

Abstract
People perform better in visual search when the target feature repeats across trials (intertrial feature priming [IFP]). Here, we investigated whether repetition of a feature singleton's color modulates stimulus-driven shifts of spatial attention by presenting a probe stimulus immediately after each singleton display. The task alternated every two trials between a probe discrimination task and a singleton search task. We measured both stimulus-driven spatial attention (via the distance between the probe and singleton) and IFP (via repetition of the singleton's color). Color repetition facilitated search performance (IFP effect) when the set size was small. When the probe appeared at the singleton's location, performance was better than at the opposite location (stimulus-driven attention effect). The magnitude of this attention effect increased with the singleton's set size (which increases its saliency) but did not depend on whether the singleton's color repeated across trials, even when the previous singleton had been attended as a search target. Thus, our findings show that repetition of a salient singleton's color affects performance when the singleton is task relevant and voluntarily attended (as in search trials). However, color repetition does not affect performance when the singleton becomes irrelevant to the current task, even though the singleton does capture attention (as in probe trials). Therefore, color repetition per se does not make a singleton more salient for stimulus-driven attention. Rather, we suggest that IFP requires voluntary selection of color singletons in each consecutive trial.
Keywords: visual search, intertrial priming, feature singleton, saliency, stimulus-driven attention, goal-driven attention
Introduction
At each moment, we must select some visual information for detailed processing and some to ignore. This is a difficult challenge as we often lack explicit knowledge regarding which locations and features in a scene are most relevant. Empirical evidence shows that recent experience implicitly guides attention, as if the brain assumes that the world is stable and the most relevant features a moment ago are likely to remain so (Awh, Belopolsky, & Theeuwes, 2012; Chun & Nakayama, 2000; Lamy & Kristjánsson, 2013; White, Rolfs, & Carrasco, 2013; Yashar & Lamy, 2010b).
Intertrial feature priming (IFP) in visual search, also known as priming of popout (PoP), provides evidence for such experience-based effects on attention. For example, when observers have to find and discriminate a target defined as a color singleton (e.g., a green item among red items), they are faster and more accurate when the target color repeats than when it switches in consecutive trials (e.g., Maljkovic & Nakayama, 1994; Yashar, Makovski, & Lamy, 2013).
The IFP effect is considered to reflect an involuntary bias of spatial attention toward items with the previously selected feature value (e.g., Asgeirsson, Kristjánsson, & Bundesen, 2014; Becker, 2008b; Chun & Nakayama, 2000; Fecteau & Munoz, 2003; Maljkovic & Nakayama, 1994; but see Huang, Holcombe, & Pashler, 2004). According to models of spatial attention, locations are prioritized based on their saliency (e.g., local feature contrast) and their task relevance (e.g., Borji & Itti, 2013; Fecteau & Munoz, 2006; White & Munoz, 2017). According to what we refer to as the saliency hypothesis of IFP, saliency values accumulate over stimulus repetition (Fecteau & Munoz, 2003; Maljkovic & Nakayama, 1994). As a result, repetition of a feature singleton increases the saliency value of its defining feature value, which leads to a stronger or faster shift of spatial attention toward the repeated feature (e.g., Becker, 2008b; Fecteau & Munoz, 2003; Lee, Mozer, & Vecera, 2009; Maljkovic & Nakayama, 1994; Theeuwes & Van der Burg, 2013). This presumably involuntary shift can be considered stimulus driven because it is driven by the stimulus sequence rather than by task goals.
A strong version of the saliency hypothesis predicts that singletons attract stimulus-driven spatial attention more strongly when their colors repeat than switch across trials, regardless of the task. Indeed, that task independence is central to what we mean by “saliency.” This prediction has been only partially tested because, on one hand, studies that investigated IFP caused by task-irrelevant stimuli did not directly measure spatial attention (Fecteau, 2007; Huang et al., 2004; Kristjánsson, 2006; Michal, Lleras, & Beck, 2014), and on the other hand, studies that directly measured spatial attention did not test task-irrelevant attention and IFP (Belopolsky, Schreij, & Theeuwes, 2010; Folk & Remington, 2008; Irons, Remington, & Folk, 2012).
A strong version of the saliency hypothesis also predicts that IFP should emerge even when the first color singleton in the pair of two consecutive trials was merely involuntarily attended. However, no study has directly tested this prediction; investigations that focused on the role of task and response during the first trial did not directly measure spatial attention on that trial (Kristjánsson, Saevarsson, & Driver, 2013; Yashar et al., 2013).
In this study, we tested this hypothesis by interleaving singleton search task trials with a probe task designed to measure stimulus-driven (involuntary) spatial attention shifts. Stimulus-driven attention is typically manipulated and measured using variations of the Posner cueing paradigm (Posner, Snyder, & Davidson, 1980): A task-irrelevant cue appears at one peripheral location, followed by a “probe” stimulus at either the same or a different location. Facilitation of probe discrimination performance on same-location trials (the relative location effect) indicates that attention was shifted by the cue (reviewed by Carrasco, 2011). Cue saliency can alter the magnitude of these effects; for instance, increasing cue contrast leads to a larger attentional effect (e.g., Fuller, Park, & Carrasco, 2009).
Recently, White, Lunau, and Carrasco (2014) extended the spatial cueing protocol to study the attentional effects of feature singletons by using singletons as task-irrelevant spatial cues. Consistent with stimulus-driven attentional capture, sensitivity to the probe was higher when it appeared in the same than in a different location as a task-irrelevant color singleton (relative location effect). Studies on IFP typically use similar singleton displays but with search tasks in which some aspect of the singleton itself must be discriminated. During such tasks, attention must shift and engage on the singleton target (e.g., Duncan & Humphreys, 1992) using both stimulus-driven (exogenous) and goal-driven (endogenous) guidance. In contrast, during the probe task used by White et al. (2014), attention only shifted to the feature singleton in a purely stimulus-driven manner, without needing to engage on it.
In this study, we directly tested whether repetition of a singleton's color per se increases stimulus-driven shifts of spatial attention regardless of the task. Specifically, we measured the effect of feature singleton repetition in a search task and in a probe task, using the same sequence of stimuli within all trials. In search trials, the effect of intertrial priming would facilitate responses to the singleton target if its color matched the singleton in the previous trial. In probe trials, if IFP could be explained by an increase in the singleton's saliency such that it captures spatial attention more strongly, then the relative location effect would be larger when the singleton's color repeats than when it switches across trials, even when it is task irrelevant. We tested for this effect in two conditions. The first condition was in probe trials that immediately follow probe trials. Although the singleton is task irrelevant in both trials, the relative location effect in such probe trials would indicate that stimulus-driven attention was directed to the singleton during both the current and the previous trials. This condition enables us to test whether stimulus-driven attention is sufficient to induce a color repetition effect. Moreover, because in such trials color repetition is within the context of the same task, this condition also enables us to ensure that the results are not due to task switching (e.g., Fecteau, 2007). Second, we also included conditions in which probe trials immediately follow search trials. In such conditions, the singleton on the previous trial was task relevant and voluntarily attended. This condition maximizes the potential for a modulation of stimulus-driven attention according to the saliency hypothesis.
Experiment 1
We adapted White et al.'s (2014) design and added conditions in which the observer actively responded to the color singleton during trial n-1: We interleaved trials of a search task between trials of the probe task. The two tasks alternated in pairs of trials (i.e., SSPPSSPP). In the search task, the probe was irrelevant and observers discriminated the orientation of the color singleton ring, which had a gap near the top. According to the classic IFP effect, search performance should improve when the singleton color repeats from a previous search trial. In the probe task, the singleton was irrelevant and observers discriminated the orientation of the probe. If singleton color repetition affects stimulus-driven attention, then color repetition would interact with the location effect in the probe task, especially when the previous trial was a search trial (such that on trial n-1, the observer had to actively discriminate the color singleton).
This design also enabled us to test alternative explanations regarding the role of the task in IFP. If IFP requires active singleton discrimination during the previous trial (Kristjánsson et al., 2013; Yashar et al., 2013), that is, during the encoding of the primed feature into short-term memory, then we should find IFP in search trials that follow other search trials but not those that follow probe trials (because singletons are to be ignored in probe trials).
Methods
Observers
Twelve observers participated (seven women, ages 19–27 years). All had normal or corrected-to-normal vision and were naïve to the purposes of the study (except coauthor W. F.). This preset sample size is similar to that in studies showing IFP on accuracy (Yashar & Lamy, 2010a). The New York University Institutional Review Board approved this study.
Apparatus
The experiment was conducted in a quiet darkened room. Stimuli were presented on a calibrated and linearized Sony GDM-F520 screen (1,280 × 960 pixels, 85-Hz refresh rate), with a chin rest 57 cm from the screen. An Apple iMac computer with MATLAB (Mathworks) and MGL (http://gru.stanford.edu/doku.php/mgl/overview) controlled stimulus presentation and collected observers' responses.
Stimuli and procedure
Figure 1 illustrates the trial sequence. The screen background was medium gray. Each trial began with the fixation display, which consisted of a black fixation mark at the center of the screen and “landmarks” at each corner of an imaginary square, 6° of visual angle (dva) from fixation, which marked the potential singleton and probe locations. Each landmark was composed of four black dots at the vertices of a 2.2-dva imaginary diamond. Along with the fixation mark, a task cue was presented at the center of the screen throughout the whole trial. The cue indicates both task type (“G” for grating in the probe task or “R” for ring in the search task) and trial number for that task (“1” for first or “2” for second).
Figure 1.

Illustration of (a) the sequence of events in the search and the probe trials in Experiment 1 and (b) the singleton displays in all four experiments, with varying set size. Tasks switched after every two trials (SSPPSSPP…), but the stimulus sequence was the same in all trials. Note that in Experiment 3, all trials were probe trials. Each trial's task was indicated by a letter near the fixation mark (R for search task and G for probe task). In all four experiments, there were only four potential singleton and probe locations, which were each marked by four landmark dots.
The singleton display was a circle of four equally spaced rings 6 dva from fixation. Each ring had a gap 20 radial degrees wide. In search trials, the gap was near the top of the ring but slightly rotated. The search task was to report the gap orientation in the color singleton: clockwise or counterclockwise of vertical. In probe trials, the gap was oriented straight down, all the colored rings were irrelevant and uninformative, and the task was to report the orientation of the Gabor probe.
After 1-s fixation, a 94-ms singleton display appeared, which consisted of the fixation display along with a circle of four equally spaced, isoeccentric rings (6 dva from fixation). Each ring was 0.4 dva thick with an outer diameter of 2.4 dva. Each ring had a gap 20 radial degrees wide. In search trials, the gap was near the top of the ring but slightly rotated. One of the rings at the landmark locations (randomly selected) was a color singleton, either a red among greens or green among reds. The singleton color was randomly determined with equal probability such that singleton color had a 50% chance to repeat or to switch across two consecutive trials.
After a 12-ms interstimulus interval, a probe display appeared for 35 ms. The probe display consisted of the fixation display and a 35% contrast Gabor stimulus (4 cpd sinusoidal grating embedded in a Gaussian envelope with 0.25 dva standard deviation, 1.75 dva diameter), which was randomly tilted right or left from vertical. On half of the trials, the Gabor location coincided with the singleton location (same-location trials). In the remaining trials, the Gabor was at the opposite location from the singleton (different-location trials). Same-location and different-location trials were randomly intermixed. The singleton's location and color were completely independent of each other. Note that the singleton did reduce uncertainty about the probe's location from four possible locations to two. However, the delay between the onsets of the singleton and the probe was very brief (106 ms), and the probe itself was of high enough contrast to be localized easily on its own. Therefore, in addition to the fact that the probe appeared far from the singleton on a random half of trials, there was little reason for the observer to attend to the singleton during the probe task. The trial intervals were also designed to be brief enough to prevent saccadic eye movements between singleton onset and probe offset (total time = 141 ms).
The Gabor stimulus was followed by a response screen, identical to the fixation display. Following the onset of the response screen, observers reported either the orientation of the Gabor probe (in probe trials) or the orientation of the color singleton ring (in search trials) by pressing one of four keys (“z,” “x” for counterclockwise or clockwise of vertical, respectively for items on the left hemifield; “<” or “>” for counterclockwise or clockwise of vertical, respectively, for items on the right hemifield). Accuracy was stressed, and response time was unlimited. As soon as the observer responded, an auditory tone indicated whether the response was correct. Note that the stimulus sequence was the same between the tasks. Observers were instructed to fixate on the “fixation mark” throughout the trial.
Design
Throughout the experiment, the probe task and the search task alternated after every two trials, allowing for a systematic manipulation of two intertrial effects: task repeated versus switched and singleton color repeated versus switched. Over three 1-hr sessions conducted on different days, each observer completed 23 blocks of 96 trials (2,208 trials).
At the beginning of the first session, observers practiced each task until performance reached above chance with orientation ≤30°. Experimenters stressed the irrelevance of the colored rings in probe trials. QUEST adaptive staircases (Watson & Pelli, 1983) were used to adjust the Gabor's (probe trials) and the ring's (search trials) tilt to reach 80% correct performance before proceeding to the main experiment. The trial sequence in the staircases was exactly the same as in the main experiment. Each observer completed two staircase blocks of 85 trials per session, and then the probe tilt magnitude was fixed at the estimated threshold.
Results
We excluded trials with response times (RTs) ≤250 ms (≤0.1%). To optimize the IFP effect, we also excluded trials that followed an error trial (13%; Lamy, Zivony, & Yashar, 2011; Yashar & Lamy, 2010a). When analyzing the geometric means of RTs, we excluded incorrect trials and trials with RTs ≥4 SDs above the observer's mean (≤0.5% of the trials).
Search task
Figure 2 depicts the color repetition effects on accuracy (top panel) and RTs (bottom panel) for each task order (first and second search trials in the pair). For both error rates and RTs, we conducted a two way (2 × 2) repeated-measures analysis of variance (ANOVA) with factors task order and color repetition (Figure 2a).
Figure 2.

Results of Experiment 1. (a) Mean accuracy (top panel) and RTs (bottom panel) for the search task as a function of color repetition and trial order (first or second search trial in the pair). The classic IFP effect can be seen as higher accuracy and lower RTs when the color repeats, in the second search trials. (b) Mean accuracy (top panels) and RTs (bottom panels) as a function of relative location and color repetition (left panel) in all probe trials and (right panel) in probe trials that followed search trials. The stimulus-driven attentional effect of the singleton can be seen as higher accuracy and lower RTs on same-location than different-location trials. That effect did not depend on probe trial order or singleton color repetition. Error bars are ±1 within-subject standard error (Morey, 2008). *p < 0.05. **p < 0.01. ***p < 0.001.
Accuracy:
Neither the main effects of color repetition, F(1, 11) = 4.10, p = 0.068, η2 = 0.27 and task order, F(1, 11) = 3.50, p = 0.088, η2 = 0.24, nor their interaction, F(1, 11) = 1.37, p > 0.2, were significant.
RT:
The main effect of color repetition was significant, F(1, 11) = 78.39, p < 0.001, η2 = 0.87, with RTs being on average 31 ms faster when the color repeated. The main effect of task order was significant, F(1, 11) = 19.77, p = 0.002, η2 = 0.64, with RTs being on average 29 ms faster on the second trial. The two effects significantly interacted, F(1, 11) = 11.39, p = 0.006, η2 = 0.5, indicating that the color repetition effect was stronger on the second search trials (which followed another search trial) than on the first search trials (which followed a probe trial).
Classic IFP: Search task following search task
Previous demonstrations of the IFP effect always used tasks in which every trial required search for the singleton. Accordingly, we conducted planned comparisons of the effect of color repetition within search trials that followed search trials (task order 2), to evaluate the IFP effect under “classic” conditions. Accuracy was higher, F(1, 11) = 6.12, p = 0.031, η2 = 0.35 (M = 3%, SE = 1%), and RTs were faster, F(1, 11) = 64.28, p < 0.001, η2 = 0.85 (M = 46 ms, SE = 5.6), when the singleton color repeated than when it switched across trials. By contrast, the effect of color repetition was not significant on accuracy (F < 1) and on RTs, F(1, 11) = 4.25, p = 0.06, η2 = 0.27, in search trials following probe trials (Figure 2a).
Probe task
Figure 2b depicts the relative location effects on accuracy (top panel) and RTs (bottom panel). Our interest was the effect of relative location and color repetition on each of the two probe task orders: (a) first probe trials that followed search trials and (2) second probe trials that followed probe trials. We conducted a three-way (2 × 2 × 2) ANOVA with factors Task Order, Color Repetition, and Relative Location.
Accuracy:
Only the main effect of relative location was significant, F(1, 10) = 4.98, p = 0.049, η2 = 0.31. None of the other main effects and interactions were significant (all ps > 0.1).
RT:
The main effects of relative location, F(1, 11) = 45.90, p < 0.001, η2 = 0.8, and task order, F(1, 11) = 14.73, p = 0.01, η2 = 0.57, were significant. The main effect of color repetition was not significant (F < 1). Neither the three-way nor the two-way interactions were significant (all ps > 0.3).
Bayes factors:
Because it is risky to accept the null hypothesis in standard significance testing, we also analyzed the interaction between the relative location effect and color repetition with a Bayesian model selection approach (Masson, 2011). This approach uses simple transformations of the sums of squares from the ANOVAs to generate Bayesian information criterion probabilities (pBIC) of the null (H0) and alternative (H1) hypotheses given the data set D. For accuracy, the Bayes factor favored the null hypothesis over the interaction between relative location and color repetition with an odds of 2.33 to 1: pBIC(H1|D) = 0.3 and pBIC(H0|D) = 0.7. For RTs, the Bayes factor yielded odds of 2.7 to 1: pBIC(H1|D) = 0.27 and pBIC(H0|D) = 0.73. In each case, the probability of the alternative hypothesis is much smaller than what is typically considered even a weak effect: A value of pBIC(H1|D) between 0.5 and 0.75 is considered a weak effect (Raftery, 1999; Dienes, 2011). A Bayes factor greater than 3 provides additional support for the null hypothesis (Raftery, 1999). These analyses therefore confirmed that there is no effect of color repetition on the relative location effect in both accuracy and RTs.
Probe task following search task
In the search task, color repetition affected performance mainly in trials that followed search trials. Therefore, to be sure that IFP does not affect bottom-up attention, we conducted an ANOVA with factors Color Repetition and Relative Location only for probe trials that followed a search trial (Figure 2b).
The main effect of probe location was significant, both on accuracy, F(1, 11) = 6.99, p = 0.02, η2 = 0.38, and on RTs, F(1, 11) = 58.66, p < 0.001, η2 = 0.84), with higher accuracy and faster RTs on singleton-probe same-location than different-location trials. Neither the main effect of color repetition nor its interaction with probe location were significant, for both accuracy and RTs (all ps > 0.1). Therefore, the relative location effect was not dependent on color repetition.
Bayes factors:
For accuracy, the Bayes factor favored the null hypothesis over the interaction between relative location and color repetition with an odds of 2.1 to 1: pBIC(H1|D) = 0.32 and pBIC(H0|D) = 0.68. For RTs, the Bayes factor yielded odds of 3.16 to 1: pBIC(H1|D) = 0.24 and pBIC(H0|D) = 0.76. These analyses confirmed that there is no effect of color repetition on the relative location effect on both accuracy and RTs.
Probe task following probe task
Given that in some cases IFP does not transfer across tasks (Fecteau, 2007), we ruled out the possibility that the IFP could have been eliminated in the probe trials because of the task switch. We conducted an ANOVA with factors Color Repetition and Relative Location only for probe trials that followed a probe trial (for which there was no task switch). The main effect of probe location was significant on RTs but not accuracy, F(1, 11) = 21.96, p < 0.001, η2 = 0.70, and p > 0.5, respectively, with faster RTs on singleton-probe same-location than different-location trials. Neither the main effect of color repetition nor its interaction with probe location were significant for either accuracy or RTs, all ps > 0.2. This confirms that the relative location effect was not dependent on color repetition, even when task-switching costs are not a factor.
Experiment 2
In the second experiment, we tested how increasing the singleton's saliency by increasing set size from four to eight would affect IFP in the search task and the stimulus-driven attentional shifts toward the color singleton in the probe task. On one hand, previous research suggests that increasing set size weakens the IFP effects in singleton search (Meeter & Olivers, 2006). On the other hand, we predicted that increasing set size would strengthen the stimulus-driven attentional effects in the probe task, if that task is sensitive to singleton salience. Note that although we increased the singleton set size from four to eight, there were still just four possible locations of the singleton and the probe, marked by outline landmarks. Thus, the probabilities for same- and different-location trials remain the same across experiments. This ensured that variation in set size modulated saliency alone while keeping target predicability the same.
Manipulating set size also enables us to rule out the possibility that the lack of interaction between color repetition and the relative location effect was because the latter is insensitive to any manipulations of singleton saliency. If saliency modulates the relative location effect, then we predict a larger effect in Experiment 2 (set size eight, high singleton saliency) than in Experiment 1 (set size four, lower singleton saliency). A statistical test of the effect of set size among all experiments is reported at the end of the Results and Discussion subsections of the Experiment 4 section.
Method
Observers
Eleven observers (seven women, ages 20–29 years) with normal or corrected-to-normal vision participated. All observers (except coauthor W. F.) were naïve to the purposes of the study.
Apparatus, stimuli, procedure, and design
Figure 1 illustrates the trial sequence and stimuli. All were the same as in Experiment 1 except that the singleton display contained eight equally spaced rings (as before, 6 dva from fixation).
Results and discussion
We excluded from all analyses trials with RTs ≤250 ms (≤0.1% of all trials). To optimize the IFP effect, we excluded trials that followed an error trial (about 10% of all trials). When analyzing the geometric means of RTs, we excluded incorrect trials and trials with RTs ≥4 SDs above the observer's mean (≤0.5% of trials).
Search task
Figure 3 depicts the color repetition effects for each task order (first search trial following a probe trial vs. second search trial following a search trial) on accuracy (top panel) and RTs (bottom panel). For both accuracy and RTs, we conducted a two-way (2 × 2) repeated-measures ANOVA with factors Task Order and Color Repetition (Figure 3a).
Figure 3.

Results of Experiment 2. (a) Mean accuracy (top panel) and RTs (bottom panel) in the search task as a function of color repetition for search trials that followed probe trials (“1st”) and search trials that followed search trials (“2nd”). These data show no IFP effect. (b) Mean accuracy (top panels) and RTs (bottom panels) in the probe task. The left panels plot the effect of singleton-probe relative location and task order. The right panels plot the effect of singleton-probe relative location and color repetition, only for probe trials that followed search trials. The stimulus-driven attentional effect of the singleton can be seen as higher accuracy and lower RTs on same-location than different-location trials. Again, that effect did not depend significantly on probe trial order or singleton color repetition. Error bars are ±1 within-subject standard error (Morey, 2008). *p < 0.05. **p < 0.01.
Accuracy:
None of the effects were significant (all ps > 0.1).
RT:
The main effect of task order was significant, F(1, 10) = 95.96, p < 0.001, η2 = 0.38, indicating that the second trial was faster than the first. The main effect of color repetition was not significant, F(1, 10) = 3.94, p = 0.075, η2 = 0.28. The two effects did not interact (p > 0.3).
Classic IFP: Search task following search task
All previous demonstrations of the IFP effect used tasks in which every trial required search for the singleton. Accordingly, we conducted planned comparisons between repeated and switched singleton color in search trials that followed search trials (trial order 2). The color repetition effect was not significant either on accuracy, F(1, 10) = 1.69, p > 0.2, or on RTs (F < 1), indicating no “classic” IFP effect.
In summary, increasing the set size from four to eight eliminated the IFP effect in search trials when the singleton was the task-relevant target. Color repetition had no effect on accuracy or RTs in search trials regardless of whether they followed a probe trial or a search trial.
Probe task
Figure 3b depicts the relative location effects on accuracy (top panel) and RTs (bottom panel). For both accuracy and RTs, we conducted a three-way (2 × 2 × 2) ANOVA with factors Task Order, Color Repetition, and Relative Location.
Accuracy:
The main effect of color repetition was not significant (F < 1). The main effect of relative location was significant, F(1, 10) = 5.01, p = 0.049, η2 = 0.33, with higher accuracy for the same than different location. There was also a main effect of task order, F(1, 10) = 7.48, p = 0.021, η2 = 0.88, with higher accuracy on the second than the first trials. The two-way interaction between task order and relative location was not significant, F(1, 10) = 3.31, p = 0.09, η2 = 0.24. The location effect in probe trials that followed a search trial was significant, F(1, 10) = 8.02, p = 0.017, η2 = 0.44; Figure 3b, right panel) but not in the probe trials that followed a probe trial, F(1, 10) = 1.24, p > 0.2. The two-way interaction was dependent on color repetition, as indicated by the significant three-way interaction among Relative Location, Task Order, and Color Repetition, F(1, 10) = 14.86, p = 0.003, η2 = 0.59. None of the other two-way interactions were significant (F < 1).
RT:
The main effect of color repetition was not significant (F < 1). The main effects of relative location, F(1, 10) = 25.86, p < 0.001, η2 = 0.72, and task order, F(1, 10) = 91.57, p < 0.001, η2 = 0.84, were significant. Neither the three-way nor the two-way interactions were significant (all Fs < 1). Indeed, the relative location effect was significant both on probe trials following a search trial, F(1, 10) = 18.23, p = 0.001, η2 = 0.64, and on probe trials following a probe trial, F(1, 10) = 27.30, p < 0.001, η2 = 0.73.
Bayes factors:
For accuracy, the Bayes factor favored the null hypothesis over the interaction between relative location and color repetition with odds 2.33 to 1: pBIC(H1|D) = 0.3 and pBIC(H0|D) = 0.7. For RTs, the Bayes factor gave odds of 3.16 to 1: pBIC(H1|D) = 0.24 and pBIC(H0|D) = 0.76. These analyses confirmed that there is no effect of color repetition on the relative location effect in both accuracy and RTs.
In summary, the probe task results show higher accuracy and faster RTs when the probe appeared at the same than at a different location as the color singleton, demonstrating that spatial attention was captured to the location of the task-irrelevant singleton. The relative location effect did not interact with color repetition: The singleton cue was effective regardless of whether its color repeated or switched from the previous trial.
Experiment 3
We next tested whether the independence of the relative location effect and IFP in Experiment 1 was due to the task switching required by interleaving pairs of trials of different tasks. To eliminate possible interference from task switching within each block, which could have modulated the color repetition effect, the two tasks—search and probe discrimination—were conducted in separate blocks of 80 trials rather than intermixed within a block. Set size was four, as in Experiment 1. If task switching underlies the pattern of results in Experiment 1, then the blocked design here should reveal an interaction between color repetition and the relative location effect.
Method
Observers
Twelve observers (eight women, ages 20–37 years) with normal or corrected-to-normal vision participated. All observers were naïve to the purposes of the study.
Apparatus, stimuli, procedure, and design
Figure 1 illustrates the trial sequence and stimuli. All were the same as in Experiment 1 except that search and probe task trials were administered in separate blocks. There were 12 blocks of 80 trials each (960 trials overall). Half of the blocks were search trials and half probe trials, in alternating order. The first block's task was counterbalanced across observers.
Results and discussion
We excluded trials with RTs ≤250 ms (≤0.1%). As in the previous experiments, we excluded trials that followed an error trial (about 13% of all trials). When analyzing the geometric means of RTs, we excluded incorrect trials and trials with RTs ≥4 SDs above the observer's mean (≤0.5% of the trials).
Search task
Figure 4a depicts the color repetition effects on accuracy (top panel) and RTs (bottom panel). RTs were faster when the singleton color repeated than when it switched, t(23) = 4.57, p < 0.001, Cohen's D = 0.47 (M = 32ms, SE = 5 ms), replicating the classic IFP effect. There was no effect or tradeoff with accuracy (t < 1).
Figure 4.
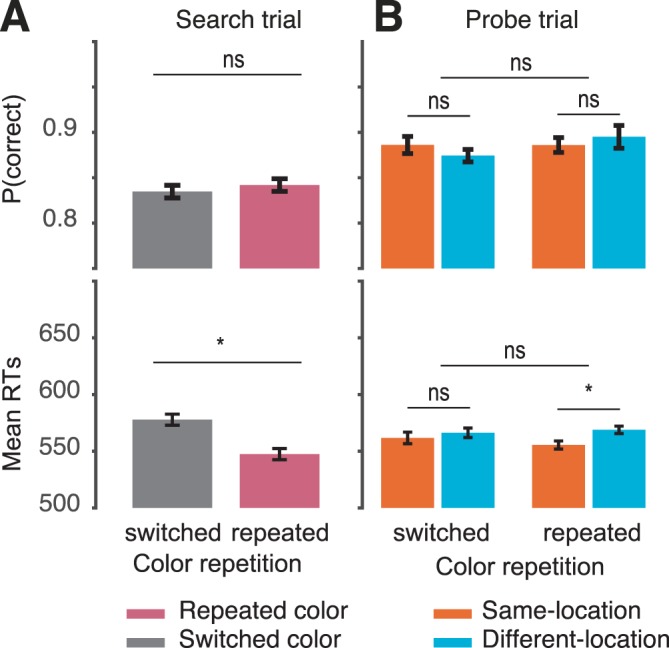
Results of Experiment 3. (a) Mean accuracy (top left panel) and RTs (bottom panel) for the search task as a function of color repetition. The classic IFP effect can be seen in the faster search RTs when the color repeats than switches. (b) Mean accuracy (top panel) and RTs (bottom panel) in probe trials as a function of relative location and color repetition. The stimulus-driven attentional effect, which did not interact with color repetition, can be seen in the faster probe RTs on same-location than different-location trials. Error bars are ±1 within-subject standard error (Morey, 2008). *p < 0.05.
Probe task
Figure 4b depicts the relative location effects on accuracy (top panel) and RTs (bottom panel). We conducted a two-way (2 × 2) ANOVA with factors Color Repetition and Relative Location. For accuracy, there was no effect of color repetition, relative location, or interaction, all ps > 0.12. Only the main effect of relative location was significant on RT, F(1, 11) = 5.37, p = 0.04, η2 = 0.35 (M = 9 ms, SE = 3 ms). Both the main effect of color and the interaction between color repetition and relative location were not significant on RTs (both ps > 0.3).
Bayes factors:
For accuracy, the Bayes factor equally favored the null hypothesis and the alternative hypothesis that there is an interaction between relative location and color repetition with an odds of 1 to 1: pBIC(H1|D) = 0.5 and pBIC(H0|D) = 0.5. However, mean accuracy showed a numerical trend toward higher relative location effect when the singleton color switched. This is the opposite to the main alternative hypothesis, that is, that the relative location effect increases with color repetition. Therefore, it is safe to say that these results show that color repetition does not increase the relative location effect. For RTs, the Bayes factor favored the null hypothesis over the interaction between relative location and color repetition with an odds of 1.94 to 1: pBIC(H1|D) = 0.34 and pBIC(H0|D) = 0.66. These analyses therefore confirmed that the color repetition does not increase the relative location effect in either accuracy or RTs.
In summary, the blocked design eliminated any possible task-switching effect, and yet the results replicated the key findings of Experiment 1. In search trials, repetition of the color singleton decreased RTs (IFP), and in probe trials, RTs were faster when the probe appeared at the same than at a different location as the color singleton (relative location effect), demonstrating attentional capture by the color singleton. Importantly, stimulus-driven attention was independent of IFP; the relative location effect did not interact with repetition of the color singleton. The results confirm that that IFP does not modulate stimulus-driven attention.
The lack of effects on accuracy is perhaps due to the fact that observers here tend to emphasize speed during response over accuracy; indeed, RTs were faster than in Experiment 1. The relative location effect on RTs in the probe task was also quite small, consistent with the pattern across experiments for weak stimulus-driven attentional capture by the singleton when set size is low (see below).
Experiment 4
We tested whether increasing singleton saliency further can produce a strong relative location effect even when the singleton display is task irrelevant in all trials. To do so, we increased the set size from four to 16 and, given that IFP in search is known to disappear with high set size, we used only blocks of probe trials (no search trials). We also included a condition with no singleton in the array of discs (i.e., homogenous colors), as a neutral condition against which benefits and costs of same- and different-location trials could be evaluated. Again, although the singleton set size was 16, there were still just four possible locations of the singleton and the probe, marked by outline landmarks (as in all previous experiments). Thus, the probabilities for same- and different-location trials remain the same across experiments.
Method
Observers
Sixteen observers (nine female, ages 18–23 years) with normal or corrected-to-normal vision participated. All observers were naïve to the purposes of the study.
Apparatus, stimuli, procedure, and design
Figure 5a illustrates the trial sequence and stimuli. All were the same as in Experiment 3, except for the following changes. Only the probe task trials were administered. The singleton display appeared for 71 ms, and the probe appeared after a 35-ms interstimulus interval. The probe was followed by a postcue, a small line (0.5 dva) extending from the fixation mark pointing toward the probe location. The postcue eliminated any location uncertainty during decision making. Over three 1-hr sessions conducted on different days, each observer completed ∼4,200 trials. At the beginning of the first session, observers practiced the task until performance reached above chance with orientation ≤30°.
Figure 5.

Trial sequence and results from Experiment 4. (a) Illustration of the sequence of events within a trial. (b) Mean accuracy and RTs as a function of singleton-probe locations (bottom-up attention effect) and color repetition. Accuracy was highest and RTs lowest when the probe appeared at the same location as the singleton, demonstrating that the irrelevant singleton captured attention. That effect did not interact with color repetition. Error bars are ±1 within-subject standard error (Morey, 2008). **p < 0.001.
Results and discussion
We excluded trials with RTs ≤250 ms (≤0.1% of all trials). To optimize the IFP effect, we also excluded trials that followed an error trial (about 18.5% of all trials). When analyzing the geometric means of RTs, we excluded incorrect trials and trials with RTs ≥4 SDs above the observer's mean (≤0.5% of the trials).
When analyzing the effect of singleton color repetition, we included only singleton trials that followed other singletons trials (excluding “neutral” trials with no singleton in the homogenous array of rings). When analyzing color repetition among trials with no singleton, we included only trials that followed trials that also had no singleton.
Mean accuracy and RTs are depicted in Figure 5b. For both measures, we conducted a repeated-measures ANOVA with factors Probe Location (same vs. different as singleton) and Singleton Color Repetition (switched vs. repeated).
Accuracy
The main effect of relative location was significant, F(2, 30) = 8.54, p = 0.001, η2 = 0.89. On average, observers were 82.7% correct on same-location trials and 80.3% on different-location trials. Planned comparisons between no-singleton and same-location trials (benefit) and between no-singleton and different-location trials (cost) reveled higher accuracy in no-singleton trials than in different-location trials, t(15) = 3.19, p = 0.006, ΔM = 2%, SE = 0.5%. There was no significant difference between no-singleton and same-location trials (t < 1), suggesting that the relative location effect was mainly due to cost in different-location trials. Neither the main effect of color repetition nor its interaction with probe location were significant (both Fs < 1).
RT
The relative location effect was significant, F(2, 30) = 28.59, p < 0.001, η2 = 0.96. On average, correct responses were 33 ms faster when the probe appeared at the singleton's location than at the opposite location. Planned comparisons reveled faster RTs in no-singleton trials than in different-location trials, t(15) = 4.22, p < 0.001, ΔM = 15 m, SE = 3 ms, and in same-location trials than in no-singleton trials, t(15) = 3.8, p = 0.001, ΔM = 18 ms, SE = 5 ms, suggesting that the location effect on RTs reflected both cost and benefit. The main effect of color repetition and the interaction between relative location and color repetition were not significant (both Fs < 1). This analysis ruled out any speed-accuracy tradeoff.
Bayes factors
For accuracy, the Bayes factor favored the null hypothesis over the interaction between relative location and color repetition with an odds of 3 to 1: pBIC(H1|D) = 0.25 and pBIC(H0|D) = 0.75. For RTs, the Bayes factor also favored the null hypothesis over the interaction between relative location and color repetition with odds of 3.34 to 1: pBIC(H1|D) = 0.23 and pBIC(H0|D) = 0.77. These analyses confirmed that there is no effect of color repetition on the relative location effect in both accuracy and RTs.
The effect of the relative location of the singleton and probe (i.e., the cueing effect) was reliable in both accuracy (mainly due to the cost in different-location trials) and RTs (due to both cost and benefit). Critically, it was independent of whether the singleton's color repeated or switched, confirming the results of Experiments 1–3.
Effect of set size on the IFP and relative location effects
We conducted a between-experiments analysis to evaluate all these manipulations. In this analysis, we used a combined measure of accuracy and response time: efficiency = mean RT/p(correct) (Townsend & Ashby, 1983) to succinctly summarize all the effects. We found IFP in search trials only when set size was four, which makes the singleton's salience low (in Experiment 1 and Experiment 3; Figure 6). We found the relative location effect in probe trials of all four experiments (Figure 6). To test whether the magnitude of the relative location effect (in units of efficiency) was modulated by saliency and task switching, we conducted a two-way ANOVA with color singleton saliency (set size high [Experiment 2 and Experiment 4] vs. low [Experiment 1 and Experiment 3]) and task switching (blocked [Experiment 3 and Experiment 4] vs. interleaved [Experiment 1 and Experiment 2]). We collapsed set sizes 16 and eight into “high” saliency because that allowed for a 2 × 2 statistical test for main effects and interactions of saliency and task switching, and both conditions showed large relative location effects. Both high saliency and interleaved task switching increased the relative location effect on efficiency, F(1, 47) = 15.89, p < 0.001, and F(1, 47) = 7.24, p = 0.009, respectively, with no significant interaction (F < 1).
Figure 6.
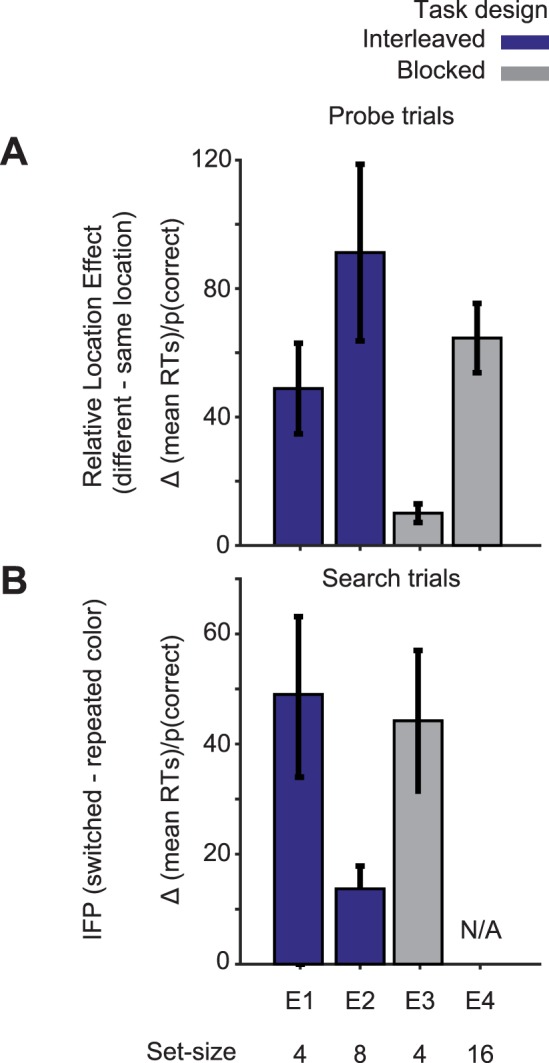
The role of task-switching and saliency across experiments in the magnitudes of (a) the relative location effects in probe trials and (b) the IFP effect in search trials. In Experiment 4 (E4), there was no search task, hence the lack of IFP value. Relative location effects and IFP were measured in units of efficiency (RT/p[correct]) (Townsend & Ashby, 1983). Error bars are ±1 standard error.
In summary, both IFP in search trials and stimulus-driven attention in probe trials were sensitive to singleton saliency when manipulated via set size, but in opposite directions. IFP is stronger when salience is low, and the attention effect is stronger when salience is high. The presence or absence of task switching (interleaved vs. blocked trials) also affected probe task performance but not its modulation by singleton color repetition.
General discussion
Summary of findings
Using identical stimulus displays, we measured two classic effects that did not interact. First, IFP in singleton search with set size four: Color repetition across pairs of search trials increased discrimination accuracy and reduced RTs. The size of that IFP effect on RT (∼46 ms) is similar to previous findings (e.g., Becker, 2008b; Kristjánsson et al., 2013; Lamy, Antebi, Aviani, & Carmel, 2008; Maljkovic & Nakayama, 1994). Second, the stimulus-driven spatial attention effect: Probes were discriminated more accurately and more quickly when their locations were precued by irrelevant singletons. This result is consistent with a previous study that used a similar protocol (White et al., 2014). However, we found that the stimulus-driven attention effect was not modulated by whether the singleton's color repeated or switched.
We conclude that IFP from trial n to trial n+1 depends on the feature singleton being task relevant and voluntarily attended on both trials. Therefore, the present findings cast doubt on the strong version of the saliency hypothesis of IFP. According to this hypothesis, repetition of an attended color singleton makes the color more salient such that it automatically draws spatial attention in a stimulus-driven fashion, that is, independently of what the observer is looking for. This hypothesis predicts that the spatial attention effect on probe discrimination performance should be larger when the singleton's color repeats than when it switches across trials, which we did not find.
Across all four experiments, we included four types of trial pairs, defined by the tasks on each trial. They each contribute one piece of evidence for our overall conclusion.
Search, search (Experiments 1–3). The singleton is voluntarily attended and discriminated on both trials, and we see the classic IFP effect as long as the set size is low.
Probe, probe (Experiments 1–4). In both trials, the singleton is task irrelevant but draws spatial attention involuntarily, as seen in the relative location effect on probe performance. Color repetition does not modulate the magnitude of that relative location effect. Therefore, involuntary attention to the singletons is not sufficient to cause IFP.
Probe, search (Experiments 1–2). The singleton is voluntarily attended only in the second trial. IFP was weak, if present at all. Therefore, voluntary attention toward the singleton in the first trial is needed for color repetition to facilitate search for the singleton in the second trial.
Search, probe (Experiments 1–2). The singleton is voluntarily attended only in the first trial. In the second trial, it is task irrelevant but draws spatial attention involuntarily. Nonetheless, color repetition does not modulate the magnitude of that stimulus-driven attention effect, contrary to the prediction of the saliency hypothesis. Therefore, voluntary attention is also needed in the second trial to have an effect of color repetition.
These four pieces of evidence lead us to conclude that color repetition per se does not modulate stimulus-driven shifts of attention toward the singleton. Voluntary attention to the first singleton is required to encode a memory trace of its color. That memory trace then influences how similar items in the future are voluntarily attended, localized, discriminated, and/or responded to but does not influence how future items automatically draw involuntary (stimulus-driven) attention in virtue of their salience in the display.
Support for our conclusion also comes from a recent study that assessed the effect of feature repetition on attentional saliency (Amunts, Yashar, & Lamy, 2014). Given that in visual search, the level of target saliency is inferred by the slope of search RTs as a function of set size, the saliency view of IFP predicts reduction of the slope in repeated trials compared with switch trials. However, repetition of the target feature did not modulate the slope of search RTs, a finding the authors considered as evidence against the saliency view.
Role of task switching
Note that the last two types of trial pairs (3 and 4, above) involve a switch of task. One study has suggested that task switching “resets” the system and eliminates the IFP effect (e.g., Fecteau, 2007). This could be why we found no effect of color repetition in probe trials that followed search trials. In other words, it is possible that following one search trial, there is an involuntary increase in the salience of that target's color, but it has no effect when the task switches. Under this hypothesis, which we cannot definitively rule out, true priming effects on singleton salience are very weak and would be difficult to measure. However, others have shown that the encoded information that underlies IFP is not automatically lost after observers switch to another task (Thomson & Milliken, 2011). For instance, Yashar and Lamy (2010a) demonstrated IFP from one orientation discrimination task (trial n) to a similar task two trials later (trial n+2), despite a different task (localization) on the interleaved trial (n+1). This finding shows that merely switching to another task does not reset the priming effect.
In any case, we note that the singleton-probe relative location effect did not depend on color repetition regardless of whether the task switched across trials (see trial pair types 2 and 4, above). Even when the probe trials were conducted in blocks with all the same task (Experiments 3 and 4), there was no effect of color repetition on how strongly the singletons captured spatial attention. From this finding, we conclude that the stimulus-driven shift of attention to a salient item operates independently of its similarity to previous items and shows no signs of priming.
Also regarding task switching, there are two pieces of evidence indicating that observers were not able to completely stop attending to the singletons when the task switched from search to probe discrimination, which make it more surprising that we found no effect of color repetition in probe performance. First, the relative location effect in probe trials was larger overall when the two tasks were interleaved (Experiment 1) than blocked (Experiment 3). Second, the relative location effect was also larger when probe trials followed interleaved search trials (the condition that maximizes IFP in the search task) than when they followed probe trials (Experiments 1 and 2). These results suggest that observers may not have fully ignored the singleton in probe trials that followed search trials. Nonetheless, those singletons did not capture attention more strongly when their color was the same as the previous task-relevant singleton than when their color switched. In fact, in some cases, the trend was in the opposite direction (larger relative location effects when the color switched; Experiment 1).
Alternative explanations
One possible alternative explanation for the lack of interaction between singleton color repetition and the relative location effect is that the cueing effect in our probe task may not be susceptible to any manipulations of singleton saliency. To test that, we directly manipulated saliency via the set size of the singleton array. Lower set sizes make the singleton less salient and increase the magnitude of the IFP effect (Meeter & Olivers, 2006). Note that in all experiments, we used the same four potential singleton and probe locations (indicated by the four “landmarks”), such that variations in set size manipulated saliency but did not vary the predictability of the singleton location with respect to the probe location. The results show that the relative location effect is modulated by set size but not by color repetition. Our probe task, therefore, was sensitive enough to reveal modulation of the stimulus-driven attentional effect by saliency. Moreover, whereas increasing saliency with set size potentiated the stimulus-driven attentional effect, it reduced or even eliminated the IFP effect in search trials (Figure 6). These findings further suggest that the two effects are independent.
A second alternative is that feature priming is not strong enough to modulate the spatial attention effect with just one repetition. In general, the magnitude of the IFP effect increases with the number of consecutive trials with the same color (Maljkovic & Nakayama, 1994). Our design and main dependent measure (accuracy) did not enable a reliable statistical analysis of more than one trial back. Further research is required to determine whether increasing the number of repetitions can lead eventually to an interaction between color repetition and the relative location effect. Importantly, however, we do show that a single repetition is sufficient to induce IFP in search trials but nonetheless does not modulate the relative location effect in probe trials.
Relation to other studies about IFP and attention
Previous studies on this topic have tested how IFP interacts with various forms of attention measured in a variety of ways, but they all involved top-down guidance or voluntary attention. For instance, some studies manipulated goal-driven rather than stimulus-driven attention to the singletons and did not have an independent manipulation of saliency (Belopolsky et al., 2010; Biderman, Biderman, Zivony, & Lamy, in press; Folk & Remington, 2008; Irons et al., 2010). Other studies did make the singletons task irrelevant, but they neither manipulated nor measured spatial attention, merely inferring its effects or lack thereof (Fecteau, 2007; Huang et al., 2004; Kristjánsson, 2006; Michal et al., 2014). In contrast, by using search and probe tasks with the same displays, we measured the classic IFP effect and directly manipulated stimulus-driven spatial attention (by using task-irrelevant singletons as exogenous “cues”; White et al., 2014).
Moreover, electrophysiological studies considered to support the involvement of attention in IFP do not necessarily entail that feature repetition modulates saliency or stimulus-driven attention. For instance, intertrial feature repetition correlates with the N2pc, an event-related potential measured from the scalp that is associated with spatial selection (Hickey, Olivers, Meeter, & Theeuwes, 2011; Töllner, Gramann, Müller, Kiss, & Eimer, 2008). But there is also evidence that the N2pc is related to target engagement or voluntary selection rather than merely a shift of spatial attention (Kiss, Van Velzen, & Eimer, 2008).
It is also possible to argue that color repetition increases the saliency or attentional priority of singleton targets because planned eye movements are facilitated when the target color repeats (e.g., Bichot & Schall, 2002; Becker, 2008a, 2008b; McPeek, Maljkovic, & Nakayama, 1999; Meeter & Van der Stigchel, 2013). Top-down guidance must be involved to direct the eyes to the singleton in such tasks, so those results are not in conflict with our conclusion. Our conclusion is also consistent with the finding that there is no IFP when a 100% valid cue appears before the search display (Goolsby & Suzuki, 2001). That finding actually favors the notion that IFP modulates top-down rather than stimulus-driven attention, because the cue eliminates the need for top-down guidance to find the color singleton. Moreover, our finding is in line with other studies showing a reduction of IFP when the repeated feature is task irrelevant (Fecteau, 2007; Huang et al., 2004; Kristjánsson, 2006; Michal et al., 2014).
Finally, we note that our results do not contradict the general view that IFP involves attentional and/or perceptual processes. In fact, we found an IFP effect on both search accuracy and RTs, which indicates an effect on perceptual processing. Previous studies have also demonstrated such an IFP effect on accuracy with very short display durations (Asgeirsson et al., 2014;Yashar & Lamy, 2010a). However, we note that such demonstrations do not necessarily indicate an effect on saliency. Yashar and Lamy (2010a) have shown that the effect of IFP on accuracy emerges only when observers select and engage on the singleton target to discriminate its feature and not when observers had to localize only the singleton target position. Similarly, here we show that the color repetition effect emerges when observers engage on the target singleton (as in search trials) but not when attention is passively drawn to the singleton's location (as in probe trials).
Attentional mechanisms in probe discrimination performance
We designed our probe task to measure the effects of stimulus-driven spatial attention, induced by the color singleton, on perceptual sensitivity. We strove to minimize the role of higher-level decision processes and response preparation with two design features: First, we minimized spatial uncertainty by using a single probe stimulus with suprathreshold luminance contrast, making it easily localizable. (In Experiment 4, there was additionally a postcue that always indicated the probe's location.) Therefore, it is unlikely that the attentional effects in the probe task were due to reduction of location uncertainty on same-location trials or biased competition in memory (as there was only one probe stimulus). Second, we measured discrimination accuracy (with difficulty levels well below ceiling) in addition to response times. Previous studies investigating attentional capture and IFP usually measured only RTs as the main dependent variable. RTs can be influenced by criterion shifts or response preparation (e.g., Carrasco & McElree, 2001; Grubb, White, Heeger, & Carrasco, 2015; Wickelgren, 1977), and some researchers consider the distracting effect of irrelevant singletons on RTs to be a “filtering cost” rather than a true shift of spatial attention (e.g., Folk & Remington, 1998). In contrast, measuring the singleton's effects on probe discrimination accuracy enabled us to infer an effect of spatial attention on perceptual sensitivity.
We also must consider whether our relative location effects in the probe task are due to a purely stimulus-driven (i.e., exogenous) attentional mechanism. We borrowed the probe task design from the study by White et al. (2014), who isolated stimulus-driven attentional effects with several design choices. First, the color singleton was task irrelevant during all probe trials. As noted above, observers may not have been able to fully ignore the singletons on probe trials that followed intermixed search trials (in Experiments 1 and 2), but the relative location effect persisted in Experiments 3 and 4 when the probe trials were tested in blocks with only irrelevant singletons. Second, the singleton provided no information that could have enabled observers to produce the relative location effect voluntarily. Although the singleton location reduced the number of probe potential locations from four to two, the probe was equally likely to appear at the singleton's location or at the opposite side (the other hemifield). Thus, the predictability of the probe cannot account for the relative location effect. The fact that the singleton does not predict the probe hemifield also means that it could not bias eye movements toward the probe. Moreover, the time between the singleton onset and the probe onset was only 106 ms, too short for an endogenous shift of attention to the two possible probe locations before the probe appears (e.g., Liu, Stevens, & Carrasco, 2007; Nakayama & Mackeben, 1989). The short delay, the 50% chance that the probe would be diagonally opposite to the singleton, and the fact that the probe was high enough contrast to be easily localized on its own together minimized any incentive to attend to the singleton on probe trials and led us to conclude that the attentional effects are stimulus driven or exogenous.
This issue relates to an unresolved debate in the literature as to whether attentional capture by feature singletons is in fact purely stimulus driven (e.g., Theeuwes, 1994, 2010; Theeuwes & Chen, 2005) or dependent on the top-down attentional state (e.g., Bacon & Egeth, 1994; Egeth, Leonard, & Leber, 2010; Folk & Remington, 1998, 1999; Folk, Remington, & Johnston, 1992; Folk, Remington, & Wright, 1994; Lamy, Tsal, & Egeth, 2003; Yantis & Egeth, 1999). Our study cannot resolve this debate about “contingent capture,” in part because we did not test whether our relative location effect could be modulated in magnitude by changes in the top-down attentional state (White et al., 2014). Nonetheless, we do argue that bottom-up factors in our singleton displays were sufficient to drive a shift of spatial attention, which improved probe performance on same-location trials.
Conclusion
We found that intertrial repetition of a singleton's color does not modulate the stimulus-driven shift of attention toward the singleton. Nonetheless, manipulating singleton saliency via set size does modulate that spatial attention effect, and color repetition does facilitate singleton search with the same displays. We therefore argue against the hypothesis that IFP is due to an increase in the stimulus-driven saliency of repeated features. In addition, the effect of color repetition on search performance was stronger when the previous singleton had also been attended as a search target. Both results suggest that recent experience biases the voluntary selection of visual information, as in search, but not involuntary spatial selection, as in the relative location effect. Therefore, to the extent that the brain assumes a stable external world, it applies that assumption only to potentially task-relevant stimuli.
Acknowledgments
This study was supported by The Israel Science Foundation (Grant 111/15) to AY, NIH training T32 EY007136 to NYU (to ALW), and NIH Grant R01 EY016200 to MC. We would like to thank Rachel Denison, Ian Donovan, and Ricardo Max for their comments on a draft of this article.
Commercial relationships: none.
Corresponding author: Amit Yashar.
Email: amit.yashar@nyu.edu.
Address: Department of Psychology, New York University, New York, NY, USA.
Contributor Information
Amit Yashar, Email: amit.yashar@nyu.edu.
Alex L. White, alexlw@uw.edu, http://alexlwhite.com.
Wanghaoming Fang, Email: fangwan1@msu.edu.
Marisa Carrasco, marisa.carrasco@nyu.edu, https://sites.google.com/site/carrascolabnyu/.
References
- Amunts, L., Yashar, A., Lamy, D.. (2014). Inter-trial priming does not affect attentional priority in asymmetric visual search. Frontiers in Psychology, 5, 1– 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asgeirsson, A. G., Kristjánsson, A., Bundesen, C.. (2014). Independent priming of location and color in identification of briefly presented letters. Attention, Perception & Psychophysics, 76, 40– 48. [DOI] [PubMed] [Google Scholar]
- Awh, E., Belopolsky, A. V., Theeuwes, J.. (2012). Top-down versus bottom-up attentional control: a failed theoretical dichotomy. Trends in Cognitive Sciences, 16, 437– 443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bacon, W. F., Egeth, H. E.. (1994). Overriding stimulus-driven attentional capture. Perception & Psychophysics, 55, 485– 496. [DOI] [PubMed] [Google Scholar]
- Becker, S. I. (2008a). The mechanism of priming: Episodic retrieval or priming of pop-out? Acta Psychologica, 127 2, 324– 339. [DOI] [PubMed] [Google Scholar]
- Becker, S. I. (2008b). The stage of priming: Are intertrial repetition effects attentional or decisional? Vision Research, 48, 664– 684. [DOI] [PubMed] [Google Scholar]
- Belopolsky, A. V., Schreij, D., Theeuwes, J.. (2010). What is top-down about contingent capture? Attention, Perception, & Psychophysics, 72, 326– 341. [DOI] [PubMed] [Google Scholar]
- Bichot, N. P., Schall, J. D.. (2002). Priming in macaque frontal cortex during popout visual search: Feature-based facilitation and location-based inhibition of return. Journal of Neuroscience, 22, 4675– 4685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biderman, D., Biderman, N., Zivony, A., Lamy, D.. (in press). Contingent capture is weakened in search for multiple features from different dimensions. Journal of Experimental Psychology: Human Perception and Performance, in press. [DOI] [PubMed]
- Borji, A., Itti, L.. (2013). State-of-the-art in visual attention modeling. IEEE Transactions on Pattern Analysis and Machine Intelligence, 35, 185– 207. [DOI] [PubMed] [Google Scholar]
- Carrasco, M. (2011). Visual attention: The past 25 years. Vision Research, 51, 1484– 1525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrasco, M., McElree, B.. (2001). Covert attention accelerates the rate of visual information processing. Proceedings of the National Academy of Sciences, USA, 98, 5363– 5367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chun, M. M., Nakayama, K.. (2000). On the functional role of implicit visual memory for the adaptive deployment of attention across scenes. Visual Cognition, 7, 65– 81. [Google Scholar]
- Dienes, Z. (2011). Bayesian versus orthodox statistics: Which side are you on? Perspectives on Psychological Science, 6, 274– 290. [DOI] [PubMed] [Google Scholar]
- Duncan, J., Humphreys, G.. (1992). Beyond the search surface: Visual search and attentional engagement. Journal of Experimental Psychology: Human Perception and Performance, 18, 578– 588. [DOI] [PubMed] [Google Scholar]
- Egeth, H. E., Leonard, C. J., Leber, A. B.. (2010). Why salience is not enough: Reflections on top-down selection in vision. Acta Psychologica, 135, 130– 132, doi:10.1016/j.actpsy. 2010.05.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fecteau, J. H. (2007). Priming of pop-out depends upon the current goals of observers. Journal of Vision, 7 6: 1, 1– 11, doi:10.1167/7.6.1. [PubMed] [Article] [DOI] [PubMed] [Google Scholar]
- Fecteau, J. H., Munoz, D. P.. (2003). Exploring the consequences of the previous trial. Nature Reviews Neuroscience, 4, 435– 443. [DOI] [PubMed] [Google Scholar]
- Fecteau, J. H., Munoz, D. P.. (2006). Salience, relevance, and firing: A priority map for target selection. Trends in Cognitive Sciences, 10, 382– 390. [DOI] [PubMed] [Google Scholar]
- Folk, C. L., Remington, R.. (1998). Selectivity in distraction by irrelevant featural singletons: Evidence for two forms of attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 24, 847– 858. [DOI] [PubMed] [Google Scholar]
- Folk, C. L., Remington, R.. (1999). Can new objects override attentional control settings? Perception & Psychophysics, 61, 727– 739. [DOI] [PubMed] [Google Scholar]
- Folk, C. L., Remington, R. W.. (2008). Bottom-up priming of top-down attentional control settings. Visual Cognition, 16, 215– 231. [Google Scholar]
- Folk, C. L., Remington, R. W., Johnston, J. C.. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 18, 1030– 1044. [PubMed] [Google Scholar]
- Folk, C. L., Remington, R. W., Wright, J. H.. (1994). The structure of attentional control: Contingent attentional capture by apparent motion, abrupt onset, and color. Journal of Experimental Psychology: Human Perception and Performance, 20, 317– 329. [DOI] [PubMed] [Google Scholar]
- Fuller, S., Park, Y. Carrasco, M.. (2009). Cue contrast modulates the effects of bottom-up attention on appearance. Vision Research, 49, 1825– 1837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goolsby, B. A., Suzuki, S.. (2001). Understanding priming of color-singleton search: Roles of attention at encoding and “retrieval”. Attention, Perception, & Psychophysics, 63 6, 929– 944. [DOI] [PubMed] [Google Scholar]
- Grubb, M. A., White, A. L., Heeger, D. J., Carrasco, M.. (2015). Interactions between voluntary and involuntary attention modulate the quality and temporal dynamics of visual processing. Psychonomic Bulletin & Review, 22, 437– 444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hickey, C., Olivers, C., Meeter, M., Theeuwes, J.. (2011). Feature priming and the capture of visual attention: Linking two ambiguity resolution hypotheses. Brain Research, 1370, 175– 184. [DOI] [PubMed] [Google Scholar]
- Huang, L. Q., Holcombe, A. O., Pashler, H.. (2004). Repetition priming in visual search: Episodic retrieval, not feature priming. Memory & Cognition, 32, 12– 20. [DOI] [PubMed] [Google Scholar]
- Irons, J. L., Folk, C. L., Remington, R. W.. (2012). All set! Evidence of simultaneous attentional control settings for multiple target colors. Journal of Experimental Psychology: Human Perception and Performance, 38, 758– 775. [DOI] [PubMed] [Google Scholar]
- Kiss, M., Van Velzen, J., Eimer, M.. (2008). The N2pc component and its links to attention shifts and spatially selective visual processing. Psychophysiology, 45, 240– 249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kristjánsson, A. (2006). Simultaneous priming along multiple feature dimensions in a visual search task. Vision Research, 46 16, 2554– 2570. [DOI] [PubMed] [Google Scholar]
- Kristjánsson, Á., Saevarsson, S., Driver, J.. (2013). The boundary conditions of priming of visual search: From passive viewing through task-relevant working memory load. Psychonomic Bulletin & Review, 20, 514– 521. [DOI] [PubMed] [Google Scholar]
- Lamy, D., Antebi, C., Aviani, N., Carmel, T.. (2008). Priming of pop-out provides reliable measures of target activation and distractor inhibition in selective attention. Vision Research, 48, 30– 41. [DOI] [PubMed] [Google Scholar]
- Lamy, D., Kristjánsson, A.. (2013). Is goal-directed attentional guidance just intertrial priming? A review. Journal of Vision, 13 3: 14, 1– 19, doi:10.1167/13.3.14. [PubMed] [Article] [DOI] [PubMed] [Google Scholar]
- Lamy, D., Zivony, A., Yashar, A.. (2011). The role of search difficulty in intertrial feature priming. Vision Research, 51, 2099– 2109. [DOI] [PubMed] [Google Scholar]
- Lamy, D., Tsal, Y., Egeth, H. E.. (2003). Does a salient distractor capture attention early in processing? Psychonomic Bulletin & Review, 10, 621– 629. [DOI] [PubMed] [Google Scholar]
- Lee, H., Mozer, M. C., Vecera, S. P.. (2009). Mechanisms of priming of pop-out: Stored representations or feature-gain modulations? Attention, Perception, & Psychophysics, 71, 1059– 1071. [DOI] [PubMed] [Google Scholar]
- Liu, T., Stevens, S. T., Carrasco, M.. (2007). Comparing the time course and efficacy of spatial and feature-based attention. Vision Research, 47, 108– 113. [DOI] [PubMed] [Google Scholar]
- Maljkovic, V., Nakayama, K.. (1994). Priming of pop-out: I. Role of features. Memory & Cognition, 22 6, 657– 672. [DOI] [PubMed] [Google Scholar]
- Masson, M. E. (2011). A tutorial on a practical Bayesian alternative to null-hypothesis significance testing. Behavior Research Methods, 43, 679– 690. [DOI] [PubMed] [Google Scholar]
- McPeek, R. M., Maljkovic, V., Nakayama, K.. (1999). Saccades require focal attention and are facilitated by a short-term memory system. Vision Research, 39 8, 1555– 1566. [DOI] [PubMed] [Google Scholar]
- Meeter, M., Olivers, C. N. L.. (2006). Intertrial priming stemming from ambiguity: A new account of priming in visual search. Visual Cognition, 13, 202– 222. [Google Scholar]
- Meeter, M., Van der Stigchel, S.. (2013). Visual priming through a boost of the target signal: Evidence from saccadic landing positions. Attention, Perception, & Psychophysics, 75, 1336– 1341. [DOI] [PubMed] [Google Scholar]
- Michal, A. L., Lleras, A., Beck, D. M.. (2014). Relative contributions of task-relevant and task-irrelevant dimensions in priming of pop-out. Journal of Vision, 14 12: 14, 1– 12, doi:10.1167/14.12.14. [PubMed] [Article] [DOI] [PubMed] [Google Scholar]
- Morey, R. D. (2008). Confidence intervals from normalized data: A correction to Cousineau (2005). Tutorial in Quantitative Methods in Psychology, 4 2, 61– 64. [Google Scholar]
- Nakayama, K., Mackeben, M.. (1989). Sustained and transient components of focal visual attention. Vision Research, 29, 1631– 1647. [DOI] [PubMed] [Google Scholar]
- Posner, M. I., Snyder, C. R., Davidson, B. J.. (1980). Attention and the detection of signals. Journal of Experimental Psychology, 109, 160– 174. [PubMed] [Google Scholar]
- Raftery, A. E. (1999). Bayes factors and BIC. Sociological Methods & Research, 27, 411– 417. [Google Scholar]
- Theeuwes, J. (1994). Stimulus-driven capture and attentional set: selective search for color and visual abrupt onsets. Journal of Experimental Psychology: Human Perception and Performance, 20, 799. [DOI] [PubMed] [Google Scholar]
- Theeuwes, J. (2010). Top–down and bottom–up control of visual selection. Acta Psychologica, 135, 77– 99. [DOI] [PubMed] [Google Scholar]
- Theeuwes, J., Van der Burg, E.. (2013). Priming makes a stimulus more salient. Journal of Vision, 13 3: 21, 1– 11, doi:10.1167/13.3.21. [PubMed] [Article] [DOI] [PubMed] [Google Scholar]
- Theeuwes, J., Chen, C. Y. D.. (2005). Attentional capture and inhibition (of return): The effect on perceptual sensitivity. Attention, Perception, & Psychophysics, 67, 1305– 1312. [DOI] [PubMed] [Google Scholar]
- Thomson, D. R., Milliken, B.. (2011). A switch in task affects priming of pop-out: Evidence for the role of episodes. Attention, Perception, & Psychophysics, 73, 318– 333. [DOI] [PubMed] [Google Scholar]
- Töllner, T., Gramann, K., Müller, H. J., Kiss, M., Eimer, M.. (2008). Electrophysiological markers of visual dimension changes and response changes. Journal of Experimental Psychology: Human Perception and Performance, 34, 531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Townsend, J. T., Ashby, F. G.. (1983). Stochastic modeling of elementary psychological processes. Cambridge, UK: Cambridge University Press. [Google Scholar]
- Watson, A. B., Pelli, D. G.. (1983). Quest: A Bayesian adaptive psychometric method. Perception & Psychophysics, 33, 113– 120. [DOI] [PubMed] [Google Scholar]
- White, A. L., Lunau, R., Carrasco, M.. (2014). The attentional effects of single cues and color singletons on visual sensitivity. Journal of Experimental Psychology: Human Perception and Performance, 40, 639– 652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- White, A. L., Rolfs, M., Carrasco, M.. (2013). Adaptive deployment of spatial and feature-based attention before saccades. Vision Research, 85, 26– 35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- White, B. J., Munoz, D. P.. (2017). Neural mechanisms of saliency, attention, and orienting: Computational and cognitive neuroscience of vision. Zhao Q. (Ed.) Singapore: Springer Science+Business Media. [Google Scholar]
- Wickelgren, W. (1977). Speed accuracy tradeoff and information processing dynamics. Acta Psychologica, 41, 67– 85. [Google Scholar]
- Yantis, S., Egeth, H. E.. (1999). On the distinction between visual salience and stimulus-driven attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 25, 661– 676. [DOI] [PubMed] [Google Scholar]
- Yashar, A., Lamy, D.. (2010a). Intertrial repetition affects perception: The role of focused attention. Journal of Vision, 10 14: 3, 1– 8, doi:10.1167/10.14.3. [PubMed] [Article] [DOI] [PubMed] [Google Scholar]
- Yashar, A., Lamy, D.. (2010b). Intertrial repetition facilitates selection in time: Common mechanisms underlie spatial and temporal search. Psychological Science, 21, 243– 251. [DOI] [PubMed] [Google Scholar]
- Yashar, A., Makovski, T., Lamy, D.. (2013). The role of motor response in implicit encoding: Evidence from intertrial priming in pop-out search. Vision Research, 93, 80– 87. [PubMed] [Google Scholar]