

Abstract
The incidence of malignant melanoma is rapidly increasing and current medicine is offering only limited options for treatment of the advanced disease. For B-Raf mutated melanomas, treatment with mutation-specific drug inhibitors may be used. Unfortunately, tumors frequently acquire resistance to the treatment. Tumor microenvironment, namely cancer-associated fibroblasts, largely influence this acquired resistance. In the present study, fibroblasts were isolated from a patient suffering from acrolentiginous melanoma (Breslow, 4.0 mm; Clark, IV; B-Raf V600E mutated). The present study focused on the expression of structural and functional markers of fibroblast activation in melanoma-associated fibroblasts (MAFs; isolated prior to therapy initiation) as well as in autologous control fibroblasts (ACFs) of the same patient isolated during B-Raf inhibitor therapy, yet before clinical progression of the disease. Analysis of gene transcription was also performed, as well as DNA methylation status analysis at the genomic scale of both isolates. MAFs were positive for smooth muscle actin (SMA), which is a marker of myofibroblasts and the hallmark of cancer stoma. Surprisingly, ACF isolated from the distant uninvolved skin of the same patient also exhibited strong SMA expression. A similar phenotype was also observed in control dermal fibroblasts (CDFs; from different donors) exclusively following stimulation by transforming growth factor (TGF)-β1. Immunohistochemistry confirmed that melanoma cells potently produce TGF-β1. Significant differences were also identified in gene transcription and in DNA methylation status at the genomic scale. Upregulation of SMA was observed in ACF cells at the protein and transcriptional levels. The present results support recent experimental findings that tumor microenvironment is driving resistance to B-Raf inhibition in patients with melanoma. Such an activated microenvironment may be viable for the growth of circulating melanoma cells.
Keywords: melanoma, cancer-associated fibroblast, genome, transcriptome, gene methylation, stroma, smooth muscle actin, α-actin-2, B-Raf inhibitor, transforming growth factor-β1, stroma-driven resistance
Introduction
The incidence of malignant melanoma is increasing rapidly worldwide (1). Despite notable progress in melanoma research, treatment options for advanced stages of the disease remain limited and the mortality rate in advanced melanoma is significantly higher than in other skin tumors (1).
The tumor stroma represents an important component in the structure of malignant tumors. This microenvironment may significantly influence their biological properties, including metastatic potential (2). The powerful effect of this microenvironment has also been observed in malignant melanoma via convincing models, where melanoma cells acquired properties of melanocyte precursors (neural crest cells) following grafting to vertebrate embryos (3). The embryonic microenvironment here eliminates their ability to form tumors (3). Intercellular crosstalk between different cell types occurring in melanoma (4,5) may be relevant for tumor biology in general.
As fibroblasts represent a fundamental component of various tissues, they also have an indispensable role in cancer (2). The broad biological activity of melanoma-associated fibroblasts (MAFs) has been described and notably, they are also able to stimulate other cancer types than melanoma in vitro (4,6-9). MAFs, as well as other cancer-associated fibroblasts, express smooth muscle actin (SMA) (10,11) in the majority of tumor types (2). The transition of fibroblasts to SMA-expressing myofibroblasts is primarily stimulated by transforming growth factor-β1 (TGF-β1), which is frequently elevated in sera of melanoma patients. An additive effect of endogenous lectin, galectin-1, has also been reported in this context (10-12).
The present report details the comparison of MAFs prepared from a cutaneous melanoma metastasis and autolo-gous control fibroblasts (ACF) from non-cancerous skin of the same patient. The former was isolated prior to therapy initiation, and the latter was isolated during B-Raf inhibitor therapy, yet before the onset of computed tomography (CT)-documented disease progression. Such a matched pair of stromal fibroblasts of human origin is notably rare for analysis. The data obtained from cell cultures are compared with immunohistochemical analysis of sections from the melanoma metastasis including the expression of TGF-β1. The effect of this cytokine on normal dermal fibroblasts prepared from the skin of healthy donors was also examined.
Material and methods
Case report, tissue collection and processing
The patient, a 76 year-old Caucasian female who was diagnosed with acrolentiginous melanoma of the left sole (Breslow, 4.0 mm; Clark, IV) (13) in 2004, was recruited in January 2011 at the Department of Dermatology and Venereology, First Faculty of Medicine, Charles University (Prague, Czech Republic). CT scanning was used throughout observation of the patient (Fig. 1), and 7 years following wide surgical excision, multiple metastases were detected on the lateral parts of the chest and in axillary lymph nodes (Fig. 1A). One cutaneous metastasis on the chest was surgically removed for histologic confirmation (Figs. 1A and 2) of the disease relapse and also for B-Raf V600E mutation screening. MAFs were isolated from the same tumor mass using a previously described method (14,15). B-Raf V600E mutation was confirmed (Fig. 3) and treatment with the B-Raf inhibitor vemurafenib was initiated. Following 3 months of vemurafenib therapy, unaffected skin from the anatomically comparable site (Fig. 1B) was harvested via punch biopsy to isolate ACFs. At this stage, partial response was achieved using RECIST criteria (16). Following further 6 months, CT documented rapid disease progression and the patient succumbed to mortality 5 months later.
Figure 1.

CT scan of the patient's chest. (A) Skin metastasis of melanoma used for the analysis (white circle). (B) Control CT scan after 3 months on vemurafenib treatment; distant uninvolved skin (white circle), a biopsy from this site was used for isolation of autologous control fibroblasts. CT, computed tomography.
Figure 2.

(A) Histology of malignant melanoma metastasis (biopsy from Fig. 1A) with hematoxylin and eosin staining; magnification, ×200. (B) Immunohistochemical analysis (hematoxylin counterstained) confirmed smooth muscle actin-positive myofibroblasts (brown) of tumor stroma between tumor lobules and also (C) transforming growth factor β1intensely positive melanoma cells in the lobules of melanoma metastasis. (D) Negative control.
Figure 3.
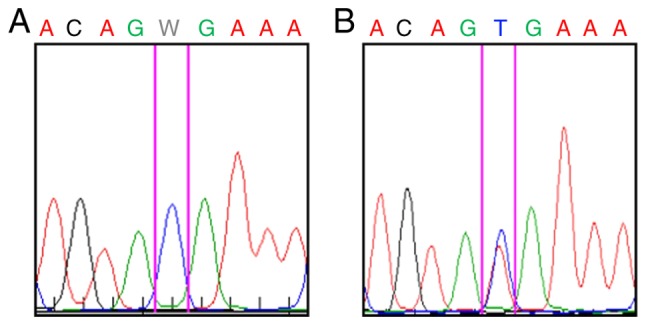
(A) B-Raf V600E mutation was not detected in melanoma-associated fibroblasts. (B) However, this mutation was confirmed in the same metastasis. W denotes A/T mutation.
Control dermal fibroblasts (CDF) were prepared from anatomically comparable chest skin of three healthy Caucasian women donors (aged 35-42 years), undergoing aesthetic surgery (recruited at the same department between March 2011 and September 2013). All tissue samples used in the present study were obtained following the provision of explicit written informed consent, with a protocol reviewed by a local ethics committee (FWA 00003027; General University Hospital, Prague, Czech Republic) with full respect to the Declaration of Helsinki. Institutional consent for publication and data maintenance was also issued.
Cell culture
MAFs, ACFs and CDFs were isolated from tissue samples as described previously (14,15). Passage 3 or 4 cells were used in the present study for analysis. For immunocytochemistry, RNA and DNA studies, fibroblasts were cultured for 5 days before they reached subconfluent monolayers in Dulbecco's modified Eagle's medium containing 10% fetal bovine serum and antibiotics (penicillin/streptomycin; 10.0 U/ml and 10.0 μg/ml, respectively; all, Biochrom, Ltd., Cambridge, UK) at 37°C and 5% CO2. To evaluate the induction of SMA in CDF, TGF-β1 (R&D Systems, Inc., Minneapolis, MN, USA) was supplemented to the culture medium at 10 ng/ml (11).
B-Raf sequencing
For DNA extraction, paraffin-embedded formalin-fixed (4% formaldehyde in PBS for 24-48 h at room temperature) tissue sections (thickness, 5 μm) were deparaffinized using sequential washes with xylene and absolute ethanol. Sections were digested using proteinase K and DNA was extracted using NucleoSpin FFPE DNA (both, Machery-Nagel GmbH, Düren, Germany), according to the manufacturer's protocol. DNA extraction from fibroblasts was performed using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA) according to the manufacturer's protocol. The concentration of DNA was measured using a fluorometer Qubit 2.0 (Invitrogen; Thermo Fisher Scientific, Inc.).
DNA extracted from MAFs was selectively amplified to generate a 224-bp sequence of BRAF exon 15 using polymerase chain reaction (PCR) with the following primers: Forward, 5′-TCATAATGCTTGCTCTGATAGGA-3′ and reverse, 5′-GGCCAAAAATTTAATCAGTGGA-3′ (15). A total of 30 ng DNA was amplified with 0.1 μM each primer, 160 μM deoxynucleotide triphosphates, 2 mM MgCl2, 0.3 U Gold AmpliTaq Perkin Polymerase, and 1X PCR Buffer II (10 mM Tris-HCl pH 8.3, 50 mM KCl) in a final volume of 15 μl (both from Thermo Fisher Scientific, Inc). Cycling conditions were as follows: Denaturation step at 94°C for 2 min and at 95°C for 20 sec followed by 5 cycles of denaturation at 95°C for 20 sec, annealing at 62.5°C for 30 sec and primer extension at 72°C for 1 min; 35 cycles of denaturation at 95°C for 20 sec, annealing at 57.5°C for 30 sec and primer extension at 72°C for 1 min; and one final run-off extension at 72°C for 10 min (16). PCR was performed in a TGradient 96 thermocycler (Biometra GmbH, Göttingen, Germany). PCR products were visualized via 1.5% agarose gel electrophoresis in 0.5X Tris-borate-EDTA buffer (Thermo Fisher Scientific, Inc). Subsequently, the PCR products were purified using an Agencourt AMpure XP (Beckman Coulter, Inc., Brea, CA, USA) and sequenced using forward primers of the PCR amplification and a DTCS Quick Start kit according to the manufacturer's protocol (Beckman Coulter, Inc). The cycle sequencing conditions consisted of 30 cycles of 96°C for 20 sec, 50°C for 20 sec, and 60°C for 4 min. The PCR products were re-purified using an Agencourt CleanSEQ kit (Beckman Coulter, Inc.) according to the manufacturer's protocols, sequenced on a CEQ 8000 sequencing machine (Beckman Coulter, Inc.), and analysed using CEQ 8000 sequencing software. Sequence traces were compared with BRAF reference sequence (NM_004333.4; https://www.ncbi.nlm.nih.gov/nuccore/NM_004333).
Immunohistochemistry and immunocytochemistry
Immunohistochemistry was routinely performed using the avidin-biotin complex method. Briefly, tissue was fixed for 24-48 h in 4% formaldehyde (in PBS) at room temperature and embedded in paraffin. Tissue sections (5-um tick; rehydrated through xylene and ethanol) were washed with PBS with 0.2% Tween-20, and heat-induced epitope retrieval was performed in citrate buffer (pH 6.0, in autoclave at 120°C for 3 min with slow gradual cooling for 60 min. All chemicals for immunohistochemistry were supplied by Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Tissue sections were blocked using Protein Block system (Dako; Agilent Technologies, Inc., Santa Clara, CA, USA; cat. no. X0909) at room temperature for 10 min according to the manufacturer's protocol, followed by 3% hydrogen peroxide (in PBS) treatment for 20 min (all from Sigma-Aldrich; Merck KGaA). Consequently sections were incubated overnight at 4°C with biotinylated antibodies directed against SMA (1:100; Dako; Agilent Technologies, Inc.; cat. no. M0851; clone 1A4), and TGF-β1 (1:100, Bioss Antibodies, Inc., Woburn, MA, USA; cat. no. bs-0086). A Dako Streptavidin Peroxidase kit (including 50X 3,3′-diaminobenzidine- horseradish peroxidase substrate buffer) was used for visualization of immunohistochemical reaction according to the manufacturer's protocol (Dako; Agilent Technologies, Inc.; cat. no. K5001). Nuclei were counterstained with hematoxylin and mounted in permanent mountant.
The cultured cells on coverslips were briefly fixed in 2% paraformaldehyde in PBS for 5 min at room temperature and permeabilized with Triton X-100 (Sigma-Aldrich; Merck KGaA). Antibodies against SMA (1:100; mouse monoclonal; Dako; Agilent Technologies, Inc., cat.no. M0851; clone 1A4) and fibronectin (1:1,000; rabbit polyclonal; Dako; Agilent Technologies, Inc.; cat. no. A0245) were used to detect protein levels, via incubations for 2 h at room temperature. Blocking was performed in 10% goat serum for 30 min at room temperature. Results of immunocytochemical reaction were visualized using tetramethylrhodamine-conjugated goat anti-rabbit (cat. no. T5393; 1:250) and fluorescein isothiocyanate-conjugated goat anti-mouse antibody (cat. no. AP307F; 1:250; both, Sigma-Aldrich; Merck KGaA) for 60 min at room temperature). Nuclei were counterstained with DAPI for 1 min at room temperature (Sigma-Aldrich; Merck KGaA).
The negative control of all staining procedures was performed by replacement of primary antibody with appropriate isotype control (Universal Negative Control; mouse cat. no. N1698; rabbit cat. no. N1699; Dako; Agilent Technologies, Inc.). Following mounting in Vectashield (Vector Laboratories, Inc., Burlingame, CA, USA) samples were analyzed at ×200 magnification using an ECLIPSE-90i fluorescence microscope (Nikon Corp., Tokyo, Japan) equipped with suitable filter blocks, a charge-couple device camera (Vosskühler, Osnabrück, Germany) and a system for computer-assisted image analysis (LUCIA 5.1, Laboratory Imaging s.r.o., Prague, Czech Republic).
Transcriptome analysis
Total RNA was isolated from MAFs, ACFs and CDFs using an RNeasy Micro kit (Qiagen Sciences, Inc., Gaithersburg, MD, USA) according to the manufacturer's protocol. Quality and concentration of RNA were measured with a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Inc.). RNA integrity was analysed using an Agilent Bioanalyzer 2100 (Agilent Technologies, Inc.). Only samples with an intact RNA profile were used for expression profiling analyses (RNA Integrity Number >9).
Illumina HumanHT-12 v4 Expression BeadChips (Illumina, Inc., San Diego, CA, USA) were used for the microarray analysis following the standard protocol. Briefly, 200 ng RNA was amplified with the Illumina TotalPrep RNA Amplification kit (Ambion; Thermo Fisher Scientific, Inc.) and 750 ng labelled RNA was hybridized on the chip according to the manufacturer's protocol. Analysis was performed in four (CDF, 2 donors) and two (MAF and ACF) replicates per group. The raw data were preprocessed using GenomeStudio software (version 1.9.0.24624; Illumina, Inc.) and the limma package (19) of Bioconductor (20), as described previously (21). The transcription profiles were background corrected using a normal-exponential model, quantile normalized and variance stabilized using base 2 logarithmic transformation. Moderated t-test was used to detect differentially expressed transcripts within limma (19). False discovery rates (FDR) were used to select significantly differentially transcribed genes (FDR <0.05), as presented in Table I. Principal components analysis (PCA) was performed using the made4 (22) package of the Bioconductor on expression data of the genes that were found differentially expressed in at least one of the comparisons.
Table I.
Significantly deregulated genes that are differentially expressed in comparisons of MAFs, ACFs and CDFs.
| A, Upregulated genes, MAF vs. CDF
| |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Entrez gene ID | EnsEMBL gene ID | Symbol | Definition | Descriptive GO BP term | FDR | CDF log2-intensity | MAF log2-intensity | ACF log2-intensity | |
| 3036 | ENSG00000105509 | HAS1 | Hyaluronan synthase 1 | GO:0085029 | 8E-23 | 4.50 | 7.62 | 4.50 | |
| 221303 | ENSG00000183807 | FAM162B | Family with sequence similarity 162, member B | – | 3E-22 | 4.48 | 7.85 | 5.10 | |
| 3875 | ENSG00000111057 | KRT18 | Keratin 18 | GO:0009653 | 2E-20 | 5.45 | 9.76 | 8.93 | |
| 894 | ENSG00000118971 CC | ND2 | Cyclin D2 | GO:0008284 | 2E-20 | 5.00 | 9.10 | 6.85 | |
| 8076 | ENSG00000197614 | MFAP5 | Microfibrillar associated protein 5 | GO:0030198a | 2E-19 | 7.73 | 12.06 | 12.24 | |
| 11341 | ENSG00000164106 | SCRG1 | Stimulator of chondrogenesis 1 | GO:0007399 | 2E-19 | 4.55 | 8.26 | 7.07 | |
| 5909 | ENSG00000076864 | RAP1GAP | RAP1 GTPase activating protein | GO:0043087 | 2E-19 | 4.50 | 7.59 | 5.10 | |
| 353139 | ENSG00000187173 | LCE2A | Late cornified envelope 2A | GO:0030216 | 3E-19 | 4.96 | 7.77 | 5.34 | |
| 1776 | ENSG00000163687 D | NASE1L3 | Deoxyribonuclease I-like 3 | GO:0006309 | 7E-18 | 4.56 | 7.51 | 5.79 | |
| 284085 | ENSG00000265480 | KRT18P55 | Keratin 18 pseudogene 55 | – | 1E-17 | 4.68 | 8.37 | 7.04 | |
| 3202 | ENSG00000106004 | HOXA5 | Homeobox A5 | GO:0048704a | 2E-17 | 7.30 | 10.72 | 7.39 | |
| 1829 | ENSG00000046604 D | SG2 | Desmoglein 2 | GO:0007155 | 6E-17 | 4.65 | 7.78 | 4.92 | |
| 3887 | ENSG00000205426 | KRT81 | keratin 81 | GO:0005198 | 7E-17 | 5.67 | 8.93 | 6.94 | |
| B, Downregulated genes, MAF vs. CDF
| ||||||||
|---|---|---|---|---|---|---|---|---|
| Entrez gene ID | EnsEMBL gene ID | Symbol | Definition | Descriptive GO BP term | FDR | CDF log2-intensity | MAF log2-intensity | ACF log2-intensity |
| 404266 | ENSG00000233101 | HOXB-AS3 | HOXB cluster antisense RNA 3 (non- protein coding) | – | 2E-21 | 9.54 | 5.92 | 9.78 |
| 8988 | ENSG00000169271 | HSPB3 | Heat shock 27 kDa protein 3 | GO:0006986 | 6E-18 | 10.12 | 5.73 | 9.35 |
| 6387 | ENSG00000107562 C | XCL12 | Chemokine (C-X-C motif) ligand 12 | GO:0001666a | 9E-18 | 10.36 | 4.97 | 10.19 |
| 1396 | ENSG00000213145 C | RIP1 | Cysteine-rich protein 1 (intestinal) | GO:0008283 | 1E-17 | 11.03 | 7.10 | 8.73 |
| 160364 | ENSG00000172322 | CLEC12A | C-type lectin domain family 12, member A | – | 2E-17 | 5.99 | 4.53 | 4.48 |
| 4147 | ENSG00000132561 | MATN2 | Matrilin 2 | GO:0008347a | 5E-17 | 10.57 | 6.86 | 9.11 |
| 160364 | ENSG00000172322 | CLEC12A | C-type lectin domain family 12, member A, transcript variant 2 | – | 8E-17 | 6.64 | 4.51 | 4.55 |
| C, Upregulated genes, MAF vs. ACF
| ||||||||
|---|---|---|---|---|---|---|---|---|
| Entrez gene ID | EnsEMBL gene ID | Symbol | Definition | Descriptive GO BP term | FDR | CDF log2-intensity | MAF log2-intensity | ACF log2-intensity |
| 3036 | ENSG00000105509 | HAS1 | Hyaluronan synthase 1 | GO:0085029 | 1E-22 | 4.50 | 7.62 | 4.50 |
| 221303 | ENSG00000183807 | FAM162B | Family with sequence similarity 162, member B | – | 3E-19 | 4.48 | 7.85 | 5.10 |
| 3202 | ENSG00000106004 | HOXA5 | Homeobox A5 | GO:0048704a | 7E-17 | 7.30 | 10.72 | 7.39 |
| 353139 | ENSG00000187173 | LCE2A | Late cornified envelope 2A | GO:0030216 | 1E-16 | 4.96 | 7.77 | 5.34 |
| 5909 | ENSG00000076864 | RAP1GAP | RAP1 GTPase activating protein | GO:0043087 | 5E-16 | 4.50 | 7.59 | 5.15 |
| 1829 | ENSG00000046604 D | SG2 | Desmoglein 2 | GO:0007155 | 5E-15 | 4.65 | 7.78 | 4.92 |
| 55966 | ENSG00000196581 | AJAP1 | Adherens junctions associated protein 1 | GO:0007155 | 1E-13 | 4.60 | 6.16 | 4.37 |
| 1300 | ENSG00000123500 C | OL10A1 | Collagen, type X, α1 | GO:0030198a | 2E-14 | 5.11 | 7.78 | 4.93 |
| 3778 | ENSG00000156113 | KCNMA1 | Potassium large conductance calcium- activated channel, subfamily M, α member 1 | GO:0001666a | 2E-14 | 9.64 | 9.46 | 6.95 |
| 794 | ENSG00000172137 C | ALB2 | Calbindin 2 | GO:0005509 | 8E-14 | 6.78 | 8.33 | 4.98 |
| D, Downregulated genes, MAF vs. ACF
| ||||||||
|---|---|---|---|---|---|---|---|---|
| Entrez gene ID | EnsEMBL gene ID | Symbol | Definition | Descriptive GO BP term | FDR | CDF log2-intensity | MAF log2-intensity | ACF log2-intensity |
| 404266 | ENSG00000233101 | HOXB-AS3 | HOXB cluster antisense RNA 3 (non-protein coding) | – | 3E-21 | 9.54 | 5.92 | 9.78 |
| 116154 | ENSG00000087495 | PHACTR3 | Phosphatase and actin regulator 3 | GO:0043086 | 2E-19 | 7.40 | 5.32 | 10.49 |
| 72 | ENSG00000163017 | ACTG2 | Actin, γ2, smooth muscle, enteric | GO:0090131 | 2E-18 | 4.84 | 4.58 | 12.18 |
| 70 | ENSG00000159251 | ACTC1 | Actin, α, cardiac muscle 1 | GO:0090131 | 7E-17 | 9.89 | 7.27 | 12.76 |
| 6387 | ENSG00000107562 C | XCL12 | Chemokine (C-X-C motif) ligand 12 | GO:0001666a | 7E-17 | 10.36 | 4.97 | 10.19 |
| 154197 | ENSG00000146453 | PNLDC1 | Poly(A)-specific ribonuclease (PARN)-like domain containing 1 | – | 3E-16 | 4.49 | 4.45 | 5.51 |
| 8988 | ENSG00000169271 | HSPB3 | Heat shock 27 kDa protein 3 | GO:0006986 | 8E-15 | 10.12 | 5.73 | 9.35 |
| 90139 | ENSG00000157570 | TSPAN18 | Tetraspanin 18 | GO:0030198 | 8E-15 | 5.69 | 4.90 | 8.26 |
| 668 | ENSG00000183770 | FOXL2 | Forkhead box L2 | GO:0030154 | 3E-14 | 7.66 | 4.79 | 8.16 |
| 5121 | ENSG00000183036 | PCP4 | Purkinje cell protein 4 | GO:0007417 | 1E-13 | 4.53 | 4.44 | 7.16 |
| E, Upregulated genes, ACF vs. CDF
| ||||||||
|---|---|---|---|---|---|---|---|---|
| Entrez gene ID | EnsEMBL gene ID | Symbol | Definition | Descriptive GO BP term | FDR | CDF log2-intensity | MAF log2-intensity | ACF log2-intensity |
| 8076 | ENSG00000197614 | MFAP5 | Microfibrillar associated protein 5 | GO:0030198a | 2E-18 | 7.73 | 12.06 | 12.24 |
| 72 | ENSG00000163017 | ACTG2 | Actin, γ2, smooth muscle, enteric | GO:0090131 | 4E-17 | 4.84 | 4.58 | 12.18 |
| 3875 | ENSG00000111057 | KRT18 | Keratin 18 | GO:0009653 | 4E-16 | 5.45 | 9.76 | 8.93 |
| 85409 | ENSG00000145506 | NKD2 | Naked cuticle homolog 2 (Drosophila) | GO:0030178 | 7E-16 | 4.45 | 7.27 | 7.70 |
| 154197 | ENSG00000146453 | PNLDC1 | Poly(A)-specific ribonuclease (PARN)-like domain containing 1 | – | 2E-15 | 4.49 | 4.45 | 5.51 |
| 8322 | ENSG00000174804 | FZD4 | Frizzled family receptor 4 | GO:0060070 | 3E-15 | 6.85 | 8.31 | 9.66 |
| 64798 | ENSG00000155792 | DEPTOR | DEP domain containing MTOR-interacting protein | GO:0035556 | 1E-13 | 4.79 | 6.71 | 8.64 |
| 11341 | ENSG00000164106 | SCRG1 | Stimulator of chondrogenesis 1 | GO:0007399 | 2E-13 | 4.55 | 8.26 | 7.07 |
| 54518 | ENSG00000077420 | APBB1IP | Amyloid beta (A4) precursor protein-binding. family B, member 1 interacting protein | GO:0045785a | 2E-13 | 7.60 | 9.39 | 9.57 |
| 157506 | ENSG00000121039 | RDH10 | Retinol dehydrogenase 10 (all-trans) | GO:0014032 | 3E-13 | 7.38 | 9.37 | 10.58 |
| 5121 | ENSG00000183036 | PCP4 | Purkinje cell protein 4 | GO:0007417 | 3E-13 | 4.53 | 4.44 | 7.16 |
| 3751 | ENSG00000184408 | KCND2 | Potassium voltage-gated channel. Shal-related subfamily, member 2 | GO:0071456 | 3E-13 | 4.42 | 6.28 | 6.14 |
| F, Downregulated genes, ACF vs. CDF
| ||||||||
|---|---|---|---|---|---|---|---|---|
| Entrez gene ID | EnsEMBL gene ID | Symbol | Definition | Descriptive GO BP term | FDR | CDF log2-intensity | MAF log2-intensity | ACF log2-intensity |
| 4885 | ENSG00000106236 | NPTX2 | Neuronal pentraxin II | – | 4E-17 | 9.10 | 6.41 | 5.30 |
| 100133941 | ENSG00000272398 | CD24 | CD24 molecule | GO:0002237a | 3E-16 | 10.02 | 7.09 | 6.68 |
| 160364 | ENSG00000172322 | CLEC12A | C-type lectin domain family 12. member A | – | 4E-16 | 5.99 | 4.53 | 4.48 |
| 160364 | ENSG00000172322 | CLEC12A | C-type lectin domain family 12. member A. transcript variant 2 | – | 1E-15 | 6.64 | 4.51 | 4.55 |
| 3778 | ENSG00000156113 | KCNMA1 | Potassium large conductance calcium-activated channel. Subfamily M. α member 1 | GO:0001666a | 9E-15 | 9.64 | 9.46 | 6.95 |
| 84675 | ENSG00000147573 | TRIM55 | Tripartite motif containing 55 | GO:0007165 | 3E-13 | 6.31 | 4.70 | 4.47 |
| 6228 | ENSG00000186468 | RPS23 | Ribosomal protein S23 | GO:0005840 | 6E-13 | 11.47 | 8.85 | 8.83 |
| 85352 | ENSG00000138944 | KIAA1644 | KIAA1644 | – | 7E-13 | 9.68 | 6.61 | 7.14 |
The 20 most marked DEGs are listed.
Listed among GO terms enriched by DEGs as identified by gene set enrichment analysis. Significance level, FDR<0.05. MAF, melanoma-associated fibroblast; ACF, autologous control fibroblast; CDF, control dermal fibroblast; FDR, false discovery rate; DEG, differentially expressed gene; GO BP, gene ontology biological process.
Genome methylation analysis
Genomic DNA was isolated using the aforementioned standard techniques and purified using a Qiagen DNeasy kit (Qiagen Sciences, Inc.) according to the manufacturer's protocol. Quality and concentration of DNA were measured with a NanoDrop 1000 spectrophotometer (Thermo Fisher Scientific, Inc.). Bisulfite conversion was performed using an EZ DNA Methylation kit (Zymo Research Corp., Irvine, CA, USA).
Illumina Infinium HumanMethylation27 BeadChips (Illumina, Inc.) were used for microarray analysis following the manufacturer's protocol. In brief, 400 ng bisulphite-converted DNA was amplified using an Infinium Methylation Assay kit (Illumina, Inc.) and 750 ng of labelled DNA was hybridized on the chip according to the manufacturer's procedure. The analysis was performed in two replicates per group (MAF, ACF and CDF). The raw data were preprocessed using GenomeStudio software to obtain methylated and unmethylated probe intensities, and the Bioconductor (20) package methylumi to background correct and normalize the data, and to calculate M-values according to the following formula: M=log2((methylated_probe_intensity + 100)/(unmethylated_ probe_intensity + 100)). The positions of CpG islands were previously retrieved in human genomic DNA by Wu et al (23). CpG shores (and shelves) were defined as 2-kbp regions that flanked CpG islands (or shores, respectively).
Moderated t-test implemented in the limma package (19) was used to detect differentially methylated genomic regions (DMR) based on change in M-value and false discovery rate (FDR<0.1, δM>1). PCA of the methylation data was performed using the made4 package on M-values of regions that were considered DMR in at least one comparison.
The MIAME compliant expression and methylation data were subsequently deposited to the ArrayExpress database (accessions E-MTAB-4964 and E-MTAB-4965; https://www.ebi.ac.uk/arrayexpress/browse.html).
Gene set enrichment analysis (GSEA)
GSEA was performed using Enrichr (24). Briefly, differentially expressed genes and differentially methylated genomic regions, respectively, that were specific for MAF or CDF were uploaded to the Enrichr web interface (http://amp.pharm.mssm.edu/Enrichr/). The analysis was performed with default parameters and with equal weight on each gene (or genomic region, in cases of methylation). Analysis of various gene set collections was performed, including the gene ontology (GO) terms and KEGG pathways, only the analysis of the GO terms, and in particular the biological process ontology, are reported. Other analyses resulted in the identification of similar gene sets. For expression data, the 20 best terms (i.e. those with the greatest significance) for each comparison are reported in Table II. The selection roughly corresponds to the terms with an adjusted P-value of the enrichment test of P<0.03. The analysis of DMRs did not result in any significantly enriched GO terms.
Table II.
Results of gene set enrichment analysis performed on differentially expressed genes and GO BP terms.
| A, (MAF and ACF) vs. CDF
| |||
|---|---|---|---|
| GO BP ID | Term | Overlap | Adjusted P-value |
| GO:0030198 | Extracellular matrix organization | 35/359 | 0.00091 |
| GO:0043062 | Extracellular structure organization | 35/360 | 0.00091 |
| GO:0001936 | Regulation of endothelial cell proliferation | 15/88 | 0.00335 |
| GO:0010574 | Regulation of vascular endothelial growth factor production | 8/27 | 0.01791 |
| GO:0002237 | Response to molecule of bacterial origin | 24/243 | 0.01791 |
| GO:0009611 | Response to wounding | 19/167 | 0.01791 |
| GO:0051272 | Positive regulation of cellular component movement | 27/296 | 0.01791 |
| GO:0040017 | Positive regulation of locomotion | 27/304 | 0.02116 |
| GO:2000147 | Positive regulation of cell motility | 26/287 | 0.02116 |
| GO:0001503 | Ossification | 15/116 | 0.02116 |
| GO:0008347 | Glial cell migration | 6/15 | 0.02116 |
| GO:0045765 | Regulation of angiogenesis | 19/179 | 0.02490 |
| GO:0032496 | Response to lipopolysaccharide | 22/228 | 0.02490 |
| GO:0030335 | Positive regulation of cell migration | 25/280 | 0.02511 |
| GO:0030111 | Regulation of Wnt signaling pathway | 21/214 | 0.02515 |
| GO:0060828 | Regulation of canonical Wnt signaling pathway | 17/152 | 0.02544 |
| GO:0051894 | Positive regulation of focal adhesion assembly | 6/18 | 0.02978 |
| GO:1901890 | Positive regulation of cell junction assembly | 6/18 | 0.02978 |
| GO:0001937 | Negative regulation of endothelial cell proliferation | 7/27 | 0.02978 |
| B, (CDF and ACF) vs. MAF
| |||
|---|---|---|---|
| GO BP ID | Term | Overlap | Adjusted P-value |
| GO:0048729 | Tissue morphogenesis | 44/358 | 0.00023 |
| GO:0051272 | Positive regulation of cellular component movement | 39/296 | 0.00023 |
| GO:0030198 | Extracellular matrix organization | 44/359 | 0.00023 |
| GO:0043062 | Extracellular structure organization | 44/360 | 0.00023 |
| GO:2000147 | Positive regulation of cell motility | 37/287 | 0.00055 |
| GO:0030335 | Positive regulation of cell migration | 36/280 | 0.00068 |
| GO:0048598 | Embryonic morphogenesis | 45/403 | 0.00094 |
| GO:0040017 | Positive regulation of locomotion | 37/304 | 0.00118 |
| GO:0030155 | Regulation of cell adhesion | 39/336 | 0.00158 |
| GO:0002009 | Morphogenesis of an epithelium | 35/296 | 0.00312 |
| GO:0048562 | Embryonic organ morphogenesis | 20/121 | 0.00400 |
| GO:0048704 | Embryonic skeletal system morphogenesis | 16/81 | 0.00403 |
| GO:0045785 | Positive regulation of cell adhesion | 24/171 | 0.00507 |
| GO:0036293 | Response to decreased oxygen levels | 30/245 | 0.00507 |
| GO:0009887 | Organ morphogenesis | 42/405 | 0.00507 |
| GO:0070482 | Response to oxygen levels | 31/259 | 0.00516 |
| GO:0071294 | Cellular response to zinc ion | 7/14 | 0.00770 |
| GO:0001666 | Response to hypoxia | 29/241 | 0.00770 |
| GO:0022617 | Extracellular matrix disassembly | 18/116 | 0.01315 |
| GO:0048705 | Skeletal system morphogenesis | 17/106 | 0.01380 |
The 'best' 20 ontology terms are provided for each comparison (i.e. corresponds to the terms with an adjusted P-value of the enrichment test ≥0.03). GO BP, gene ontology biological process; MAF, melanoma-associated fibroblast; ACF, autologous control fibroblast; CDF, control dermal fibroblast.
Results
Properties of fibroblasts isolated from the patients
Fibroblasts in cutaneous metastasis (Fig. 2A) exhibited SMA (Fig. 2B) and immunohistochemistry also revealed that melanoma cells in this metastasis were rich in TGF-β1 (Fig. 2C). MAFs isolated from this metastasis were not harbouring the B-Raf V600E mutation (Fig. 3A). The isolated MAFs in culture exhibited a high cytoplasmic signal of fibronectin and were also sporadically positive for SMA (Fig. 4A).
Figure 4.

(A) Melanoma-associated fibroblasts exhibited fibronectin (green) in the cytoplasm and sporadic cells were SMA positive (red). (B) Autologous control fibroblasts with high incidence of SMA-positive myofibroblasts in a pool of autologous fibroblasts. (C) CDFs did not contain SMA-positive cells and produced only sparse fibro fibres. (D) Stimulation of CDFs by transforming growth factor-β in culture mediaincreased transition of the fibroblasts to SMA-positive myofibroblasts, which also produced a dense fibro-positive extracellular matrix meshwork. Scale bar represents 50 mm. SMA, smooth muscle actin; fibro, fibronectin; CDF, control dermal fibroblast.
This observation was in contrast with ACFs isolated from the normal skin of the same patient following 3 months on vemurafenib, where numerous clusters of SMA-positive myofibroblasts were present in vitro (Fig. 4B). The expression of fibronectin was similar in MAFs (Fig. 4A) and ACFs (Fig. 4B).
In the control experiment performed with CDFs, sparse extracellular fibres of fibronectin were observed, but no myofibroblasts were detected (Fig. 4C). As TGF-β1 was detected in the melanoma metastasis and because the elevation of TGF-β1 in sera of melanoma patients was documented previously (10), the effect of TGF-β1 on CDFs was assessed. TGF-β1 was able to markedly stimulate the transition of fibroblasts to myofibro-blasts and also increase fibronectin production (Fig. 4D).
Comparison of expression profiles of MAFs, ACFs, and CDFs
The expression profiles of MAFs, ACFs and CDFs were compared using microarrays (Fig. 5A–D and Table I). The expression profiles differed between MAFs and ACFs in the expression of 1,800 genes, and between MAFs and CDFs there were 2,701 deregulated genes. Finally, there were 1,314 deregulated genes between ACFs and CDFs (FDR<0.05). Among the genes upregulated in the comparison of MAF and CDF cells, hyaluronan synthase 1 and desmoglein 2, which are both associated with extracellular matrix (ECM) formation, and cytokeratins 18 and 81 were identified. MAF cells exhibited decreased expression of chemokine CXCL12. In comparison of MAFs with ACFs, upregulation of hyaluronan synthase 1 and desmoglein 2 was observed, together with another ECM component, COL10A1. ACF showed upregulation of CXCL12 and cardiac-muscle and smooth-muscle actins (ACTC1 and ACTG2). ACTG2 was also upregulated in ACFs compared with CDFs, where the upregulation of the following Wntsignalling pathway related genes was also observed: Frizzled family receptor 4 and naked cuticle homolog 2.
Figure 5.

MAFs, ACFs and CDFs differ profoundly both in their gene expression profiles and in their genome methylation profiles. (A) Projections of the expression data on the first two PCs, as determined by a PC analysis performed on expression profiles of all differentially expressed genes. PC1 reflects differences between MAF and CDF, with ACF being placed between the stromal and normal fibroblasts. (B and C) Venn diagrams display overlaps of upregulated and downregulated genes. (D) Heatmaps with a comparative analysis of MAF, ACF and CDF present a clear separation of the different fibroblast groups both in their expression profiles. (E) Projections of the methylation data on the first two PCs were plotted as determined by a PC analysis performed on methylation profiles of all differentially methylated genes. (F and G) Venn diagrams display overlaps of hyper-methylated and hypo-methylated regions in various studied comparisons. (H) MAF, ACF and CDF methylation profiles. Only differentially expressed genes and differentially methylated regions are presented. MAF, melanoma-associated fibroblast; ACF, autologous control fibroblast; CDF, control dermal fibroblast; PC, principal component.
Principal component analysis was performed to visualize associations between the analysed groups (Fig. 5A): The location of the groups in the plot and the number of deregulated genes between the groups demonstrated that ACFs differ greatly from CDFs. The association between ACFs and other analysed groups were further analyzed (Fig. 5B and C). Two sets of deregulated genes were defined: a) Genes deregulated in both MAFs and ACFs, and in both MAFs and CDFs; and b) genes deregulated in both ACFs and CDFs, and in both MAFs and CDFs. To identify biological processes deregulated in a) MAFs and b) ACFs and MAFs, GSEA was performed on the respective sets (Table II).
Genes deregulated similarly in ACFs and in CDFs (in comparison with MAFs; set a) participate in control of biological processes related to cell migration and motility (GO:0051272, 2000147, 0030335 and 0048598), morphogenesis, ECM structure (GO:0048729, 0030198 and 0043062), and response to stress factors, including decreased oxygen level (GO:0070482, 0036293, 0071294 and 0001666). At the gene level, the activity of, e.g. CXCL12 and FGF13, were upregulated and was CXCL5 downregulated in ACFs and CDFs in comparison with MAFs.
ACFs share deregulated genes with MAFs (in comparison with CDFs; gene set b), which are associated with changes in angiogenesis and endothelial cell proliferation (GO:0001525, 0001936, 0001936, 0045765, 0045765, and 0001937), ECM structure (GO:0030198 and 0043062), response to wounding (GO:0002237), positive regulation of cell motility and locomotion (GO:2000147 and 0040017), ossification (GO:0001503), and regulation of the Wnt signaling pathway (GO:0030111 and 0060828). At the gene level, marked upregulation of interleukin (IL)6, VEGFA, and TGFB3 genes and downregulation of TGFA was observed in MAFs and ACFs as compared to CDFs. The relevant differentially regulated genes were consequently validated directly at the protein level, therefore quantitative-PCR was not used for validation.
Changes in DNA methylation in MAFs, ACFs and CDFs
To evaluate whether the changes in transcription activity of differently expressed genes were associated with epigenomic changes of the fibroblasts, differentially methylated genomic regions (DMRs) were detected between the cultured fibroblasts using microarray technology. A total of 2,300 DMRs were detected between MAFs and ACFs (787 genes affected in CpG islands, 985 genes affected in CpG shores), between MAFs and CDFs there were 2,908 DMRs (966 genes affected in CpG islands, 1159 genes affected in CpG shores), and finally there were 1,835 DMRs between ACFs and CDFs (649 genes affected in CpG islands, 751 genes affected in CpG shores; FDR<0.1, |logFC|>1; Fig. 5E–H; Table III). PCA analysis (Fig. 5E) distinguished ACF cells from CDFs and indicated that they differ from either MAFs or CDFs, respectively (Fig. 5H). MAFs displayed hyper-methylation of genomic regions associated with an SMA (ACTA2) and fibroblast growth factor binding protein 2 (FGFBP2). The latter was specifically hyper-methylated in MAF cells. ACTA2 was hypo-methylated in normal CDF cells when compared with both MAF and ACF cells.
Table III.
DMRs between fibroblast groups.
| A, Hyper-methylated genes in MAF vs. CDF
| ||||||||
|---|---|---|---|---|---|---|---|---|
| RefSeq ID | Entrez Gene ID | Symbol | Definition | Descriptive GO BP term | FDR | CDF M-value | MAF M-value | ACF M-value |
| NM_001141945 | 59 | ACTA2 | Actin, α2, smooth muscle, aorta | GO:0090131 | 6.0E-07 | −3.48 | 3.43 | 1.72 |
| NM_006308 | 8988 | HSPB3 | Heat shock 27 kDa protein 3 | – | 7.6E-07 | −3.32 | 3.07 | 2.09 |
| NM_001202233 | 3164 | NR4A1 | Nuclear receptor subfamily 4. group A, member 1 | GO:0044344 | 7.6E-07 | 0.07 | 4.86 | 4.06 |
| NM_031950 | 83888 | FGFBP2 | Fibroblast growth factor binding protein 2 | – | 7.6E-07 | −3.13 | 1.69 | −2.15 |
| NM_001307983 | 130013 | ACMSD | Aminocarboxymuconate semialdehyde decarboxylase | GO:1905004 | 7.6E-07 | −2.15 | 3.15 | 1.12 |
| NM_001256552 | 79661 | NEIL1 | Nei-like DNA glycosylase 1 | GO:0010730 | 7.6E-07 | −1.86 | 3.19 | 0.87 |
| NM_001039469 | 2011 | MARK2 | MAP/microtubule affinity-regulating kinase 2 | GO:0016055 | 7.6E-07 | −3.05 | 1.15 | 1.01 |
| NM_001193341 | 221421 | RSPH9 | Radial spoke head 9 homolog (Chlamydomonas) | GO:0044458 | 7.6E-07 | −3.74 | 0.46 | −2.33 |
| NM_033199 | 90226 | UCN2 | Urocortin 2 | GO:0031669 | 7.6E-07 | −3.43 | 1.19 | 0.14 |
| B, Hypo-methylated genes in MAF vs. CDF
| ||||||||
|---|---|---|---|---|---|---|---|---|
| RefSeq ID | Entrez Gene ID | Symbol | Definition | Descriptive GO BP term | FDR | CDF M-value | MAF M-value | ACF M-value |
| NM_001199505 | 116154 | PHACTR3 | Phosphatase and actin regulator 3 | GO:0043086 | 7.6E-07 | 3.37 | −2.73 | 0.74 |
| NM_144719 | 152206 CCDC | 13 | Coiled-coil domain containing 13 | GO:0035058 | 7.6E-07 | 2.05 | −2.57 | −0.17 |
| NM_000438 | 5077 | PAX3 | Paired box 3 | GO:0009887a | 7.6E-07 | 2.96 | −1.53 | 2.98 |
| NM_002151 | 3249 | HPN | Hepsin | GO:0010719 | 7.6E-07 | 4.09 | −1.15 | 1.34 |
| NM_153038 | 151278 CCDC | 140 | Coiled-coil domain containing 140 | – | 7.6E-07 | 2.40 | −2.86 | 2.87 |
| NM_000384 | 338 | APOB | Apolipoprotein B | GO:0032496a | 7.6E-07 | 2.90 | −2.93 | 0.66 |
| C, Hyper-methylated genes in MAF vs. ACF
| ||||||||
|---|---|---|---|---|---|---|---|---|
| RefSeq ID | Entrez Gene ID | Symbol | Definition | Descriptive GO BP term | FDR | CDF M-value | MAF M-value | ACF M-value |
| NM_001242805 | 676 | BRDT | Bromodomain, testis-specific | GO:0006338 | 4.2E-06 | 1.15 | 3.62 | −1.43 |
| NM_001300905 | 11176 | BAZ2A | Bromodomain adjacent to zinc finger domain, 2A | GO:0006338 | 5.1E-06 | −4.52 | −0.9 | −4.29 |
| NM_018593 | 117247 | SLC16A10 | Solute carrier family 16 (aromatic amino acid transporter). Member 10 | GO:0006865 | 4.2E-06 | −3.50 | 1.46 | −3.98 |
| NM_005992 | 6899 | TBX1 | T-box 1 | GO:0001525 | 4.2E-06 | 0.79 | 1.60 | −2.20 |
| NM_178818 | 146223 | CMTM4 | CKLF-like MARVEL transmembrane domain containing 4 | GO:0005125 | 4.2E-06 | −2.70 | 1.35 | −2.46 |
| NM_000663 | 18 | ABAT | 4-aminobutyrate aminotransferase | GO:0001666a | 4.2E-06 | −4.14 | −0.07 | −4.11 |
| NM_006829 | 10974 | ADIRF | Adipogenesis regulatory factor | GO:0030154 | 4.2E-06 | −4.29 | 0.31 | −4.32 |
| NM_006181 | 4917 | NTN3 | Netrin 3 | GO:0007411 | 4.2E-06 | −4.54 | −1.14 | −4.78 |
| NM_031950 | 83888 | FGFBP2 | Fibroblast growth factor binding protein 2 | – | 4.7E-06 | −3.13 | 1.69 | −2.15 |
| NM_001720 | 656 | BMP8B | Bone morphogenetic protein 8b | GO:0001503a | 4.7E-06 | −4.66 | −0.79 | −4.55 |
| NM_005731 | 10109 | ARPC2 | Actin related protein 2/3 complex. subunit 2, 34 kDa | GO:1900026 | 5.1E-06 | −3.37 | −0.84 | −4.31 |
| D, Hypo-methylated genes in MAF vs. ACF
| ||||||||
|---|---|---|---|---|---|---|---|---|
| RefSeq ID | Entrez Gene ID | Symbol | Definition | Descriptive GO BP term | FDR | CDF M-value | MAF M-value | ACF M-value |
| NM_000192 | 6910 | TBX5 | T-box 5 | GO:0002009a | 1.3E-06 | −2.27 | −3.24 | 3.75 |
| NM_153038 | 151278 CCDC | 140 | Coiled-coil domain containing 140 | – | 3.4E-06 | 2.40 | −2.86 | 2.87 |
| NM_000438 | 5077 | PAX3 | Paired box 3 | GO:0009887a | 4.2E-06 | 2.96 | −1.53 | 2.98 |
| NM_001053 | 6755 | SSTR5 | Somatostatin receptor 5 | GO:0007186 | 4.2E-06 | −0.75 | −4.83 | −0.62 |
| NM_152739 | 3205 | HOXA9 | Homeobox A9 | GO:0007275 | 4.2E-06 | 0.09 | −3.76 | 0.28 |
| NM_015916 | 51063 C | ALHM2 | Calcium homeostasis modulator 2 | GO:0005261 | 5.1E-06 | −4.79 | −4.84 | −1.16 |
| E, Hyper-methylated genes in ACF vs. CDF
| ||||||||
|---|---|---|---|---|---|---|---|---|
| RefSeq ID | Entrez Gene ID | Symbol | Definition | Descriptive GO BP term | FDR | CDF M-value | MAF M-value | ACF M-value |
| NM_000192 | 6910 | TBX5 | T-box 5 | GO:0002009a | 3.6E-06 | −2.27 | −3.24 | 3.75 |
| NM_001141945 | 59 | ACTA2 | Actin. α2, smooth muscle, aorta | GO:0090131 | 3.6E-06 | −3.48 | 3.43 | 1.72 |
| NM_006308 | 8988 | HSPB3 | Heat shock 27 kDa protein 3 | – | 3.6E-06 | −3.32 | 3.07 | 2.09 |
| NM_001039469 | 2011 | MARK2 | MAP/microtubule affinity-regulating kinase 2 | GO:0016055 | 3.6E-06 | −3.05 | 1.15 | 1.01 |
| NM_001202233 | 3164 | NR4A1 | Nuclear receptor subfamily 4. group A. member 1 | GO:0044344 | 4.0E-06 | 0.07 | 4.86 | 4.06 |
| NM_000129 | 2162 | F13A1 | Coagulation factor XIII. A1 polypeptide | GO:0007596 | 4.0E-06 | −4.97 | −0.78 | −1.12 |
| NM_001286555 | 56940 | DUSP22 | Dual specificity phosphatase 22 | GO:0008283 | 5.9E-06 | −3.92 | −0.45 | −0.51 |
| NM_015916 | 51063 | CALHM2 | Calcium homeostasis modulator 2 | GO:0005261 | 5.9E-06 | −4.79 | −4.84 | −1.16 |
| NM_001012631 | 9235 | IL32 | Interleukin 32 | GO:0005125 | 5.9E-06 | −3.35 | 0.37 | 1.29 |
| NM_000504 | 2159 | F10 | Coagulation factor X | GO:0030335a | 6.2E-06 | −0.42 | 3.59 | 3.68 |
| F, Hypo-methylated genes in ACF vs. CDF
| ||||||||
|---|---|---|---|---|---|---|---|---|
| RefSeq ID | Entrez Gene ID | Symbol | Definition | Descriptive GO BP term | FDR | CDF M-value | MAF M-value | ACF M-value |
| NM_001297436 | 3036 | HAS1 | Hyaluronan synthase 1 | GO:0085029 | 4.0E-06 | 1.38 | −4.34 | −4.22 |
| NM_000965 | 5915 | RARB | Retinoic acid receptor, beta | GO:0007165 | 4.0E-06 | −0.57 | −4.43 | −4.66 |
| NM_001166247 | 2898 | GRIK2 | Glutamate receptor, ionotropic, kainate 2 | GO:0046328 | 4.0E-06 | 0.73 | −3.46 | −3.40 |
| NM_001207011 | 1271 | CNTFR | Ciliary neurotrophic factor receptor | GO:0060538 | 5.8E-06 | 2.87 | −2.6 | −1.69 |
| NM_001128423 | 255027 | MPV17L | MPV17 mitochondrial membrane protein-like | GO:0010730 | 6.3E-06 | −0.75 | −3.91 | −4.53 |
The 15 most marked DMRs with FDR<0.1 and |log2FC|>1 are listed.
Listed among GO terms enriched by differentially expressed genes as identified by gene set enrichment analysis. DMR, differentially methylated genomic region; FDR, false discovery rate; M-value, methylation M-value; GO BP, gene ontology biological process; MAF, melanoma-associated fibroblast; ACF, autologous control fibroblast; CDF, control dermal fibroblast.
As in the gene expression analysis, the association between ACFs and other analysed groups was evaluated (Fig. 5F and G) and two sets of deregulated regions were defined: a) Those differentially methylated in both MAFs and ACFs, and MAFs and CDFs, and b) the genes differentially methylated in both ACFs and CDFs, and MAFs and CDFs. GSEA was performed on the respective gene sets. Notably, although there is a large number of DMRs and associated genes, GSEA did not yield significant enrichment of any single GO term (except the general term 'behavior', which is unrelated to the studied phenomenon). Still, it was observed that the genes attributed to the GO terms affected by changes in gene transcription were widely present among DMRs. For MAF specific genes (set a), the GO terms typically contained 25 DMRs (min 23, max 70), with the exception of a single GO term (GO:0071294) with only 3 DMRs. For CDF specific genes (set b), the terms typically contained 17 DMRs (min 7, max 34), with the exception of the GO term GO:0001937 with two DMRs only. Therefore, it may be concluded that, although changes in methylation are not specifically targeted on biological processes manifested by changes in the expression activity, they target these processes to a large degree.
At the gene level, the largest changes in methylation of these selected genes were observed: Actins ACTA1 and ACTA2, the genes coding for SMA; growth factors FGF11, FGF22, and BMP8B; and chemokines and interleukins CXCL12, IL6, and IL32.
Finally, the expression profiles (Fig. 6A) and methylation profiles (Fig. 6B) were evaluated, together with the selected genes with strongest changes in gene expression: Actins ACTC1 and ACTG2; growth factors FGF13, BMP6, and TGFA; chemokines and interleukins CXCL5, -6, -12, IL1B, IL6, IL8, IL11, and IL24; and keratin KRT7. Notably, the expression profiles of the selected genes in ACF samples are typically closer to expression profiles observed in CDF samples, whereas their methylation status is, in general, closer to MAFs (Fig. 6).
Figure 6.

Changes in gene expression and methylation of selected genes with the strongest changes in gene expression and methylation. For the selected genes, heatmaps of (A) gene expression and (B) methylation of genomic regions overlapping the genes loci are presented. Methylation is expressed in terms of M-values. M=−4 [(A) 0; (B) 4] corresponds to ~5% [(A) 50%; (B) 95%] of the DNA in the methylated genomic region. MAF, melanoma-associated fibroblast; ACF, autologous control fibroblast; CDF, control dermal fibroblast.
Discussion
In skin, normal fibroblasts represent a number of distinct differentiated mesenchymal cell types that have different origins, locations and functions (25,26). Furthermore, similar heterogeneity is observed in tumor stromal tissues. The myofibroblast (27), which is a hallmark of cancer stroma, remains one of the most poorly understood cell types. It is clear that current understanding of the myofibroblast (its origins, functions and molecular regulation) will have a profound influence on the future effectiveness of regenerative medicine, tissue engineering and cancer therapy.
To exclude site-specific differences in e.g. gene transcription or DNA methylation, low passage fibroblasts obtained from the comparable anatomically defined region, the upper trunk, were used. Based on this restriction, relatively high homogeneity in gene transcription of CDFs, even from various donors, can be demonstrated.
In the present study, MAFs did not share B-Raf mutations with melanoma cells from skin metastasis. This suggests a likely local origin from the dermis, rather than the epithelial-to-mesenchymal transition from malignant cells. This also excludes MAFs as the primary target of the direct effect of B-Raf inhibitor.
Via the comparison of MAFs and CDFs from healthy donors, distinctive differences were observed. Recruitment of normal dermal cells in the vicinity of melanoma and their conversion to MAFs is classically associated with TGF-β release from melanoma cells in a paracrine manner (27). Notably, ACFs from the same melanoma patient also greatly differed from CDFs from the general population. All gene expression profile differences were confirmed directly on the protein level, therefore omitting the necessity of quantitative-PCR.
The comparison of expression profiles of MAFs, ACFs and CDFs indicated that ACFs exhibit higher activity of the genes responsible for activation of fibroblasts and cancer cells, namely important mediators such as TGF-β or IL6. ACFs also demonstrated deregulation of ACTG2 expression, and of another gene associated with smooth muscle, KCNMA1.
The presence of activated fibroblasts in normal dermis far from the melanoma lesions supports that melanoma is a systemic disease. This idea was initially proposed by Krasagakis et al (12). Furthermore, the present patient was treated with a B-Raf inhibitor which is known to induce cytokine release from melanoma cells in vitro (27). The observed unusual activation of ACF cells may be explained in a similar manner as activation of MAF via production of bioactive substances, namely TGF-β, IL6, IL8, and matrix metalloproteinase-1 by cancer cells in consequence to vemurafenib treatment (28-32).
TGF-β signalling is one of the major pathways controlling cell and tissue behaviour in development and in various human pathologies. During tumorigenesis, TGF-β has a dual role as tumor suppressor and tumor promoter (33). Important functions of this pathway have previously been described in a context-dependent manner both in epithelial cancer cells and in the tumor microenvironment during tumor progression (28,30,33). The TGF-β system signals via protein kinase receptors and SMAD mediators. Alterations of the TGF-β signaling pathway are implicated in numerous types of human cancer (33).
Notably, differential expression of genes in the TGF-β signalling pathway (e.g. SMAD molecules, etc.) have not been observed, despite the fact that the TGF-β signalling pathway serves an important and key role in the tumor microenvironment and cancer progression. This reflects the dependence of SMADs on their phosphorylation status, rather than actual quantity. Furthermore, TGF-β signalling is always reported as 'context-dependent' (27,28,30). In such cases, the critical point is availability of the cytokine (TGF-β) in the tumor tissue.
In a recent study, Fedorenko et al (28) concluded that B-Raf inhibitor treatment escape in melanoma is due to short-term adaptation in which cells evade the immediate effects of the drug in the supportive microenvironment. However, confirmation of relevancy of this murine model for human pathology was (to the best of our knowledge) missing. The present study offers a comparison of this murine model to paired dermal fibroblasts isolated from a single melanoma patient prior to systemic therapy (B-Raf inhibitor) initiation and on treatment. Such paired material is rarely available and the present data is of relevance to the above-mentioned animal model.
The present data suggests a more sustained effect as the ACF cultures were stored without vemurafenib for a substantial duration (≥2 months) during expansion in vitro. Maintenance of this activated phenotype would require more profound cellular changes, e.g. on the epigenetic level. Although dependence of the gene methylation and its expression is complex, and both gene upregulation and downregulation may occur with gene methylation (33), the observed increase of IL6 expression in ACFs may be associated with the observed demethylation of this gene in ACFs. Similarly, an increased expression of keratin 7 was observed in ACFs and MAFs accompanied by its decreased methylation. This unusual presence of simple type keratins in activated fibroblasts has also been described elsewhere (34).
Notably, observed upregulation of SMA expression in ACF cells at the protein level and the transcriptional activation of ACTA2, the gene coding for SMA, is consistent with abrupt hypermethylation of ACTA2 in both ACF and MAF cells. This observation may indicate the association of epigenetic changes in the activation of ACF cells.
Cancer-associated fibroblasts, including MAFs, produce numerous bioactive substances that actively influence cancer cell proliferation, differentiation and migration (15) and are able to strongly influence melanoma cells. Furthermore, cancer-associated fibroblasts, or MAFs specifically, are important drivers of tumor progression, including metastatic spread, and resistance to vemurafenib, as proposed recently (27,30,37). Based on the presented similarities of MAFs and ACFs in the present patient on B-Raf inhibitor treatment, it may be hypothesized that vemurafenib therapy is able to influence not only B-Raf mutated tumor cells, but presumably may also shape the whole landscape of tissue microenvironment in the human body, to some extent. Vast differences have also been documented in control fibroblasts (ACF vs. CDF) on the level of DNA methylation, RNA transcription and also on the protein level. Such an activated microenvironment may be suitable for growth of circulating melanoma cells (2,4,6). The fibronectin rich 'safe haven' may be created either by melanoma-associated fibroblasts or by distant fibroblasts activated by TGF-β1 released from B-Raf therapy-stressed melanoma cells (27-31). Mechanistically, this phenomenon may participate in the induction of resistance to the therapy as depicted in the case of the presented patient.
The key question for the future is whether the present knowledge of fibroblast heterogeneity may be of clinical use in personalized cancer therapy. As indicated above, the ACFs resemble normal dermal fibroblasts of a healthy individual stimulated by TGF-β1. Therefore, it may be hypothesized that therapeutic attenuation of TGF-β activity (38,39), blocking of transition of fibroblasts to myofibroblasts (39) and/or therapeutic blockade of their function (40) may be used to prevent resistance to B-Raf inhibitor therapy in the future. Mechanistically, this blockade would disrupt bi-directional cross-talk between the melanoma cells and stromal fibroblasts which allow the tumors to amplify a drug-resistant niche. Genomic studies have clearly demonstrated the evolution of genetic heterogeneity in melanoma in the course of tumor progression and metastasis formation (41,42). The tumor microenvironment seems to participate in the tumor evolution by the formation of a suitable cellular ecosystem supporting its progression (43).
The present study represents a single clinical case analysis that may be easily linked to the latest development in preclinical melanoma research on animal models (28-31,37,38). Larger cohort-based studies are necessary for the near future for selection of proper candidate additional targets for further improvement of personalized melanoma therapy.
Acknowledgments
The present study is a result of the project implementation: 'The equipment for metabolomic and cell analyses' (registration no. CZ.1.05/2.1.00/19.0400) supported by Research and Development for Innovations Operational Program (RDIOP) co-financed by the European Regional Development Fund and the state budget of the Czech Republic, MH CZ-DRO (Institute of Endocrinology grant no. EU 00023761). The present study was also supported by the Grant Agency of the Czech Republic (grant no. 16-05534S), the Ministry of Health (AZV nos. 16-29032A, 16-32665A and 16-30954A), Charles University (Project of Specific University Research grant nos. PRVOUK-27 and UNCE 23014) and by the Ministry of Education, Youth and Sports of CR within the National Sustainability Program II (Project BIOCEV-FAR reg. no. LQ1604) and by the project BIOCEV (grant no. CZ.1.05/1.1.00/02.0109). Access to computing and storage facilities owned by parties and projects contributing to the National Grid Infrastructure MetaCentrum were provided under the programme 'Projects of Large Research, Development, and Innovations Infrastructures' (grant no. CESNET LM2015042), is greatly appreciated. The present authors are grateful to Marie Jindráková for technical assistance and also to Ms. Pearl Harris for English language editing during manuscript preparation and consequent proofreading.
Glossary
Abbreviations
- ACF
autologous control fibroblasts
- CDF
control dermal fibroblasts
- CT
computed tomography
- MAF
melanoma associated fibroblasts
- PCR
polymerase chain reaction
- SMA
smooth muscle actin
- TGF-β1
transforming growth factor-β1
Footnotes
Competing interests
The authors declare that they have no competing interests.
References
- 1.Forsea AM, Del Marmol V, de Vries E, Bailey EE, Geller AC. Melanoma incidence and mortality in Europe: New estimates, persistent disparities. Br J Dermatol. 2012;167:1124–1130. doi: 10.1111/j.1365-2133.2012.11125.x. [DOI] [PubMed] [Google Scholar]
- 2.Smetana K, Jr, Dvoøánková B, Szabo P, Strnad H, Koláø M. Role of stromal fibroblasts in cancer originated from squamous epithelia. In: Bai X, editor. Dermal Fibroblasts: Histological perspectives, characterization and role in disease. Nova Sciences Publishers; New York, NY: 2013. pp. 83–94. [Google Scholar]
- 3.Kulesa PM, Kasemeier-Kulesa JC, Teddy JM, Margaryan NV, Seftor EA, Seftor RE, Hendrix MJ. Reprogramming metastatic melanoma cells to assume a neural crest cell-like phenotype in an embryonic microenvironment. Proc Natl Acad Sci USA. 2006;103:3752–3757. doi: 10.1073/pnas.0506977103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kodet O, Dvořánková B, Krejčí E, Szabo P, Dvořák P, Štork J, Krajsová I, Dundr P, Smetana K, Jr, Lacina L. Cultivation-dependent plasticity of melanoma phenotype. Tumour Biol. 2013;34:3345–3355. doi: 10.1007/s13277-013-0905-x. [DOI] [PubMed] [Google Scholar]
- 5.Kodet O, Lacina L, Krejčí E, Dvořánková B, Grim M, Štork J, Kodetová D, Vlček Č, Šáchová J, Kolář M, et al. Melanoma cells influence the differentiation pattern of human epidermal keratinocytes. Mol Cancer. 2015;14:1. doi: 10.1186/1476-4598-14-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Li L, Dragulev B, Zigrino P, Mauch C, Fox JW. The invasive potential of human melanoma cell lines correlates with their ability to alter fibroblast gene expression in vitro and the stromal microenvironment in vivo. Int J Cancer. 2009;125:1796–1804. doi: 10.1002/ijc.24463. [DOI] [PubMed] [Google Scholar]
- 7.Comito G, Giannoni E, Di Gennaro P, Segura CP, Gerlini G, Chiarugi P. Stromal fibroblasts synergize with hypoxic oxidative stress to enhance melanoma aggressiveness. Cancer Lett. 2012;324:31–41. doi: 10.1016/j.canlet.2012.04.025. [DOI] [PubMed] [Google Scholar]
- 8.Dvořánková B, Szabo P, Lacina L, Kodet O, Matoušková E, Smetana K., Jr Fibroblasts prepared from different types of malignant tumors stimulate expression of luminal marker keratin 8 in the EM-G3 breast cancer cell line. Histochem Cell Biol. 2012;137:679–685. doi: 10.1007/s00418-012-0918-3. [DOI] [PubMed] [Google Scholar]
- 9.Yin M, Soikkeli J, Jahkola T, Virolainen S, Saksela O, Hölttä E. TGF-β signaling, activated stromal fibroblasts, and cysteine cathepsins B and L drive the invasive growth of human melanoma cells. Am J Pathol. 2012;181:2202–2216. doi: 10.1016/j.ajpath.2012.08.027. [DOI] [PubMed] [Google Scholar]
- 10.Krasagakis K, Thölke D, Farthmann B, Eberle J, Mansmann U, Orfanos CE. Elevated plasma levels of transforming growth factor (TGF)-beta1 and TGF-beta2 in patients with disseminated malignant melanoma. Br J Cancer. 1998;77:1492–1494. doi: 10.1038/bjc.1998.245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Dvořánková B, Szabo P, Lacina L, Gal P, Uhrova J, Zima T, Kaltner H, André S, Gabius HJ, Sykova E, Smetana K., Jr Human galectins induce conversion of dermal fibroblasts into myofibroblasts and production of extracellular matrix: Potential application in tissue engineering and wound repair. Cells Tissues Organs. 2011;194:469–480. doi: 10.1159/000324864. [DOI] [PubMed] [Google Scholar]
- 12.Krasagakis K, Garbe C, Schrier PI, Orfanos CE. Paracrine and autocrine regulation of human melanocyte and melanoma cell growth by transforming growth factor beta in vitro. Anticancer Res. 1994;14:2565–2571. [PubMed] [Google Scholar]
- 13.Balch CM, Gershenwald JE, Soong SJ, Thompson JF. Update on the melanoma staging system: The importance of sentinel node staging and primary tumor mitotic rate. J Surg Oncol. 2011;104:379–385. doi: 10.1002/jso.21876. [DOI] [PubMed] [Google Scholar]
- 14.Lacina L, Smetana K, Jr, Dvoránková B, Pytlík R, Kideryová L, Kucerová L, Plzáková Z, Stork J, Gabius HJ, André S. Stromal fibroblasts from basal cell carcinoma affect phenotype of normal keratinocytes. Br J Dermatol. 2007;156:819–829. doi: 10.1111/j.1365-2133.2006.07728.x. [DOI] [PubMed] [Google Scholar]
- 15.Kolář M, Szabo P, Dvořánková B, Lacina L, Gabius HJ, Strnad H, Sáchová J, Vlček C, Plzák J, Chovanec M, et al. Upregulation of IL-6, IL-8 and CXCL-1 production in dermal fibroblasts by normal/malignant epithelial cells in vitro: Immunohistochemical and transcriptomic analyses. Biol Cell. 2012;104:738–751. doi: 10.1111/boc.201200018. [DOI] [PubMed] [Google Scholar]
- 16.Therasse P, Arbuck SG, Eisenhauer EA, Wanders J, Kaplan RS, Rubinstein L, Verweij J, Van Glabbeke M, van Oosterom AT, Christian MC, Gwyther SG. New guidelines to evaluate the response to treatment in solid tumors European organization for research and treatment of cancer, National Cancer institute of the United States, National Cancer Institute of Canada. J Natl Cancer Inst. 2000;92:205–216. doi: 10.1093/jnci/92.3.205. [DOI] [PubMed] [Google Scholar]
- 17.Salvatore G, Chiappetta G, Nikiforov YE, Decaussin-Petrucci M, Fusco A, Carney JA, Santoro M. Molecular profile of hyalinizing trabecular tumours of the thyroid: High prevalence of RET/PTC rearrangements and absence of B-raf and N-ras point mutations. Eur J Cancer. 2005;41:816–821. doi: 10.1016/j.ejca.2005.01.004. [DOI] [PubMed] [Google Scholar]
- 18.Sykorova V, Dvorakova S, Ryska A, Vcelak J, Vaclavikova E, Laco J, Kodetova D, Kodet R, Cibula A, Duskova J, et al. BRAFV600E mutation in the pathogenesis of a large series of papillary thyroid carcinoma in Czech Republic. J Endocrinol Invest. 2010;33:318–324. doi: 10.1007/BF03346593. [DOI] [PubMed] [Google Scholar]
- 19.Smyth GK. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat Appl Genet Mol Biol. 2004;3 doi: 10.2202/1544-6115.1027. Article 3. [DOI] [PubMed] [Google Scholar]
- 20.Gentleman RC, Carey VJ, Bates DM, Bolstad B, Dettling M, Dudoit S, Ellis B, Gautier L, Ge Y, Gentry J, et al. Bioconductor: Open software development for computational biology and bioinformatics. Genome Biol. 2004;5:R80. doi: 10.1186/gb-2004-5-10-r80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Valach J, Fík Z, Strnad H, Chovanec M, Plzák J, Cada Z, Szabo P, Sáchová J, Hroudová M, Urbanová M, et al. Smooth muscle actin-expressing stromal fibroblasts in head and neck squamous cell carcinoma: Increased expression of galectin-1 and induction of poor prognosis factors. Int J Cancer. 2012;131:2499–2508. doi: 10.1002/ijc.27550. [DOI] [PubMed] [Google Scholar]
- 22.Culhane AC, Thioulouse J, Perrière G, Higgins DG. MADE4: An R package for multivariate analysis of gene expression data. Bioinformatics. 2005;21:2789–2790. doi: 10.1093/bioinformatics/bti394. [DOI] [PubMed] [Google Scholar]
- 23.Wu H, Caffo B, Jaffee HA, Irizarry RA, Feinberg AP. Redefining CpG islands using hidden Markov models. Biostatistics. 2010;11:499–514. doi: 10.1093/biostatistics/kxq005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Chen EY, Tan CM, Kou Y, Duan Q, Wang Z, Meirelles GV, Clark NR, Ma'ayan A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinformatics. 2013;14:128. doi: 10.1186/1471-2105-14-128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Driskell RR, Watt FM. Understanding fibroblast heterogeneity in the skin. Trends Cell Biol. 2015;25:92–99. doi: 10.1016/j.tcb.2014.10.001. [DOI] [PubMed] [Google Scholar]
- 26.Tomasek JJ, GAbbiani G, Hinz B, Chaponnier C, Brown RA. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat Rev Mol Cell Biol. 2002;3:349–363. doi: 10.1038/nrm809. [DOI] [PubMed] [Google Scholar]
- 27.Whipple CA, Brinckerhoff CE. BRAF(V600E) melanoma cells secrete factors that activate stromal fibroblasts and enhance tumourigenicity. Br J Cancer. 2014;111:1625–1633. doi: 10.1038/bjc.2014.452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Fedorenko IV, Wargo JA, Flaherty KT, Messina JL, Smalley KSM. BRAF inhibition generates a host-tumor niche that mediates therapeutic escape. J Invest Dermatol. 2015;135:3115–3124. doi: 10.1038/jid.2015.329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hirata E, Girotti MR, Viros A, Hooper S, Spencer-Dene B, Matsuda M, Larkin J, Marais R, Sahai E. Intravital imaging reveals how BRAF inhibition generates drug-tolerant microenvi-ronments with high integrin β1/FAK signaling. Cancer Cell. 2015;27:574–588. doi: 10.1016/j.ccell.2015.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Fedorenko IV, Smalley KS. The complexity of microenvironment-mediated drug resistance. Genes Cancer. 2015;6:367–368. doi: 10.18632/genesandcancer.82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.De Wever O, Hendrix A, De Boeck A, Eertmans F, Westbroek W, Braems G, Bracke ME. Single cell and spheroid collagen type I invasion assay. Methods Mol Biol. 2014;1070:13–35. doi: 10.1007/978-1-4614-8244-4_2. [DOI] [PubMed] [Google Scholar]
- 32.Varley KE, Gertz J, Bowling KM, Parker SL, Reddy TE, Pauli-Behn F, Cross MK, Williams BA, Stamatoyannopoulos JA, Crawford GE, et al. Dynamic DNA methylation across diverse human cell lines and tissues. Genome Res. 2013;23:555–567. doi: 10.1101/gr.147942.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Drabsch Y, ten Dijke P. TGF-β signalling and its role in cancer progression and metastasis. Cancer Metastasis Rev. 2012;31:553–568. doi: 10.1007/s10555-012-9375-7. [DOI] [PubMed] [Google Scholar]
- 34.Guo L, Kuroda N, Nakayama H, Miyazaki E, Hayashi Y, Toi M, Hiroi M, Enzan H. Cytokeratin positive subserosal positive subserosal myofibroblasts in gastroduodenal ulcer; another type of myofibroblasts. Histol Histopathol. 2006;21:697–704. doi: 10.14670/HH-21.697. [DOI] [PubMed] [Google Scholar]
- 35.Seip K, Fleten KG, Barkovskaya A, Nygaard V, Haugen MH, Engesæter BØ, Mælandsmo GM, Prasmickaite L. Fibroblast-induced switching to the mesenchymal-like phenotype and PI3K/mTOR signaling protects melanoma cells from BRAF inhibitors. Oncotarget. 2016;7:19997–20015. doi: 10.18632/oncotarget.7671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Johnson DB, Menzies AM, Zimmer L, Eroglu Z, Ye F, Zhao S, Rizos H, Sucker A, Scolyer RA, Gutzmer R, et al. Acquired BRAF inhibitor resistance: A multicenter meta-analysis of the spectrum and frequencies, clinical behaviour, and phenotypic associations of resistance mechanisms. Eur J Cancer. 2015;51:2792–2799. doi: 10.1016/j.ejca.2015.08.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Llopiz D, Dotor J, Casares N, Bezunartea J, Díaz-Valdés N, Ruiz M, Aranda F, Berraondo P, Prieto J, Lasarte JJ, et al. Peptide inhibitors of transforming growth factor-beta enhance the effi-cacy of antitumor immunotherapy. Int J Cancer. 2009;125:2614–2623. doi: 10.1002/ijc.24656. [DOI] [PubMed] [Google Scholar]
- 38.Morris JC, Tan AR, Olencki TE, Shapiro GI, Dezube BJ, Reiss M, Hsu FJ, Berzofsky JA, Lawrence DP. Phase I study of GC1008 (fresolimumab): A human anti-transforming growth factor-beta (TGFβ) monoclonal antibody in patients with advanced malignant melanoma or renal cell carcinoma. PLoS One. 2014;9:90353. doi: 10.1371/journal.pone.0090353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mifková A, Kodet O, Szabo P, Kučera J, Dvořánková B, André S, Koripelly G, Gabius HJ, Lehn JM, Smetana K., Jr Synthetic polyamine BPA-C8 inhibits TGF-β1-mediated conversion of human dermal fibroblast to myofibroblasts and establishment of galectin-1-rich extracellular matrix in vitro. Chembiochem. 2014;15:1465–1470. doi: 10.1002/cbic.201402087. [DOI] [PubMed] [Google Scholar]
- 40.Jobe NP, Rösel D, Dvořánková B, Kodet O, Lacina L, Mateu R, Smetana K, Brábek J. Simultaneous blocking of IL-6 and IL-8 is sufficient to fully inhibit CAF-induced human melanoma cell invasiveness. Histochem Cell Biol. 2016;146:205–217. doi: 10.1007/s00418-016-1433-8. [DOI] [PubMed] [Google Scholar]
- 41.Harbst K, Lauss M, Cirenajwis H, Winter C, Howlin J, Törngren T, Kvist A, Nodin B, Olsson E, Häkkinen J, et al. Molecular and genetic diversity in the metastatic process of melanoma. J Pathol. 2014;233:39–50. doi: 10.1002/path.4318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Patrick E, Schramm SJ, Ormerod JT, Scolyer RA, Mann GJ, Mueller S, Yang JY. A multi-step classifier addressing cohort heterogeneity improves performance of prognostic biomarkers in three cancer types. Oncotarget. 2017;8:2807–2815. doi: 10.18632/oncotarget.13203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Maley CC, Aktipis A, Graham TA, Sottoriva A, Boddy AM, Janiszewska M, Silva AS, Gerlinger M, Yuan Y, Pienta KJ, et al. Classifying the evolutionary and ecological features of neoplasms. Nat Rev Cancer. 2017;17:605–619. doi: 10.1038/nrc.2017.69. [DOI] [PMC free article] [PubMed] [Google Scholar]