalek RA, El Halas H, Greige L, et al. HLA class II allele frequencies in the Lebanese population. Gene. 2012; 506(2): 396–9. doi: 10.1016/j.gene.2012.06.063 [DOI] [PubMed] [Google Scholar]
- 46.Elbjeirami WM, Abdel-Rahman F, Hussein AA. Probability of finding an HLA-matched donor in immediate and extended families: the Jordanian experience. Biology of Blood and Marrow Transplantation. 2013; 19(2): 221–6. doi: 10.1016/j.bbmt.2012.09.009 [DOI] [PubMed] [Google Scholar]
- 47.Mourad J, Monem F. HLA-DRB1 allele association with rheumatoid arthritis susceptibility and severity in Syria. Revista Brasileira De Reumatologia. 2013; 53(1): 47–56. [PubMed] [Google Scholar]
- 48.Djidjik R, Allam I, Douaoui S, Meddour Y, Cherguelaîne K, Tahiat A, et al. Association study of human leukocyte antigen-DRB1 alleles with rheumatoid arthritis in Algerian patients. International Journal of Rheumatic Diseases. 2014. doi: 10.1111/1756-185X.12272 [DOI] [PubMed] [Google Scholar]
- 49.Hajeer AH, Al Balwi MA, AytülUyar F, Alhaidan Y, Alabdulrahman A, Al Abdulkareem I, et al. HLA-A, -B, -C, -DRB1 and -DQB1 allele and haplotype frequencies in Saudis using next generation sequencing technique. Tissue Antigens. 2013; 82(4): 252–8. doi: 10.1111/tan.12200 [DOI] [PubMed] [Google Scholar]
- 50.Hajeer AH, Sawidan FA, Bohlega S, Saleh S, Sutton P, Shubaili A, Tahan AA, Al Jumah M. HLA class I and class II polymorphisms in Saudi patients with myasthenia gravis. International Journal of Immunogenetics. 2009; 36(3): 169–72. doi: 10.1111/j.1744-313X.2009.00843.x [DOI] [PubMed] [Google Scholar]
- 51.Albalushi KR, Sellami MH, Alriyami H, varghese M, Boukef MK, Hmida S. HLA Class II (DRB1 and DQB1) Polymorphism in Omanis. Journal of Transplantation Technologies and Research 2014; 4: 134 https://doi.org/10.4172/2161-0991.1000134 [Google Scholar]
- 52.Haider MZ, Shaltout A, Alsaeid K, Qabazard M, Dorman J. Prevalence of human leukocyte antigen DQA1 and DQB1 alleles in Kuwaiti Arab children with type 1 diabetes mellitus. Clinical Genetics. 1999; 56(6): 450–6. https://doi.org/10.1034/j.1399-0004.1999.560608.x [DOI] [PubMed] [Google Scholar]
- 53.Haider MZ, Zahid MA, Dalal HN, Razik MA. Human leukocyte antigen (HLA) DRB1 alleles in Kuwaiti Arabs with schizophrenia.American Journal of Medical Genetics. 2000; 96(6): 870–2. https://doi.org/10.1002/1096-8628(20001204)96:6<870::AID-AJMG36>3.0.CO;2-L . [DOI] [PubMed] [Google Scholar]
- 54.Arnaiz-Villena A, Palacio-Grüber J, Muñiz E, Campos C, Alonso-Rubio J, Gomez-Casado E, et al. Genetic HLA Study of Kurds in Iraq, Iran and Tbilisi (Caucasus, Georgia): Relatedness and Medical Implications. PLoS One. 2017. January 23; 12(1): e0169929 doi: 10.1371/journal.pone.0169929 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Nassar MY, Al-Shamahy HA, Masood HA. The Association between Human Leukocyte Antigens and Hypertensive End-Stage Renal Failure among Yemeni Patients. Sultan Qaboos University Medical Journal. 2015; 15(2): e241–249. [PMC free article] [PubMed] [Google Scholar]
- 56.Middleton D, Williams F, Meenagh A, Daar AS, Gorodezky C, Hammond M, et al. Analysis of the distribution of HLA-A alleles in populations from five continents. Human Immunology. 2000; 61(10): 1048–52. https://doi.org/10.1016/S0198-8859(00)00178-6 [DOI] [PubMed] [Google Scholar]
- 57.Williams F, Meenagh A, Darke C, Acosta A, Daar AS, Gorodezky C, et al. Analysis of the distribution of HLA-B alleles in populations from five continents. Human Immunology. 2001; 62(6): 645–50. https://doi.org/10.1016/S0198-8859(01)00247-6 [DOI] [PubMed] [Google Scholar]
- 58.Jazairi B, Khansaa I, Ikhtiar A, Murad H. Frequency of HLA-DRB1 and HLA-DQB1 Alleles and Haplotype Association in Syrian Population. Immunological Investigation. 2016; 45(2): 172–9. doi: 10.3109/08820139.2015.1131293 [DOI] [PubMed] [Google Scholar]
- 59.Hajjej A, Hajjej G, Almawi WY, Kaabi H, El-Gaaied A, Hmida S. HLA class I and class II polymorphism in a population from south-eastern Tunisia (Gabes Area). International Journal of Immunogenetics. 2011; 38(3): 191–9. doi: 10.1111/j.1744-313X.2011.01003.x [DOI] [PubMed] [Google Scholar]
- 60.Hajjej A, Kâabi H, Sellami MH, Dridi A, Jeridi A, El borgi W, et al. The contribution of HLA class I and II alleles and haplotypes to the investigation of the evolutionary history of Tunisians. Tissue Antigens. 2006; 68(2): 153–62. doi: 10.1111/j.1399-0039.2006.00622.x [DOI] [PubMed] [Google Scholar]
- 61.Hajjej A, Almawi WY, Hattab L, El-Gaaied A, Hmida S. HLA Class I and Class II Alleles and Haplotypes Confirm the Berber Origin of the Present Day Tunisian Population. PLoS One. 2015; 10(8): e0136909 doi: 10.1371/journal.pone.0136909 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Hajjej A, Almawi WY, Hattab L, El-Gaaied A, Hmida S. The investigation of the origin of Southern Tunisians using HLA genes. Journal of Human Genetics. 2017; 62(3): 419–429. doi: 10.1038/jhg.2016.146 [DOI] [PubMed] [Google Scholar]
- 63.Ayed K, Ayed-Jendoubi S, Sfar I, Labonne MP, Gebuhrer L. HLA class-I and HLA class-II phenotypic, gene and haplotypic frequencies in Tunisians by using molecular typing data. Tissue Antigens. 2004; 64(4): 520–32. doi: 10.1111/j.1399-0039.2004.00313.x [DOI] [PubMed] [Google Scholar]
- 64.Oumhani K, Canossi A, Piancatelli D, Di Rocco M, Del Beato T, Liberatore G, et al. Sequence-Based analysis of the HLA-DRB1 polymorphism in Metalsa Berber and Chaouya Arabic-speaking groups from Morocco. Human Immunology. 2002; 63(2): 129–38. https://doi.org/10.1016/S0198-8859(01)00370-6 [DOI] [PubMed] [Google Scholar]
- 65.Canossi A, Piancatelli D, Aureli A, Oumhani K, Ozzella G, Del Beato T, et al. Correlation between genetic HLA class I and II polymorphisms and anthropological aspects in the Chaouya population from Morocco (Arabic speaking). Tissue Antigens. 2010; 76(3): 177–193. doi: 10.1111/j.1399-0039.2010.01498.x [DOI] [PubMed] [Google Scholar]
- 66.Roitberg-Tambur A, Witt CS, Friedmann A, Safirman C, Sherman L, Battat S, Nelken D, Brautbar C. Comparative analysis of HLA polymorphism at the serologic and molecular level in Moroccan and Ashkenazi Jews. Tissue Antigens. 1995; 46(2): 104–10. https://doi.org/10.1111/j.1399-0039.1995.tb02485.x [DOI] [PubMed] [Google Scholar]
- 67.Arnaiz-Villena A, Benmamar D, Alvarez M, Diaz-Campos N, Varela P, Gomez-Casado E, et al. HLA allele and haplotype frequencies in Algerians. Relatedness to Spaniards and Basques. Human Immunology. 1995; 43(4): 259–68. https://doi.org/10.1016/0198-8859(95)00024-X [DOI] [PubMed] [Google Scholar]
- 68.Imanishi T, Akaza T, Kimura A, Tokunaga K, Gjobori T. Allele and haplotype frequencies for HLA and complement loci in various ethnic groups In, eds. HLA 1991. VOL 1 Oxford: Oxford University Press; 1992; 1065–220. [Google Scholar]
- 69.Arnaiz-Villena A, Iliakis P, González-Hevilla M, Longás J, Gómez-Casado E, Sfyridaki K, et al. The origin of Cretan populations as determined by characterization of HLA alleles. Tissue Antigens. 1999; 53(3): 213–26. https://doi.org/10.1034/j.1399-0039.1999.530301.x [DOI] [PubMed] [Google Scholar]
- 70.Comas D, Mateu E, Calafell F, Pérez-Lezaun A, Bosch E, Martínez-Arias R, et al. HLA class I and class II DNA typing and the origin of Basques. Tissue Antigens. 1998; 51(1): 30–40. https://doi.org/10.1111/j.1399-0039.1998.tb02944.x [DOI] [PubMed] [Google Scholar]
- 71.Grimaldi MC, Crouau-Roy B, Amoros JP, Cambon-Thomsen A, Carcassi C, Orru S, et al. West Mediterranean islands (Corsica, Balearic Islands, Sardinia) and the Basque population: contribution of HLA class I molecular markers to their evolutionary history. Tissue Antigens. 2001; 58(5): 281–92. https://doi.org/10.1034/j.1399-0039.2001.580501.x [DOI] [PubMed] [Google Scholar]
- 72.Renquin J, Sanchez-Mazas A, Halle L, Rivalland S, Jaeger G, Mbayo K, et al. HLA class II polymorphism in Aka Pygmies and Bantu Congolese and a reassessment of HLA-DRB1 African diversity. Tissue Antigens. 2001; 58(4): 211–22. https://doi.org/10.1034/j.1399-0039.2001.580401.x [DOI] [PubMed] [Google Scholar]
- 73.Farjadian S, Ghaderi A. HLA class II genetic diversity in Arabs and Jews of Iran. Iranian Journal of Immunology. 2007; 4(2): 85–93. doi: IJIv4i2A3 [DOI] [PubMed] [Google Scholar]
- 74.Kollaee A, Ghaffarpor M, Ghlichnia HA, Ghaffari SH, Zamani M. The influence of the HLA-DRB1 and HLA-DQB1 allele heterogeneity on disease risk and severity in Iranian patients with multiple sclerosis. International Journal of Immunogenetics. 2012; 39(5): 414–22. doi: 10.1111/j.1744-313X.2012.01104.x [DOI] [PubMed] [Google Scholar]
- 75.Sayad A, Akbari MT, Pajouhi M, Mostafavi F, Zamani M. The influence of the HLA-DRB, HLA-DQB and polymorphic positions of the HLA-DRβ1 and HLA-DQβ1 molecules on risk of Iranian type 1 diabetes mellitus patients. International Journal of Immunogenetics. 2012; 39(5): 429–36. doi: 10.1111/j.1744-313X.2012.01116.x [DOI] [PubMed] [Google Scholar]
- 76.Sulcebe G, Sanchez-Mazas A, Tiercy JM, Shyti E, Mone I, Ylli Z, et al. HLA allele and haplotype frequencies in the Albanian population and their relationship with the other European populations. International Journal of Immunogenetics. 2009; 36(6): 337–43. doi: 10.1111/j.1744-313X.2009.00868.x [DOI] [PubMed] [Google Scholar]
- 77.Sanchez-Velasco P, Gomez-Casado E, Martinez-Laso J, Moscoso J, Zamora J, Lowy E, et al. HLA alleles in isolated populations from North Spain: origin of the Basques and the ancient Iberians. Tissue Antigens. 2003; 61(5): 384–92. https://doi.org/10.1034/j.1399-0039.2003.00041.x [DOI] [PubMed] [Google Scholar]
- 78.Arnaiz-Villena A, Dimitroski K, Pacho A, Moscoso J, Gómez-Casado E, Silvera-Redondo C, et al. HLA genes in Macedonians and the sub-Saharan origin of the Greeks. Tissue Antigens. 2001; 57(2): 118–27. https://doi.org/10.1034/j.1399-0039.2001.057002118.x [DOI] [PubMed] [Google Scholar]
- 79.Arnaiz-Villena A, Karin M, Bendikuze N, Gomez-Casado E, Moscoso J, Silvera C, et al. HLA alleles and haplotypes in the Turkish population: relatedness to Kurds, Armenians and other Mediterraneans. Tissue Antigens. 2001; 57(4): 308–17. https://doi.org/10.1034/j.1399-0039.2001.057004308.x [DOI] [PubMed] [Google Scholar]
- 80.Muro M, Marín L, Torío A, Moya-Quiles MR, Minguela A, Rosique-Roman J, et al. HLA polymorphism in the Murcia population (Spain): in the cradle of the archaeologic Iberians. Human Immunology. 2001; 62(9): 910–21. https://doi.org/10.1016/S0198-8859(01)00290-7 [DOI] [PubMed] [Google Scholar]
- 81.Farjadian S, Ghaderi A. HLA class II similarities in Iranian Kurds and Azeris. International Journal of Immunogenetics. 2007; 34(6): 457–63. doi: 10.1111/j.1744-313X.2007.00723.x [DOI] [PubMed] [Google Scholar]
- 82.Mohyuddin A, Ayub Q, Khaliq S, Mansoor A, Mazhar K, Rehman S, et al. HLA polymorphism in six ethnic groups from Pakistan. Tissue Antigens. 2002; 59(6): 492–501. https://doi.org/10.1034/j.1399-0039.2002.590606.x [DOI] [PubMed] [Google Scholar]
- 83.Agrawal S, Srivastava SK, Borkar M, Chaudhuri TK. Genetic affinities of north and northeastern populations of India: inference from HLA-based study. Tissue Antigens. 2008; 72(2): 120–30. doi: 10.1111/j.1399-0039.2008.01083.x [DOI] [PubMed] [Google Scholar]
- 84.Rani R, Sood A, Goswami R. Molecular basis of predisposition to develop type 1 diabetes mellitus in North Indians. Tissue Antigens. 2004; 64(2): 145–55. doi: 10.1111/j.1399-0039.2004.00246.x [DOI] [PubMed] [Google Scholar]
- 85.Migot-Nabias F, Fajardy I, Danze PM, Everaere S, Mayombo J, Minh TN, et al. HLA class II polymorphism in a Gabonese Banzabi population. Tissue Antigens. 1999; 53(6): 580–5. https://doi.org/10.1034/j.1399-0039.1999.530610.x [DOI] [PubMed] [Google Scholar]
- 86.Arnaiz-Villena A, Muñiz E, Campos C, Gomez-Casado E, Tomasi S, Martínez-Quiles N, et al. Origin of Ancient Canary Islanders (Guanches): presence of Atlantic/Iberian HLA and Y chromosome genes and Ancient Iberian language. International Journal of Modern Anthropology. 2015; 8: 67–93. https://doi.org/10.4314/ijma.v1i8.4 [Google Scholar]
- 87.Reil A, Bein G, Machulla HK, Sternberg B, Seyfarth M. High-resolution DNA typing in immunoglobulin A deficiency confirms a positive association with DRB1*0301, DQB1*02 haplotypes. Tissue Antigens. 1997; 50(5): 501–6. https://doi.org/10.1111/j.1399-0039.1997.tb02906.x [DOI] [PubMed] [Google Scholar]
- 88.Arnaiz-Villena A, Martínez-Laso J, Gómez-Casado E, Díaz-Campos N, Santos P, Martinho A, et al. Relatedness among Basques, Portuguese, Spaniards, and Algerians studied by HLA allelic frequencies and haplotypes. Immunogenetics. 1997; 47(1): 37–43. [DOI] [PubMed] [Google Scholar]
- 89.Stearns PN. The Encyclopedia of World History: Ancient, Medieval, and Modern, Chronologically Arranged, 6 ed., Houghton Mifflin Harcourt, 2001, 2017, pp. 129–131. [Google Scholar]
- 90.Mira-Guardiola MA (2000). Cartago contra Roma. Ed.: Alderaban; Madrid, Spain. [Google Scholar]
- 91.Sellier J, Sellier A. Atlas des Peuples d’Orient. Paris, France: Editions La Decouverte, 1993 [Google Scholar]
- 92.Arnaiz-Villena A, Martinez-Laso J, Alonso-Garciá J. The Correlation Between Languages and Genes: The Usko-Mediterranean Peoples. Human Immunology. 2001; 62(9): 1051–1061. https://doi.org/10.1016/S0198-8859(01)00300-7 [DOI] [PubMed] [Google Scholar]
- 93.Arnaiz-Villena A, Gomez-Casado E, Martinez-Laso J. Population genetic relationships between Mediterranean populations determined by HLA allele distribution and a historic Perspective. Tissue Antigens. 2002; 60(2): 111–21. https://doi.org/10.1034/j.1399-0039.2002.600201.x [DOI] [PubMed] [Google Scholar]
- 94.Hachid M. Postface de l’ouvrage “aux origines de l’ecriture au Maroc. corpus des inscriptions amazighes des sites d’art rupestre du maroc” edited by: Skounti A., Lemdjidi A. and Nami M. Publication de l’institut royal de la culture amazighe; Cealpa, rabat, morocco, 2003. [Google Scholar]
- 95.Malika H. Les premier berebers entre mediterranee, tassili et nil. Edited by edisud; aix-en-provence, France: 2000 [Google Scholar]
- 96.Carter RA. Boat remains and trade in Persian Gulf during the 6th and 5th millenia BC. Antiquity. 2006; 80(307): 52–63. https://doi.org/10.1017/S0003598X0009325X [Google Scholar]
- 97.Kuhrt A. The ancient Near East (3000–330 BC). Vol II Barcelona, Editorial Critica, 2001. [Google Scholar]
- 98.Hitti PK. History of Syria: Including Lebanon and Palestine, 2004, p26 [Google Scholar]
- 99.Encyclopaedia Britannica: https://www.britannica.com/
- 100.Sartre M, D'Alexandre à Zénobie: Histoire du Levant antique, IVe siècle avant Jésus-Christ-IIIe siècle après Jésus-Christ, Fayard, 2001. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOC)
Populations’ data were taken from references detailed in Tables 1 and 2. Bootstrap values from 1.000 replicates are shown.
(TIF)
Populations’ data were taken from references detailed in Tables 1 and 2. Bootstrap values from 1.000 replicates are shown.
(TIF)
(DOC)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.