

Abstract
Global climatic transitions and Tibetan Plateau uplifts are hypothesized to have profoundly impacted biodiversity in southeastern Asia. To further test the hypotheses related to the impacts of these incidents, we investigated the diversification patterns of the newt genus Tylototriton sensu lato, distributed across the mountain ranges of southeastern Asia. Gene-tree and species-tree analyses of two mitochondrial genes and two nuclear genes revealed five major clades in the genus, and suggested several cryptic species. Dating estimates suggested that the genus originated in the early-to-middle Miocene. Under different species delimitating scenarios, diversification analyses with birth-death likelihood tests indicated that the genus held a higher diversification rate in the late Miocene-to-Pliocene era than that in the Pleistocene. Ancestral area reconstructions indicated that the genus originated from the northern Indochina Peninsula. Accordingly, we hypothesized that the Miocene Climatic Transition triggered the diversification of the genus, and the reinforcement of East Asian monsoons associated with the stepwise uplifts of the Tibetan Plateau promoted the radiation of the genus in southeastern Asia during the Miocene-to-Pliocene period. Quaternary glacial cycles likely had limited effects on speciation events in the genus, but mainly had contributions on their intraspecific differentiations.
Keywords: Cryptic diversity, Radiation, Diversification rate, Climate shifts, Tibetan plateau, Tylototriton
Introduction
Southeastern Asia contains several biodiversity-hotspots (Myers et al., 2000). Processes promoting origin of biodiversity in this region have been explained by several mechanisms, including vicariances due to alterations of biogeographical configurations (Peng et al., 2006; Zhang et al., 2010), rapid radiations (Liu et al., 2006; Sun et al., 2012; Wang et al., 2012a; Wang et al., 2012b; Kennedy et al., 2012) and diversification rate shifts (Leneveu, Chichvarkhin & Wahlberg, 2009; Mao et al., 2010; Yu et al., 2014) associated with the growths of the Tibetan Plateau.
Despite arguments on timing and patterns of the uplifts of the Tibetan Plateau and adjacent regions (Harrison et al., 1992; Li & Fang, 1999; An et al., 2001; Sun & Wang, 2005; Molnar, Boos & Battisti, 2010; Yao et al., 2011), this process was unequivocally estimated to have occurred within the late Tertiary, and episodes of uplifts probably continued throughout the Pliocene and even Quaternary (Wan et al., 2007; Favre et al., 2014). The uplifts of the Tibetan Plateau had reinforced the East Asian monsoons (EAMs) several times, such as that about ∼15 Mya and ∼8 Mya (An et al., 2001; Wan et al., 2007; Yao et al., 2011). These incidents were proposed to have been promoted high levels of biodiversity transformations and radiations which created the present extraordinary high biodiversity in this region (Favre et al., 2014; Klaus et al., 2016).
Quaternary climatic oscillations have been singled out as another kind of historical forces in shaping species distributions and modulating local diversification (Hewitt, 2000). The effects of Pleistocene glaciations on promoting speciation, however, remain controversial. The refugium hypothesis (Haffer, 1969) indicates that speciation rates have increased because of vicariances among glacial refugia (Knowles, 2001; Knowles & Richards, 2005). On the contrary, the model of decreased diversification rates as a result of the increased extinction rates as related to the Pleistocene glaciations was postulated (Coope, 2004; Zink, Klicka & Barber, 2004). Additionally, the pattern with a constant diversification rate was proposed (Zink & Klicka, 2006) because diversification rates were also likely to be influenced by biotic factors (Thomas et al., 2010), ecological features (Weir & Schluter, 2007) or random episodes (Blount, Borland & Lenski, 2008). Yet in southeastern Asia, a number of evolutionary studies mainly reflect the contributions of the Pleistocene glacial cycles on the intraspecific lineage heterogeneity and revealed many local refugia for lineages especially in isolated mountains (e.g., Ding et al., 2011; Wang et al., 2012a; Wang et al., 2012b; Wang et al., 2013; Gu et al., 2013; Lin et al., 2014).
To explore diversification dynamics of a species group, inclusion of a high ratio of the total number of natural “true” species is the most important factor (Cusimano, Stadler & Renner, 2012). However, species diversity and taxonomy are still poorly known in many regions (e.g., the tropical region; Dubey & Shine, 2012). Taxonomic uncertainty may be due to inadequate biodiversity inventories, limited data to delimit taxonomic units and inconsistent criteria for species delimitation (Renner & Cusimano, 2010; Dubey & Shine, 2012). Many areas in southeastern Asia have not been thoroughly explored especially in the high-endemism-diversity regions, such as Himalayas, Hengduan Mountains and the tropical region of Indochina. In these areas, recently, a large number of new amphibian species were described (see Frost, 2016). Moreover, in many presumably well-described groups, molecular data indicated that many nominal species are not monophyletic (e.g., Tylototriton asperrimus complex; Yuan et al., 2011), and taxonomic reclassifications are needed. Even in a complete biodiversity inventory, different species concepts would produce different sets of taxonomic units (e.g., the Gynandropaa complex; Zhang et al., 2010; Fei, Ye & Jiang, 2012). Obviously, taxonomic uncertainty may influence the reconstructions of diversification patterns. Therefore, we need to examine the role of taxonomic uncertainty on exploring diversification patterns.
The newt genus Tylototriton sensu lato (s.l.) Anderson, 1871 (Salamandridae, Caudata) currently contains 24 nominal species (Fei, Ye & Jiang, 2012; Zhao et al., 2012; Frost, 2016), and is distributed across southeastern Asia. Tylototriton s.l. was supported as a monophyletic group in previous phylogenetic studies (e.g., Nishikawa et al., 2013; Nishikawa, Matsui & Nguyen, 2013). But based on morphology, this group has ever been divided into three genera, i.e., Tylototriton sensu stricto, Yaotriton and Liangshantriton (Fei, Ye & Jiang, 2012). Twelve Tylototriton s.l. species have been described in the past five years (Zhao et al., 2012; Shen, Jiang & Mo, 2012; Hou, Li & Lu, 2012; Nishikawa et al., 2013; Nishikawa, Matsui & Nguyen, 2013; Nishikawa, Matsui & Rao, 2014; Yang et al., 2014; Khatiwada et al., 2015; Phimmachak, Aowphol & Stuart, 2015). But within all nominal species, T. daweishanensis was argued to be synonym of T. yangi (Nishikawa et al., 2015), and T. shanjing was argued to be synonym of T. verrucosus (Zhang et al., 2007). These inconsistencies indicate that deep phylogenetic investigations in the group were required to investigate the relationships within Tylototriton to resolve species relationships and uncover any cryptic diversity within this group using multiple genes. Additionally, most Tylototriton s.l. species are found to be distributed allopatrically throughout the Asian monsoon climate zone (Frost, 2016), with a preference for humid environments, and usually inhabit and breed near or in ponds in the forest because many of them have limited capability on dispersal capability (Fei, Ye & Jiang, 2012). Accordingly, it is presumed that their diversification may have been easily impacted by historical climatic changes. Therefore, this genus is an ideal model to examine the effects of past climatic shifts on the diversification in southeastern Asia.
We hypothesize that the past climatic changes, especially the reinforcement of the East Asian monsoons (EAMs) associated with the stepwise uplifts of the Tibetan Plateau during the late Tertiary and Quaternary glacial oscillations, drove the speciation diversification of Tylototriton s.l.. Thus, in this study, to test the hypotheses, based on the sampling of all 24 nominal Tylototriton s.l. species and molecular data of mitochondrial and nuclear genes, we reconstruct species phylogenetic relationships and infer a time frame and diversification patterns for the Tylototriton s.l. newts.
Materials and Methods
Sampling and sequencing
A total of 108 samples was used, representing all 23 recognized species (we did not use “T. daweishanensis” because it is assigned as synonym of T. yangi) of Tylototriton s.l., which were collected from 49 localities scattering across the range of the genus in the southeastern Asia (Fig. 1A; Table S1). Based on previous studies (Steinfartz et al., 2007; Zhang et al., 2008), 23 species representing major divisions within Salamandridae were included in our phylogenetic analyses, and one Ambystoma mexicanum was used as outgroup (Table S1). In these 23 Salamandridae species, two specimens of Echinotriton chinhaiensis (Zhang et al., 2008) were collected in this study. The Animal Care and Use Committee of Chengdu Institute of Biology, CAS provided full approval for this purely observational research (Number: CIB2010031015). Field experiments were approved by the Management Office of the Kuankuoshui Nature Reserve (number: KKSNR201204002) and the Management Office of the Dabashan Nature Reserve (number: DBSNR201204002).
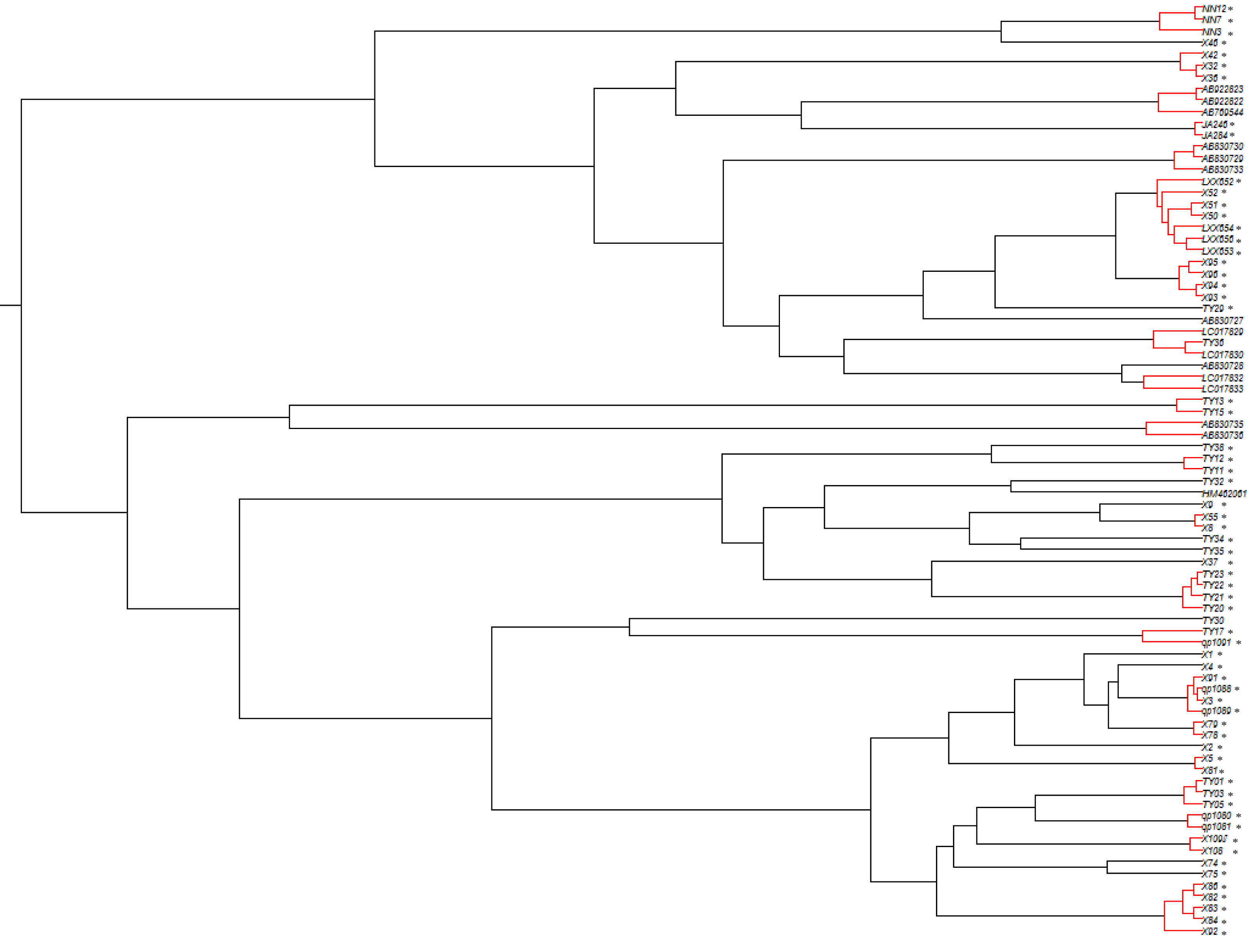
Figure 1. Sampling localities (A) and Maximum likelihood (ML) tree obtained based on mtDNA data of Tylototriton s.l. and relatives (B).

Sample number 1–108 refer to Table S1. Five clades (I, II, III, IV and V) of the genus were denoted as different colors. Bootstrap supports (bs) resulted from ML analyses and posterior probability (pp) resulted from Bayesian inference (BI) method were labeled on major nodes. Node supports ML1 and BI1 were resulted from analyses on mtDNA data, while ML2 and BI2 were from analyses on four-gene concatenated data. Black: bs > 70% or pp > 0.95, grey: bs = 50–70% or pp = 0.85–0.95, white: bs < 50% and pp < 0.85.
The genomic DNA was extracted with the Qiagen DNeasy tissue kits (Qiagen, Hilden, Germany) from tissues preserved in 95% alcohol. Fragments of two mitochondrial genes, i.e., 16S and ND2, and two nuclear genes, NCX1 and BDNF, were amplified for all samples collected in this work. Primers for amplifying ND2, NCX1 and BDNF genes were designed in this study (Table S2). The polymerase chain reaction (PCR) cycles included an initial denaturation step of 4 min at 94 °C and 35 cycles of denaturation for 30 s at 94 °C, primer annealing for 30 s at 48–58 °C, and extension for 1 min 30 s at 72 °C. Sequencing was performed in both directions using the primers in PCR on ABI3730 sequencer. New sequences were deposited in GenBank (accession numbers were shown in Table S1). ND2 sequences of 28 specimens were retrieved from GenBank (see Table S1), and ND2 sequences of two T. yangi samples were from Zhao et al. (2012).
Alignments were performed in the program MAFFT v.6 (Katoh et al., 2002) with default options. To avoid artificial bias in refining alignments, GBLOCKS v.0.91.b (Castresana, 2000) with default settings was used to extract regions of defined sequence conservation from the length-variable 16S fragment. DNA sequences of each protein-coding gene were translated into amino acid sequences, and then the amino acid sequences of each gene were aligned in MEGA v.6 (Tamura et al., 2013). The nucleotide gaps were determined according to the positions of amino-acid gaps in the alignments of the amino acid sequences. Non-sequenced fragments were defined as missing loci. Recombination in the nuclear markers was tested in the program RDP3 (Martin et al., 2010) with default setting. The test indicated that there was no recombination in the nuclear markers.
Phylogenetic analyses
Gene trees were reconstructed just for the mitochondrial DNA (mtDNA) data and four-gene concatenated data, respectively, because there were few variable sites in nuclear gene data for reconstructing reliable nuclear gene trees. Maximum likelihood (ML) and Bayesian Inference (BI) analyses were conducted in the programs RAXML v.8.1.24 (Stamatakis, 2014) and MRBAYES v.3.2.6 (Ronquist et al., 2012), respectively. To avoid under- or over-parameterization (Lemmon & Moriarty, 2004; McGuire et al., 2007), the best partition scheme and the best evolutionary model for each partition were selected for the phylogenetic analyses using the program PARTITIONFINDER v.1.1.1 (Lanfear et al., 2012). For PARTITIONFINDER analyses, 16S gene and each codon position of each protein-coding gene was defined and the “greedy” search where branch length estimates were unlinked under the Bayesian Inference Criteria (BIC) as recommended by Lanfear et al. (2012) was performed. In ML analyses, the best tree was produced from 1,000 iterations of the random addition of taxa. For the ML tree, branch supports were drawn from 10,000 non-parametric bootstrap replicates. Four-gene concatenated data was analyzed with joint branch optimization to minimize the impact of missing loci on branch lengths. For BI analyses, gaps were scored as presence/absence data in the program GAP-CODER (Young & Healy, 2003) using the procedure of (Simmons, Ochoterena & Carr, 2001). In BI analyses, the parameters for each partition were unlinked, and branch lengths were allowed to vary proportionately across partitions. Two independent runs were initiated each with four simultaneous Markov Chain Monte Carlo (MCMC) chains for 100 million generations and sampled every 1,000 generations. Convergence of chains and burn-in periods were determined using the program TRACER v.1.6 (Rambaut et al., 2014). Then the burn-in samples (the first 25% defined by Tracer) were discarded, and then a maximum bifurcating consensus tree and posterior probability (pp) were derived from the remaining samples.
To explore the possibility that the unlinked gene trees might be incongruent among each other or with the species tree due to processes such as hybridization, incomplete lineage sorting, horizontal gene transfer, or gene duplication (Maddison, 1997; Kubatko & Degnan, 2007; Degnan & Rosenberg, 2009; Edwards & Beerli, 2000), the species tree was inferred using multispecies coalescent inference. The analyses were conducted using the *BEAST option (Heled & Drummond, 2010) in the program BEAST v.2.3.0 (Bouckaert et al., 2014), as described in the following section together with dating estimations.
Dating analyses
Divergence times were estimated also based just on mitochondrial data and four-gene concatenated data, respectively, because there were few variable sites in nuclear gene data for reconstructing reliable nuclear gene trees. Time-calibrated phylogenies were reconstructed by incorporating fossil calibration points using BEAST. Four Salamandridae fossil records were used as calibration points. The first calibration point was the divergence between Tylototriton s.l. and Pleurodeles for which the minimum age was set to 44 Mya, based on a Tylototriton-related salamandrid fossil, Chelotriton (Herre, 1935; Estes, 1981; Milner, 2000). The relevant fossil of Brachycormus was not used because it was alleged to have been a neotenic amphibian or a derivate of Chelotriton (Roček, 1996a; Roček, 1996b). Secondly, the minimum age of ancestor of Taricha and Notophthalmus was presumed to be 22 Mya based on a fossil of Taricha oligocenica (Estes, 1981). Then the minimum age of the common ancestor of Triturus was given as 24 Mya based on a Triturus fossil (Böhme, 2003). Finally, based on the fossil of Procynops miocenicus closely related to Cynops (Estes, 1981), we presumed the minimum divergence time between Cynops and Paramesotriton at ∼15 Mya. A lognormal prior distribution was applied for each calibration point in view of the fact that this model allowed for uncertainty of the age estimation of the fossil and bias associated with the incompleteness of the fossil record (Forest, 2009).
For BEAST analyses, to avoid zero-length branches, each coalescent cluster was represented by one specimen. So the Generalized Mixed Yule Coalescent (GMYC) method (Pons et al., 2006; Fontaneto et al., 2007) was used to identify coalescent clusters within Tylototriton s.l. The GMYC method was proposed to potentially promote objective delimitation of coalescent clusters even based on single gene and supplies explanations for divergences between clusters by speciation models (Powell, 2012). GMYC analysis was implemented in the R package SPLITS (Species Limits by Threshold Statistic) using the single-threshold option. Because the GMYC analysis needs a time-scale tree, a preliminary BEAST analysis was conducted only based on mtDNA data set with the best partitioning scheme and the best model for each partition selected above. In this analysis, a conservative parameter setting (i.e., a relaxed lognormal clock with the mean rate set to 1 and a coalescent Yule model as tree prior) was assigned (Monaghan et al., 2009). MCMC chains were run for 20 million generations with sampling every 1,000 generations, and the first 25% generations were discarded as burn-in. GMYC analysis resulted in 39 cluster representatives (see ‘results’), which were used for the following analyses.
For BEAST analyses, the best partition scheme and the best evolutionary model for each partition were also selected for the phylogenetic analyses using the program PARTITIONFINDER. An unlinked uncorrected lognormal relaxed clock to each partition, a Yule speciation process for branching rates, and the model of four rate categories for nucleotide substitutions were defined for the analyses. A ML tree was used as a starting tree inferred by RAXML based on single-GMYC-cluster representatives. Two runs of 100 million generations sampling every 1,000 generations were conducted. The convergence of MCMC chains and burn-in periods were determined by Tracer. A maximum clade credibility (MCC) tree with posterior probability was calculated using TREEANNOTATOR.
Species tree was inferred using *BEAST. The method needs prior assignments of specimens to species. So the selection of specimens for *BEAST analyses was according to the GMYC clusters and the sampling number (>1) of each cluster. GMYC singletons and individuals with missing data genes were excluded. Because we obtained sequences of nuclear genes only for 91 Tylototriton s.l. samples and two E. chinhaiensis samples, outgroup fossil calibrations (see above) could not be utilized for the *BEAST analysis. So, the nucleotide substitution rate of each partition was set according to the rate estimations from the four-gene concatenated Beast analyses conducted above. For *BEAST analysis, mitochondrial DNA (16S + ND2) and each nuclear gene was defined as an independent partition, respectively. The best-fitting substitution model for each partition was selected by the program JMODELTEST v.2.1 (Guindon & Gascuel, 2003; Darriba et al., 2012). *BEAST analyses were carried out using the unlinked uncorrected lognormal relaxed clock to each partition and a Yule speciation process as tree prior. Lognormal prior distributions were assigned for mean rate of each partition. Two runs of 150 million generations were carried out with sampling every 1,000 generations. Convergence of MCMC chains and burn-in periods were assessed using TRACER.
Topology of the *BEAST tree with fewer number of species was almost consistent with the BEAST tree except for the position of one major clade (see ‘results’). Therefore, to produce a “complete species-time-tree”, a final topology-constrained BEAST analysis was conducted based on mtDNA of GMYC cluster representatives and all outgroups by applying a constraint of the topology as the *BEAST species tree. The fossil calibrations and parameter-settings were specified as constant as the mtDNA-BEAST analysis above.
Ancestral area reconstructions
Ancestral area was reconstructed using the statistical dispersal-vicariance analysis (S-DIVA; Yu, Harris & He, 2010) and Bayesian Binary MCMC (BBM) all implemented in the program RASP v.3.2 (Yu, Harris & He, 2012). According to the zoogeographic regions defined by Fei, Ye & Jiang (2010) using amphibian data, the distribution range of Tylototriton s.l. could be delineated into four regions: A, Northern Indochina Peninsula and South Yunnan Province of China (NIP-SY); B, Southwest China mountains and Eastern Himalaya mountains (SCM-EHM); C, Coastal areas of Southern China region and Hainan Island (CSC-HI); D, Central China region (CC).
Ancestral area reconstructions were based on the final 39-GMYC-cluster MCC tree (see ‘results’) and randomly sampled 2,000 post-burn-in MCMC trees resulted from the final topology-constrained BEAST analysis above. The maximum number of ancestral areas was restricted to two based on distributional suggestions for each species or species group (Fei, Ye & Jiang, 2012) and preliminary analyses comparing the S-DIVA values, analogous to the fit of the data to the area model, of n = 2, 3 and 4. The dispersal event was restricted to adjacent regions. The BBM analysis was conducted for 500,000 generations of 10 chains at a temperature of 0.1. Fixed state frequencies of the JC model, equal rate variation among sites and a wide root distribution were specified, and null distribution and distributions outside target regions were prohibited in any reconstructions.
Diversification analyses
Diversification analyses are dependent on the number of user-specified biodiversity units. Yet there is controversy on the criterion to delimit species-level lineages. To evaluate how robust models of diversification were to variation of number of species-level units, phylogenetic lineages were delimited according to three criteria: GMYC clusters (statistically inferred coalescent lineages), currently recognized nominal species (NS) and NS plus (NSP: NS plus independent lineages representing putative cryptic species, for example, T. wenxianensis lineage 2 and 3, see ‘results’). Two thousand trees were randomly sampled from the post-burn-in posterior trees resulted from the final topology-constrained Beast analysis. Then, MCC tree and 2,000 sampled posterior trees were pruned according to each lineage delimitating strategy. The number of lineages varied when we used different units of biodiversity: 39 GMYC-clusters, 23 NS, and 27 NSP (see results). Finally, the pruned 39-tips, 23-tips and 27-tips trees were used for diversification analyses.
To visualize the temporal accumulation of lineages, the log-transformed lineage-through-time (LTT) plots were constructed for each lineage delimitating strategy. To test if the time-lag between the origin and divergence of extant Tylototriton s.l. departed from random expectations under the birth-death model, simulations (n = 10,000) under birth-death process were run using five different sampling fractions: 100%, 90%, 80%, 70% and 50%, respectively. Parameters for the simulations included the speciation and extinction rates, the age of the stem group and the number of extant lineages all estimated from the observed data based on the birth-death model. Simulations were carried out in the R (R Development Core Team, 2008) package TREESIM (Stadler, 2011). LTT plots of the simulated trees were then constructed to compare with the average LTT plot of the empirical data.
To examine diversification rate shifts, the birth-death likelihood (BDL) test (Rabosky, 2006) was carried out in R packages LASER (Rabosky, 2006) and GEIGER (Harmon et al., 2008). In BDL analysis, six models (pure birth, birth-death, yule2rate, DDL (diversity-dependent linear decrease) and DDX (diversity-dependent exponential decrease)) were fitted to the observed data under the Akaike’s information criterion (AIC). The best model was selected by calculating the difference (ΔAIC) in AIC scores between the best rate-variable (RV) models and the best rate-constant (RC) models (Rabosky, 2006). The significance of the observed ΔAICRC statistic was assessed by one-tailed tests (Rabosky & Lovette, 2008a). Additionally, other three diversification models were also tested. SPVAR permits speciation rate to vary over time, EXVAR permits extinction rate to vary over time, and BOTHVAR allows both to vary over time (Rabosky & Lovette, 2008b). Finally, Akaike weights (Turkheimer, Hinz & Cunningham, 2003) were calculated for each model.
Results
Phylogenetic assignments and dating estimation
A total of 133 samples was included, and 108 individuals representing all currently nominal species of Tylototriton s.l. (Table S1). Alignments resulted in 1,035 bps for ND2, 501 bps for NCX1 and 648 bps for BDNF, and GBLOCKS analysis resulted in 433 bps for 16S. In the ND2 alignment, there were few numbers of gaps existing in several outgroup species.
ML and BI analyses both supported the monophyly of the genus Tylototriton s.l., and all revealed five major clades in it (Fig. 1B). ML and BI analyses supported the relationships among clades as ((I, II), (III, IV, V)). Note that, the presently so-called “T. asperrimus”, “T. wenxianensis” and “T. dabienicus” taxa were not resolved as monophyletic, and contained two, three and two polyphyletic lineages, respectively, which were indicated as containing cryptic species.
GMYC analysis resulted in 39 independent coalescent clusters within Tylototriton s.l. (P < 0.01), consisting of 15 singletons and 24 clusters (Fig. S1). After excluding GMYC singletons and individuals with missing data genes, 19 GMYC clusters were used for species-tree analyses. Species tree had almost consistent framework with the gene trees except for one instance of incongruence, i.e., T. vietnamensis representing Clade III was clustered at the basal position of the genus in species tree, but in gene trees, this relationships was not resolved (Figs. 1 and 2).
Figure 2. Time-calibrated gene trees and species tree of Tylototriton s.l.

(A) BEAST gene tree of mtDNA; (B) BEAST gene tree of four-gene concatenated data; (C) species tree. Node number was denoted near each node. Node 2* in gene trees and node 2 in species tree had different topology. Bayesian posterior probabilities (pp) were denoted on nodes. Black circles: pp = 0.95–1.0; open circles: pp < 0.95.
The chronograms inferred from the gene-tree and species-tree approaches based on different data sets presented similar divergence time estimates (Table 1; Figs. 2 and 3). All dating estimations recovered a middle Miocene-to-present timeframe for Tylototriton s.l. (Table 1; Figs. 2 and 3; Fig. S2). Generally, the mean nodal ages estimated using the species-tree approach were little younger than the estimations resulted from the gene-tree approaches, although their 95% highest posterior densities (HPD) overlapped for each node. Except for the divergence of T. dabienicus lineage 1 and T. wenxianensis lineage 3, and that of T. shanjing and T. verrucosus, speciation of all nominal species basically predated the Pleistocene.
Table 1. Mean node age estimates and 95% highest posterior densities obtained from gene-tree (BEAST) and species-tree approaches (*BEAST).
The difference between nodes 2 and 2* refer to Fig. 2.
| Node | mtDNA Beast | Four-gene concatenated Beast | *BEAST | Topology-constrained Beast |
|---|---|---|---|---|
| 1 | 14.1 (11.8–16.6) | 14.6 (12.8–16.7) | 13.7 (10.4–17.5) | 14.4 (12.0–17.0) |
| 2 | / | / | 11.3 (8.8–14.3) | 13.7 (11.6–16.2) |
| 2* | 12.5 (10.4–14.8) | 12.9 (11.2–14.7) | / | / |
| 3 | 10.4 (8.2–12.6) | 10.3 (8.7–12.1) | 8.0 (6.1–10.2) | 10.0 (8.1–12.1) |
| 4 | 2.6 (1.4–3.9) | 2.7 (1.7–3.7) | 2.3 (1.2–3.5) | 2.6 (1.4–4.0) |
| 5 | 7.6 (6.2–9.4) | 7.5 (6.4–8.7) | 5.9 (4.5–7.5) | 7.6 (6.1–9.2) |
| 6 | 6.6 (4.9–8.3) | 6.3 (5.1–7.5) | 5.1 (3.5–6.7) | 6.4 (4.9–8.1) |
| 7 | 5.1 (3.5–6.8) | 4.9 (3.7–6.2) | / | 5.0 (3.5–6.6) |
| 8 | 5.7 (4.4–7.1) | 5.6 (4.6–6.5) | / | 5.6 (4.4–7.0) |
| 9 | 5.0 (3.7–6.3) | 5.3 (3.8–5.6) | / | 5.2 (4.0–6.6) |
| 10 | 4.0 (2.7–5.3) | 4.7 (4.4–6.2) | / | 4.4 (3.2–5.7) |
| 11 | 3.4 (2.3–4.7) | 3.5 (2.6–4.4) | / | 3.5 (2.4–4.8) |
| 12 | 2.7 (1.7–3.7) | 2.7 (2.0–3.6) | 2.4 (1.5–3.4) | 2.7 (1.7–3.8) |
| 13 | 1.0 (0.5–1.6) | 1.2 (0.7–1.8) | 1.2 (0.6–1.8) | 1.0 (0.5–1.7) |
| 14 | 11.6 (9.5–13.8) | 11.9 (10.3–13.7) | 10.1 (7.7–12.7) | 12.4 (10.1–14.7) |
| 15 | 6.0 (4.7–7.5) | 6.1 (5.1–7.2) | 4.5 (3.0–6.5) | 6.2 (4.8–7.8) |
| 16 | 5.3 (4.1–6.5) | 5.6 (4.6–6.6) | / | 5.4 (4.3–6.8) |
| 17 | 3.3 (2.2–4.6) | 3.6 (2.6–4.6) | 3.0 (1.8–4.3) | 3.4 (2.2–4.7) |
| 18 | 4.5 (3.4–5.8) | 4.9 (4.0–5.9) | / | 4.7 (3.5–6.0) |
| 19 | 2.8 (2.0–3.8) | 3.1 (2.4–3.9) | / | 2.9 (2.0–3.9) |
| 20 | 8.3 (6.5–10.3) | 8.7 (7.3–10.3) | 7.6 (4.8–9.4) | 8.6 (6.7–10.7) |
| 21 | 6.2 (4.4–8.2) | 6.6 (5.2–8.1) | / | 6.4 (4.5–8.4) |
| 22 | 4.1 (3.2–5.1) | 4.2 (3.4–5.1) | 3.8 (2.8–5.0) | 4.2 (3.3–5.3) |
| 23 | 3.3 (2.4–4.2) | 3.3 (2.6–4.0) | 3.2 (2.3–4.3) | 3.4 (2.5–4.3) |
| 24 | 3.0 (2.3–3.9) | 3.0 (2.4–3.8) | 3.1 (2.2–4.1) | 3.1 (2.3–4.1) |
| 25 | 2.7 (1.9–3.5) | 2.7 (2.1–3.4) | 2.6 (1.6–3.7) | 2.4 (2.0–3.7) |
| 26 | 1.9 (1.2–2.7) | 2.1 (1.5–2.7) | 1.9 (1.0–2.8) | 2.0 (1.3–2.8) |
| 27 | 3.1 (2.2–4.0) | 3.2 (2.5–4.0) | / | 3.2 (2.3–4.1) |
| 28 | 10.0 (7.7–12.6) | 10.5 (8.6–12.6) | / | 11.5 (8.7–14.4) |
Figure 3. Chronogram and ancestral area reconstructions of Tylototriton s.l.

This tree was pruned from the final topology-constrained BEAST tree. Grey bars at nodes show 95% highest posterior density intervals of divergence times. Node number was denoted near each node. Black circles, Bayesian posterior probabilities (pp) = 0.95–1.0; open circles, pp < 0.95. Clades I V refer to Fig. 1. Two vertical bars on branch showed the relative probabilities of alternative ancestral distributions obtained by the S-DIVA (first bar) and BBM (second bar), and the first two areas with the highest probability are shown on each bar corresponding to relative probability. Four significant dispersals were highlighted near corresponding nodes using arrows linking the originated region to the dispersed region. (insert) four regions was denoted as different color: (A) Northern Indochina Peninsula and South Yunnan Province of China; (B) Southwest China mountains and Eastern Himalaya mountains; (C) Coastal areas of Southern China region and Hainan Island; (D) Central China region.
Ancestral areas
Ancestral area reconstructions from S-DIVA and BMM analyses based on the “complete species-time” Beast trees were largely consistent (Fig. 3). The probability resulting from two methods for the occurrence of each node was greater than 80%. Bothe methods indicated NIP-SY as the ancestral area of Tylototriton s.l. Similarly, the ancestors of Clades II, III and IV were also indicated as originating from NIP-SY. Only Clade I originated from SCM-EHM and Clade V from CC. At least four dispersal events in the late Miocene were supported (Fig. 3). During the middle Miocene ∼12.5 Mya, a dispersal event from NIP-SY to CC was at the origin of the formation and expansion of Clade V. During the middle Miocene ∼10 Mya, a dispersal event from NIP-SY to SCM-EHM was at the origin of the formation of high plateau endemic species (i.e., T. taliangensis and T. pseudoverrucosus). Then the dispersal from NIP-SY to SCM-EHM during the late Miocene ∼7.5 Mya can explain the speciation of T. kweichowensis widely distributed in the Guizhou plateau of China. Finally, a dispersal event from NIP-SY to CSC-HI during the late Miocene can explain the formation of species presently living in the South China.
Diversification analyses
LTT plots for 39-GMYC-cluster, 23-NS and 27-NSP data were presented in Fig. 4. The appearance variations of LTT plots for different lineage delimitating dispositions were mainly derived from variations of the number of terminal tips. Using different undersampling scenarios, 10,000 trees were simulated for different lineage delimitating strategies on the observed number of extant species (n = 39, 27 and 23) and the observed fixed time since origin (stem age = 22.46 Ma) under the birth-death process (speciation rates and extinction rates for each delimitating strategies were shown in Fig. S3). Under full sampling or more than 50% sampling fraction scenarios, the observed data did not depart from random expectations (Fig. S3).
Figure 4. Lineage through time plots based on different lineage delimitating strategies.

Different lineage delimitating strategies: (A) 39 statistically inferred coalescent lineages using the Generalized Mixed Yule Coalescent method (GMYC); (B) 23 currently nominal species (NS); and (C) 23 NS plus four independent lineages representing putative cryptic species (NSP). All trees were pruned from the final topology-constrained BEAST analysis. The maximum credibility tree was denoted by a thick black line, and 2,000 random trees from the posterior distribution were denoted by thin grey lines.
BDL tests based on AIC scores resulted in positive ΔAIC values (3.10868 for 39-GMYC-cluster data, 5.03466 for 23-NS data, and 8.98177 for 27-NSP data) being significantly departed from the null hypothesis of constant rate model (all P < 0.03). BDL tests (Table S3; Fig. 5) indicated the yule2rate model as the best-fitting model for the 39-GMYC-cluster data and 27-NSP data, but the DDL model as the best-fitting model for the 23-NS data. Yet the yule2rate was selected as the second fitting model for the 23-NS data, as well as the DDL model was selected as the second fitting model for the 39-GMYC-cluster and 27-NSP datasets. For 39-GMYC-cluster data, the diversification rates have sharply decreased around 1 Mya, but for 27-NSP data and 23-NS data, it was at the beginning of Pleistocene, ∼2.6 Mya (Table S3).
Figure 5. Information theory approach comparing best-fitting diversification rate models.

Shown are the Akaike weights of eight diversification rate models (pure birth, birth death, DDL, DDX, yule2rate, SPVAR, EXVAR, and BOTHVAR) and data sets defined by different lineage delimitating strategies: GMYC, 39 statistically inferred coalescent lineages using the Generalized Mixed Yule Coalescent method; NS, 23 currently nominal species; and NSP, 23 NS plus four independent lineages representing putative cryptic species.
Discussion
Considerations for uncertainties and cryptic diversity
Our results on the divergence times of the genus Tylototriton s.l. and salamandrid major clades depend on the calibrations that we used for estimating these times (Fig. S2). In this case, previous phylogenetic investigations on Salamandridae using these fossils also proposed broadly consistent timeframes for its major clades (Steinfartz et al., 2007; Zhang et al., 2008; Shen et al., 2016). Also for Tylototriton s.l., using these fossil calibrations, dating estimates based on complete mitochondrial genome data (Kurabayashi et al., 2012) were largely consistent with our results based on incorporating data of mitochondrial and nuclear genes (Figs. 2 and 3).
Alternatively, because of some methodological flaw, the dating estimations may be argued to be overestimated. For example, actual divergence times were tended to be overestimated by gene trees because a fraction of gene divergence may pre-date the species divergences (Edwards & Beerli, 2000; Arbogast et al., 2002), and meanwhile, dating time may be underestimated by species-tree approaches in some works due to the uncounted presence of gene flow among lineages (McCormack et al., 2010). In our study, divergence times resulted from gene-tree and species-tree approaches did not differ substantially (Figs. 2 and 3). Similar findings were proposed in many animal groups, such as true crocodiles (Oaks, 2011), iguanian lizards (Townsend et al., 2011), and spiders (Bidegaray-Batista, Ferrández & Arnedo, 2014). Certainly, using large number of unlinked nuclear loci may increase the accuracy of species-tree estimations (Heled & Drummond, 2010). In this study, use of only two nuclear genes could be argued as providing not enough information to obtain “true” time-tree. But in another sense, a low number of nuclear loci combined with mitochondrial genes has the possibility of similar effects on dating estimations as the use of more nuclear markers (Sánchez-Gracia & Castresana, 2012). Tylototriton s.l. has likely diversified during a large timescale, i.e., middle Miocene-to-Pleistocene era (Table 1), and therefore, major conclusions could be considered broadly.
Testing diversification rate shifts is considered to be sensitive to species delimitation in a phylogeny (Smith et al., 2013; Renner & Cusimano, 2010). For the genus Tylototriton s.l., either based on biological species or statistically inferred species, BDL tests selected the rate-slowdown model as the best-fitting diversification model. Moreover, different lineage delimitating strategies likely just impacted the terminal characteristics of lineage accumulation (Fig. 4), indicating that our sampling of the “old” clades might be sufficient. This point was verified by the tree-simulation method, which suggested that the simulated data did not depart from the observed data just until under high undersampling scenarios (≥ 50%; see Fig. S3). Although species diversity of the genus will presumedly increase, the rate-slowdown model is still solidly based. It is expected that speciation time of the foreseeable newly-found species less likely will all fall in the recent Pleistocene period but may well spread over several epochs, just like the time of origin of many recently recognized species (e.g., T. liuyangensis, T. panhai, T. ziegleri and T. yangi; Fig. 3). Even for the independent lineages that were indicated as putative cryptic species, the dating estimates also suggested that origin of them generally predated the Pleistocene (Fig. 3). On the other side, if it is assumed that our phylogenetic framework has sampled all substantial species, and then we truncated potential synonyms of several species, such as “T. shanjing” sometimes indicated as T. verrucosus by phylogeographical investigations (Zhang et al., 2007; Khatiwada et al., 2015) and “T. dabienicus” previously classified as “T. wenxianensis dabienicus” (Chen, Wang & Tao, 2010), our empirical data will more closely resemble the declining rate model because the time of origin of these “redundant” lineages all fall in Pleistocene (Fig. 3).
Of course, evolution of Tylototriton s.l. has also been probably confined by other potential factors, such as density-dependent cladogenesis (Rabosky & Lovette, 2008a). Our diversification analyses selected the DDL model as the best-fitting model for 23-NS data, and selected the model as the second fitting model for 39-GMYC-cluster and 27-NSP data (Fig. 5; Table S3). Density-dependent cladogenesis also usually leads to early burst of diversification in old epochs and rate-slowdown appearances in the recent period (Phillimore & Price, 2008). In the DDL model, speciation slows down often since ecological opportunities and geographical space place limits on clade growth (Phillimore & Price, 2008). In regard of Tylototriton s.l., most species prefer humid habitats in mountain forests, and their populations usually show high local endemism (Li, Zou & Luo, 2011; Fei, Ye & Jiang, 2012), indicating that Tylototriton s.l. newts probably have strict ecological selection, and niches of most species were probably narrow and easily filled. Presumably, in the Miocene-to-Pliocene epoch, the genus had slowly radiated in the southeastern Asia and increasing number of species had probably slowly filled the niches, and then in recent era, species biodiversity of the genus has been under niche restrictions leading to less speciation. Also, in the present “colder” epoch rather than in the early Miocene (Böhme, 2003), the newts had a low capacity to persist in new locations, most likely preventing populations from evolving into species.
Additionally, the results would be cautiously interpreted for lack of external information about extinction (Rabosky, 2010). Our results indicated that Tylototriton s.l. had occupied a long-term constant diversification rate before Pleistocene (Figs. 4 and 5; Table S3). Generally, “long” branches were not expected in the null model assuming a constant diversification rate in an evolutionary phase (Magallón & Sanderson, 2001). But the phylogenetic tree of Tylototriton s.l. had several quite long branches (e.g., T. vietnamensis and T. panhai), possessing basal evolutionary positions and inhabiting the ancestral region, i.e., the northern Indochina (Fig. 3). In the long branches, the pattern of high extinction rate, lack of speciation events and replacements as a result of species turn-over was difficult to be differentiated by only incorporating contemporary taxa (Magallón & Sanderson, 2001). Unfortunately, as there are no ingroup fossil known to now, even the parameter-rich methods might be less powerful for exploring influences from changes of extinction rates and random events (Rabosky, 2010; Quental & Marshall, 2010).
Radiation of Tylototriton s.l. and climatic shifts
Our findings suggested that the genus had originated from the tropical area in NIP-SY, and divergence times between major clades had all fall in the middle-to-late Miocene epochs (Fig. 3). The extensive dispersals from NIP-SY to north especially the mainland of China during the middle-to-late Miocene (Fig. 3) indicate that Tylototriton s.l. newts had adaptive radiations in southeastern Asia. It is presumed that diversification of Tylototriton s.l. should be closely related with significant climatic and tectonic transformations in the area, just like the diversification history of amount of numerous organisms which was considered to be modulated by the Tibetan Plateau uplifts and the intensifications of EAMs in the late Tertiary (Favre et al., 2014).
Origin of Tylototriton s.l. was dated back to ∼15 Mya in the middle Miocene (Fig. 3), when several dramatic climatic transitions had taken place in Asia. Around 16 ∼15 Mya, global climate had been marked by the Miocene Climate Optimum (MCO; Böhme, 2003), but from 15 Mya, climate begun deteriorating, especially in Eurasia; this climate transition led to an increase of seasonality and aridity (Eronen et al., 2009; Bruch et al., 2011). At the same time, the central Tibetan Plateau potentially had a main-body uplift (Spicer et al., 2003; Harris, 2006). These incidents enhanced the EAMs that directly led to the warm and humid climate in the South China (Wan et al., 2007). The climatic shifts further led to turnovers of fauna and flora in Asia (e.g., groups in Klaus et al., 2016). Correspondingly, the northward extensive dispersals from NIP-SY to CC in common ancestors of major clades of Tylototriton s.l. (Fig. 3) were much likely caused by the prosperity of the suitable subtropical monsoon climates in the mainland.
Evidence from the thermochronology, sedimentology, oceanography, and palaeoclimatology supported a rapid uplift of the Tibetan Plateau during the late Miocene about 10∼7 Mya that further enhanced the EAMs (An et al., 2001). Diversification of biodiversity in southeastern Asia was reported to be promoted by these remarkable events (Mao et al., 2010; Sun et al., 2012; Wang et al., 2012a; Wang et al., 2012b; Yu et al., 2014). Likewise, origin of most clades in Tylototriton s.l. were also estimated from the late Miocene (Fig. 2), indicating the impact of these historical climatic shifts on diversification of the newt genus.
Radiation of Tylototriton s.l. in southeastern Asia could also be shown by the fact that the Miocene-derived clades occupy natural environments distinct from those of the common ancestor of the genus. The common ancestor of Tylototriton s.l. and most clades likely shared habitats in the tropical area or near-tropical area in the northern Indochina Peninsula with the putative exceptions of clades I and V (Fig. 3). Clade I, containing two species (i.e., T. taliangensis and T. pseudoverrucosus; Figs. 1 and 3), is of particular endemism to several sky-island mountains in eastern Hengduan Mountains, where most populations inhabit high plateau (about 2,200–3,300 m; Li, Zou & Luo, 2011; Fei, Ye & Jiang, 2012). Their habitats are generally located in the northern subtropical or even temperate zones (Fei, Ye & Jiang, 2012). Furthermore, most species in clade V occupy the northernmost distribution range (Fig. 1A). In particular, newts in these clades undergo a dormant phase in the longer-term “cold” seasons by hiding themselves in soil caves or underground tree holes. Our results suggest that the evolution of these “specialized” lineages have been triggered the dispersal events from tropical area to the subtropics and/or the temperate zone during the late Miocene. Moreover, these species show very distinct morphological characters (Fei, Ye & Jiang, 2012) and reproductive behavior (such as T. taliangensis). All these points support their origin through Miocene radiations to the subtropics and temperate regions. Of course, reinforcement of the EAMs following the rapid uplifts of the Tibetan Plateau during the late Miocene (An et al., 2001) already changing biodiversity profiles in the southeastern Asia had certainly contributed to the radiations of these lineages to subtropics and plateau zones.
Anyhow, the empirical data generally support that diversification rates of the genus have decreased at beginning of Pleistocene which seems to reject the Pleistocene speciation hypothesis in this group. The rate-slowdown model was also previously observed in other animal groups, such as reptiles (Rawlings et al., 2008), birds and mammals (Weir & Schluter, 2007) and beetles (McKenna & Farrell, 2006). Numerous studies on amphibians occurring in the southeastern Asia also revealed that interspecific and even intraspecific divergences observed predate the Pleistocene (Wang et al., 2012a; Wang et al., 2012b; Wang et al., 2013). They supported the hypothesis that speciation of many animal lineages need a relatively long-lasting time at least before the glaciations (Zink & Slowinski, 1995). On the other hand, it is speculated that the glaciations might promote extinction rates that would slow down the diversification rates (Zink, Klicka & Barber, 2004). Based on our empirical findings, it is expected to have caused a drastic increase in extinction rates at the beginning, followed by depressed diversification rates due to the smaller areas that remained or became available as the habitat for the newts. As mentioned above, any attempt to estimate or simulate diversification rates from data from extant organisms alone is bound to underestimate both extinction and diversification rates, because whole radiations have gone extinct.
Conclusions
Our findings provided evidence for the profound influences of historical climate shifts especially associated with the uplift of the Tibetan Plateau on the diversification of animals occurring in the southeastern Asia. The results basically confirmed the reports on plant lineages, such as Rheum (Sun et al., 2012), Lepisorus (Wang et al., 2012a; Wang et al., 2012b), Juniperus (Mao et al., 2010) and Isodon (Yu et al., 2014), and on animal groups, such as Chiastocheta Pokorny, 1889 (Anthomyiidae, Diptera, Insecta; Espíndola, Buerki & Alvarez, 2012) and spiny frogs (Dicroglossidae, Anura, Amphibia; Che et al., 2010). Yet the present study further supplied some other indications. Primarily, although diversification of the newt group was promoted by the climatic shifts, several factors, for example, niche limits and their intrinsic low dispersal capacity, might have contributed to their Pleistocene rate-slowdown diversification. In addition, this study provided a putative “complete” species tree for Tylototriton s.l. to date. The inclusion of many putative cryptic species in the diversification analyses seems to be effective in finding diversification models of the group. But at present, wild populations of Tylototriton s.l. show a significant decline due to many reasons, such as climate changes, human capture and habitat deterioration. Further deep investigations of undetected cryptic lineages might supply the basic requirement for disclosing “true” diversification history of the taxa.
Supplemental Information
“*” sequences were downloaded from GenBank. “#” sequences were retrieved form Zhao et al. (2012). CIB, Chengdu Institute of Biology, Chinese Academy Science; HNNU, Henan Normal University; IEBR, Institute of Ecology and Biological Resources; VNUH, Vietnam National UniversityHanoi; VNMN, Vietnam National Museumof Nature; KUHE, Graduate School of Human and Environmental Studies of Kyoto University; CSUFT, Central South University of Forestry and Technology.
Different lineage delimitating strategies: GMYC, 39 statistically inferred coalescent lineages using the Generalized Mixed Yule Coalescent method; NS, 23 currently nominal species; NSP, 23 NS plus four independent lineages representing putative cryptic species. DDL and DDX, logistic and exponential density-dependent speciation models, respectively; yule2rate, multi-rate variant of the Yule model; SPVAR,time-varying speciation only, with a constant extinction rate; EXVAR, time-varying extinction only; BOTHVAR, both speciation and extinction vary over time; r1–r2, net diversification rate (speciation event per million years); a, extinction fraction; st, time of rate shift (Myr); κ, parameter in the logistic density dependent model; x, parameter in the density-dependent exponential model; κ, initial speciation rate for SPVAR model; κ, parameter of the exponential change in speciation rate for BOTHVAR model; z, parameter of exponential change in extinction rate; l, final extinction rate; Ln(L), loglikelihood; AIC, Akaike information criterion; deltaAIC, difference in AIC scores between Pure Birth model and the best-fit model.
Red shading branches indicate GMYC clusters. “*” individuals were used in *BEAST analyses.
{kind=link}
The tree was resulted from the topology-constrained Beast analysis. Fossil calibrations C1–C4 were given in materials for details. Grey bars at nodes show 95% highest posterior density intervals of divergence times.
{kind=link}
Different lineage delimitating strategies: GMYC, 39 statistically inferred coalescent lineages using the Generalized Mixed Yule Coalescent method; NS, 23 currently nominal species; and NSP, 23 NS plus four independent lineages representing putative cryptic species. Trees were simulated under the observed fixed time since origin (stem age = 22.6 Ma) using the constant rate birth–death process, with speciation and extinction rate values estimated from the empirical data (see Table S3 for different delimitating strategies). Simulations run under full sampling scenario, 10% missing extant lineage, 20% missing extant lineages, 30% missing extant lineages and 30% missing extant lineages. Thick bars correspond to the observed age of the common ancestor of extant Tylototriton s.l., with grey rectangles representing the 95% highest posterior density (HPD) interval.
{kind=link}
Acknowledgments
We thank JR Teng, J Wang, TN Vu, TT Nguyen, QK Le, DQ Rao and HJ Gu on helping collection of samples. We are much indebted to the editor and referees for their valuable comments.
Funding Statement
This study was supported by the following foundations: the Strategic Priority Research Program of the Chinese Academy of Sciences, Grant No. XDPB0202; the National Natural Sciences Foundation of China (NSFC-31201702, 31471964); Biodiversity Conservation Key Laboratory of Guizhou province Education Department, Guiyang College; the Alexander von Humboldt Foundation (VIE 114344); the project from the Ministry of Education of Japan (No. 20770066); the Kyoto University Foundation in 2008; JSPS AA Science Platform program in 2011; Science and technology Innovation Team Project of iupanshui Normal University LPSSYKJTD201602); Joint Science Foundation of Guizhou Province (Qiankehe-J-LKLS[2013]06). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Additional Information and Declarations
Competing Interests
The authors declare there are no competing interests.
Author Contributions
Bin Wang conceived and designed the experiments, performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, prepared figures and/or tables.
Kanto Nishikawa conceived and designed the experiments, performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, authored or reviewed drafts of the paper.
Masafumi Matsui conceived and designed the experiments, analyzed the data, contributed reagents/materials/analysis tools, authored or reviewed drafts of the paper.
Truong Quang Nguyen performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, authored or reviewed drafts of the paper.
Feng Xie analyzed the data, contributed reagents/materials/analysis tools, prepared figures and/or tables, authored or reviewed drafts of the paper.
Cheng Li and Janak Raj Khatiwada analyzed the data, contributed reagents/materials/analysis tools, authored or reviewed drafts of the paper.
Baowei Zhang, Dajie Gong, Yunming Mo, Gang Wei, Xiaohong Chen, Youhui Shen and Daode Yang contributed reagents/materials/analysis tools, authored or reviewed drafts of the paper.
Rongchuan Xiong analyzed the data, contributed reagents/materials/analysis tools.
Jianping Jiang conceived and designed the experiments, analyzed the data, prepared figures and/or tables, authored or reviewed drafts of the paper.
Animal Ethics
The following information was supplied relating to ethical approvals (i.e., approving body and any reference numbers):
The Animal Care and Use Committee of Chengdu Institute of Biology, CAS provided full approval for this purely observational research (CIB2010031015).
Field Study Permissions
The following information was supplied relating to field study approvals (i.e., approving body and any reference numbers):
Field experiments were approved by the Management Office of the Kuankuoshui Nature Reserve (number: KKSNR201204002) and the Management Office of the Dabashan Nature Reserve (number: DBSNR201204002).
DNA Deposition
Data Availability
The following information was supplied regarding data availability:
The raw data has been provided as a Supplemental File.
References
- An et al. (2001).An ZS, Kutzbach JE, Prell WL, Porter SC. Evolution of Asian monsoons and phased uplift of the Himalaya-Tibetan plateau since Late Miocene times. Nature. 2001;411:62–66. doi: 10.1038/35075035. [DOI] [PubMed] [Google Scholar]
- Arbogast et al. (2002).Arbogast BS, Edwards SV, Wakeley J, Beerli P, Slowinski JB. Estimating divergence times from molecular data on phylogenetic and population genetic timescales. Annual Review of Ecology and Systematics. 2002;33:707–740. doi: 10.1146/annurev.ecolsys.33.010802.150500. [DOI] [Google Scholar]
- Bidegaray-Batista, Ferrández & Arnedo (2014).Bidegaray-Batista L, Ferrández MÁ, Arnedo MA. Winter is coming: miocene and Quaternary climatic shifts shaped the diversification of Western-Mediterranean Harpactocrates (Araneae, Dysderidae) spiders. Cladistics. 2014;30:428–446. doi: 10.1111/cla.12054. [DOI] [PubMed] [Google Scholar]
- Blount, Borland & Lenski (2008).Blount ZD, Borland .CZ, Lenski RE. Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America. 2008;105:7899–7906. doi: 10.1073/pnas.0803151105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Böhme (2003).Böhme M. The miocene climatic optimum: evidence from ectothermic vertebrates of Central Europe. Palaeogeography Palaeoclimatology Palaeoecology. 2003;195:389–401. doi: 10.1016/S0031-0182(03)00367-5. [DOI] [Google Scholar]
- Bouckaert et al. (2014).Bouckaert R, Heled J, Kühnert D, Vaughn T, Wu CH, Xie D, Suchard MA, Rambaut A, Drummond AJ. BEAST 2: a software platform for Bayesian evolutionary analysis. PLOS Computational Biology. 2014;10(4):e1003537. doi: 10.1371/journal.pcbi.1003537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruch et al. (2011).Bruch AA, Utescher T, Mosbrugger V, NECLIME members Precipitation patterns in the Miocene of Central Europe and the development of continentality. Palaeogeography Palaeoclimatology Palaeoecology. 2011;304:202–211. doi: 10.1016/j.palaeo.2010.10.002. [DOI] [Google Scholar]
- Castresana (2000).Castresana J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Molecular Biology and Evolution. 2000;17:540–552. doi: 10.1093/oxfordjournals.molbev.a026334. [DOI] [PubMed] [Google Scholar]
- Che et al. (2010).Che J, Zhou WW, Hu JS, Yan F, Papenfuss TJ, Wake DB, Zhang YP. Spiny frogs (Paini) illuminate the history of the Himalayan region and Southeast Asia. Proceedings of the National Academy of Sciences of the United States of America. 2010;107:13765–13770. doi: 10.1073/pnas.1008415107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, Wang & Tao (2010).Chen XH, Wang XW, Tao J. A new subspecies of genus Tylototriton from China (Caudata: Salamandridae) Acta Zootaxonomica Sinica. 2010;35:666–670. [Google Scholar]
- Coope (2004).Coope GR. Several million years of stability among insect species because of, or in spite of, Ice Age climatic instability? Philosophical Transactions of the Royal Society B: Biological Sciences. 2004;359:209–214. doi: 10.1098/rstb.2003.1393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cusimano, Stadler & Renner (2012).Cusimano N, Stadler T, Renner SS. A new method for handling missing species in diversification analysis applicable to randomly or nonrandomly sampled phylogenies. Systematic Biology. 2012;61:785–792. doi: 10.1093/sysbio/sys031. [DOI] [PubMed] [Google Scholar]
- Darriba et al. (2012).Darriba D, Taboada GL, Doallo R, Posada D. jModelTest 2: more models, new heuristics and parallel computing. Nature Methods. 2012;9(8):772. doi: 10.1038/nmeth.2109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Degnan & Rosenberg (2009).Degnan JH, Rosenberg NA. Gene tree discordance, phylogenetic inference and the multispecies coalescent. Trends in Ecology & Evolution. 2009;24:332–340. doi: 10.1016/j.tree.2009.01.009. [DOI] [PubMed] [Google Scholar]
- Ding et al. (2011).Ding L, Gan XN, He SP, Zhao EM. A phylogeographic, demographic and historical analysis of the short-tailed pit viper (Gloydius brevicaudus): evidence for early divergence and late expansion during the Pleistocene. Molecular Ecology. 2011;20:1905–1922. doi: 10.1111/j.1365-294X.2011.05060.x. [DOI] [PubMed] [Google Scholar]
- Dubey & Shine (2012).Dubey S, Shine R. Are reptile and amphibian species younger in the Northern Hemisphere than in the Southern Hemisphere? Journal of Evolutionary Biology. 2012;25:220–226. doi: 10.1111/j.1420-9101.2011.02417.x. [DOI] [PubMed] [Google Scholar]
- Edwards & Beerli (2000).Edwards SV, Beerli P. Perspective: gene divergence, population divergence, and the variance in coalescence time in phylogeographic studies. Evolution. 2000;54:1839–1854. doi: 10.1111/j.0014-3820.2000.tb01231.x. [DOI] [PubMed] [Google Scholar]
- Eronen et al. (2009).Eronen JT, Ataabadi MM, Micheels A, Karme A, Bernor RL, Fortelius M. Distribution history and climatic controls of the Late Miocene Pikermian chronofauna. Proceedings of the National Academy of Sciences of the United States of America. 2009;106:11867–11871. doi: 10.1073/pnas.0902598106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Espíndola, Buerki & Alvarez (2012).Espíndola A, Buerki S, Alvarez N. Ecological and historical drivers of diversification in the fly genus Chiastocheta Pokorny. Molecular Phylogenetics and Evolution. 2012;63:466–474. doi: 10.1016/j.ympev.2012.01.018. [DOI] [PubMed] [Google Scholar]
- Estes (1981).Estes R. Gymnophiona, Caudata. HandBuch Der Paloherpetologie. 1981;2:1–115. [Google Scholar]
- Favre et al. (2014).Favre A, Packert M, Pauls SU, Jahnig SC, Uhl D, Michalak I, Muellner-Riehl AN. The role of the uplift of the Qinghai-Tibetan Plateau for the evolution of Tibetan biotas. Biological Reviews of the Cambridge Philosophical Society. 2014;90(1):236–253. doi: 10.1111/brv.12107. [DOI] [PubMed] [Google Scholar]
- Fei, Ye & Jiang (2010).Fei L, Ye CY, Jiang JP. Progress and prospects for studies on Chinese amphibians. Asian Herpetological Research. 2010;2:1–22. [Google Scholar]
- Fei, Ye & Jiang (2012).Fei L, Ye CY, Jiang JP. Colored atlas of Chinese amphibians and their distributions. Sichuan Publishing House of Science and Technology; Chengdu: 2012. pp. 1–620. [Google Scholar]
- Fontaneto et al. (2007).Fontaneto D, Herniou EA, Boschetti C, Caprioli M, Melone G, Ricci C, Barraclough TG. Independently evolving species in asexual bdelloid rotifers. PLOS Biology. 2007;5:e87. doi: 10.1371/journal.pbio.0050087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forest (2009).Forest F. Calibrating the tree of life: fossils, molecules and evolutionary timescales. Annals of Botany. 2009;104:789–794. doi: 10.1093/aob/mcp192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frost (2016).Frost DR. American museum of natural history. New York, USA: 2016. [Google Scholar]
- Gu et al. (2013).Gu L, Liu Y, Que P, Zhang Z. Quaternary climate and environmental changes have shaped genetic differentiation in a Chinese pheasant endemic to the eastern margin of the Qinghai-Tibetan Plateau. Molecular Phylogenetics and Evolution. 2013;67:129–139. doi: 10.1016/j.ympev.2012.12.013. [DOI] [PubMed] [Google Scholar]
- Guindon & Gascuel (2003).Guindon S, Gascuel O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Systematic Biology. 2003;52:696–704. doi: 10.1080/10635150390235520. [DOI] [PubMed] [Google Scholar]
- Haffer (1969).Haffer J. Speciation in Amazonian forest birds. Science. 1969;165:131–137. doi: 10.1126/science.165.3889.131. [DOI] [PubMed] [Google Scholar]
- Harmon et al. (2008).Harmon LJ, Weir JT, Brock CD, Glor RE, Challenger W. GEIGER: investigating evolutionary radiations. Bioinformatics. 2008;24:129–131. doi: 10.1093/bioinformatics/btm538. [DOI] [PubMed] [Google Scholar]
- Harris (2006).Harris N. The elevation history of the Tibetan Plateau and its implications for the Asian monsoon. Palaeogeography Palaeoclimatology Palaeoecology. 2006;241:4–15. doi: 10.1016/j.palaeo.2006.07.009. [DOI] [Google Scholar]
- Harrison et al. (1992).Harrison TM, Copeland P, Kidd WS, Yin A. Raising tibet. Science. 1992;255:1663–1670. doi: 10.1126/science.255.5052.1663. [DOI] [PubMed] [Google Scholar]
- Heled & Drummond (2010).Heled J, Drummond AJ. Bayesian inference of species trees from multilocus data. Molecular Biology and Evolution. 2010;27:570–580. doi: 10.1093/molbev/msp274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herre (1935).Herre W. Die Schwanzlurche der mitteleocänen (oberlutetischen) Braunkohle des Geiseltales und die Phylogenie der Urodelen unter Einschluss der fossilen Formen. Zoologica. 1935;33:1–85. [Google Scholar]
- Hewitt (2000).Hewitt G. The genetic legacy of the Quaternary ice ages. Nature. 2000;405:907–913. doi: 10.1038/35016000. [DOI] [PubMed] [Google Scholar]
- Hou, Li & Lu (2012).Hou M, Li PP, Lu SQ. Morphological research development of genus Tylototriton and primary confirmation of the status of four cryptic populations. Journal of Huangshan University. 2012;14:61–65. [Google Scholar]
- Katoh et al. (2002).Katoh K, Misawa K, Kuma K, Miyata T. MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Research. 2002;30:3059–3066. doi: 10.1093/nar/gkf436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kennedy et al. (2012).Kennedy JD, Weir JT, Hooper DM, Tietze DT, Martens J, Price TD. Ecological limits on diversification of the Himalayan core Corvoidea. Evolution. 2012;66:2599–2613. doi: 10.1111/j.1558-5646.2012.01618.x. [DOI] [PubMed] [Google Scholar]
- Khatiwada et al. (2015).Khatiwada JR, Wang B, Ghimire S, Vasudevan K, Paudel S, Jiang JP. A new species of the genus Tylototriton (Amphibia: Urodela: Salamandridae) from eastern Himalaya. Asian Herpetological Research. 2015;6:245–256. [Google Scholar]
- Klaus et al. (2016).Klaus S, Morley RJ, Plath M, Zhang YP, Li JT. Biotic interchange between the Indian subcontinent and mainland Asia through time. Nature Communications. 2016;7:12132. doi: 10.1038/ncomms12132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knowles (2001).Knowles LL. Did the pleistocene glaciations promote divergence? Tests of explicit refugial models in montane grasshopprers. Molecular Ecology. 2001;10:691–701. doi: 10.1046/j.1365-294x.2001.01206.x. [DOI] [PubMed] [Google Scholar]
- Knowles & Richards (2005).Knowles LL, Richards CL. Importance of genetic drift during Pleistocene divergence as revealed by analyses of genomic variation. Molecular Ecology. 2005;14:4023–4032. doi: 10.1111/j.1365-294X.2005.02711.x. [DOI] [PubMed] [Google Scholar]
- Kubatko & Degnan (2007).Kubatko LS, Degnan JH. Inconsistency of phylogenetic estimates from concatenated data under coalescence. Systematic Biology. 2007;56:17–24. doi: 10.1080/10635150601146041. [DOI] [PubMed] [Google Scholar]
- Kurabayashi et al. (2012).Kurabayashi A, Nishitani T, Katsuren S, Oumi S, Sumida M. Mitochondrial genomes and divergence times of crocodile newts: inter-islands distribution of Echinotriton andersoni and the origin of a unique repetitive sequence found in mt genomes Tylototriton. Genes & Genetic Systems. 2012;87:39–51. doi: 10.1266/ggs.87.39. [DOI] [PubMed] [Google Scholar]
- Lanfear et al. (2012).Lanfear R, Calcott B, Ho SY, Guindon S. Partitionfinder: combined selection of partitioning schemes and substitution models for phylogenetic analyses. Molecular Biology and Evolution. 2012;29:1695–1701. doi: 10.1093/molbev/mss020. [DOI] [PubMed] [Google Scholar]
- Lemmon & Moriarty (2004).Lemmon AR, Moriarty EC. The importance of proper model assumption in Bayesian phylogenetics. Systematic Biology. 2004;53:265–277. doi: 10.1080/10635150490423520. [DOI] [PubMed] [Google Scholar]
- Leneveu, Chichvarkhin & Wahlberg (2009).Leneveu J, Chichvarkhin A, Wahlberg N. Varying rates of diversification in the genus Melitaea (Lepidoptera: Nymphalidae) during the past 20 million years. Biological Journal of the Linnean Society. 2009;97:346–361. doi: 10.1111/j.1095-8312.2009.01208.x. [DOI] [Google Scholar]
- Li & Fang (1999).Li J, Fang X. Uplift of the Tibetan Plateau and environmental changes. Chinese Science Bulletin. 1999;44:2117–2124. doi: 10.1007/BF03182692. [DOI] [Google Scholar]
- Li, Zou & Luo (2011).Li B, Zou LK, Luo Y. The survey of Tylototriton taliangensis habitat and its conservation. Sichuan Journal of Zoology. 2011;30:964–966. [Google Scholar]
- Lin et al. (2014).Lin AQ, Csorba G, Li LF, Jiang TL, Lu GJ, Thong VD, Soisook P, Sun KP, Feng J. Phylogeography of Hipposideros armiger (Chiroptera: Hipposideridae) in the oriental region: the contribution of multiple pleistocene glacial refugia and intrinsic factors to contemporary population genetic structure. Journal of Biogeography. 2014;41:317–327. doi: 10.1111/jbi.12163. [DOI] [Google Scholar]
- Liu et al. (2006).Liu JQ, Wang YJ, Wang AL, Hideaki O, Abbott RJ. Radiation and diversification within the Ligularia-Cremanthodium-Parasenecio complex (Asteraceae) triggered by uplift of the Qinghai-Tibetan Plateau. Molecular Phylogenetics and Evolution. 2006;38:31–49. doi: 10.1016/j.ympev.2005.09.010. [DOI] [PubMed] [Google Scholar]
- Maddison (1997).Maddison WP. Gene trees in species trees. Systematic Biology. 1997;46:523–536. doi: 10.1093/sysbio/46.3.523. [DOI] [Google Scholar]
- Magallón & Sanderson (2001).Magallón S, Sanderson MJ. Absolute diversification rates in angiosperm clades. Evolution. 2001;55:1762–1780. doi: 10.1111/j.0014-3820.2001.tb00826.x. [DOI] [PubMed] [Google Scholar]
- Mao et al. (2010).Mao K, Hao G, Liu J, Adams RP, Milne RI. Diversification and biogeography of Juniperus (Cupressaceae): variable diversification rates and multiple intercontinental dispersals. New Phytologist. 2010;188:254–272. doi: 10.1111/j.1469-8137.2010.03351.x. [DOI] [PubMed] [Google Scholar]
- Martin et al. (2010).Martin DP, Lemey P, Lott M, Moulton V, Posada D, Lefeuvre P. RDP3: a flexible and fast computer program for analyzing recombination. Bioinformatics. 2010;26:2462–2463. doi: 10.1093/bioinformatics/btq467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCormack et al. (2010).McCormack JE, Heled J, Delaney KS, Peterson AT, Knowles LL. Calibrating divergence times on species trees versus gene trees: implications for speciation history of Aphelocoma jays. Evolution. 2010;65:184–202. doi: 10.1111/j.1558-5646.2010.01097.x. [DOI] [PubMed] [Google Scholar]
- McGuire et al. (2007).McGuire JA, Witt CC, Altshule DL, Remsen JV. Phylogenetic systematics and biogeography of hummingbirds: bayesian and maximum likelihood analyses of partitioned data and selection of an appropriate partitioning strategy. Systematic Biology. 2007;56:837–856. doi: 10.1080/10635150701656360. [DOI] [PubMed] [Google Scholar]
- McKenna & Farrell (2006).McKenna DD, Farrell BD. Tropical forests are both evolutionary cradles and museums of leaf beetle diversity. Proceedings of the National Academy of Sciences of the United States of America. 2006;103:10947–10951. doi: 10.1073/pnas.0602712103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milner (2000).Milner A. Mesozoic and tertiary Caudata and Albanerpetontidae. Amphibian Biology. 2000;4:1412–1444. [Google Scholar]
- Molnar, Boos & Battisti (2010).Molnar P, Boos WR, Battisti DS. Orographic controls on climate and paleoclimate of Asia: thermal and mechanical roles for the Tibetan Plateau. Annual Review of Earth and Planetary Sciences. 2010;38:77–102. doi: 10.1146/annurev-earth-040809-152456. [DOI] [Google Scholar]
- Monaghan et al. (2009).Monaghan MT, Wild R, Elliot M, Fujisawa T, Balke M, Inward DJG, Lees DC, Ranaivosolo R, Eggleton P, Barraclough TG. Accelerated species inventory on Madagascar using coalescent-based models of species delineation. Systematic Biology. 2009;58:298–311. doi: 10.1093/sysbio/syp027. [DOI] [PubMed] [Google Scholar]
- Myers et al. (2000).Myers N, Mittermeier RA, Mittermeier CG, Da Fonseca GA, Kent J. Biodiversity hotspots for conservation priorities. Nature. 2000;403:853–858. doi: 10.1038/35002501. [DOI] [PubMed] [Google Scholar]
- Nishikawa et al. (2013).Nishikawa K, Khonsue W, Pomchote P, Matsui M. Two new species of Tylototriton from Thailand (Amphibia: Urodela: Salamandridae) Zootaxa. 2013;3737:261–279. doi: 10.11646/zootaxa.3737.3.5. [DOI] [PubMed] [Google Scholar]
- Nishikawa, Matsui & Nguyen (2013).Nishikawa K, Matsui M, Nguyen TT. A new species of Tylototriton from northern Vietnam (Amphibia: Urodela: Salamandridae) Current Herpetology. 2013;32:34–49. doi: 10.5358/hsj.32.34. [DOI] [Google Scholar]
- Nishikawa, Matsui & Rao (2014).Nishikawa K, Matsui M, Rao DQ. A new species of Tylototriton (Amphibia: Urodela: Salamandridae) from central Myanmar. Natural History Bulletin of the Siam Society. 2014;60:9–22. [Google Scholar]
- Nishikawa et al. (2015).Nishikawa K, Rao DQ, Matsui M, Eto K. Taxonomic relationship between Tylototriton daweishanensis Zhao, Rao, Liu, Li and Yuan, 2012 and T. yangi (Hou, Li & Lu, 2012) (Amphibia: Urodela: Salamandridae) Current Herpetology. 2015;34:67–74. doi: 10.5358/hsj.34.67. [DOI] [Google Scholar]
- Oaks (2011).Oaks JR. A time-calibrated species tree of Crocodylia reveals a recent radiation of the true crocodiles. Evolution. 2011;65:3285–3297. doi: 10.1111/j.1558-5646.2011.01373.x. [DOI] [PubMed] [Google Scholar]
- Peng et al. (2006).Peng Z, Ho SY, Zhang Y, He S. Uplift of the Tibetan plateau: evidence from divergence times of glyptosternoid catfishes. Molecular Phylogenetics and Evolution. 2006;39:568–572. doi: 10.1016/j.ympev.2005.10.016. [DOI] [PubMed] [Google Scholar]
- Phillimore & Price (2008).Phillimore AB, Price TD. Density-dependent cladogenesis in birds. PLOS Biology. 2008;6:e71. doi: 10.1371/journal.pbio.0060071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phimmachak, Aowphol & Stuart (2015).Phimmachak S, Aowphol A, Stuart BL. Morphological and molecular variation in Tylototriton (Caudata: Salamandridae) in Laos, with description of a new species. Zootaxa. 2015;4006:285–310. doi: 10.11646/zootaxa.4006.2.3. [DOI] [PubMed] [Google Scholar]
- Pons et al. (2006).Pons J, Barraclough T, Gomez-Zurita J, Cardoso A, Duran D, Hazell S, Kamoun S, Sumlin W, Vogler A. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Systematic Biology. 2006;55:595–609. doi: 10.1080/10635150600852011. [DOI] [PubMed] [Google Scholar]
- Powell (2012).Powell JR. Accounting for uncertainty in species delineation during the analysis of environmental DNA sequence data. Methods in Ecology and Evolution. 2012;3:1–11. doi: 10.1111/j.2041-210X.2011.00122.x. [DOI] [Google Scholar]
- Quental & Marshall (2010).Quental TB, Marshall CR. Diversity dynamics: molecular phylogenies need the fossil record. Trends in Ecology & Evolution. 2010;25:434–441. doi: 10.1016/j.tree.2010.05.002. [DOI] [PubMed] [Google Scholar]
- R Development Core Team (2008).R Development Core Team . R Foundation for Statistical Computing. Vienna: 2008. R: a language and environment for statistical computing. [Google Scholar]
- Rabosky (2006).Rabosky DL. LASER: a maximum likelihood toolkit for detecting temporal shifts in diversification rates from molecular phylogenies. Evolutionary Bioinformatics Online. 2006;2:273–276. [PMC free article] [PubMed] [Google Scholar]
- Rabosky (2010).Rabosky DL. Extinction rates should not be estimated from molecular phylogenies. Evolution. 2010;64:1816–1824. doi: 10.1111/j.1558-5646.2009.00926.x. [DOI] [PubMed] [Google Scholar]
- Rabosky & Lovette (2008a).Rabosky DL, Lovette IJ. Density-dependent diversification in North American wood warblers. Proceedings of the Royal Society B: Biological Sciences. 2008a;275:2363–2371. doi: 10.1098/rspb.2008.0630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rabosky & Lovette (2008b).Rabosky DL, Lovette IJ. Explosive evolutionary radiations: decreasing speciation or increasing extinction through time? Evolution. 2008b;62:1866–1875. doi: 10.1111/j.1558-5646.2008.00409.x. [DOI] [PubMed] [Google Scholar]
- Rambaut et al. (2014).Rambaut A, Suchard MA, Xie D, Drummond AJ. Tracer v1.6. 2014. http://beast.community/tracer http://beast.community/tracer
- Rawlings et al. (2008).Rawlings LH, Rabosky DL, Donnellan SC, Hutchinson MN. Python phylogenetics: inference from morphology and mitochondrial DNA. Biological Journal of the Linnean Society. 2008;93:603–619. doi: 10.1111/j.1095-8312.2007.00904.x. [DOI] [Google Scholar]
- Renner & Cusimano (2010).Renner SS, Cusimano N. Slowdowns in diversification rates from real phylogenies may not be real. Systematic Biology. 2010;59:458–464. doi: 10.1093/sysbio/syq032. [DOI] [PubMed] [Google Scholar]
- Roček (1996a).Roček Z. Skull of the neotenic salamandrid amphibian Triturus alpestris and abbreviated development in the Tertiary Salamandridae. Journal of Morphology. 1996a;230:187–197. doi: 10.1002/(SICI)1097-4687(199611)230:2<187::AID-JMOR6>3.0.CO;2-E. [DOI] [PubMed] [Google Scholar]
- Roček (1996b).Roček Z. The salamander Brachycormus noachicus from the Oligocene of Europe, and the role of neoteny in the evolution of salamanders. Palaeontology. 1996b;39:477–495. [Google Scholar]
- Ronquist et al. (2012).Ronquist F, Teslenko M, Van der Mark P, Ayres DL, Darling A, Hohna S, Larget B, Liu L, Suchard MA, Huelsenbeck JP. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology. 2012;61:539–542. doi: 10.1093/sysbio/sys029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sánchez-Gracia & Castresana (2012).Sánchez-Gracia A, Castresana J. Impact of deep coalescence on the reliability of species tree inference from different types of DNA markers in mammals. PLOS ONE. 2012;7:e30239. doi: 10.1371/journal.pone.0030239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen, Jiang & Mo (2012).Shen YH, Jiang JP, Mo XY. A new species of the genus Tylototriton (Amphibia, Salamandridae) from Hunan, China. Asiatic Herpetological Research. 2012;3:21–30. doi: 10.3724/SP.J.1245.2012.00021. [DOI] [Google Scholar]
- Shen et al. (2016).Shen XX, Liang D, Chen MY, Mao RL, Wake DB, Zhang P. Enlarged multilocus data set provides surprisingly younger time of origin for the Plethodontidae, the largest family of salamanders. Systematic Biology. 2016;65:66–81. doi: 10.1093/sysbio/syv061. [DOI] [PubMed] [Google Scholar]
- Simmons, Ochoterena & Carr (2001).Simmons MP, Ochoterena H, Carr TG. Incorporation, relative homoplasy, and effect of gap characters in sequence-based phylogenetic analysis. Systematic Biology. 2001;50:454–462. doi: 10.1080/10635150120427. [DOI] [PubMed] [Google Scholar]
- Smith et al. (2013).Smith BT, Ribas CC, Whitney BW, Hernández-Baños BE, Klicka J. Identifying biases at different spatial and temporal scales of diversification: a case study using the Neotropical parrotlet genus Forpus. Molecular Ecology. 2013;22:483–494. doi: 10.1111/mec.12118. [DOI] [PubMed] [Google Scholar]
- Spicer et al. (2003).Spicer RA, Harris NB, Widdowson M, Herman AB, Guo S, Valdes PJ, Wolfe JA, Kelley SP. Constant elevation of southern Tibet over the past 15 million years. Nature. 2003;421:622–624. doi: 10.1038/nature01356. [DOI] [PubMed] [Google Scholar]
- Stadler (2011).Stadler T. Simulating trees with a fixed number of extant species. Systematic Biology. 2011;60:676–684. doi: 10.1093/sysbio/syr029. [DOI] [PubMed] [Google Scholar]
- Stamatakis (2014).Stamatakis A. RAxML Version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014;30:1312–1313. doi: 10.1093/bioinformatics/btu033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinfartz et al. (2007).Steinfartz S, Vicario S, Arntzen JW, Caccone A. A Bayesian approach on molecules and behavior: reconsidering phylogenetic and evolutionary patterns of the Salamandridae with emphasis on Triturus newts. Journal of Experimental Zoology. 2007;308B:139–162. doi: 10.1002/jez.b.21119. [DOI] [PubMed] [Google Scholar]
- Sun & Wang (2005).Sun X, Wang P. How old is the Asian monsoon system?—Palaeobotanical records from China. Palaeogeography Palaeoclimatology Palaeoecology. 2005;222:181–222. doi: 10.1016/j.palaeo.2005.03.005. [DOI] [Google Scholar]
- Sun et al. (2012).Sun Y, Wang A, Wan D, Wang Q, Liu J. Rapid radiation of Rheum (Polygonaceae) and parallel evolution of morphological traits. Molecular Phylogenetics and Evolution. 2012;63:150–158. doi: 10.1016/j.ympev.2012.01.002. [DOI] [PubMed] [Google Scholar]
- Tamura et al. (2013).Tamura K, Glen S, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Molecular Biology and Evolution. 2013;30:2725–2729. doi: 10.1093/molbev/mst197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas et al. (2010).Thomas JA, Welch JJ, Lanfear R, Bromham L. A generation time effect on the rate of molecular evolution in invertebrates. Molecular Biology and Evolution. 2010;27:1173–1180. doi: 10.1093/molbev/msq009. [DOI] [PubMed] [Google Scholar]
- Townsend et al. (2011).Townsend TM, Mulcahy DG, Noonan BP, Sites JW, Kuczynski CA, Wiens JJ, Reeder TW. Phylogeny of iguanian lizards inferred from 29 nuclear loci, and a comparison of concatenated and species-tree approaches for an ancient, rapid radiation. Molecular Phylogenetics and Evolution. 2011;61:263–380. doi: 10.1016/j.ympev.2011.07.008. [DOI] [PubMed] [Google Scholar]
- Turkheimer, Hinz & Cunningham (2003).Turkheimer FE, Hinz R, Cunningham VJ. On the undecidability among kinetic models: from model selection to model averaging. Journal of Cerebral Blood Flow & Metabolism. 2003;23:490–498. doi: 10.1097/01.WCB.0000050065.57184.BB. [DOI] [PubMed] [Google Scholar]
- Wan et al. (2007).Wan S, Li A, Clift PD, Stuut JBW. Development of the East Asian monsoon: mineralogical and sedimentologic records in the northern South China Sea since 20 Ma. Palaeogeography Palaeoclimatology Palaeoecology. 2007;254:561–582. doi: 10.1016/j.palaeo.2007.07.009. [DOI] [Google Scholar]
- Wang et al. (2012a).Wang B, Jiang J, Xie F, Li C. Postglacial colonization of the Qinling Mountains: phylogeography of the swelled vent frog (Feirana quadranus) PLOS ONE. 2012a;7:e41579. doi: 10.1371/journal.pone.0041579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang et al. (2013).Wang B, Jiang J, Xie F, Li C. Phylogeographic patterns of mtDNA variation revealed multiple glacial refugia for the frog species Feirana taihangnica endemic to the Qinling Mountains. Journal of Molecular Evolution. 2013;76:112–128. doi: 10.1007/s00239-013-9544-5. [DOI] [PubMed] [Google Scholar]
- Wang et al. (2012b).Wang L L, Schneider H, Zhang XC, Xiang QP. The rise of the Himalaya enforced the diversification of SE Asian ferns by altering the monsoon regimes. BMC Plant Biology. 2012b;12:210. doi: 10.1186/1471-2229-12-210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weir & Schluter (2007).Weir JT, Schluter D. The latitudinal gradient in recent speciation and extinction rates of birds and mammals. Science. 2007;315:1574–1576. doi: 10.1126/science.1135590. [DOI] [PubMed] [Google Scholar]
- Yang et al. (2014).Yang DD, Jiang JP, Shen YH, Fei DB. A new species of the genus Tylototriton (Urodela: Salamandridae) from northeastern Hunan Province, China. Asian Herpetological Research. 2014;5:1–11. doi: 10.3724/SP.J.1245.2014.00001. [DOI] [Google Scholar]
- Yao et al. (2011).Yao YF, Bruch AA, Mosbrugger V, Li CS. Quantitative reconstruction of Miocene climate patterns and evolution in Southern China based on plant fossils. Palaeogeography Palaeoclimatology Palaeoecology. 2011;304:291–307. doi: 10.1016/j.palaeo.2010.04.012. [DOI] [Google Scholar]
- Young & Healy (2003).Young ND, Healy J. GapCoder automates the use ofindel characters in phylogenetic analysis. BMC Bioinformatics. 2003;4:6. doi: 10.1186/1471-2105-4-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu, Harris & He (2010).Yu Y, Harris AJ, He X. S-DIVA (Statistical Dispersal-Vicariance Analysis): a tool for inferring biogeographic histories. Molecular Phylogenetics and Evolution. 2010;56:848–850. doi: 10.1016/j.ympev.2010.04.011. [DOI] [PubMed] [Google Scholar]
- Yu, Harris & He (2012).Yu Y, Harris AJ, He XJ. RASP (Reconstruct Ancestral State in Phylogenies) 2.1 beta. 2012. http://mnh.scu.edu.cn/soft/blog/RASP/ http://mnh.scu.edu.cn/soft/blog/RASP/ [DOI] [PubMed]
- Yu et al. (2014).Yu XQ, Maki M, Drew BT, Paton AJ, Li HW, Zhao JL, Conran JG, Li J. Phylogeny and historical biogeography of Isodon (Lamiaceae): rapid radiation in south-west China and Miocene overland dispersal into Africa. Molecular Phylogenetics and Evolution. 2014;77:183–194. doi: 10.1016/j.ympev.2014.04.017. [DOI] [PubMed] [Google Scholar]
- Yuan et al. (2011).Yuan ZY, Jiang K, Lu SQ, Yang JX, Nguyen QT, Nguyen TT, Jin JQ, Che J. A phylogeny of the Tylototriton asperrimus group (Caudata: Salamandridae) based on a mitochondrial study: suggestions for a taxonomic revision. Zoological Research. 2011;32:577–584. doi: 10.3724/SP.J.1141.2011.06577. [DOI] [PubMed] [Google Scholar]
- Zhang et al. (2010).Zhang DR, Chen MY, Murphy RW, Che J, Pang JF, Hu JS, Luo J, Wu SJ, Ye H, Zhang YP. Genealogy and palaeodrainage basins in Yunnan Province: phylogeography of the Yunnan spiny frog, Nanorana yunnanensis (Dicroglossidae) Molecular Ecology. 2010;19:3406–3420. doi: 10.1111/j.1365-294X.2010.04747.x. [DOI] [PubMed] [Google Scholar]
- Zhang et al. (2008).Zhang P, Papenfuss TJ, Wake MH, Qu L, Wake DB. Phylogeny and biogeography of the family Salamandridae (Amphibia: Caudata) inferred from complete mitochondrial genomes. Molecular Phylogenetics and Evolution. 2008;49:586–597. doi: 10.1016/j.ympev.2008.08.020. [DOI] [PubMed] [Google Scholar]
- Zhang et al. (2007).Zhang MW, Rao DQ, Yu GH, Yang JXT. The validity of red knobby newt (Tylototriton shanjing) species status based on mitochondrial cytb gene. Zoological Research. 2007;28:430–436. [Google Scholar]
- Zhao et al. (2012).Zhao YT, Rao DQ, Liu N, Li B, Yuan SQ. Molecular phylogeny analysis of Tylototriton verrucosus group and description of new species. Journal of West China Forestry Science. 2012;41:86–89. [Google Scholar]
- Zink & Klicka (2006).Zink RM, Klicka J. The tempo of avian diversification: a comment on Johnson and Cicero. Evolution. 2006;60:411–414. doi: 10.1111/j.0014-3820.2006.tb01118.x. [DOI] [PubMed] [Google Scholar]
- Zink, Klicka & Barber (2004).Zink RM, Klicka J, Barber BR. The tempo of avian diversification during the Quaternary. Philosophical Transactions of the Royal Society B: Biological Sciences. 2004;359:215–220. doi: 10.1098/rstb.2003.1392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zink & Slowinski (1995).Zink RM, Slowinski JB. Evidence from molecular systematics for decreased avian diversification in the pleistocene Epoch. Proceedings of the National Academy of Sciences of the United States of America. 1995;92:5832–5835. doi: 10.1073/pnas.92.13.5832. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
“*” sequences were downloaded from GenBank. “#” sequences were retrieved form Zhao et al. (2012). CIB, Chengdu Institute of Biology, Chinese Academy Science; HNNU, Henan Normal University; IEBR, Institute of Ecology and Biological Resources; VNUH, Vietnam National UniversityHanoi; VNMN, Vietnam National Museumof Nature; KUHE, Graduate School of Human and Environmental Studies of Kyoto University; CSUFT, Central South University of Forestry and Technology.
Different lineage delimitating strategies: GMYC, 39 statistically inferred coalescent lineages using the Generalized Mixed Yule Coalescent method; NS, 23 currently nominal species; NSP, 23 NS plus four independent lineages representing putative cryptic species. DDL and DDX, logistic and exponential density-dependent speciation models, respectively; yule2rate, multi-rate variant of the Yule model; SPVAR,time-varying speciation only, with a constant extinction rate; EXVAR, time-varying extinction only; BOTHVAR, both speciation and extinction vary over time; r1–r2, net diversification rate (speciation event per million years); a, extinction fraction; st, time of rate shift (Myr); κ, parameter in the logistic density dependent model; x, parameter in the density-dependent exponential model; κ, initial speciation rate for SPVAR model; κ, parameter of the exponential change in speciation rate for BOTHVAR model; z, parameter of exponential change in extinction rate; l, final extinction rate; Ln(L), loglikelihood; AIC, Akaike information criterion; deltaAIC, difference in AIC scores between Pure Birth model and the best-fit model.
Red shading branches indicate GMYC clusters. “*” individuals were used in *BEAST analyses.
The tree was resulted from the topology-constrained Beast analysis. Fossil calibrations C1–C4 were given in materials for details. Grey bars at nodes show 95% highest posterior density intervals of divergence times.
Different lineage delimitating strategies: GMYC, 39 statistically inferred coalescent lineages using the Generalized Mixed Yule Coalescent method; NS, 23 currently nominal species; and NSP, 23 NS plus four independent lineages representing putative cryptic species. Trees were simulated under the observed fixed time since origin (stem age = 22.6 Ma) using the constant rate birth–death process, with speciation and extinction rate values estimated from the empirical data (see Table S3 for different delimitating strategies). Simulations run under full sampling scenario, 10% missing extant lineage, 20% missing extant lineages, 30% missing extant lineages and 30% missing extant lineages. Thick bars correspond to the observed age of the common ancestor of extant Tylototriton s.l., with grey rectangles representing the 95% highest posterior density (HPD) interval.
Data Availability Statement
The following information was supplied regarding data availability:
The raw data has been provided as a Supplemental File.