

Abstract
The aim of this work has been to assess the effect of diet density [control (CON) or 15% diluted (DIL)] and growth curve [recommended by the genetic line (RBW) or 15% heavier (HBW)] and their interaction on BW uniformity, feeding motivation, behavior, and body composition of broiler breeder pullets. A total of 3,000 one-day-old female breeders Ross 308, distributed in 20 pens, was randomly assigned to each treatment. Feed allowance was weekly adjusted to reach the desired BW. Feed was provided as pelleted (zero to 3 wk) and crumble (4 to 19 wk). Time eating was measured at 7, 11, and 19 weeks. A feeding rate test was performed after 11 weeks. Behavior was observed at 9 and 15 wk, by visual scan. At 6, 13, and 19 wk of age, one bird/pen was slaughtered for weighing different organs and analyzing the composition of empty whole bodies. Treatments did not affect BW uniformity; relative weights of the ovary, oviduct, or gizzard; or protein content of empty BW. Time eating varied with the growth curve at 19 wk (P < 0.05), HBW pullets spent 19 more min eating than RBW pullets. DIL led to 4 and 8 more min eating at 19 wk for pullets of RBW and HBW (P < 0.05), respectively. Pullets fed DIL consumed 30% (P < 0.05) less during the feeding rate test when kept on a restricted regimen, and they had lower compensatory energy intake after ad libitum feeding than those fed CON, indicating lower feeding motivation. Behavior was affected by the age and by the time of the d measured, but it did not change with the treatments. Birds spent most time pecking objects (50%), feeding (28%), and drinking (17%). Pullets fed DIL had 8% lower breast yield at different ages and higher empty digestive tracts at 6 weeks. Body composition varied with age; fat content increased from 12.7 to 15.9 to 19.8% for 6, 13, and 19 wk, respectively. The lowest body fat was observed for RBW pullets fed DIL (P = 0.003) at 19 weeks. Feeding DIL diets to HBW pullets could be done to increase the time spent eating and reduce their feeling of hunger without negative effects on body composition. However, its influence on behavior and BW uniformity was not proved.
Keywords: breeder pullet, diet density, growth curve, body composition, eating motivation
INTRODUCTION
In broiler breeders feed intake is restricted due to the inability of these birds to adequately self-regulate feed intake during growth and development to achieve the optimal body weight and body composition needed to support efficient hatching-egg production (Richards, 2003). Female broiler breeders that are fed ad libitum decrease their total egg output and number of chicks hatched per breeder hen (Yu et al., 1992; Robinson et al., 1993). In contrast, very severe feed restriction can result in poor egg production and hatchability as well (Wilson and Harms, 1986).
Broiler genetics companies are continually improving the body weight gain and meat yield of their final products (Havenstein et al., 2003a,b). These improvements have negative side effects on reproduction traits (Rauw et al., 1998) that try to be solved by an increased feed restriction of the broiler breeders. Feed restriction during the rearing phase, estimated at between 1/3 and 1/2 of ad libitum broiler breeder intake at a similar age or BW (De Jong et al., 2002), is a focal point of concerns in terms of broiler breeder welfare and later performance. The feeding program of broiler breeders most commonly applied has the highest degree of feed restriction from 8 to 16 wk of age (Hocking, 1993). Due to the growth, metabolic, reproductive, and health problems that are alleviated under feed restriction conditions, feed intake patterns and body weight growth curves provided by the breeder companies are considered the most welfare-friendly alternative to ad libitum feeding (Renema and Robinson, 2004). However, restriction may result in considerable feeling of hunger that could be negatively appreciated by public opinion. Under commercial restrictions, it is possible to observe behaviors associated with frustration, such as nonfood pecking activities, which sometimes can lead to feather pecking and aggression (Mench, 2002). The potential to reduce the degree of feed restriction and hunger through diet dilution (Hocking, 2004; De Jong et al., 2005), relaxed feed restriction (Hocking et al., 2002; Bruggeman et al., 2005), and the introduction of alternative genetic stocks (Heck et al., 2004; Bruggeman et al., 2005) has been assessed with variable results. It was hypothesized that dilution of the diets and allowing pullets to reach higher body weights led to a less severe restriction that might reduce the feeling of hunger and its negative welfare impact.
The aim of the current study has been to assess the effect of diet density (dilution) and growth curve (target body weight) and their interaction on body weight uniformity, eating time, feeding motivation, behavior, and body composition of broiler breeders in the rearing phase.
MATERIALS AND METHODS
Birds, Husbandry, and Feeding Program
All experimental procedures were approved by the animal Ethics Committee of the Trouw Nutrition Poultry Research Center, in compliance with the Spanish guidelines for the care and use of animals in research (Boletín Oficial del Estado, 2013).
There were 3,000 one-day-old female broiler breeders Ross 308 (Aviagen Spain S.A., Riells del Fai, Spain) randomly distributed in 20 pens (150 chicks/pen) in the broiler breeder facility of the Trouw Nutrition Poultry Research Center (El Viso de San Juan, Toledo, Spain). Each pen had an area of 30 m2 (6 × 5 m) and contained a squared automatic chain feeder (20 linear m, 10 m x 2 sides), 20 nipple drinkers, and 24 individual nests placed in 2 levels that were kept closed throughout the trial. Wood shavings were utilized as litter material. When pullets were 11 wk old, plastic slats were placed in front of the closed nests. The slat area covered a surface of 4.8 m2 (1.2 × 4 m) and was 60 cm height above floor.
Animals were beak trimmed and vaccinated at the hatchery against Marek disease and infectious bronchitis. A standard vaccination program adapted to the area of production was followed during the rearing period.
The lighting program was 24 h of light during the first 2 d and then, gradually reduced to 8 h until d 10. From d 10 to the end of the trial (19 wk), the lighting program was 8 h of light (from 0700 to 1500 h). Light intensity at the animal level was set at 20 lux. The temperature during the first 2 d was maintained at 33 °C, and then gradually reduced to reach 20 °C at 28 d and maintained at 20 °C thereafter. Water was provided ad libitum throughout the trial.
Experimental Design
The study was conducted as a 2 × 2 factorial completely randomized design with 2 nutrient densities: control (CON) and 15% diluted (DIL), which contained 15% lower nutritional value (AMEn, crude protein and amino acids, digestible phosphorous, and calcium) and 2 target body weights: recommended (RBW; Aviagen, 2012) and 15% heavier (HBW) from 3 wk onwards. Target BW at 19 wk was set at 2,185 and 2,513 g, for RBW and HBW treatments, respectively. Feed allowance to achieve the different target weights was weekly calculated based on the energy requirements provided by the breeding company (RBW) or 15% higher (HBW) and the AMEn content of the tested diets (CON and DIL).
Feed was provided ad libitum during the first 2 wk in order to ensure proper growth at the beginning. During the third wk, feed intake was set to obtain the differences in BW determined by each treatment. From 4 to 19 wk of age, animals were fed restricted, according to the treatment assigned. A 4-phase feeding program was followed: starter-1 (from zero to 3 wk of age), starter-2 (from 4 to 7 wk), grower-1 (from 8 to 16 wk), and grower-2 (until the end of the study). Pelleted feed (2 mm) was provided from zero to 3 wk of age and in crumbles (3.5 mm) afterwards. Raw material and nutritional composition of diets are shown in Table 1.
Table 1.
Ingredient and nutritional (%) composition of the experimental diets.
| Starter-2 | Grower-1 | Grower-2 | |||||
|---|---|---|---|---|---|---|---|
| Raw materials | Starter1 | Control | Diluted | Control | Diluted | Control | Diluted |
| Corn | 30.0 | 25.0 | 20.0 | 20.0 | 18.8 | 28.0 | 30.8 |
| Wheat | 25.8 | 18.8 | 3.2 | 21.3 | – | 29.1 | 2.0 |
| Barley | – | 15.0 | 15.0 | 15.0 | 12.2 | 10.0 | 18.0 |
| Oat | – | 10.0 | 15.0 | 10.0 | 10.0 | – | – |
| Wheat bran | 5.0 | – | 17.3 | 10.0 | 27.0 | 7.7 | 21.0 |
| Gluten feed | – | – | – | – | 10.0 | – | – |
| Sunflower meal | 4.7 | 8.5 | 10.0 | 9.0 | 11.0 | 10.0 | 11.9 |
| Soybean meal | 25.3 | 15.5 | 5.6 | 8.2 | – | – | – |
| Rapeseed | – | – | – | – | – | 7.7 | – |
| Sunflower seeds | 2.3 | – | – | – | – | – | – |
| Soybean hulls | – | – | 5.0 | – | – | – | – |
| Oat hulls | – | – | – | – | – | – | 4.0 |
| Cereal Straw | – | 1.8 | 2.0 | 2.0 | 5.0 | 3.0 | 6.0 |
| Alfalfa | – | 2.0 | – | 1.2 | – | – | |
| Blend Fat | 2.0 | – | – | – | – | – | – |
| Soya oil | – | 1.5 | 1.5 | – | – | 1.0 | 1.0 |
| Sodium bicarbonate | 0.23 | 0.17 | 0.25 | 0.11 | 0.17 | 0.15 | 0.18 |
| Calcium Carbonate | 1.61 | 1.59 | 1.23 | 1.16 | 1.14 | 1.40 | 1.52 |
| Monocalcium phosphate | 1.21 | 1.01 | 0.66 | 1.10 | 0.64 | 0.92 | 0.68 |
| Sepiolite | 0.84 | – | – | 1.25 | 2.00 | – | 2.00 |
| Salt | 0.21 | 0.26 | 0.14 | 0.26 | 0.11 | 0.27 | 0.19 |
| L-Lysine | 0.06 | 0.17 | 0.28 | 0.07 | 0.16 | 0.19 | 0.21 |
| DL-Methionine | 0.15 | 0.17 | 0.16 | 0.04 | 0.02 | – | 0.02 |
| L-Threonine | – | 0.06 | 0.15 | – | 0.02 | 0.02 | 0.04 |
| Premix | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 |
| Calculated values, % | |||||||
| AMEn Kcal/kg | 2800 | 2700 | 2295 | 2550 | 2170 | 2650 | 2253 |
| dig Lys | 0.89 | 0.75 | 0.64 | 0.55 | 0.47 | 0.55 | 0.47 |
| dig Met | 0.42 | 0.36 | 0.31 | 0.23 | 0.20 | 0.23 | 0.21 |
| dig M+C | 0.70 | 0.60 | 0.51 | 0.46 | 0.39 | 0.47 | 0.39 |
| dig Thre | 0.60 | 0.51 | 0.43 | 0.41 | 0.35 | 0.41 | 0.35 |
| dig Trp | 0.21 | 0.17 | 0.13 | 0.15 | 0.12 | 0.14 | 0.11 |
| Crude protein | 20.0 | 17.0 | 14.5 | 15.0 | 12.8 | 15.0 | 12.8 |
| Crude fiber | 4.2 | 6.6 | 10.8 | 7.0 | 9.6 | 7.1 | 10.1 |
| Crude fat | 4.9 | 3.8 | 4.0 | 2.4 | 3.1 | 3.2 | 3.4 |
| Ash | 7.2 | 6.0 | 5.9 | 7.0 | 8.1 | 5.4 | 7.7 |
| Calcium | 1.00 | 0.90 | 0.77 | 0.80 | 0.77 | 0.80 | 0.77 |
| dig Phosphorous | 0.40 | 0.34 | 0.29 | 0.38 | 0.31 | 0.32 | 0.27 |
| Analyzed values, % | |||||||
| DM | 89.4 | 89.5 | 89.6 | 89.3 | 89.9 | 92.8 | 93.3 |
| Crude protein | 20.8 | 16.8 | 14.2 | 14.7 | 12.4 | 14.9 | 13.0 |
| Crude fiber | 4.6 | 7.1 | 10.5 | 7.4 | 10.0 | 6.8 | 10.2 |
| Crude fat | 3.5 | 3.6 | 4.0 | 2.2 | 2.6 | 3.1 | 3.0 |
| Ash | 7.5 | 5.9 | 5.8 | 6.1 | 7.4 | 5.3 | 7.9 |
| Starch | – | 41.2 | 33.1 | 41.5 | 35.0 | 44.3 | 38.0 |
Determinations
Feed Intake, Body Weight, Uniformity, and Mortality.
Feed per pen was daily weighed and supplied. Feed allowance was adjusted to obtain the defined growth curves (RBW or HBW) and calculated per week. Body weight was weekly recorded per pen (except for 16 wk of age). Thus, a defined number of animals within a range (20 to 30 as reference) were individually weighed per pen to calculate the weekly body weight. Those animals were randomly caught by using catching frames, and all birds rounded up were weighed in order to eliminate any selective bias. At 19 wk of age, all animals per pen were individually weighed. Body weight uniformity was calculated at 19 wk of age, as the percentage of animals within ± 10% the average BW, and as the coefficient of variation of the BW of each pen. Mortality was daily recorded and reported as accumulated mortality till 19 wk of age.
Time Eating.
At 7, 11, and 19 wk of age, the time required (min/d) for consuming total supplied feed was recorded. Time eating was defined as the time difference between feeder filling and emptying.
Feeding Rate Test.
The principle behind the test is that the level of hunger is reflected in the speed of eating. The feeding rate was determined according to Nielsen et al. (2011), between 74 and 87 d of age. At 74 d of age, 2 birds were removed from each pen at 1300 h, weighed, and placed in different floor pens, keeping the feeding regimen during one wk (adaptation to the new pen characteristics). At 81 d of age, the feeder was filled with a known amount of the corresponding diet at 0730 h, as always, allowing birds to eat for 2 min, and, afterwards, remaining feed and birds were weighed. The difference of feed weight was registered as feeding rate at restriction. Subsequently, the birds had ad libitum access to the feed, with feed intake estimated by weighing the feeder after 24 and 48 h (d 82 and 83 of age) considered compensatory intake, at which points the birds also were weighed. Compensatory intake was calculated as the daily energy consumed per metabolic body weight after 24 h (first d) and 48 h (second d) of ad libitum consumption. After a total period of 5 d of ad libitum feeding, the feed was weighed and the feeders elevated (86 d of age). Following 24 h of feed withdrawal, birds were re-tested by giving access to the feed for 2 min (87 d of age), being registered as the feeding rate in ad libitum conditions.
Behavior.
Behavior was measured adapting the procedure described by De Jong et al. (2005). At 9 and 15 wk of age, behavior of the birds was observed by visual scan (2 people) following the ethogram shown in Table 2. The observers were previously trained for this. For each pen of 150 birds, behavior was scored by counting the number of birds engaged in different activities in a quarter of the pen, which was established in advance and kept fixed during the entire study. This area included a quarter of the feeders and drinkers and included all the elements of a pen, such as slat, walls, and closed nests. Behavior was observed 8 times around feeding from 0730 to 1030 h and another 8 times after feeding from 1130 to 1430 h on 2 consecutive days. For each time period, the total number of animals in the quarter also was recorded to calculate later the percentage of animals engaged in the different activities.
Table 2.
Ethogram of the behavioral measurements.
| Behavior | Description |
|---|---|
| Walking | Walking or running with no other discernable activity |
| Drinking | Drinking or releasing water from the nipple drinkers |
| Feeding | Feeding from the feeder |
| Pecking objects | Pecking at objects in the pen, including the empty feeder |
| Pecking other birds | Pecking birds, including the tail feathers or tail region of other birds |
| Foraging | Pecking, scratching the litter |
| Comfort | All other comfort behaviors, such as preening, scratching or wing stretching, sitting, standing and sand bathing |
Body Composition.
At 6, 13, and 19 wk of age, one bird per pen was selected within a BW range from 95 to 105% of the average BW per pen in that week. Pullets were slaughtered by cervical dislocation and weighed. The gastrointestinal tract of each pullet was emptied and weights of the empty gastrointestinal tract, empty gizzard, breast meat, oviduct, and ovary (at 19 wk only) were recorded. Weights of each part of the bird were expressed as percentage of BW, except for the oviduct and the ovary, which were reported as g/ 100 kg BW, due to their low values. After weighing, organs, tissues, and the empty body of each animal were frozen together for posterior analysis. Whole bodies without gastrointestinal content but including the feathers and blood were ground with a blade cutter (Cruells Cutter 157375, tri-phase motor at 220 V with 15 L rotator plate at 18 rpm and rotator knives at 2600 rpm). A representative sample of each pullet was collected and dry matter (DM), ash, crude protein (CP), and ether extract (EE) determined. Energy and protein efficiencies were calculated as the ratio of energy and protein retained (considering the body weight and its composition at 19 wk of age) and the AMEn and CP ingested during rearing. A caloric value of body fat and protein of 9.385 and 5.686 kcal/g, respectively, was used for energy efficiency estimations (Sakomura et al., 2003).
Chemical Analyses
Feeds and pullet body samples were analyzed according to AOAC International 18th edition (2005) for moisture by oven-drying (934.01), total ash (942.05), nitrogen by combustion (990.03) using a LECO analyzer, and ether extract (960.39). Feeds also were analyzed for starch by the α-amylase glucosidase method (996.11), crude fiber by sequential extraction with diluted acid and alkali (962.09), and specific weight (985.19).
Statistical Analysis
The experimental design was completely randomized with 4 treatments arranged in a factorial array (2 diet densities x 2 growth curves). Main effects (diet density and growth curve) and the interaction were analyzed by using the GLM and MIXED procedures of SAS® 9.3. (SAS Institute Inc., 2012). Behavior observations were analyzed considering a binomial distribution using GLIMMIX. These results were log transformed. Body weight, diet, and the moment of the d (morning or afternoon) were analyzed as fixed effects, with age as a repetitive measure and pen as a random effect. The variance homogeneity of the data for all traits was tested by the Levene´s Test (Hovtest option of the GLM procedure).
RESULTS
Feed Intake, Body Weight, Uniformity, and Mortality
Feed intake and body weight between zero and 19 wk of age are shown in Figure 1 (A and B respectively). To meet target BW of each treatment, feed supply was increased in CON-HBW, DIL-RBW, and DIL-HBW breeder pullets by 15, 14, and 28%, respectively, compared to birds in the control (CON-RBW) treatment. Accumulated feed intake per breeder pullet was on average 7,258, 8,343, 8,300, and 9,306 g for treatments CON-RBW, CON-HBW, DIL-RBW, and DIL-HBW, respectively. After weekly adjustment of feed supply to pullet requirements of each treatment, feed intake was 8.3% lower, on average for all treatments, than the calculated before starting the trial. However, the target weights were achieved. At the end of the trial, HBW breeder pullets had, on average, 14% higher BW compared to the RBW treatments (2,449 vs. 2,151 g; P < 0.0001). Breeder females eating diluted diet had 1.6% lower body weight (P = 0.039) than the ones fed the control diet (2,281 vs. 2,318 g).
Figure 1.

Weekly average feed intake (A) and body weight (B) of broiler breeder pullets from one to 19 wk of age per treatment (means of 5 pens per treatment ± SEM). Treatments are identified in the graphs as CON/RBW: control diet and recommended growing curve; CON/HBW: control diet and reared to reach a 15% heavier body weight compared to the recommended; DIL/RBW: pullets fed 15% diluted diet and recommended growing curve; DIL/HBW: 15% diluted diet and 15% heavier body weight. In figure 1 (B), the pointed lines show the target body weight for the treatments with the RBW () and HBW ().
As shown in Table 3, body weight coefficient of variation (BW cv), uniformity, and accumulated mortality at 19 wk were not affected by any of the effects studied (P > 0.1) and averaged 13.6, 53.7, and 5.3%, respectively. Nevertheless, breeder pullets fed diluted diets had lower BW cv and higher uniformity compared to the animals eating the control diet (13.1 and 14.2, P = 0.107; and 56.0 and 51.6%, P = 0.113, respectively).
Table 3.
Effect of diet density and growing curve and their interaction on body weight, body weight coefficient of variation (cv), and body weight uniformity (%) at 19 wk of age and the accumulated mortality from 0 to 19 wk of age.
| Effects | BW, g | BW cv | BW uniformity, | Ac. mortality, |
|---|---|---|---|---|
| % | % | |||
| Diet density | ||||
| CON | 2318 | 14.2 | 51.6 | 6.0 |
| DIL | 2281 | 13.1 | 56.0 | 4.6 |
| SEM (n = 10) | 12 | 0.5 | 1.9 | 0.9 |
| Target body weight | ||||
| RBW | 2151 | 14.1 | 53.1 | 4.7 |
| HBW | 2449 | 13.2 | 54.4 | 5.8 |
| SEM (n = 10) | 12 | 0.5 | 1.9 | 0.9 |
| Treatments | ||||
| CON / REC | 2161 | 14.6 | 51.2 | 6.3 |
| CON / HEAVY | 2475 | 13.7 | 51.9 | 5.6 |
| DIL / REC | 2140 | 13.6 | 55.0 | 3.1 |
| DIL / HEAVY | 2422 | 12.6 | 56.9 | 6.0 |
| SEM (n = 5) | 16 | 0.6 | 2.6 | 1.3 |
| P-values | ||||
| Diet | 0.039 | 0.107 | 0.113 | 0.299 |
| Target body weight | <.0001 | 0.174 | 0.624 | 0.407 |
| Interaction | 0.353 | 0.872 | 0.809 | 0.192 |
Time Eating
Time eating varied along the trial (Table 4) from 85 and 75 to 102 min a d, on average, for breeder pullets of 7, 11, and 19 wk of age, respectively. At 7 and 11 wk of age, no differences between treatments were observed. However, at 19 wk of age, breeder pullets reared to obtain HBW spent 17% more time eating compared to those with RBW (P = 0.008; 92.6 vs. 111.3 min). An interaction showed that diet dilution led to 4 and 8 more min eating for pullets of RBW and HBW, respectively.
Table 4.
Time eating (min) and eating rate (g feed/min) in broiler breeder pullets at 7, 11, and 19 wk of age.
| 7 wk | 11 wk | 19 wk | ||||
|---|---|---|---|---|---|---|
| Effects | Eating time | Eating rate | Eating time | Eating rate | Eating time | Eating rate |
| Diet density | ||||||
| CON | 83.7 | 0.64 | 73.6 | 0.90 | 98.9 | 0.89 |
| DIL | 86.0 | 0.72 | 76.9 | 0.98 | 105.0 | 0.95 |
| SEM (n = 10) | 4.6 | 0.04 | 5.3 | 0.07 | 4.7 | 0.05 |
| Target body weight | ||||||
| RBW | 81.8 | 0.65 | 70.3 | 0.94 | 92.6 | 0.95 |
| HBW | 87.9 | 0.70 | 80.2 | 0.94 | 111.3 | 0.88 |
| SEM (n = 10) | 4.6 | 0.04 | 5.3 | 0.07 | 4.7 | 0.05 |
| Treatments | ||||||
| CON / RBW | 81.0 | 0.61 | 69.2 | 0.88 | 90.6 | 0.91 |
| CON / HBW | 86.4 | 0.67 | 78.0 | 0.91 | 107.2 | 0.86 |
| DIL / RBW | 82.6 | 0.70 | 71.4 | 1.00 | 94.6 | 0.99 |
| DIL / HBW | 89.4 | 0.73 | 82.4 | 0.96 | 115.4 | 0.91 |
| SEM (n = 5) | 6.5 | 0.06 | 7.5 | 0.10 | 6.6 | 0.06 |
| P-values | ||||||
| Diet | 0.729 | 0.174 | 0.376 | 0.414 | 0.368 | 0.322 |
| Target BW | 0.364 | 0.415 | 0.341 | 0.974 | 0.008 | 0.313 |
| Interaction | 0.810 | 0.841 | 0.418 | 0.721 | 0.050 | 0.812 |
Feeding Rate Test
The feeding rate test (Figure 2) showed significant effect of diet dilution (P < 0.0001) but not any statistical effect of target weight. Figure 2B shows the effect of diet dilution on feed intake during 2 min after 24-hour feed withdrawal in birds on restricted feeding and the same birds after 5 days of ad libitum feeding. Breeder pullets on restricted feeding consumed 71% more feed during the first 2 min of feeding than those pullets after 5 d of ad libitum regime (26.1 vs. 15.3 g, on average, respectively). Breeders eating the diluted diet consumed 30% (P < 0.05) less than those fed the control diet during their first access to the feed when kept in restricted regimen. After 5 d of ad libitum feeding, birds eating the diluted diet ate 24% less amount of feed than those fed the control diet, but this difference is not statistically different. The compensatory AME intake consumed after 24 and 48 h of ad libitum feeding resulted in 14% lower, on average, for animals fed the DIL than the CON diet, but this effect was clearer the first 24 h (16%) than from 24 to 48 h (12%) of ad libitum feeding (Figure 2B).
Figure 2.
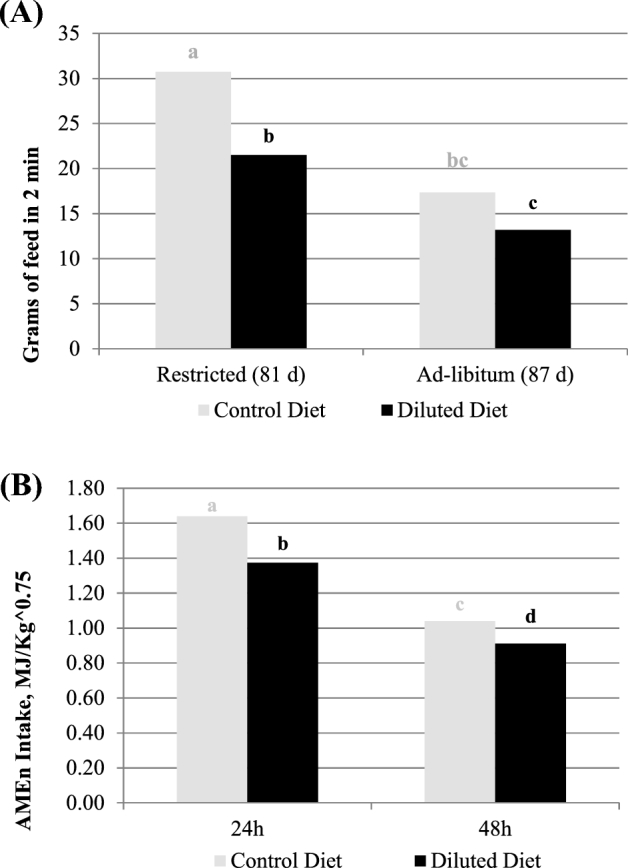
Feeding rate test (g of feed consumed in 2 min) after feed withdrawal of 24 h in pullets subjected to daily feed restriction (81 d) or ad libitum feeding (87 d) (A) and compensatory energy intake (MJ/kg 0.75) 24 h (first d) and 48 h (secong d) after ad libitum feed supplied (B) between the different diets (control vs. diluted).
Behavior
No significant interactions among the different effects studied have been observed, so they were removed from the statistical analysis. Figure 3 shows the percentage of birds of each treatment observed in the different behavior activities (A) and those activities that were differently affected by the age of the breeder pullet (B) or the time of the d (C). Among treatments, no statistical differences were obtained. Birds spent most time pecking objects, feeding, drinking, and walking, based on the percentage of animals performing these activities (49.9, 27.6, 17.6, and 12.6% on average, respectively), whereas for activities related to comfort, pecking other birds and foraging amounted to less time (2 to 5% of birds performing these activities). The age of the animals affected (P < 0.05) some of these activities. At 7 wk of age, more animals were observed walking (13.4 vs. 11.8%), drinking (18.4 vs. 16.7%), and foraging (3.2 vs 1.0%), whereas less were pecking other birds (0.9 vs. 8.1%) and performing comfort activities than those at 15 wk of age. In the same way, some activities differed depending on the time of day. Thus, feeding was exclusively performed during the morning, and more birds were drinking in the morning than in the evening (20.0 vs. 15.4%), whereas pecking objects and comfort activities were mainly performed in the evening (26.9 vs. 58.0% and 4.4 vs 7.4%, respectively).
Figure 3.

Percentage of animals (%) performing different activities per treatment (A), per wk of age (7 and 15 wk of age) (B), and per time of the d (morning: with feed in the feeder; afternoon: without feed) (C).
Body Composition.
The effects of diet density and growth curve on some muscles and organ weights at different ages are shown in Table 5. Breast yield increased from 10.4 to 19.1% at 6 and 19 wk of age, on average, respectively, whereas the digestive tract decreased from 6.8 to 3.1%. Breast yield was 8% higher, on average, in breeder pullets fed the control diet than those fed the diluted diet at every age (P < 0.10). At 13 wk of age, pullets fed diluted diets had lighter breast yield when they were grown according to the recommended target weight (P = 0.0027) than 15% over it. Empty digestive tract yield was affected only at 6 wk of age by diet dilution, in such a way that pullets fed the DIL diet yielded 15% higher digestive tract than those fed CON ones. But, the effect of diet dilution was more pronounced in the digestive tract yield of pullets following HBW (19% increase) than RBW (11% increase). However, relative gizzard weight was not affected by treatments, but it was reduced with age from 3.4% at 6 wk to 2.0% at 19 wk of age, on average. The relative weight of the oviduct and ovaries rendered 16.5 and 31.7 g/100 kg, respectively, at 19 wk of age, on average, and they did not respond differently among treatments.
Table 5.
Effects of diet density and growing curve (target body weight) and their interaction on breast meat yield, digestive tract, and gizzard (% BW) at 6, 13, and 19 wk of age and oviduct and ovary (g/100 kg BW) at 19 wk of age.
| Breast meat yield | Empty digestive tract | Gizzard | Oviduct | Ovary | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Effects | 6 wk | 13 wk | 19 wk | 6 wk | 13 wk | 19 wk | 6 wk | 13 wk | 19 wk | 19 wk | 19 wk |
| Diet density | |||||||||||
| CON | 10.9 | 15.2 | 19.6 | 6.37 | 3.26 | 3.04 | 3.31 | 2.36 | 1.96 | 17.5 | 29.4 |
| DIL | 9.92 | 13.6 | 18.6 | 7.33 | 3.35 | 3.21 | 3.40 | 2.49 | 2.06 | 15.7 | 33.9 |
| SEM (n = 10) | 0.4 | 0.4 | 0.3 | 0.17 | 0.10 | 0.08 | 0.18 | 0.09 | 0.07 | 1.2 | 2.2 |
| Target body weight | |||||||||||
| RBW | 10.5 | 13.7 | 19.1 | 7.05 | 3.36 | 3.14 | 3.42 | 2.45 | 2.04 | 16.0 | 33.3 |
| HBW | 10.3 | 15.0 | 19.2 | 6.66 | 3.25 | 3.11 | 3.30 | 2.40 | 1.98 | 17.3 | 30.1 |
| SEM (n = 10) | 0.4 | 0.4 | 0.3 | 0.17 | 0.10 | 0.08 | 0.18 | 0.09 | 0.07 | 1.2 | 2.2 |
| Treatments | |||||||||||
| CON / RBW | 10.8 | 15.1a | 19.7 | 6.67a,b | 3.20 | 3.04 | 3.31 | 2.35 | 2.01 | 16.7 | 28.9 |
| CON / HBW | 10.9 | 15.2a | 19.6 | 6.08b | 3.32 | 3.04 | 3.52 | 2.56 | 2.08 | 18.4 | 29.9 |
| DIL / RBW | 10.2 | 12.4b | 18.6 | 7.42a | 3.53 | 3.25 | 3.30 | 2.38 | 1.91 | 15.3 | 37.6 |
| DIL / HBW | 9.70 | 14.7a,b | 18.7 | 7.24a,b | 3.18 | 3.17 | 3.29 | 2.42 | 2.05 | 16.1 | 30.2 |
| SEM (n = 5) | 0.5 | 0.6 | 0.5 | 0.24 | 0.14 | 0.11 | 0.25 | 0.12 | 0.10 | 1.6 | 3.1 |
| P-value | |||||||||||
| Diet | 0.084 | 0.010 | 0.039 | 0.001 | 0.505 | 0.146 | 0.702 | 0.319 | 0.331 | 0.308 | 0.194 |
| Target BW | 0.744 | 0.042 | 0.940 | 0.120 | 0.425 | 0.748 | 0.633 | 0.676 | 0.550 | 0.471 | 0.347 |
| Interaction | 0.312 | 0.003 | 0.217 | 0.002 | 0.232 | 0.488 | 0.913 | 0.608 | 0.708 | 0.787 | 0.223 |
a,bLeast square means not sharing a common superscript differed as compared using Tukey's Honestly Significant Difference test (Tukey, 1953).
Neither diet density nor growth curve affected crude protein and moisture content (Table 6) of body weight at different ages. Pullets’ body protein content was 65.3, 62.7, and 63.5%, on average, at 6, 13, and 19 wk of age. However, some significant effects were observed for ash and body fat composition. At 6 wk of age, an interaction of diet density and growth curve was observed in ash content (P = 0.012). Pullets with RBW fed the DIL diet had the lowest amount of body ashes (10.7%), whereas those fed the CON diet had the highest body ash concentration (13.7%); however, heavier pullets had intermediate and similar body ash content (12.5%) independent of the diet fed. Afterwards, at 13 wk of age, HBW pullets tended (P = 0.074) to have lower ash content (12.2%) than the RBW ones (14.5%). However, this effect was not shown at 19 wk of age, when the average body ash content was 11.6%. Body fat content increased with age from 12.8% at wk 6 to 15.9% at wk 13 and to 19.8% at 19 wk of age, on average. The effect of treatments was observed only at wk 19, when body fat content was higher in pullets fed CON than DIL diets (22.2 vs. 17.4% respectively, P = 0.009). However, when the DIL diet was supplied to HBW pullets, the body fat content was as high as that obtained with the CON diet; meanwhile, when supplied to RBW, the body fat content was much lower (14.0%, interaction P = 0.003).
Table 6.
Effects of diet density and growing curve and their interaction on body composition (% body weight on DM basis) of broiler breeder pullets at 6, 13, and 19 wk of age.
| Crude protein, % DM | Ash, % DM | Fat, % DM | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Effects | 6 wk | 13 wk | 19 wk | 6 wk | 13 wk | 19 wk | 6 wk | 13 wk | 19 wk |
| Diet density | |||||||||
| CON | 65.6 | 64.4 | 62.4 | 12.8 | 13.5 | 12.0 | 12.9 | 15.2 | 22.2 |
| DIL | 66.0 | 61.0 | 64.6 | 11.8 | 13.2 | 11.3 | 12.6 | 16.7 | 17.4 |
| SEM (n = 10) | 1.2 | 1.8 | 1.2 | 0.5 | 0.9 | 0.7 | 1.1 | 1.1 | 1.2 |
| Target body weight | |||||||||
| RBW | 67.2 | 64.3 | 62.9 | 12.2 | 14.5 | 11.5 | 12.4 | 16.4 | 18.6 |
| HBW | 64.4 | 61.2 | 64.1 | 12.5 | 12.2 | 11.8 | 13.1 | 15.4 | 21.0 |
| SEM (n = 10) | 1.2 | 1.8 | 1.2 | 0.5 | 0.9 | 0.7 | 1.1 | 1.1 | 1.2 |
| Treatments | |||||||||
| CON / RBW | 67.2 | 67.8 | 61.0 | 13.7a | 15.1 | 12.0 | 12.8 | 16.0 | 23.1a |
| CON / HBW | 64.0 | 61.1 | 63.8 | 12.0a,b | 11.9 | 12.0 | 12.9 | 14.3 | 21.3a,b |
| DIL / RBW | 67.2 | 60.8 | 64.9 | 10.7b | 13.8 | 11.0 | 12.1 | 16.8 | 14.0b |
| DIL / HBW | 64.8 | 61.3 | 64.3 | 13.0a,b | 12.6 | 11.6 | 13.2 | 16.5 | 20.8a,b |
| SEM (n = 5) | 1.6 | 2.5 | 1.7 | 0.6 | 1.2 | 1.0 | 1.5 | 1.5 | 1.8 |
| P-values | |||||||||
| Diet | 0.805 | 0.178 | 0.210 | 0.122 | 0.783 | 0.515 | 0.877 | 0.323 | 0.009 |
| Target BW | 0.095 | 0.218 | 0.509 | 0.678 | 0.074 | 0.767 | 0.685 | 0.492 | 0.166 |
| Interaction | 0.407 | 0.120 | 0.410 | 0.012 | 0.271 | 0.892 | 0.961 | 0.713 | 0.003 |
a,bLeast square means not sharing a common superscript differed as compared using Tukey's Honestly Significant Difference test (Tukey, 1953).
DISCUSSION
Effect of Diet Density
Energy intake was similar among the different dietary treatments (8.71 and 8.80 kcal AMEn/ g BW in the DIL and CON diet, respectively); accordingly, differences in feed intake were expected to reach the desired body weights. Thus, FCR was 14.6% higher with diluted diets (3.85) compared to the control diets (3.36), whereas calculated energy efficiency was only 7% lower with the DIL diet (0.190 and 0.203 energy retained/AMEn consumed, respectively) and determined protein efficiency slightly higher (0.384 and 0.375 CP retained/CP consumed, respectively). Zuidhof et al. (2015) using oat hulls at 25% to dilute the diets obtained 22% higher FCR, but the energy and protein consumed/g BW were 4.8 and 6.0% lower than the control, suggesting a better energy and feed efficiency.
Opposite our expectation, the current study did not show reduced BW cv at 19 wk of age, when supplied diets were 15% diluted, nor more time spent eating at any age. The lack of differences could be explained, because our feed was produced in crumbles, which may reduce differences among the diets’ densities (59.9 ± 2.5 and 54.0 ± 1.5 kg/Hl for CON and DIL diets, respectively) and reduced particles, which may produce a high eating rate, 0.916 g/min on average. On the other hand, the feeding rate test showed that the animals under diluted diets were less motivated to eat both after a period of restriction and ad libitum. These differences in the feeding rate test cannot be explained by density, because the volume ingested in 2 min by animals fed DIL was 22 and 16% lower than those fed CON, in restricted and ad libitum conditions. These results agree with those of Nielsen et al. (2011) and Savory and Maros (1993), which obtained lower feed motivation (30% in feed intake reduction) with diluted diets. Moreover, Nielsen (1999) stated that hunger is associated with faster feed intake. Considering our results and his assumption, we could say that animals fed with the 15% diluted diet felt less hunger than the pullets reared with the control diet. The lower compensatory feed intake with DIL corroborates that the diluted diets may result in lower feeding motivation, according to De Jong et al. (2003), who considered the compensatory intake at 48 h as an indicator of hunger in broiler breeders.
Contrary to what was found by other authors (Qaisrani et al., 2013; Morrissey et al., 2014), no differences in the pullets’ behavior related to positive or negative welfare indicators were found in our work. The differences in the methodology used, visual scan against video camera records, could partially explain the limited significant results observed in these variables. However, we observed that activities changed with the age and with the time of d, and these changes seem to be related to social interactions (pecking other birds) and feeding. Feed allowance is an important factor that determines the activities, and as feed was provided during the morning, it steered the differences among the morning and the evening.
Diet dilution affected final BW conformation and composition and the weight distribution. Thus, breast meat yield and body fat composition at 19 wk were lower in pullets fed the DIL diet than those fed the CON diet; however, no differences were observed in the body protein content. At the same time, digestive tract and gizzard weights of pullets fed the DIL diet were consistently higher at the 3 ages in which they were measured, as also was observed by Enting et al. (2007). An excess of feed restriction could lead to low fat deposition, and although in this study nutrients supplied were similar among the different diets, the DIL diet provoked lower fat content, especially in RBW pullets. A clear explanation was not found as according the protein and energy intake, the nutrient efficiency among diets was not different. However, the variation obtained in the carcass traits got wider than that reported in other studies (Renema et al., 1999; Robinson et al., 2007).
Effect of Target Body Weight
Breeder pullets with HBW consumed, on average, 12% more than those following the RBW curve. Feed intake was, on average for both treatments, 8.3% lower than calculated from the energy requirements provided by the genetics company. This difference could be explained by a higher efficiency of the breeder pullets compared to the energy recommendations or due to a deviation between the energy system used to calculate animal requirements and that applied to evaluate the diet, or both. The amount of feed consumed per 100 g of body weight was consistent for both treatments, 361 g of FI/ 100 g BW, which corresponded to 876 kcal/ 100 g BW. Renema et al. (2001) and van Emous et al. (2013) observed similar figures for several target body weights (ranging from about 2,100 to 2,400 g) of the breeder pullets at 20 weeks.
Target body weight influenced the time eating, which was on average 14% longer in pullets with higher body weight, and this effect was more pronounced when they ate diluted diets. Van Emous et al. (2013) fed mash diets to standard and 9% heavier pullets, showing that eating time was 17% longer in heavier pullets. This effect was partially explained by the higher quantity of feed required to obtain the higher body weights. In our study, feed was presented as crumble, which is consumed faster than mash (Vilariño et al., 1996) and, consequently, could also explain the lower response compared to the published data by van Emous et al. (2013). However, in our study, this longer time eating of HBW than RBW did not cause any significant response in the feeding rate test and behavior observations.
Body composition and conformation results revealed small or no differences among treatments. At 13 wks of age, pullets with HBW had greater breast meat yield than hens following the RBW and fed the DIL diet. However this result was not consistent throughout time. Van Emous et al. (2013) did not observe differences in breast meat yield when comparing standard and 9% heavier body weights, but heavier pullets had more abdominal fat at 20 weeks. Likewise, Renema et al. (1999), comparing pullets with standard and 20% higher body weight, did not observe any difference in body composition at 21 wks of age. Nevertheless, Robinson et al. (2007) observed that pullets fed to obtain 50% higher body weight had increased breast meat yields and abdominal fat (% of BW) from 6 to 18 wk of age compared to recommended body weights.
It was hypothesized that rearing breeder pullets to higher BW and feeding them diluted diets could have a positive impact in some of the traits measured. Although the interactions observed are far from being statistically significant, some results suggest that this type of strategy may be of interest. The BW coefficient of variation was the lowest in HBW pullets fed the diluted diets. The body fat composition that was reduced with diluted diets is increased when breeder pullets are grown to HBW. This is an important finding, as the body fat content at rearing is related to the egg persistency in the second half of the production phase (van Emous et al., 2015). The fact that the feed allowance of the treatment DIL-HBW was 28% higher compared to CON-RBW makes us expect higher differences than observed in eating time and behavior. However, only a non-significant 20% reduction in the percentage of animals pecking objects was observed (53 vs. 42%, on average, for CON-RBW and DIL-HBW treatments, respectively).
CONCLUSIONS
Feeding low-density diets to breeder pullets, formulated with rich fiber raw materials and produced as crumble, does not have an impact on body weight uniformity at the end of the rearing phase, nor on behavioral activities. Diluted diets reduce feeding motivation. It can contribute to control breast meat yield growth and results in leaner carcasses without affecting the protein content of the whole body. Rearing animals to HBW resulted in higher feed intake and time eating without reducing their feeding motivation or changing their behavior. Finally, it can be concluded that feeding DIL diets to HBW pullets could be used for increasing the time spent eating and reducing their feeling of hunger without negative effects on body composition. The impact of these strategies should be followed and analyzed during the next production phase.
REFERENCES
- AOAC International 2005. Official Methods of Analysis of Association of Official Analytical Chemists International. 18th ed AOAC Int., Gaithersburg, MD. [Google Scholar]
- Aviagen 2012. Broiler Management Manual: Ross 308. Aviagen, Ltd., Huntsville, AL. [Google Scholar]
- Boletín Oficial del Estado 2013. Real Decreto 53/2013 sobre protección de los animales utilizados para experimentación y otros fines científicos.
- Bruggeman V., Onagbesan O., Ragot O., Metayer S., Cassy S., Favreau F., Jego Y., Trevidy J., Tona K., Williams J., Decuypere E., Picard M.. 2005. Feed allowance-genotype interactions in broiler breeder hens. Poult. Sci. 84:298–306. [DOI] [PubMed] [Google Scholar]
- De Jong I. C., Voorst S. V., Ehlhardt D. A., Blokhuis H. J.. 2002. Effects of restricted feeding on physiological stress parameters in growing broiler breeders. Br. Poult. Sci. 43:157–168. [DOI] [PubMed] [Google Scholar]
- De Jong I. C., Van Voorst S., Blokhuis H. J.. 2003. Parameters for quantification of hunger in broiler breeders. Physiol. Behav. 78:773–783. [DOI] [PubMed] [Google Scholar]
- De Jong I., Enting H., Van Voorst A., Blokhuis H.. 2005. Do low-density diets improve broiler breeder welfare during rearing and laying? Poult. Sci. 84:194–203. [DOI] [PubMed] [Google Scholar]
- Enting H., Veldman A., Verstegen M. W. A., Van Der Aar P. J.. 2007. The effect of low-density diets on broiler breeder development and nutrient digestibility during the rearing period. Poult. Sci. 86:720–726. [DOI] [PubMed] [Google Scholar]
- Havenstein G., Ferket P., Qureshi M.. 2003a. Growth, livability, and feed conversion of 1957 versus 2001 broilers when fed representative 1957 and 2001 broiler diets. Poult. Sci. 82:1500–1508. [DOI] [PubMed] [Google Scholar]
- Havenstein G., Ferket P., Qureshi M.. 2003b. Carcass composition and yield of 1957 versus 2001 broilers when fed representative 1957 and 2001 broiler diets. Poult. Sci. 82:1509–1518. [DOI] [PubMed] [Google Scholar]
- Heck A., Onagbesan O., Tona K., Metayer S., Putterflam J., Jego Y., Trevidy J. J., Decuypere E., Williams J., Picard M., Bruggeman V.. 2004. Effects of ad libitum feeding on performance of different strains of broiler breeders. Br. Poult. Sci. 45:695–703. [DOI] [PubMed] [Google Scholar]
- Hocking P. M. 1993. Welfare of broiler breeder and layer females subjected to food and water control during rearing: Quantifying the degree of restriction. Br. Poult. Sci. 34:53–64. [Google Scholar]
- Hocking P. M., Maxwell M. H., Robertson G. W., Mitchell M. A.. 2002. Welfare assessment of broiler breeders that are food restricted after peak rate of lay. Br. Poult. Sci. 43:5–15. [DOI] [PubMed] [Google Scholar]
- Hocking P. M. 2004. Roles of body weight and feed intake in ovarian follicular dynamics in broiler breeders at the onset of lay and after a forced molt. Poult. Sci. 83:2044–2050. [DOI] [PubMed] [Google Scholar]
- Mench J. A. 2002. Broiler breeders: Feed restriction and welfare. World's Poult. Sci. 58:23–29. [Google Scholar]
- Morrissey K. L. H., Widowski T., Leeson S., Sandilands V., Arnone A., Torrey S.. 2014. The effect of dietary alterations during rearing on growth, productivity, and behavior in broiler breeder females. Poult. Sci. 93:285–295. [DOI] [PubMed] [Google Scholar]
- Nielsen B. L. 1999. On the interpretation of feeding behaviour measures and the use of feeding rate as an indicator of social constraint. Appl. Anim. Behav. Sci. 63:79–91. [Google Scholar]
- Nielsen B. L., Thodberg K., Malmkvist J., Steenfeldt S.. 2011. Proportion of insoluble fibre in the diet affects behaviour and hunger in broiler breeders growing at similar rates. Anim. 5:1247–1258. [DOI] [PubMed] [Google Scholar]
- Qaisrani S. N., Van Krimpen M. M., Kwakkel R. P.. 2013. Effects of dietary dilution source and dilution level on feather damage, performance, behavior, and litter condition in pullets. Poult. Sci. 92:591–602. [DOI] [PubMed] [Google Scholar]
- Rauw W. M., Kanis E., Noordhuizen-Stassen E. N., Grommers F. J.. 1998. Undesirable side effects of selection for high production efficiency in farm animals: A review. Livest. Prod. Sci. 56:15–33. [Google Scholar]
- Renema R. A., Robinson F. E., Newcombe M., McKay R. I.. 1999. Effects of body weight and feed allocation during sexual maturation in broiler breeder hens. I. Growth and carcass characteristics. Poult. Sci. 78:619–628. [DOI] [PubMed] [Google Scholar]
- Renema R. A., Robinson F. E., Goerzen P. R.. 2001. Effects of altering growth curve and age at photostimulation in female broiler breeders. 1. Reproductive development. Can. J. Anim. Sci. 81:467–476. [Google Scholar]
- Renema R. A.,, Robinson F. E.. 2004. Defining normal: Comparison of feed restriction and full feeding of female broiler breeders. World's Poult. Sci. 60:508–522. [Google Scholar]
- Richards M. 2003. Genetic regulation of feed intake and energy balance in poultry. Poult. Sci. 82:907–916. [DOI] [PubMed] [Google Scholar]
- Robinson F. E., Wilson J. L., Yu M. W., Fasenko G. M., Hardin R. T.. 1993. The relationship between body weight and reproductive efficiency in meat-type chickens. Poult. Sci. 72:912–922. [Google Scholar]
- Robinson F. E., Zuidhof M. J., Renema R. A.. 2007. Reproductive efficiency and metabolism of female broiler breeders as affected by genotype, feed allocation and age at photostimulation. I. Pullet growth and development. Poult. Sci. 86:2256–2266. [DOI] [PubMed] [Google Scholar]
- Sakomura N., Silva R., Couto H., Coon C., Pacheco C.. 2003. Modeling metabolizable energy utilization in broiler breeder pullets. Poult. Sci. 82:419–427. [DOI] [PubMed] [Google Scholar]
- SAS Institute Inc 2012. SAS 9.4. SAS Institute Inc., Cary, NC. [Google Scholar]
- Savory C. J., Maros K.. 1993. Influence of degree of food restriction, breeder chickens age and time of day on behaviour of broiler. Behav. Processes 29:179–189. [DOI] [PubMed] [Google Scholar]
- Van Emous R. A., Kwakkel R., Van Krimpen M., Hendriks W.. 2013. Effects of growth pattern and dietary protein level during rearing on feed intake, eating time, eating rate, behavior, plasma corticosterone concentration, and feather cover in broiler breeder females during the rearing and laying period. Appl. Anim. Behav. Sci. [Google Scholar]
- Van Emous R. A., Kwakkel R. P., Van Krimpen M. M., Hendriks W. H.. 2015. Effects of dietary protein levels during rearing and dietary energy levels during lay on body composition and reproduction in broiler breeder females. Poult. Sci. 94:1030–1042. [DOI] [PubMed] [Google Scholar]
- Vilariño M., Picard M. L., Melcion J. P., Faure J. M.. 1996. Behavioural adaptation of laying hens to dilution of diets under mash and pellet form. Br. Poult. Sci. 37:895–907. [DOI] [PubMed] [Google Scholar]
- Wilson H. R.,, Harms R. H.. 1986. Performance of broiler breeders as affected by body weight during the breeding season. Poult. Sci. 65:1052–1057. [Google Scholar]
- Yu M. W., Robinson F. E., Charles R. G., Weingardt R.. 1992. Effect of feed allowance during rearing and breeding on female broiler breeders: 2. Ovarian morphology and production. Poult. Sci. 71:1750–1761. [DOI] [PubMed] [Google Scholar]
- Zuidhof M. J., Holm D. E., Renema R. A., Jalal M. A., Robinson F. E.. 2015. Effects of broiler breeder management on pullet body weight and carcass uniformity. Poult. Sci. 94:1389–1397. [DOI] [PubMed] [Google Scholar]