

Abstract
The success of Staphylococcus aureus, as both a human and animal pathogen, stems from its ability to rapidly adapt to a wide spectrum of environmental conditions. Two-component systems (TCSs) play a crucial role in this process. Here, we describe a novel staphylococcal virulence factor, SpdC, an Abi-domain protein, involved in signal sensing and/or transduction. We have uncovered a functional link between the WalKR essential TCS and the SpdC Abi membrane protein. Expression of spdC is positively regulated by the WalKR system and, in turn, SpdC negatively controls WalKR regulon genes, effectively constituting a negative feedback loop. The WalKR system is mainly involved in controlling cell wall metabolism through regulation of autolysin production. We have shown that SpdC inhibits the WalKR-dependent synthesis of four peptidoglycan hydrolases, SceD, SsaA, LytM and AtlA, as well as impacting S. aureus resistance towards lysostaphin and cell wall antibiotics such as oxacillin and tunicamycin. We have also shown that SpdC is required for S. aureus biofilm formation and virulence in a murine septicemia model. Using protein-protein interactions in E. coli as well as subcellular localization in S. aureus, we showed that SpdC and the WalK kinase are both localized at the division septum and that the two proteins interact. In addition to WalK, our results indicate that SpdC also interacts with nine other S. aureus histidine kinases, suggesting that this membrane protein may act as a global regulator of TCS activity. Indeed, using RNA-Seq analysis, we showed that SpdC controls the expression of approximately one hundred genes in S. aureus, many of which belong to TCS regulons.
Author summary
Staphylococcus aureus is a major human pathogen, and has become a significant worldwide health concern due to the rapid emergence of antibiotic resistant strains. Like most bacteria, S. aureus adapts to its environment by adjusting its genetic expression through sensing and regulatory systems. We show here that the SpdC membrane protein is a novel virulence factor of S. aureus, controlling biofilm formation and pathogenesis. We show that SpdC interacts with the WalK histidine kinase to inhibit activity of the WalKR two-component system. SpdC also interacts with nine other histidine kinases of S. aureus, suggesting it acts as a pleiotropic global regulator.
Introduction
Two-component systems (TCSs) are composed of a histidine kinase, usually membrane-bound and acting as an environmental sensor, which phosphorylates a coupled response regulator, often controlling gene transcription. Although these systems have been extensively studied and play an essential role in bacterial adaptation to the environment, the signal(s) to which they respond and additional factors positively or negatively controlling their activities remain mostly unknown. Staphylococcus aureus, a major human pathogen, causes diseases ranging from superficial cutaneous abscesses to life-threatening infections affecting all major organs [1]. S. aureus is also a commensal bacterium, colonizing approximately half the human population asymptomatically, essentially within the anterior nares [2]. In addition to its considerable arsenal of virulence factors, S. aureus must rapidly adapt to environmental conditions encountered during host colonization. Among the 16 TCSs encoded by the S. aureus genome [3], the SaeSR, AgrCA, and WalKR systems are particularly important for controlling virulence and innate immune evasion factors [4–8].
The WalKR system is the only S. aureus TCS shown to be essential for cell viability, suggesting that its activity may respond not only to environmental conditions but could also be controlled by intrinsic bacterial factors [9–11]. Indeed, it is becoming increasingly apparent that so-called two-component systems frequently involve additional proteins regulating the phosphorylation levels of the response regulator [12, 13]. These include accessory phosphatases such as CheZ, Spo0E or RapA, respectively dephosphorylating the E. coli CheY [14], B. subtilis Spo0A [15] and Spo0F [16] response regulators, or antikinases such as KipI and Sda that inhibit B. subtilis KinA [17, 18] and FixT which inhibits the Sinorhizobium meliloti FixJ kinase [19]. A sub-class of histidine kinases, known as intra-membrane sensing kinases [20], require the permease component of an associated ABC transporter for signal sensing, such as the S. aureus BraS [21] and GraS [22] kinases. Many TCS histidine kinases act as so-called bifunctional sensors, acting on their cognate response regulators both as kinases and phosphoprotein phosphatases [23]. Accordingly, several accessory proteins act by binding to the histidine kinase to inhibit its kinase activity or stimulate its phosphatase activity towards the response regulator. These include the PII protein acting on the NtrB kinase to control nitrogen assimilation by dephosphorylating NtrC [24], the Streptococcus agalactiae Abx1 Abi-domain membrane protein which interacts with the CovS histidine kinase to inhibit activity of the CovR response regulator [25], and the SaePQ protein complex, which stimulates the phosphatase activity of the SaeS histidine kinase in S. aureus [26].
In Bacillus subtilis, the WalK histidine kinase is thought to coordinate cell wall plasticity with cell division, with two membrane-bound accessory proteins, WalH and WalI, inhibiting WalK kinase activity [11, 27–29]. However, the WalH and WalI proteins of S. aureus share no significant sequence similarities with those of B. subtilis and their role is not as clear-cut. Indeed, although they are also membrane proteins and interact with the WalK kinase, WalH and WalI do not seem to play a major role in negatively controlling WalKR activity, suggesting that their functions have evolved [30].
In an effort to identify additional factors controlling the WalKR system, we showed that the S. aureus SpdC Abi-domain protein negatively affects WalK activity and expression of WalKR-regulated genes. We showed that SpdC, previously identified as playing a role in the display of surface proteins [31], forms a complex with the WalK histidine kinase and that the two membrane proteins preferentially localize at the division septum, suggesting that this interaction regulates WalK histidine kinase activity. The ΔspdC mutant displays a pleiotropic phenotype, including altered resistance towards compounds targeting the cell wall, as well as strongly diminished biofilm formation and virulence. Using RNA-Seq analysis, we showed that SpdC controls the expression of approximately one hundred genes in S. aureus. Indeed, SpdC activity appears to extend well beyond the WalKR system, since we have shown it also interacts with several other S. aureus histidine kinases suggesting it could be involved in controlling multiple regulatory/adaptive pathways.
Results
Expression of spdC is controlled by the WalKR system
We previously performed an extensive transcriptome analysis in order to define the scope of the S. aureus WalKR regulon [6]. Our results showed that expression of the spdC gene (SAOUHSC_02611) was increased 3.5-fold in a S. aureus strain producing a constitutively active form of the WalR response regulator (D55E) [6]. SpdC, a membrane-anchored protein with 8 predicted transmembrane segments and an Abi domain (CAAX protease self-immunity), (Fig 1A), was previously reported as playing a role in the display of surface proteins such as protein A [31]. In order to confirm that spdC is a member of the WalKR regulon, we used quantitative real-time PCR (qRT-PCR) to measure its expression in a S. aureus strain where the walRKHI operon is placed under the control of the IPTG-inducible Pspac promoter [9]. Cells were grown overnight in TSB with 0.05 mM IPTG, and cultures were inoculated at OD600nm = 0.05, with or without different IPTG concentrations (0.05 and 1 mM) to induce expression from the Pspac promoter. RNA samples were prepared from exponentially growing cells harvested at OD600nm = 0.5, before cessation of growth of the culture lacking IPTG, and walR and spdC mRNA levels were measured by qRT-PCR. As shown in Fig 1B, walR transcription is increased approximately 4-fold when cells are grown with 0.05 mM IPTG and 8-fold at 1 mM IPTG. Under the same conditions, spdC expression followed that of walR, and was increased 2-fold and 5-fold with 0.05 or 1 mM IPTG, respectively (Fig 1B), confirming positive regulation by the WalKR system.
Fig 1. The WalKR two-component system controls expression of spdC, encoding a Abi-domain membrane protein.

A. Predicted membrane topology of the SpdC protein generated using the Protter prediction tool (http://wlab.ethz.ch/protter/start/). The eight predicted transmembrane segments are numbered. Amino acids forming the putative Abi domain (based on Pfam database annotation) are shaded in green. B. Relative levels of walR and spdC transcripts were measured by qRT-PCR during growth of the HG001 PspacwalRKHI strain (ST1017). Bacteria were grown to exponential phase in TSB with or without increasing concentrations of IPTG to induce expression from the Pspac promoter. Expression levels were normalized using 16S rRNA as an internal standard and are indicated as n-fold change with respect to the control condition (absence of IPTG).
SpdC is a pleiotropic regulator of gene expression
As shown above, the WalKR TCS controls spdC expression. In Streptococcus agalactiae, another bacterial Abi-domain protein, Abx1, has been shown to inhibit activity of the CovSR two-component system [25]. In order to determine whether SpdC has a regulatory role in S. aureus, we generated a ΔspdC mutant in strain HG001 and performed a comparative RNA-Seq analysis. The ΔspdC mutant did not display any gross morphological changes or growth defects. Indeed, although it had a slight lag during the first hour post inoculation, the growth rate and final OD600nm were not significantly different from those of the parental strain (S1 Fig). Doubling times (http://www.doubling-time.com/compute.php) calculated during the exponential growth phase (S1 Fig, 90 min to 210 min) gave identical values of 32 min for both strains. The HG001 strain and ΔspdC mutant were grown in TSB until early exponential phase (OD600nm = 1) and total RNA was extracted for RNA-Seq analysis (See Materials and Methods). We verified that spdC is well expressed under these conditions using a lacZ reporter fusion with the spdC promoter region (S2 Fig).
Three biological replicates were analyzed by RNA-Seq for each strain and the data presented as the mean fold-change. Using a value cut-off greater than 2 with a P value less than 0.05, we found that the expression of 42 genes was lowered in the ΔspdC mutant strain and that of 65 increased (Table 1). In order to perform a general analysis of the transcriptomic data we generated an ontological grouping of SpdC-regulated genes (Fig 2A). Among the genes positively controlled by SpdC, 10 are known virulence factors, suggesting that SpdC may influence S. aureus pathogenicity, and 11 are involved in capsular biosynthesis. The S. aureus capsule is known to impede phagocytosis and promote host colonization, however although the HG001 strain used in this study carries the serotype 5 capsule gene cluster, a missense mutation in the cap5E gene prevents capsular biosynthesis [32–34].
Table 1. Genes differentially expressed in the ΔspdC mutant compared to the parental HG001 strain.
| SpdC-activated genes | |||
| Id(a) | Gene name/function | Fold change(b) | Also regulated by(c) |
| Virulence | |||
| 00069* | spa | -59.69 | VraSR, WalKR |
| 02709* | hlgC | -5.53 | SaeSR, WalKR |
| 02710* | hlgB | -4.97 | ArlSR, GraSR, SaeSR, WalKR |
| 00545* | sdrD | -4.19 | ArlSR, GraSR, AirSR |
| 02169* | chp | -2.86 | SaeSR, GraSR, WalKR |
| 01939* | splC | -2.16 | BraSR, SaeSR, WalKR |
| 01941* | splB | -2.49 | ArlSR, SaeSR, WalKR |
| 02171* | sak | -2.03 | SaeSR |
| 01112* | formyl peptide receptor-like 1 inhibitory prot. | -2.23 | SaeSR, WalKR |
| 01110* | fibrinogen-binding prot. | -2.04 | SaeSR, WalKR |
| Regulators | |||
| 00070* | sarS | -2.55 | WalKR, GraSR, BraSR |
| 00961 | comK | -2.12 | |
| 01879 | rot | -2.03 | ArlSR, GraSR, |
| ABC transporters | |||
| 02820 | ABC transporter ATP binding prot. | -5.94 | VraSR,GraSR, BraSR |
| 02821 | ABC transporter permease | -5.27 | ArlSR, GraSR, BraSR |
| 01389 | PstS phosphate ABC transporter | -3.39 | |
| Amino acid metabolism | |||
| 01452 | alanine dehydrogenase | -4.29 | ArlSR, SaeSR, GraSR, BraSR, |
| 01451* | threonine dehydratase | -5.32 | ArlSR, SaeSR, GraSR, BraSR, WalKR |
| 01450 | putative AA permease | -4.42 | |
| Carbohydrate metabolism | |||
| 00051 | 1-phosphatidylinositol phosphodiesterase | -3.27 | |
| 00187 | formate acetyl transferase | -2.92 | SaeSR |
| 00188* | pyruvate formate-lyase 1 activating enzyme | -3.04 | SaeSR, WalKR |
| 00608 | alcohol dehydrogenase | -2.75 | SaeSR |
| 00113 | bifunctional acetaldehyde-CoA/alcohol dehydrogenase | -2.72 | BraSR, GraSR |
| 02830* | D-lactate dehydrogenase | -2.20 | GraSR, WalKR |
| 02922 | L-lactate dehydrogenase | -2.11 | BraSR |
| Other | |||
| 01448 | putative transporter/multidrug resistance prot. | -3.31 | ArlSR, GraSR, SaeSR |
| 02108 | FtnA ferritin | -2.21 | SaeSR, GraSR |
| 02862* | clpC | -2.15 | WalKR |
| 02127 | staphopain thiol protease | -2.05 | |
| 00625 | monovalent cation/H+ antiporter subunit A | -2.02 | |
| 02941 | anaerobic ribonucleoside-triphosphate reductase activating prot. (nrdG) | -2.08 | |
| HP without homology | |||
| 02781 | -2.85 | GraSR | |
| 00966 | -2.82 | ||
| 00203 | -2.61 | BraSR | |
| 01123 | -2.52 | ||
| 02782 | -2.51 | ||
| 00094* | -2.31 | WalKR, BraSR | |
| 02294 | -2.20 | ||
| 02973 | -2.17 | ||
| 02842 | -2.14 | GraSR, WalKR | |
| 01798 | -2.10 | BraSR | |
| SpdC-repressed genes | |||
| Id(a) | Gene name/function | Fold change(b) | Also regulated by(c) |
| Cell wall metabolism | |||
| 02333* | sceD | 10.80 | GraSR, WalKR |
| 02576* | ssaA | 5.64 | GraSR, WalKR, SaeSR |
| 00248* | lytM | 3.81 | WalKR |
| 00994* | atlA | 2.21 | GraSR, WalKR |
| ABC transporters | |||
| 02430 | ABC transporter substrate-binding prot. | 2.69 | GraSR |
| 02153 | ABC-2 transporter family prot. | 2.20 | |
| 02154 | ABC transporter ATP-binding prot. | 2.19 | |
| 02152 | ABC transporter ATP-binding prot. | 2.13 | |
| Φ12 phage proteins | |||
| 01535 | HP | 4.00 | |
| 01573 | HP | 3.02 | |
| 01538 | phage terminase large subunit | 2.93 | |
| 01536 | scaffolding protease | 2.66 | |
| 01515 | peptidoglycan hydrolase | 2.66 | |
| 01516 | holin prot. | 2.63 | |
| 01530 | HP | 2.60 | |
| 01532 | SLT orf 110-like prot. | 2.50 | |
| 01570 | PVL orf 37-like prot. | 2.43 | |
| 01566 | phi APSE P51-like prot. | 2.43 | |
| 01531 | SLT orf 123-like prot. | 2.42 | |
| 01519 | SLT orf 129-like prot. | 2.37 | |
| 01520 | SLT orf 488-like prot. | 2.33 | |
| 01571 | SLT orf 71-like prot. | 2.28 | |
| 01533 | HP | 2.17 | |
| 01527 | HP | 2.13 | |
| 01529 | major tail prot. | 2.03 | |
| Φ13 phage proteins | |||
| 02234 | repressor-like prot. | 4.83 | |
| 02233 | phi PVL orf 32-like prot. | 3.77 | |
| 02221 | HP | 3.19 | |
| 02220 | phi ETA orf 18-like prot. | 2.71 | |
| 02228 | HP | 2.61 | |
| 02211 | phi PVL orf 50-like prot. | 2.58 | |
| 02225 | HP | 2.26 | |
| 02219 | phi ETA orf 20-like prot. | 2.21 | |
| 02216 | phage DnaC-like prot. | 2.20 | |
| 02223 | phi PVL orf 39-like prot. | 2.06 | |
| 02222 | HP | 2.05 | |
| Nucleotide metabolism | |||
| 01172* | PyrE orotate phosphoribosyltransferase | 8.02 | SaeSR, WalKR |
| 01171* | orotidine 5'-phosphate decarboxylase | 7.99 | SaeSR, WalKR |
| 01170* | CarB carbamoyl phosphate synthase large subunit | 7.04 | SaeSR, WalKR, BraSR |
| 01166* | pyrB | 3.53 | SaeSR, WalKR |
| 01017 | purH | 2.19 | GraSR, SaeSR |
| 02558 | UreA urease subunit gamma | 2.14 | ArlSR, GraSR , SaeSR |
| 01016 | phosphoribosylglycinamide formyltransferase | 2.12 | GraSR, SaeSR |
| 01018 | phosphoribosylamine—glycine ligase | 2.07 | GraSR, SaeSR |
| 01015 | phosphoribosylaminoimidazole synthetase | 2.00 | GraSR, SaeSR |
| Nitrogen metabolism | |||
| 02685 | nitrite reductase transcriptional regulator NirR | 2.48 | BraSR |
| 02684 | assimilatory nitrite reductase [NAD(P)H] large subunit | 2.03 | VraSR |
| 02681 | nitrate reductase subunit alpha | 2.03 | BraSR |
| Amino acid metabolism | |||
| 01367 | anthranilate synthase component II | 2.49 | |
| Capsular biosynthesis | |||
| 00116 | Cap5C | 2.40 | |
| 00117 | Cap5D | 2.37 | BraSR, ArlSR |
| 00122 | Cap5I | 2.32 | BraSR, GraSR |
| 00114 | Cap5A | 2.28 | ArlSR |
| 00115 | Cap5B | 2.28 | |
| 00128 | Cap5O | 2.19 | |
| 00120 | Cap5G | 2.17 | BraSR |
| 00121 | Cap5H | 2.13 | BraSR, GraSR |
| 00123 | Cap5J | 2.12 | GraSR |
| 00125 | Cap5L | 2.09 | BraSR |
| 00126 | Cap5M | 2.03 | |
| HP without homology | |||
| 01173 | 3.70 | SaeRS | |
| 01583 | 2.75 | BraSR, SaeSR | |
| 01084 | 2.71 | ||
| 01584 | 2.54 | ||
| 02863 | 2.06 | ||
*: Genes also regulated by the WalKR system
(a): Id indicates the S. aureus NCTC 8325 genome sequence SAOUHSC _ annotation.
(b): Fold change was determined as the ratio of the signal values between the ΔsdpC strain and the parental strain. Negative values indicate the gene is less expressed in the ΔspdC mutant strain and positive values that it is more highly expressed in the mutant strain. Only fold changes >2 with a P value <0.05 for the three biological replicates were considered.
(c): SpdC-regulated genes identified by RNA-Seq data analysis were compared with publicly available transcriptome analyses of S. aureus TCS mutants (http://www.satmdorg/) and TCSs with which SpdC was shown to interact are listed. Names in bold indicate positive regulation by the TCS in question as opposed to negative regulation.
Fig 2. SpdC is involved in gene regulation.

A. Ontological grouping of SpdC-regulated genes according to their annotated functions. Variations in gene expression were identified by RNA-Seq analysis of a ΔspdC strain compared to the HG001 parental strain grown in TSB until mid-exponential phase. B. qRT-PCR comparison of gene expression in a ΔspdC strain and the HG001 parental strain. Strains were grown in TSB until mid-exponential phase and RNA was extracted and treated as described in Materials and Methods. Expression levels were normalized using 16S rRNA as internal standard and presented as the n-fold change of the ΔspdC mutant strain compared to the HG001 parental strain. C. Western blot analysis of LytM (upper panel) and Spa (lower panel) production. Crude extracts (LytM) and cell wall extracts (Spa) were prepared from stationary phase cultures. Lanes: 1: Purified Staphylococcus aureus Protein A (50 ng) 2: HG001 strain; 3: ΔspdC mutant strain; 4: ΔspdC/pMK4Pprot-spdC complemented strain.
Expression of several S. aureus prophage genes was also increased in the ΔspdC mutant strain: 11 for phage Φ13 and 17 for Φ12, (Table 1).
SpdC negatively controls WalKR regulon genes
As shown above, spdC expression is controlled by the WalKR system. The RNA-Seq data analysis of SpdC-regulated genes reveals that 25 of these belong to the WalKR regulon (indicated by an asterisk in Table 1). In particular, the expression of 4 WalKR-dependent cell wall hydrolase genes (sceD, ssaA, lytM and atlA) is increased in the ΔspdC mutant, suggesting that SpdC negatively controls WalKR activity (Table 1). It is interesting to note that among the genes positively controlled by SpdC, many are also WalKR-activated genes (Table 1). All of these are classified as virulence genes, however they are not preceded by the WalR consensus binding site, and we have previously shown that several of these are not directly regulated by the WalKR system but through the SaeRS two-component system instead [6].
We used qRT-PCR to verify SpdC-dependent regulation for 3 positively (spa, hlgC, sdrD) and 3 negatively (atlA, sceD, lytM) controlled genes, in the ΔspdC strain compared to the HG001 parental strain, grown under the same conditions as for the RNA-Seq analysis (Fig 2B). We observed a perfect correlation with the RNA-Seq data: expression of the spa gene encoding protein A was very strongly lowered in the ΔspdC mutant strain (about one hundred-fold), while sdrD and hlgC expression levels were 5- to 10-fold less. The atlA, sceD and lytM cell wall hydrolase genes were more highly expressed in the absence of SpdC (2-, 12- and 5-fold, respectively) in agreement with the RNA-Seq analysis (Fig 2B).
In order to confirm SpdC-dependent regulation at the protein level, we chose two genes that were positively or negatively controlled by SpdC, spa and lytM, respectively, and performed Western blot analyses. Whole cell extracts were prepared from cultures of strains HG001, the ΔspdC mutant and the complemented mutant strain (ΔspdC/pMK4Pprot-spdC) and subjected to SDS-PAGE and immunoblotting. As shown in the top panel of Fig 2C, LytM levels are higher in the ΔspdC strain compared to the parental strain (lane 2), and in the complemented strain the LytM level is reduced to a level lower than in the parental strain (lane 3), likely reflecting the higher production of SpdC in the complemented strain. Indeed, under these conditions, spdC mRNA levels were increased more than 100-fold as measured by qRT-PCR as compared to the parental HG001 strain.
For studying levels of protein A, known to be covalently anchored to the cell wall (LPxTG sortase motif), identical quantities of cell wall fractions of the HG001 parental strain, ΔspdC deletion mutant, and complemented strain (ΔspdC/pMK4Pprot-spdC) were subjected to SDS/PAGE and compared by Western blot. As expected, protein A levels were significantly lower in the ΔspdC mutant than in the parental and complemented strains (Fig 2C, lower panel), in agreement with the RNA-Seq and qRT-PCR results.
These data indicate that SpdC is involved in controlling gene expression through potential interactions with regulatory systems, and the WalKR two-component system in particular.
SpdC is localized at the division septum and interacts with the WalK histidine kinase
As shown above, we have uncovered a regulatory link indicating that SpdC negatively controls activity of the WalKR two-component system, strongly suggesting that the proteins interact. In order to test possible interactions between SpdC and the WalKR proteins, we used the bacterial adenylate cyclase two-hybrid system (BACTH) [35]. We fused the full-length membrane-bound WalK histidine kinase or the WalR cytoplasmic response regulator to the C-terminal domain of the T25 subunit of the Bordetella pertussis adenylate cyclase and full-length SpdC to the C-terminal domain of the T18 subunit, using plasmids pKT25 and pUT18c respectively. To probe putative interactions, E. coli strain DHT1 was co-transformed with combinations of the pKT25 and pUT18c derivatives carrying the translational fusions. Upon protein-protein interactions, the close proximity between the T18 and T25 subunits restores adenylate cyclase activity, leading to cAMP synthesis and activation of the lactose operon. Interactions were tested both by spotting the resulting strains on LB plates containing X-Gal and by measuring β-galactosidase activity. To determine pair-wise interactions, we chose a cut-off value of 100 Miller Units as indicating a positive interaction between the protein fusions. As shown in Fig 3A, strong β-galactosidase activity was only observed for the plasmid combination co-producing the membrane anchored proteins SpdC and WalK while no interactions between SpdC and the cytoplasmic regulator WalR could be detected. SpdC is annotated as an Abi domain protein (CAAX protease self-immunity) in genome databases. The S. aureus HG001 genome encodes 4 Abi domain proteins, three of which, SpdA (SAOUHSC_01900), SpdB (SAOUHSC_02587), and SpdC, have been reported as being involved in surface protein display, whereas the fourth (SAOUHSC_02256) has no known function [31]. In order to test whether WalK also interacts with the other three Abi domain proteins, we constructed translational fusions for the remaining Abi proteins with the T18 domain of adenylate cyclase. As shown in Fig 3A, the combinations of SpdA, SpdB or SAOUHSC_02256 with WalK did not generate significant levels of β-galactosidase activity, demonstrating that SpdC is the only S. aureus Abi domain protein specifically interacting with WalK.
Fig 3. The SpdC membrane protein forms a complex with the WalK histidine kinase at the division septum.

A: The SpdC protein was tested for pairwise interactions with the WalK histidine kinase and the WalR response regulator using the BACTH complementation assay by translationally fusing the full length corresponding coding sequences with those of the T25 or T18 adenylate cyclase domains (upper left panel). Specificity of the SpdC-WalK complex was confirmed by testing interactions with three other S. aureus Abi-domain proteins: SpdB, SpdA and SAOUHSC_02256 (lower left panel). Cultures of E. coli DHT1 strains co-transformed with the indicated plasmid combinations were spotted on LB agar plates with X-Gal as an indicator of β-galactosidase activity (see Materials and Methods). Quantitative β-galactosidase activity assays for each strain are indicated in the right panel (expressed in Miller Units). The shaded cell indicates significant β-galactosidase activity resulting from positive protein-protein interactions. ND: Not determined. B: S. aureus HG001 cells producing a fluorescent SpdC-GFP protein fusion (HG001/pOLSA-spdC) were grown in TSB with 0.25 μM CdCl2 and observed in exponential phase by fluorescence microscopy. Fluorescence ratios (septum/lateral membrane) were quantified for two biological replicates (33 cells from each) using ImageJ software and plotted using GraphPad Prism. The horizontal line indicates the mean fluorescence ratio and the error bars represent the SEM.
We have previously shown that the S. aureus WalK histidine kinase is mainly localized at the division septum [30]. Since SpdC and WalK interact, we studied the subcellular localization of SpdC in S. aureus by constructing a translational fusion with the GFP fluorescent protein using the pOLSA vector (See Materials and Methods), under the control of the cadmium-inducible Pcad promoter. The resulting plasmid, pOLSA-spdC was then introduced into the HG001 strain. Expression of the gene fusion was induced by addition of CdCl2 (0.25 μM), cells were harvested during exponential growth (OD600nm = 1.5) and observed by fluorescence microscopy. As shown in Fig 3B, SpdC is preferentially localized at the division septum, with a mean septum/membrane fluorescence ratio of around 2.6. Taken together, our results indicate that SpdC and WalK interact and are localized at the division septum.
SpdC is involved in S. aureus resistance against compounds targeting the cell envelope
As shown above, the expression of several cell wall hydrolase genes is significantly increased in the ΔspdC mutant strain, suggesting that sensitivity to compounds targeting the cell wall might also be affected. We followed bacterial lysis during incubation in the presence of a non-anionic detergent, Triton X-100, thought to trigger cell lysis by favoring endogenous autolysin activity [36]. However, Triton X-100 induced lysis for the HG001 and ΔspdC mutant strains was not significantly different (S3 Fig).
We also tested sensitivity to lysostaphin, a glycyl-glycine endopeptidase that hydrolyzes the peptidoglycan pentaglycine interpeptide crossbridge, leading to cell lysis [37]. The HG001, ΔspdC mutant and complemented strains were grown in TSB until OD600nm ≈ 1 and cells were then harvested and resuspended in PBS in the presence of lysostaphin. As shown in Fig 4A, the ΔspdC mutant was less sensitive to lysostaphin-induced lysis than the parental HG001 and complemented ΔspdC strains, suggesting that the absence of SpdC leads to cell wall modifications.
Fig 4. SpdC impacts cell wall homeostasis.

A. The ΔspdC mutant displays increased resistance to lysostaphin-induced lysis. Cells were grown in TSB until mid-exponential phase, harvested and incubated in PBS with lysostaphin (200 ng/ml) with aeration at 37°C. Bacterial lysis was measured by monitoring OD600nm over time. Results are shown as the mean and standard deviation of three independent assays. HG001 parental strain ( ); ΔspdC mutant (■); ΔspdC/pMK4Pprot-spdC complemented strain (▲). B. The absence of SpdC leads to sensitivity to oxacillin and tunicamycin. Dilution series of the HG001, ΔspdC and ΔspdC/ pMK4Pprot-spdC strains on TSA plates with or without antibiotics. Oxacillin: 0.1 μg/ml; fosfomycin: 4 μg/ml; tunicamycin: 1 μg/ml.
); ΔspdC mutant (■); ΔspdC/pMK4Pprot-spdC complemented strain (▲). B. The absence of SpdC leads to sensitivity to oxacillin and tunicamycin. Dilution series of the HG001, ΔspdC and ΔspdC/ pMK4Pprot-spdC strains on TSA plates with or without antibiotics. Oxacillin: 0.1 μg/ml; fosfomycin: 4 μg/ml; tunicamycin: 1 μg/ml.
We then tested sensitivity to antibiotics targeting the cell wall. As shown in Fig 4B the ΔspdC mutant displayed increased sensitivity to the β-lactam antibiotic oxacillin, whereas the parental and complemented strains were able to grow at the concentration tested (0.1 μg/ml). No difference in sensitivity between the strains was seen using fosfomycin, an antibiotic inhibiting MurA, which catalyzes the very first step of peptidoglycan biosynthesis (Fig 4B). These results suggested that the ΔspdC mutant strain may either be affected in the later steps of peptidoglycan biosynthesis or may exhibit a cell wall structure modification leading to a difference in accessibility of antibiotics acting extracellularly. Wall teichoic acids (WTAs) are anionic sugar rich cell surface polymers that can alter accessibility to the cell wall. We therefore tested resistance to tunicamycin, an antibiotic targeting biosynthesis of WTAs. As shown in Fig 4B the ΔspdC mutant strain was highly sensitive to tunicamycin, in contrast to the parental and complemented strains.
Taken together, these results suggest that the S. aureus cell envelope structure is altered in the absence of SpdC.
SpdC is required for biofilm formation
Since our results indicate that the ΔspdC mutation may modify the S. aureus cell surface, we tested whether the absence of SpdC may have an effect on biofilm formation. Strains were grown statically in TSB, supplemented with glucose and NaCl, for 24 h at 37°C in PVC microplates. As shown in Fig 5, biofilm formation was strongly decreased in the absence of SpdC (approximately 7-fold). Complementation of the ΔspdC mutant with the pMK4Pprot-spdC plasmid restored biofilm formation to levels comparable to those of the parental HG001 strain (Fig 5). These results are consistent with a modification of the S. aureus cell surface in the absence of SpdC, which could influence resistance against antimicrobial compounds targeting cell surface structures as well as the capacity to form biofilms. In order to determine which biofilm component is affected, biofilm detachment experiments were carried out (S4 Fig) by treatment with proteinase K, DNaseI and sodium metaperiodate (a carbohydrate-modifying agent). Under our conditions, biofilm production was lowered three-fold after treatment with DNaseI, and more than 10-fold when treated with proteinase K, but not significantly modified after treatment with sodium metaperiodate. Thus, biofilms formed under our conditions by the HG001 parental strain are essentially protein-based, and, to a lesser extent, due to extracellular DNA. We observed reduced biofilm formation for the ΔspdC mutant even after DNAseI treatment, but not after proteinase K treatment (S4 Fig), suggesting that SpdC affects the production of proteins important for biofilm formation.
Fig 5. SpdC is required for biofilm formation.

Biofilm assays were performed by growing cells in static cultures in PVC microtiter plates (TSB plus 0.75% glucose and 3.5% NaCl). Plates were incubated at 37°C for 24h and adherent biomass was quantified as described in Materials and Methods. Data were normalized to the OD600nm of each culture and indicated as the n-fold change of the ΔspdC mutant strain compared to the HG001 parental strain. ***, P<0.0005 (Student’s t-test).
SpdC is a novel virulence factor
Cell surface modifications are known to impact virulence. Likewise, the capacity to form robust biofilms favors bacterial colonization of the host. Additionally, our RNA-Seq analysis revealed that the expression of at least 10 genes directly involved in bacterial virulence upon infection was lowered in the ΔspdC mutant, strongly suggesting that SpdC plays a role in virulence. We used a murine sepsis model to compare virulence of the HG001 and ΔspdC strains. SWISS mice were infected intravenously with 5.107 cfu and mortality was monitored over 9 days post infection. As shown in Fig 6 virulence of the ΔspdC mutant was strongly diminished. Indeed, following infection with the HG001 parental strain, significant mortality occurred in the first 3 days post-infection (greater than 60%), whereas only a single mouse out of 14 died within five days after infection with the ΔspdC mutant. After the sixth day, a moderate mortality was observed for the group infected with the ΔspdC mutant, with only 36% mortality at the end of the assay (compared to 72% mortality for mice infected with the parental HG001 strain). This significant difference indicates that SpdC is a novel virulence factor in S. aureus.
Fig 6. SpdC is a novel S. aureus virulence factor.
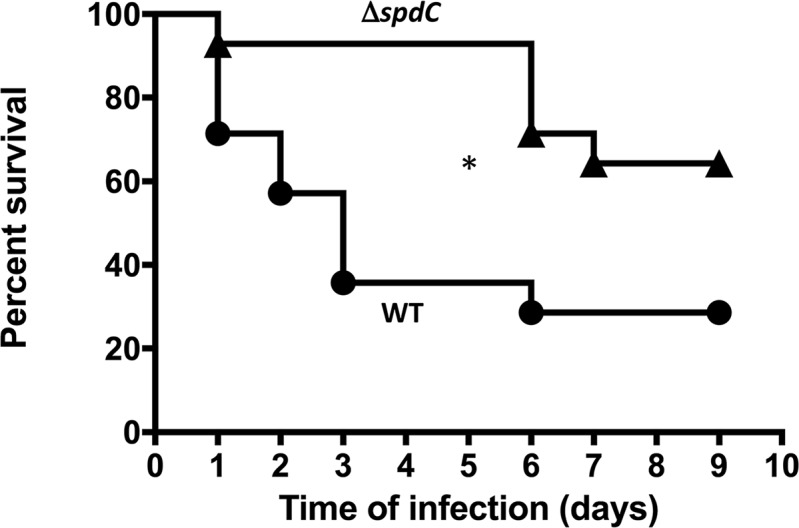
Kaplan-Meier survival curves of RjOrl:SWISS mice infected with either the HG001 parental strain (circles) or the ΔspdC strain (triangles) by i.v. route (5.107 cfu/injection). A total of 14 mice were used in each group in two independent experiments. *, P<0.05 (Wilcoxon test).
SpdC interacts with several histidine kinases through their transmembrane domains
As shown above, SpdC localizes at the division septum and interacts with the WalK histidine kinase, negatively controlling WalKR activity. However, many of the SpdC-regulated genes identified by RNA-Seq do not belong to the WalKR regulon, but are known to be controlled by other two-component systems (Table 1) suggesting SpdC may interact with other TCS regulatory pathways. The S. aureus HG001 genome encodes 16 two-component systems [33] and we constructed translational fusions for each of the histidine kinase genes with the carboxy-terminal region of the adenylate cyclase T25 domain. Each of the resulting plasmids was co-transformed in combination with the pUT18c-spdC plasmid into E. coli strain DHT1. As shown in Fig 7, in addition to WalK, we detected interactions between SpdC and nine additional histidine kinases: YesM, GraS, SaeS, DesK, ArlS, SrrB, PhoR, VraS, and BraS. These interactions appear to be specific, since no interactions were detected between SpdC and the remaining six histidine kinases, as shown in Fig 7 (LytS, AirS, AgrC, KdpD, HssS, and NreB).
Fig 7. Protein interaction network between SpdC and S. aureus histidine kinases.

The BACTH assay was used to test protein-protein interactions between SpdC and each of the S. aureus TCS histidine kinases. A. DHT1 E. coli strains co-transformed with each combination of plasmids (pUT18c-spdC and pKT25 derivatives carrying the genes for the designated kinases) were spotted on LB agar plates with X-gal as an indicator of β-galactosidase activity. Histidine kinases showing a positive result for interaction with SpdC are underlined. B. β-galactosidase activity of DHT1 strains co-transformed with the pUT18c-spdC and pKT25 derivatives carrying each of the histidine kinase encoding genes. β-galactosidase activity (Miller Units) of the strain carrying the empty vectors (pKT25-pUT18c) was arbitrarily fixed at 0 and that of the positive control (pKT25zip-pUT18czip) at 1000.
Among the genes positively regulated by SpdC (Table 1), the expression of at least fifteen is also activated by the SaeSR TCS, including the spl operon, sak, the hlgBC operon and chp [7, 38–40]. This suggests that SpdC activates the SaeSR TCS, in contrast to its role in negatively controlling activity of the WalKR system.
Histidine kinases have different combinations of signaling domains such as HAMP or PAS domains [41] in addition to the conserved H, N, G1, F and G2 boxes of the phosphoacceptor/dimerization (HisKA) and catalytic (HATPase_C) domains [42–44]. We have shown that SpdC interacts with 10 of the 16 S. aureus histidine kinases, which do not share any strong amino acid sequence similarities other than the conserved histidine kinase HisKA and HATPase_C domains. Since SpdC interacts with some, but not all of the S. aureus histidine kinases, this protein-protein contact must involve some other domain.
We focused our analysis on the WalK and SaeS histidine kinases. WalK has two transmembrane domains (amino acids 14–34 and 183–203), with an extracellular loop of 148 amino acid residues, a HAMP domain involved in signal transduction (204–256), a PAS domain (261–331) a PAC domain (314–378) and a histidine kinase domain (382–600; Fig 8A). In contrast, SaeS is a member of the intra-membrane sensing kinases [45], with two transmembrane domains (amino acids 9–29 and 40–60) separated by only ten amino acids, as well as a HAMP domain (61–114) and a histidine kinase domain (129–348; Fig 8A). We tested the interactions of SpdC with the N-terminal domains of WalK and SaeS containing the transmembrane regions (WalK1-203 and SaeS1-64, respectively). The truncated proteins were fused to the T25 domain of adenylate cyclase, and the resulting plasmids were co-transformed into E. coli strain DHT1 together with the pUT18c-spdC plasmid. As shown in Fig 8B, the first 203 amino acids of WalK are sufficient to allow stable interactions with SpdC. For SaeS, the N-terminal domain containing only the two transmembrane segments (SaeS1-64) did not lead to interaction with SpdC, but a longer fragment of the protein (SaeS1-120) gave rise to a stable interaction with SpdC and high β-galactosidase activity (Fig 8B). These results suggest that WalK and SaeS interact with SpdC through their transmembrane domains. The negative interaction results obtained with the truncated SaeS protein containing only the transmembrane domains suggest that since this kinase lacks an extracellular loop, the HAMP domain may be required for proper membrane insertion of the fusion protein.
Fig 8. Domains involved in interaction between SpdC and the WalK and SaeS histidine kinases.

A. Domains fused to either pUT18c or pKT25 for the BACTH assay. Hatched bars indicate transmembrane regions and functional domains are annotated according to the UniProt database (http://www.uniprot.org/uniprot/Q2G2U4 for WalK, http://www.uniprot.org/uniprot/Q2G2U1 for SaeS and http://www.uniprot.org/uniprot/Q2FVT1 for SpdC). B. β-galactosidase activities of DHT1 E. coli strains co-transformed with pUT18c containing either the full-length spdC coding sequence or a fragment encoding only the first 252 amino-acids of SpdC, and pKT25 derivatives carrying DNA fragments encoding the indicated proteins. Positive interaction results are shaded. ND: Not determined.
In order to identify which domain of SpdC interacts with the WalK and SaeS kinases, we compared interactions with full-length SpdC (pUT18c-spdC) and a carboxy-terminal truncated SpdC consisting only of the eight transmembrane domains (SpdC1-252; pUT18c-spdC1-252). We noted self-interaction of SpdC following co-transformation of pKT25-spdC with either pUT18c-spdC or pUT18c-spdC1-252 (Fig 8B), indicating that the SpdC transmembrane domains are involved in these self-interactions. Similar results were obtained when testing interactions with WalK and SaeS, i.e. the transmembrane domains of SpdC are sufficient to allow interactions with the histidine kinases (Fig 8B).
Taken together, these results indicate that SpdC is a membrane-bound protein that interacts with itself and several histidine kinases through transmembrane domain contacts.
Discussion
Abi-domain proteins constitute a large family whose functions are mostly unknown. SpdC was initially designated LyrA for Lysostaphin resistance A and identified by screening a bursa aurealis transposon mutant library for increased lysostaphin resistance [46]. An independent study aiming at characterizing proteins involved in the display of surface proteins led to the identification of three proteins, SpdA, SpdB and SpdC (Surface protein display A, B and C), playing a role in protein A levels at the staphylococcal cell surface [31]. These proteins share a similar structural organization with 6 to 8 transmembrane domains and an Abi-domain embedded within the hydrophobic region. An additional protein with a similar organization is encoded by the S. aureus genome, SAOUHSC_02256, but appears to have no role in controlling protein A levels [31].
We previously characterized the essential WalKR two-component system in S. aureus and highlighted its major role in controlling cell wall homeostasis [9, 47]. Transcriptome analysis revealed that spdC expression is positively controlled by WalKR [6]. We show here that the SpdC membrane protein and the WalK histidine kinase interact and that SpdC negatively controls WalKR activity and expression of WalKR-regulated genes. Interaction of the WalK histidine kinase with SpdC is specific since no interaction was seen with the other Abi-proteins (SpdA, SpdB and SAOUHSC_02256). Accordingly, we also showed that SpdC and WalK are both localized preferentially at the division septum.
Interestingly, an RNA-Seq analysis of a ΔsdpC mutant revealed that the expression of 107 genes varied compared to the parental strain. Among these, 24 (more than 20%) are controlled by the WalKR system. Since SpdC appears to negatively control WalKR activity by interacting with WalK, this septal localization is consistent with the previously suggested cell wall metabolism-related activation signal of the WalK histidine kinase in Bacillus subtilis [11, 48]. Indeed, in cocci, cell wall synthesis has been shown to exclusively occur at the division septum in an FtsZ-dependent manner [49], suggesting that a peptidoglycan metabolism related signal at the septum may relieve negative control of WalK activity by SpdC.
Histidine kinases often act as phosphoprotein phosphatases towards their associated response regulator. WalK was previously classified as a kinase/phosphatase «bifunctional sensor» [23] and the PAS domain of Streptococcus pneumoniae WalK plays a role in its phosphatase activity [50, 51]. In S. aureus, we have previously shown that WalK acts as a WalR phosphoprotein phosphatase upon entry into stationary phase in order to shut off WalR activity [6]. Interactions between SpdC and WalK can either interfere with signal perception by the sensor histidine kinase, inhibit its kinase activity or increase its phosphatase activity towards WalR, thus negatively controlling WalKR-dependent gene expression. SpdC is unlikely to directly regulate gene expression since it is a membrane protein lacking any typical DNA-binding domain.
To understand how the other SpdC-dependent genes were controlled (83 of the genes identified by RNA-Seq are not regulated by the WalKR system), we tested interactions of SpdC with the other S. aureus histidine kinases and found that it interacts with 10 of the 16 encoded in the genome. No obvious structural motifs or domain sequences were common to those that interacted with SpdC compared to those that did not. Two histidine kinases, WalK and SaeS, were chosen for further analysis of their interactions with SpdC. Our results indicate that the two transmembrane domains of WalK are sufficient to allow interaction with SpdC, whereas a greater amino-terminal fragment of SaeS was required, encompassing the HAMP domain. This result suggests that although the transmembrane domains of SaeS are likely involved in interactions with SpdC, a longer fragment may be necessary to ensure proper membrane insertion of the truncated protein. We also showed that a truncated form of SpdC, containing only the eight transmembrane domains, was sufficient for self-interaction and interaction with WalK and SaeS, indicating that transmembrane domains are involved in the interactions between SpdC and the histidine kinases. Interestingly, the only two S. aureus cytoplasmic histidine kinases which lack transmembrane domains, AirS and NreB, did not interact with SpdC under our conditions, in agreement with our results indicating transmembrane segments are involved in the interactions. The only other example of an Abi domain protein interacting with a histidine kinase is Abx1 of Streptococcus agalactiae, which interacts with the CovS kinase [25]. The two transmembrane domains of CovS were shown to be necessary and sufficient for these interactions [25].
At least fifteen genes belonging to the SaeSR regulon, including the spl operon, sak, the hlgBC operon and chp [7, 38–40], are also positively controlled by SpdC (Table 1), indicating that SpdC likely activates the SaeSR TCS, in contrast to its role in negatively controlling activity of the WalKR system. Since WalR controls spdC expression, this is consistent with our previous results showing that constitutive activation of WalR generates a signal leading to activation of the SaeSR TCS and a corresponding increase in SaeSR regulon expression [6].
The localization of SpdC at the division septum and its role in gene regulation through interactions with sensor kinases of two-component systems led us to speculate that SpdC may interfere with bacterial division sensing and impact cell wall metabolism. Accordingly, the ΔspdC mutant displays increased resistance against lysis when treated with lysostaphin, in agreement with the original phenotype characterized by transposon insertion [46]. Additionally, the absence of SpdC was reported to lead to increased cross wall abundance and thickness [31]. We tested sensitivity of the ΔspdC mutant to antibiotics targeting the cell wall. The ΔspdC mutant is highly sensitive to oxacillin and tunicamycin, but not to fosfomycin, which inhibits the first step of cell wall biosynthesis. In agreement with the sensitivity of the ΔspdC mutant to tunicamycin, which inhibits wall teichoic acid synthesis, the spdC gene was also identified as a candidate using a screen to identify synthetically lethal mutations with teichoic acid biosynthesis defects [52]. We have shown that expression of cell wall hydrolase genes is increased in the ΔspdC strain (sceD, ssaA, lytM, atlA). This may lead to increased cell wall degradation, which could explain the lowered resistance to oxacillin. This could also lead to the mutant’s increased sensitivity to tunicamycin. Indeed, teichoic acids are key elements for the proper localization of AtlA to the division septa, where cell wall biosynthesis takes place, since cell wall plasticity is essential for cell division [53]. In the presence of tunicamycin, the absence of wall teichoic acids results in a delocalized distribution of AtlA across the cell surface. Since atlA is more highly expressed in the ΔspdC stain, this could explain why this strain is more sensitive to tunicamycin.
Taken together with the strong links to the WalKR TCS, these results indicate that SpdC is involved in bacterial cell envelope homeostasis. The importance of the bacterial cell envelope in host-pathogen interactions cannot be over-emphasized: it is the first layer of contact between the bacterium and its host, containing an array of cell wall-linked or associated toxins and virulence factors, the first and major bacterial line of defense against threats from the host or environment, and is also both the target of choice for antibiotic treatment and the source of many antibiotic-resistance pathways. We have shown that SpdC is required for biofilm formation, an important step in S. aureus pathogenesis. We also showed that SpdC is a novel S. aureus virulence gene, required for the infectious process in a mouse septicemia model. The loss of virulence observed with the ΔspdC mutant may involve both its diminished capacity to form biofilms as well as the lowered expression of multiple virulence genes as shown by RNA-Seq analysis.
The regulatory mechanism mediated by SpdC remains to be determined. Abi-domain-containing proteins have been extensively studied in eukaryotes. They are involved in CAAX-protein maturation by cleaving the C-terminal AAX tripeptide after addition of an isoprenyl group on a cysteine, the last step consisting in methylation of the new C- terminus. These three modification steps are termed prenylation [54]. This post-translational maturation has a crucial role in maintaining cellular homeostasis by controlling the localization and activity of a large range of proteins. In particular, by adding a lipid group at the carboxy-terminal end of proteins, it favors their interactions with membranes, which have a high concentration of signaling molecules [55]. Prenylation has been recently described in prokaryotes and a geranyltransferase, IspA, has been identified in S. aureus [56]. Putative methyltransferase and CAAX-protease encoding genes (including spdC) are also present in the S. aureus genome. Deletion of the ispA gene has pleiotropic effects such as a growth defect, increased sensitivity to oxidative stress and an altered cell envelope [56]. Of note, the absence of IspA or SpdC both lead to increased cell wall antibiotic sensitivity. The RNA-Seq transcriptome analysis of the ΔispA strain [56] shows similarities with that of the ΔspdC mutant. Indeed, one of the most regulated genes in both cases is spa, encoding the immunoglobulin G binding protein A. We also noted that 21 Φ13 genes are up-regulated in the ΔispA mutant whereas we characterized 11 Φ13 genes up-regulated in the ΔspdC mutant, with several in common. These data suggest that SpdC and IspA could be involved in the same cellular pathway.
Interestingly, the transcriptome analysis of the ΔispA mutant revealed modified expression of a large number of genes involved in regulatory circuits and particularly increased expression of the walR, walH, walI and walJ genes of the wal locus [56]. While no direct link between prenylation and Abi domain proteins has been shown in prokaryotes, there are several links with the WalKR system, either by physical interaction and negative control of activity, for SpdC, or at the transcriptional level for the IspA geranyltransferase.
This study identifies a membrane-bound protein with an Abi domain, SpdC, at the core of an interaction network that coordinates bacterial division with cell envelope metabolism and host interactions. Further studies are required in order to decipher the molecular mechanism and consequences of these interactions.
Materials and methods
Bacterial strains and growth media
Escherichia coli K12 strain DH5α (Invitrogen, Thermo Fisher Scientific, Waltham, MA) was used for cloning experiments. Staphylococcus aureus strain HG001 [57] was used for genetic and functional studies. Plasmids were first passaged through the restriction deficient S. aureus strain RN4220 before introduction into the HG001 strain. E. coli strains were grown in LB medium with ampicillin (100 μg/ml) added when required. S. aureus strains and plasmids used in this study are listed in Table 2. S. aureus strains were grown in Trypticase Soy Broth (TSB; Difco; Becton, Dickinson and Co., Franklin Lakes, NJ) supplemented with chloramphenicol (10 μg/ml) or erythromycin (1 μg/ml) as required. E. coli and S. aureus strains were transformed by electroporation using standard protocols [58] and transformants were selected on LB or Trypticase Soy Agar (TSA; Difco) plates, respectively, with the appropriate antibiotics. Expression from the Pcad promoter was induced by adding cadmium chloride (CdCl2) at a final concentration of 0.25 μM. Expression from the Pspac promoter was induced by addition of isopropyl β-D-1-thiogalactopyranoside (IPTG).
Table 2. Strains and plasmids.
| Strain or plasmid | Description | Source or reference |
|---|---|---|
| E. coli strains | ||
| DH5α | F- Φ80lacZΔM15 Δ(lacZYA-argF) U169 recA1 endA1 hsdR17 (rK–, mK+) phoA supE44 λ– thi-1 gyrA96 relA1 | Invitrogen Life Technology |
| DHT1 | F-glnV44(AS) recA1 endA1 gyrA96 (Nalr) thi-1 hsdR17 spoT1 rfbD1 cya-854 ilv-691::Tn10 | [72] |
| S. aureus strains | ||
| RN4220 | Restriction-deficient transformation recipient strain | [75] |
| HG001 | NCTC 8325 rsbU+ | [57] |
| ST1000 | RN4220 Pspac-walRKHI | [9] |
| ST1017 | HG001 Pspac-walRKHI | This study |
| ST1317 | HG001 ΔspdC | This study |
| ST1375 | HG001 ΔspdC/pMK4Pprot-spdC | This study |
| ST1386 | HG001/pSD3-41 (spdC’-lacZ) | This study |
| ST1342 | HG001/pOLSA-spdC | This study |
| Plasmids | ||
| pMAD | Allelic exchange vector | [59] |
| pMADspdC | pMAD derivative for spdC deletion | This study |
| pMK4Pprot | pMK4 derivative with the constitutive Pprot promoter for complementation | [60] |
| pMK4Pprot-spdC | pMK4Pprot derivative with the spdC gene | This study |
| pSA14 | Vector for constructing transcriptional lacZ fusions | [62] |
| pSD3-41 | pSA14 derivative carrying the spdC promoter region transcriptionally fused to lacZ | This study |
| pOLSA | Vector for expression of GFP protein fusions | [30] |
| pOLSA-spdC | pOLSA derivative containing the spdC gene translationally fused with GFP | This study |
| pKT25 | BACTH vector designed to express a polypeptide fused at its N-terminal end with the adenylate cyclase T25 fragment; p15 ori | [35] |
| pUT18c | BACTH vector designed to express a polypeptide fused at its N-terminal end with the adenylate cyclase T18 fragment; ColE1 ori |
DNA manipulations
Oligonucleotides used in this study were synthesized by Eurofins Genomics (Ebersberg, Germany) and their sequences are listed in Table 3. S. aureus chromosomal DNA was isolated using the MasterPure Gram-positive DNA purification Kit (Epicentre Biotechnologies, Madison, WI). Plasmid DNA was isolated using a QIAprep Spin Miniprep kit (Qiagen, Hilden, Germany) and PCR fragments were purified using the Qiaquick PCR purification kit (Qiagen). T4 DNA ligase and restriction enzymes, PCR reagents and Q5 high-fidelity DNA polymerase (New England Biolabs, Ipswich, MA) were used according to the manufacturer's recommendations. Nucleotide sequencing of plasmid constructs was carried out by Beckman Coulter Genomics (Danvers, MA).
Table 3. Oligonucleotides used in this study.
| Name | Sequence | Description |
|---|---|---|
| Construction of pMAD-spdC | ||
| OP375 | TGCAGGATCCGATTGTTGTTCAGCACGTGT | Upstream fragment (BamHI/XmaI) |
| OP376 | AGCACCCGGGTATATGTAACCTCCATTAGGT | |
| OP377 | AGCACCCGGGTAACAAAGCGCTTGCTAGTA | Downstream fragment (XmaI/NcoI) |
| OP378 | ATTCTCCATGGGATTACATAAATATGGGAGGC | |
| Construction of pMK4Pprot-spdC | ||
| OP404 | ACTGGATCCTATTTTGCTTGTTACCTAATGGAGG | spdC coding sequence (BamHI/SalI) |
| OP405 | AGCGTCGACTTTGTTATTATTTGTTTTTATCTG | |
| Construction of pSD3-41 (pSA14-PspdC) | ||
| OSA512 | CTTCTGCAGAGCATTTCCCCCTCTTATTTATGTG |
spdC upstream region (PstI/BamHI) |
| OSA513 | GTTGGATCCTATATGTAACCTCCATTAGGTAACAAGC | |
| Construction of pOLSA-spdC | ||
| OSA417 | CTTGTCGACTAATGGAGGTTACATATAATGAAGAAC |
spdC coding sequence (SalI/XmaI) |
| OSA404 | TGCCCGGGTTTGTTTTTATCTGAAGATTGTTCTTCAG | |
| Construction of BACTH plasmids | ||
| OSA385 | GTTGGATCCCATGAAGAACAATAAAATTTCTGGTTTTCAATGGG | spdC (saouhsc-02611) (BamHI/KpnI) |
| OSA386 | AGCGGTACCTTTGTTATTATTTGTTTTTATCTGAAG | |
| OSA379 | AGGGGATCCCATGCAAAAATTCAAAGACTTTTTTTACG | spdA (saouhsc-1900) (BamHI/KpnI) |
| OSA380 | TGAGGTACCTATGAAAGGTTATCTCAAAATTATCTCC | |
| OSA383 | GGAGGATCCCGTGGAAAATGAAAAAAAGAAATACACG | spdB (saouhsc-02587) (BamHI/KpnI) |
| OSA384 | ACTGGTACCCCCGACCTCTTTATCTACGCATAAATA | |
| OSA381 | GAGGGATCCCATGACAAGATTATGGGCATCATTGC | saouhsc-02256 (BamHI/KpnI) |
| OSA382 | AGCGGTACCATCCGTGTGTGATTCGTTTTTTTTATTATGG | |
| OSA363 | TCGGATCCCATGAAGTGGCTAAAACAACTACAATCCC | walK (saouhsc-00021) (BamHI/KpnI) |
| OSA364 | AGGGTACCTATGCTCCTTATTATTCATCCC | |
| OSA488 | AGAGGATCCCATGACGGCATACAAACCTTATAGAC | yesM (saouhsc-00185) (BamHI/EcoRI) |
| OSA489 | ATAGAATTCTCATCATCACAAATAACTACC | |
| OSA490 | ATCGGATCCCATGTTATTACTTGAGCGTGTAGG | lytS (saouhsc-00230) (BamHI/EcoRI) |
| OSA491 | CATGAATTCTGATTAATGCTTTCATATTTATTCC | |
| OSA492 | TATGGATCCCATGAATAATTTGAAATGGGTAGC | graS (saouhsc-00666) (BamHI/EcoRI) |
| OSA493 | TCAGAATTCAGTAACAAAACGCATGTTTAAAATGAC | |
| OSA397 | GAGGATCCCATGGTGTTATCAATTAGAAGTCAAATCATTATTGC | saeS (saouhsc-00714) (BamHI/KpnI) |
| OSA398 | TAGGTACCAATCGGATTATGACGTAATGTCTAATTTGTG | |
| OSA494 | GGAGGATCCCATGAAATTTTTAAAAGATACTTC | desK (saouhsc-01313) (BamHI/EcoRI) |
| OSA495 | TTTGAATTCTCTGCAATAATTAAAGATGTCATGC | |
| OSA496 | ATGGGATCCCATGACAAAACGTAAATTGCGCAATAAC | arlS (saouhsc-01419) (BamHI/EcoRI) |
| OSA497 | AAAGAATTCACTTTGATTGACGTCTCAGTCATG | |
| OSA482 | GTTGGATCCCATGATGAGCCGGCTAAATAGTG | ssrB (saouhsc-01585) (BamHI/EcoRI) |
| OSA483 | GTTGAATTCTAACTATATTCAATTTTATTCTGG | |
| OSA480 | AAGGGATCCCATGAAGTTTCACCACCGCTTAATG | phoR (saouhsc-01799) (BamHI/EcoRI) |
| OSA481 | TACGAATTCTGTTACCACTTTAATTTTTATTC | |
| OSA498 | GAGGGATCCCATGGAACAAAGGACGCGACTAGC | airS (saouhsc-01981) (BamHI/EcoRI) |
| OSA499 | TTTGAATTCATGGGTTATCTCCTTAAATCAAGC | |
| OSA484 | TGAGGATCCCATGAACCACTACATTAGAACAATTGG | vraS (saouhsc-02099) (BamHI/KpnI) |
| OSA485 | ATCGGTACCAAACAATACTTTAATCGTCATACG | |
| OSA401 | TTGGATCCCATGATATTAATGTTTACAATACCAGC | agrC (saouhsc-02264) (BamHI/KpnI) |
| OSA402 | CAGGTACCTCCTTATGGCTAGTTGTTAATAATTTCAAC | |
| OSA500 | GGTGGATCCCATGTCAAACACTGAATCGCTAAAC | kdpD (saouhsc-02314) (BamHI/EcoRI) |
| OSA501 | CTTGAATTCTCAATATTTTAGATTGCATTATACG | |
| OSA502 | TGGGGATCCCATGTTTAAAACACTCTATGCTAG | hssS (saouhsc-02644) (BamHI/EcoRI) |
| OSA503 | AAAGAATTCTTCAGGAGGTAGAGATTAAAGTG | |
| OSA504 | GGTGGATCCCATGATTAATGAGGACAGTATACAG | nreB (saouhsc-02676) (BamHI/KpnI) |
| OSA505 | TTTGGTACCCCAATGTATGTTTCAAATTGGAATG | |
| OSA506 | GGTGGATCCCATGACCTTTCTTAAAAGTATTACTC | braS (saouhsc-02955) (BamHI/EcoRI) |
| OSA507 | ACTGAATTCATTGAAAGTTTTTATTCATCTGG | |
| OSA385 | GTTGGATCCCATGAAGAACAATAAAATTTCTGGTTTTCAATGGG |
spdC 1–252 (BamHI/KpnI) |
| OSA520 | AATGGTACCGATAATTAAGCTTAAACCAATGTATCC | |
| OSA363 | TCGGATCCCATGAAGTGGCTAAAACAACTACAATCCC |
walK 1–203 (BamHI/KpnI) |
| OSA518 | AATGGTACCCGCTATAAAGAATCCTAGGATGAC | |
| OSA397 | GAGGATCCCATGGTGTTATCAATTAGAAGTCAAATCATTATTGGC |
saeS 1–120 (BamHI/KpnI) |
| OSA517 | TTCGGTACCTTCGGATTTAATTTGATTCATTTGTTGC | |
| OSA397 | GAGGATCCCATGGTGTTATCAATTAGAAGTCAAATCATTATTGGC |
saeS 1–64 (BamHI/KpnI) |
| OSA519 | AATGGTACCTATAAGTGGATTAATAAAAATACTAC | |
| qRT-PCR experiments | ||
| OSA161 | ACGTGGATAACCTACCTATAAGACTGGGAT | 16s rRNA |
| OSA162 | TACCTTACCAACTAGCTAATGCAGCG | |
| OSA391 | GCAGTAGGATACATTGGTT | spdC |
| OSA392 | CAGCCTCAGTATGATTAGTT | |
| OSA203 | AACAGCACCAACGGATTAC | atlA |
| OSA204 | CATAGTCAGCATAGTTATTCATTG | |
| OSA127 | ATCAAATACAACATTAACTGTCGTTGATGC | sdrD |
| OSA128 | CATGTTTTGCAGTCGCAATTGTTTCACC | |
| OP321 | TGAGTCAGACATTAGGATAC | hlgC |
| OP322 | TTGTTGTTCTACTTCACTTAC | |
| OP291 | AAGTGCTAACCTATTGTCAGAAG | spa |
| OP292 | TCGTCTTTAAGGCTTTGGATG | |
| OSA218 | AGCGAACAGTAATAACTACCAATG | lytM |
| OSA219 | CGATGCCACCAGACATACG | |
| OSA231 | ACAGGTACTAATGGAGCAGAC | sceD |
| OSA232 | TGTGGTTGTTGAGTTTGAGC | |
| OSA138 | GTGTACTGTGCATACGATGGTAATGATGC | walR |
| OSA139 | CGTTACATAGTCATCTGCACCTAGTTCTA | |
Plasmid and mutant construction
For construction of the ΔspdC mutant strain, two 800 bp DNA fragments were generated by PCR using oligonucleotide pairs OP375/OP376 and OP377/OP378, respectively (see Table 3), corresponding to the DNA regions located immediately upstream and downstream from the spdC gene. These DNA fragments were cloned in tandem in two consecutive steps, between the BamHI and NcoI restriction sites of the pMAD vector. The resulting plasmid was introduced by electroporation into S. aureus and transformants were selected at 30°C on TSA plates containing erythromycin (1 μg/ml) and 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside (X-Gal, 100 μg/ml). Integration and excision of the plasmid were then performed as previously described [59], yielding mutant strain ST1317 (ΔspdC). A complementation plasmid pMK4Pprot-spdC was constructed by cloning the entire spdC coding sequence (amplicon OP404/OP405) in plasmid pMK4Pprot, under the control of the constitutive Pprot promoter [60]. The plasmid was introduced into the ST1317 ΔspdC mutant, generating the ST1375 complemented strain. Expression of the walKRHI in strain HG001 was placed under the control of the IPTG-iducible Pspac promoter by Φ80α phage transduction [61] using strain ST1000 (RN4220 PspacwalRKHI; [9]) as a donor and strain HG001 as the recipient, yielding strain ST1017 (HG001 PspacwalRKHI).
β-Galactosidase assays
Plasmid pSA14 [62] is a derivative of shuttle vector pMK4 [63], carrying a promoterless E. coli lacZ gene and was used to construct transcriptional lacZ reporter fusions. The spdC promoter region was amplified by PCR using oligonucleotides OSA512/OSA513 (see Table 3) and cloned between the PstI/BamHI restriction sites of the pSA14 vector, yielding plasmid pSD3-41 (Table 2).
For β-galactosidase assays in S. aureus, strain ST1386 carrying the spdC’-lacZ fusion was grown in TSB at 37°C and cells were harvested by centrifuging 2 ml culture samples (2 min; 5,400 x g). Assays were performed as previously described [21] and β-galactosidase specific activities expressed as Miller units mg−1 protein [64]. Protein concentrations were determined using the Bio-Rad protein assay (BioRad, Hercules, CA) [65].
Total RNA extraction
Strains were grown in TSB, supplemented with IPTG when specified, at 37°C with aeration until OD600nm = 1. Cells were pelleted by centrifugation (2 min, 20,800 x g) and immediately frozen at -20°C. RNA extraction was then performed as previously described [66], followed by DNaseI treatment with the TURBO DNA-free reagent (Ambion, Austin, TX) in order to eliminate residual genomic DNA.
cDNA synthesis and quantitative real time PCRs (qRT-PCRs)
cDNA synthesis was carried out as previously described [47]. Oligonucleotides were designed with the BEACON Designer 7.91 software (Premier Biosoft International, Palo Alto, CA) in order to synthesize 100–200 bp amplicons (see Table 3). Quantitative real-time PCRs (qRT-PCRs), critical threshold cycles (CT) and n-fold changes in transcript levels were performed and determined as previously described using the SsoFast EvaGreen Supermix (Bio-Rad, Hercules, CA) and normalized with respect to 16S rRNA whose levels did not vary under our experimental conditions [47]. All assays were performed using quadruplicate technical replicates, and repeated with three independent biological samples.
RNA-Seq transcriptome analysis
Three independent biological replicates were used for RNA-Seq analysis of the parental HG001 and ΔspdC strains. Strains were grown in TSB until OD600nm = 1. Total RNA was isolated as described above, and 7 μg were treated using the MicrobExpress kit (Ambion, Austin, TX) in order to remove rRNA. The rRNA depleted fraction was used for construction of strand specific single end cDNA libraries using the Truseq Stranded Total RNA sample prep kit according to the manufacturer’s instructions (Illumina, San Diego, CA). Libraries were sequenced using an Illumina Hiseq2000 sequencer (multiplexing 6 samples in one lane) according to the manufacturer’s instructions (Illumina, San Diego, CA).
Sequences were demultiplexed using the Illumina alignment and sequence analysis pipeline (GERALD, included in CASAVA version 1.7) giving FASTQ formatted reads. Reads were cleaned by removing adapter and low quality sequences using an in-house program (https://github.com/baj12/clean_ngs). Only sequences with a minimum length of 25 nucleotides were considered for further analysis. Bowtie (version 0.12.7, -m50—chunkmbs 400 -a—best -q -e50) [67] was used for alignment with the reference Staphylococcus aureus subsp. aureus genome (gi|88193823). Only uniquely aligning reads where considered for counting. HTseq-count (version 0.5.4p5, parameters: -m intersection-nonempty, -s yes, -t CDS -I locus_tag) was used for counting genes [68].
Statistical analysis was performed with R version 3.0.2 [69] and DESeq2 version 1.2.10 [70]. Data were first normalized with DESeq2 and the default parameters. Dispersion estimation and statistical testing were performed using the Generalized Linear Model with default parameters. Independent filtering was performed with default parameters to exclude transcripts with very low count values. Raw P-values were then adjusted according to the Benjamini and Hochberg procedure [71] and transcripts were considered differentially expressed when their adjusted P-value was lower than 0.05.
BACTH protein interaction assays
For testing protein interactions using the Bacterial Adenylate Cyclase Two-Hybrid System (BACTH), genes encoding the proteins of interest were cloned into plasmids pKT25 and pUT18c leading to translational fusions with the T25 or T18 domains of the Bordetella pertussis adenylate cyclase [35]. DNA fragments corresponding to the coding sequences were amplified by PCR using chromosomal DNA from strain HG001 and specific oligonucleotide pairs (see Table 3). Fragments were digested with BamHI and EcoRI or KpnI (indicated in Table 3) for cloning into plasmids pKT25 or pUT18c. The resulting plasmids were co-transformed into E. coli strain DHT1 [72] to detect protein-protein interactions and transformants were selected on kanamycin (50 μg/ml) for pKT25 derivatives and ampicillin (100 μg/ml) for pUT18c derivatives.
The resulting strains carrying combinations of pKT25 and pUT18c derivatives were tested for cyclic AMP-dependent activation of lacZ expression. For tests on plates, strains were grown in LB liquid medium supplemented with ampicillin (100 μg/ml) and kanamycin (50 μg/ml). Overnight cultures were then spotted on LB-agar plates containing IPTG (0.5 mM), ampicillin (100 μg/ml), kanamycin (50 μg/ml), and X-Gal (100 μg/ml). Plates were incubated for 24 H at 30°C and examined for appearance of the characteristic blue color indicative of β-galactosidase activity through X-Gal hydrolysis. Quantitative β-galactosidase assays were performed on exponentially growing E. coli liquid cultures. Cells were grown in LB with IPTG (0.5 mM), ampicillin (100 μg/ml) and kanamycin (50 μg/ml) at 30°C under aeration until OD600nm = 1 and assays performed on SDS/chloroform permeabilized cells as previously described [64]. Enzymatic activities are represented relative to negative and positive controls, respectively a strain carrying the empty pKT25 and pUT18c vectors (activity = 0, arbitrary unit), and a strain with the pKT25-zip and pUT18c-zip plasmids [35] (activity = 1000, arbitrary unit).
Subcellular localization of SpdC
The pOLSA plasmid was used to produce a fluorescent SpdC-GFP fusion protein [30]. The translational fusion was constructed by PCR amplification using HG001 chromosomal DNA and oligonucleotide pair OSA417/OSA404 (Table 3). The amplicon was cloned into pOLSA between the SalI and XmaI restriction sites, allowing transcription from the Pcad promoter and production of the SpdC-GFP fusion protein.
Subcellular protein localization of SpdC was performed in S. aureus HG001 transformed with pOLSA-spdC. Fluorescence microscopy was carried out on cells grown in liquid cultures in TSB supplemented with CdCl2 (0.25 μM) to induce gene fusion expression. When cells reached OD600nm ≈ 1.5 (exponential growth phase), they were harvested and concentrated 20 times in PBS. Cell suspensions were mixed with Vectashield mounting media (Vector Laboratories, Burlingame, CA) and used for microscopic observations with a Nikon Eclipse E600. Images were acquired with a Nikon DXM1200F Digital Camera. ImageJ software was used for quantifying fluorescence (http://imagej.nih.gov/ij/index.html; [73]). Fluorescence ratios were calculated by measuring fluorescence at the division septa divided by the fluorescence at the lateral wall after subtracting background fluorescence. Quantification was performed for 33 cells and two independent biological replicates and plotted using GraphPad Prism (GraphPad Software, San Diego, CA; http://www.graphpad.com).
Sensitivity to compounds targeting the cell envelope
The HG001, ΔspdC and ΔspdC/pMK4Pprot-spdC strains were grown overnight at 37°C with aeration in TSB medium, with chloramphenicol (10 μg/ml) when required. Bacterial suspensions diluted from 10−2 to 10−7 were spotted (3 μl) onto TSA plates with the indicated antibiotic concentrations and incubated for 15 hours at 37°C.
Protein preparation and Western blotting
S. aureus whole cell lysates were prepared as previously described [47]. Briefly, 5 ml of a cell culture grown to stationary phase were harvested by centrifugation (10 min; 3,000 x g), resuspended in 2X Laemmli SDS sample buffer (0.2 ml) and heated at 99°C for 10 min. Supernatants containing SDS-soluble proteins were collected following centrifugation (10 min; 20,800 x g), and used for further analysis. Cell wall extracts were prepared from 50 ml of the same cultures. Cells were pelleted and resuspended in 4 ml of digestion buffer (50 mM Tris-HCl pH 8, 145 mM NaCl, 30% sucrose, 160 ng/ml DNaseI, 250 μg/ml lysostaphin) and incubated for 60 min at 37°C. Supernatants corresponding to cell wall extracts were then harvested by centrifugation (10 min; 3,000 x g). Protein extracts were separated by SDS-PAGE on a 12% polyacrylamide gel, followed by Coomassie Brilliant Blue staining to verify that the quality and quantity of loaded extracts was equivalent for the different strains. For immunoblotting experiments, protein extracts were transferred to a nitrocellulose membrane after SDS-PAGE using a semi-dry blotter (Bio-Rad, Hercules, CA) and the following buffer: 25 mM Tris, 192 mM glycine, 20% ethanol. The LytM protein was detected using a purified polyclonal rabbit antibody [74] and horseradish peroxidase-coupled anti-rabbit secondary antibodies (Zymed, ThermoFisher, Waltham, MA) and the Pico chemiluminescence Western blot kit (Pierce, ThermoFisher, Waltham, MA). Detection of Spa was carried out directly using the secondary antibodies. Purified Staphylococcus aureus Protein A was obtained from Sigma-Aldrich (St. Louis, MO).
Lysostaphin-induced lysis assays
Strains were grown in TSB, with 10 μg/ml chloramphenicol for the complemented strain, at 37°C under aeration. When the OD600nm reached 1, bacteria were harvested by centrifugation, (10 min; 3000 x g), washed in PBS, and resuspended in the same volume of PBS (control) or PBS containing 200 ng/ml lysostaphin followed by incubation at 37°C. Lysis was monitored by measuring the decline in OD600nm over time and indicated as a percentage of the initial OD (measured OD600nm/initial OD600nm).
Biofilm formation assay
Biofilm assays were performed by growing cells in PVC microtiter plates (200 μl per well) in TSB with 0.75% glucose and 3.5% NaCl. After 24 h static growth at 37°C, adherent biomass was rinsed twice with PBS, air dried, stained with 0.1% crystal violet for 15 min, resuspended in ethanol-acetone (80:20) and quantified by measuring OD595nm, normalized to the OD600nm of each culture (growth rates for the different strains were the same).
Murine infection experiments
Seven-week-old female RjOrl:SWISS mice (Centre d’Elevage Roger Janvier, Le Genest-St.-Isle, France) were inoculated intravenously with the S. aureus HG001 parental strain and the otherwise isogenic ΔspdC mutant. Groups of seven mice were infected with 5.107 cfu per mouse in 0.2 ml. Survival was monitored daily over 9 days post-infection and three independent experiments were carried out. Virulence of the complemented strain could not be carried out since we have shown that in vivo, in the absence of selection pressure, the complementation plasmid was lost over the assay period. Indeed, after nine days post-infection with the ST1375 complemented strain, animals were sacrificed and the kidneys removed and homogenized for determination of bacterial CFU (total and chloramphenicol resistant) per kidney, revealing that 97% of the bacteria had lost the pMK4Pprot-spdC complementation plasmid.
Ethics statement
Animal experiments were conducted at the Institut Pasteur in compliance with French legislation (Decree N° 2001–464 05/29/01) and European Union guidelines on handling of laboratory animals:
(http://ec.europa.eu/environment/chemicals/lab_animals/index_en.htm).
Animals were sacrificed by increasing carbon dioxide concentrations. Protocols were approved by the Institut Pasteur ethics committee (Authorization N° 2013–0032).
Accession numbers
The complete RNA-Seq dataset was deposited in the EMBL European Nucleotide Archive (accession number PRJEB11849) and is accessible at the following URL:
Supporting information
Bacterial cultures were grown overnight, inoculated in TSB at a calculated OD600nm of 0.05 and incubated at 37°C with aeration. Optical densities were followed over a 6-hour period. Results are shown as the mean and standard deviation of three independent growth curves. Doubling times (http://www.doubling-time.com/compute.php) were calculated during the exponential growth phase (between 90 min and 214 min) and gave identical values of 32 min for each strain. Strains: HG001 ( ); ΔspdC (○).
); ΔspdC (○).
(TIFF)
Staphylococcus aureus strain ST1386 cells carrying plasmid pSD3-41 (spdC’-lacZ fusion) were grown in TSB at 37°C and harvested at the indicated OD600 nm values by centrifuging 2 ml culture samples (2 min; 5,400 x g). β-galactosidase assays were then performed as previously described [21] and β-galactosidase specific activities expressed as Miller units mg−1 protein [64]. Protein concentrations were determined using the Bio-Rad protein assay (BioRad, Hercules, CA) [65]. Results are shown as the mean and standard deviation of three independent experiments. The background value for the strain carrying the pSA14 plasmid with the promoterless lacZ gene was less than two Miller units mg−1 protein under the same conditions.
(TIFF)
Bacteria were grown in TSB at 37°C with aeration until OD600 nm ≈ 1, pelleted (10 min; 5,400 x g), resuspended in phosphate buffered saline (PBS) with Triton X-100 (0.1%), and incubated at 37°C with aeration. Lysis was determined as the decrease in OD600 nm over time and indicated as a percentage of the initial OD (measured OD600 nm / initial OD600 nm). Results are shown as the mean and standard deviation of three independent experiments. Strains: HG001 (■); ΔspdC (▲).
(TIFF)
Biofilm assays were performed by growing cells in PVC microtiter plates (200 μl per well) in TSB with 0.75% glucose and 3.5% NaCl. After 24 h static growth at 37°C, biofilm cultures were treated with DNAseI (5 μg/ml), proteinase K (1 μg/ml), or sodium metaperiodate (25 mM) for two hours before washing and staining of biofilms. The remaining adherent biomass was rinsed twice with PBS, air dried, stained with 0.1% crystal violet for 15 min, resuspended in ethanol-acetone (80:20) and quantified by measuring OD595nm, normalized to the OD600nm of each culture. The values represent the mean of two biological replicates and seven technical replicates and standard deviations are indicated.
(TIFF)
Acknowledgments
We are grateful to Jean-Yves Coppée and Arnaud Firon for helpful discussion. We thank Dr. Gouzel Karimova for the kind gift of E. coli strain DHT1 and plasmids pKT25 and pUT18c and Dr. Magdalena M. van der Kooi-Pol for the Staphylococcus aureus cell fractionation protocol.
Data Availability
The complete RNA-Seq dataset was deposited in the EMBL European Nucleotide Archive (accession number PRJEB11849) and is accessible at the following URL: http://www.ebi.ac.uk/ena/data/view/PRJEB11849.
Funding Statement
This work was supported by funds from the Institut Pasteur (www.pasteur.fr) and CNRS (www.cnrs.fr). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Lowy FD. Staphylococcus aureus infections. N Engl J Med. 1998;339: 520–32. doi: 10.1056/NEJM199808203390806 [DOI] [PubMed] [Google Scholar]
- 2.Wertheim HF, Melles DC, Vos MC, van Leeuwen W, van Belkum A, Verbrugh HA, et al. The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect Dis. 2005;5: 751–62. doi: 10.1016/S1473-3099(05)70295-4 [DOI] [PubMed] [Google Scholar]
- 3.Kuroda M, Ohta T, Uchiyama I, Baba T, Yuzawa H, Kobayashi I, et al. Whole genome sequencing of meticillin-resistant Staphylococcus aureus. Lancet. 2001;357: 1225–40. [DOI] [PubMed] [Google Scholar]
- 4.Giraudo AT, Calzolari A, Cataldi AA, Bogni C, Nagel R. The sae locus of Staphylococcus aureus encodes a two-component regulatory system. FEMS Microbiol Lett. 1999;177: 15–22. [DOI] [PubMed] [Google Scholar]
- 5.Novick RP. Autoinduction and signal transduction in the regulation of staphylococcal virulence. Mol Microbiol. 2003;48: 1429–49. [DOI] [PubMed] [Google Scholar]
- 6.Delauné A, Dubrac S, Blanchet C, Poupel O, Mader U, Hiron A, et al. The WalKR system controls major staphylococcal virulence genes and is involved in triggering the host inflammatory response. Infect Immun. 2012;80: 3438–53. doi: 10.1128/IAI.00195-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Liu Q, Yeo WS, Bae T. The SaeRS two-component system of Staphylococcus aureus. Genes. 2016;7. [Google Scholar]
- 8.Haag AF, Bagnoli F. The role of two-component signal transduction systems in Staphylococcus aureus virulence regulation. Curr Top Microbiol Immunol. 2016: 1–54. [DOI] [PubMed] [Google Scholar]
- 9.Dubrac S, Msadek T. Identification of genes controlled by the essential YycG/YycF two-component system of Staphylococcus aureus. J Bacteriol. 2004;186: 1175–81. doi: 10.1128/JB.186.4.1175-1181.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Toledo-Arana A, Merino N, Vergara-Irigaray M, Debarbouille M, Penades JR, Lasa I. Staphylococcus aureus develops an alternative, ica-independent biofilm in the absence of the arlRS two-component system. J Bacteriol. 2005;187: 5318–29. doi: 10.1128/JB.187.15.5318-5329.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Dubrac S, Bisicchia P, Devine KM, Msadek T. A matter of life and death: cell wall homeostasis and the WalKR (YycGF) essential signal transduction pathway. Mol Microbiol. 2008;70: 1307–22. doi: 10.1111/j.1365-2958.2008.06483.x [DOI] [PubMed] [Google Scholar]
- 12.Mitrophanov AY, Groisman EA. Signal integration in bacterial two-component regulatory systems. Genes Dev. 2008;22: 2601–11. doi: 10.1101/gad.1700308 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Buelow DR, Raivio TL. Three (and more) component regulatory systems—auxiliary regulators of bacterial histidine kinases. Mol Microbiol. 2010;75: 547–66. doi: 10.1111/j.1365-2958.2009.06982.x [DOI] [PubMed] [Google Scholar]
- 14.Hess JF, Oosawa K, Kaplan N, Simon MI. Phosphorylation of three proteins in the signaling pathway of bacterial chemotaxis. Cell. 1988;53: 79–87. [DOI] [PubMed] [Google Scholar]
- 15.Ohlsen KL, Grimsley JK, Hoch JA. Deactivation of the sporulation transcription factor Spo0A by the Spo0E protein phosphatase. Proceedings of the National Academy of Sciences of the United States of America. 1994;91: 1756–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Perego M, Glaser P, Hoch JA. Aspartyl-phosphate phosphatases deactivate the response regulator components of the sporulation signal transduction system in Bacillus subtilis. Mol Microbiol. 1996;19: 1151–7. [DOI] [PubMed] [Google Scholar]
- 17.Wang L, Grau R, Perego M, Hoch JA. A novel histidine kinase inhibitor regulating development in Bacillus subtilis. Genes Dev. 1997;11: 2569–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cunningham KA, Burkholder WF. The histidine kinase inhibitor Sda binds near the site of autophosphorylation and may sterically hinder autophosphorylation and phosphotransfer to Spo0F. Mol Microbiol. 2009;71: 659–77. doi: 10.1111/j.1365-2958.2008.06554.x [DOI] [PubMed] [Google Scholar]
- 19.Garnerone AM, Cabanes D, Foussard M, Boistard P, Batut J. Inhibition of the FixL sensor kinase by the FixT protein in Sinorhizobium meliloti. J Biol Chem. 1999;274: 32500–6. [DOI] [PubMed] [Google Scholar]
- 20.Mascher T. Bacterial (intramembrane-sensing) histidine kinases: signal transfer rather than stimulus perception. Trends Microbiol. 2014;22: 559–65. doi: 10.1016/j.tim.2014.05.006 [DOI] [PubMed] [Google Scholar]
- 21.Hiron A, Falord M, Valle J, Debarbouille M, Msadek T. Bacitracin and nisin resistance in Staphylococcus aureus: a novel pathway involving the BraS/BraR two-component system (SA2417/SA2418) and both the BraD/BraE and VraD/VraE ABC transporters. Mol Microbiol. 2011;81: 602–22. doi: 10.1111/j.1365-2958.2011.07735.x [DOI] [PubMed] [Google Scholar]
- 22.Falord M, Karimova G, Hiron A, Msadek T. GraXSR proteins interact with the VraFG ABC transporter to form a five-component system required for cationic antimicrobial peptide sensing and resistance in Staphylococcus aureus. Antimicrob Agents Chemother. 2012;56: 1047–58. doi: 10.1128/AAC.05054-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Alves R, Savageau MA. Comparative analysis of prototype two-component systems with either bifunctional or monofunctional sensors: differences in molecular structure and physiological function. Mol Microbiol. 2003;48: 25–51. [DOI] [PubMed] [Google Scholar]
- 24.Keener J, Kustu S. Protein kinase and phosphoprotein phosphatase activities of nitrogen regulatory proteins NTRB and NTRC of enteric bacteria: roles of the conserved amino-terminal domain of NTRC. Proc Natl Acad Sci USA. 1988;85: 4976–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Firon A, Tazi A, Da Cunha V, Brinster S, Sauvage E, Dramsi S, et al. The Abi-domain protein Abx1 interacts with the CovS histidine kinase to control virulence gene expression in group B Streptococcus. PLoS Pathog. 2013;9: e1003179 doi: 10.1371/journal.ppat.1003179 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Jeong DW, Cho H, Jones MB, Shatzkes K, Sun F, Ji Q, et al. The auxiliary protein complex SaePQ activates the phosphatase activity of sensor kinase SaeS in the SaeRS two-component system of Staphylococcus aureus. Mol Microbiol. 2012;86: 331–48. doi: 10.1111/j.1365-2958.2012.08198.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Szurmant H, Mohan MA, Imus PM, Hoch JA. YycH and YycI interact to regulate the essential YycFG two-component system in Bacillus subtilis. J Bacteriol. 2007;189: 3280–9. doi: 10.1128/JB.01936-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Szurmant H, Bu L, Brooks CL 3rd, Hoch JA. An essential sensor histidine kinase controlled by transmembrane helix interactions with its auxiliary proteins. Proc Natl Acad Sci USA. 2008;105: 5891–6. doi: 10.1073/pnas.0800247105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Fukushima T, Furihata I, Emmins R, Daniel RA, Hoch JA, Szurmant H. A role for the essential YycG sensor histidine kinase in sensing cell division. Mol Microbiol. 2011;79: 503–22. doi: 10.1111/j.1365-2958.2010.07464.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Poupel O, Moyat M, Groizeleau J, Antunes LCS, Gribaldo S, Msadek T, et al. Transcriptional analysis and subcellular protein localization reveal specific features of the essential WalKR system in Staphylococcus aureus. PLoS One. 2016;11: e0151449 doi: 10.1371/journal.pone.0151449 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Frankel MB, Wojcik BM, DeDent AC, Missiakas DM, Schneewind O. ABI domain-containing proteins contribute to surface protein display and cell division in Staphylococcus aureus. Mol Microbiol. 2010;78: 238–52. doi: 10.1111/j.1365-2958.2010.07334.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kneidinger B, O'Riordan K, Li J, Brisson JR, Lee JC, Lam JS. Three highly conserved proteins catalyze the conversion of UDP-N-acetyl-D-glucosamine to precursors for the biosynthesis of O antigen in Pseudomonas aeruginosa O11 and capsule in Staphylococcus aureus type 5. Implications for the UDP-N-acetyl-L-fucosamine biosynthetic pathway. J Biol Chem. 2003;278: 3615–27. doi: 10.1074/jbc.M203867200 [DOI] [PubMed] [Google Scholar]
- 33.Gillaspy AF, Worrell V, Orvis J, Roe BA, Dyer DW, Iandolo JJ. The Staphylococcus aureus NCTC 8325 Genome In: Fischetti VA, Novick RP, Ferretti JJ, Portnoy DA, Rood JI, editors. Gram-Positive Pathogens. 2nd ed. Washington, DC: ASM Press; 2006. p. 381–412. [Google Scholar]
- 34.Lee CY, Lee JC. Staphylococcal capsule In: Fischetti VA, Novick RP, Ferretti JJ, Portnoy DA, Rood JI, editors. Gram-Positive Pathogens. 2nd ed. Washington, DC: ASM Press; 2006. p. 456–63. [Google Scholar]
- 35.Karimova G, Pidoux J, Ullmann A, Ladant D. A bacterial two-hybrid system based on a reconstituted signal transduction pathway. Proc Natl Acad Sci USA. 1998;95: 5752–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Raychaudhuri D, Chatterjee AN. Use of resistant mutants to study the interaction of Triton X-100 with Staphylococcus aureus. J Bacteriol. 1985;164: 1337–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Schindler CA, Schuhardt VT. Lysostaphin: a new bacteriolytic agent for the Staphylococcus. Proc Natl Acad Sci USA. 1964;51: 414–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Voyich JM, Vuong C, DeWald M, Nygaard TK, Kocianova S, Griffith S, et al. The SaeR/S gene regulatory system is essential for innate immune evasion by Staphylococcus aureus. J Infect Dis. 2009;199: 1698–706. doi: 10.1086/598967 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mainiero M, Görke C, Geiger T, Gonser C, Herbert S, Wolz C. Differential target gene activation by the Staphylococcus aureus two-component system saeRS. J Bacteriol. 2010;192: 613–23. doi: 10.1128/JB.01242-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Sun F, Li C, Jeong D, Sohn C, He C, Bae T. In the Staphylococcus aureus two-component system sae, the response regulator SaeR binds to a direct repeat sequence and DNA binding requires phosphorylation by the sensor kinase SaeS. J Bacteriol. 2010;192: 2111–27. doi: 10.1128/JB.01524-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Galperin MY, Nikolskaya AN, Koonin EV. Novel domains of the prokaryotic two-component signal transduction systems. FEMS Microbiol Lett. 2001;203: 11–21. [DOI] [PubMed] [Google Scholar]
- 42.Kofoid EC, Parkinson JS. Transmitter and receiver modules in bacterial signaling proteins. Proc Natl Acad Sci USA. 1988;85: 4981–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Parkinson JS, Kofoid EC. Communication modules in bacterial signaling proteins. Annu Rev Genet. 1992;26: 71–112. doi: 10.1146/annurev.ge.26.120192.000443 [DOI] [PubMed] [Google Scholar]
- 44.Dutta R, Qin L, Inouye M. Histidine kinases: diversity of domain organization. Mol Microbiol. 1999;34: 633–40. [DOI] [PubMed] [Google Scholar]
- 45.Mascher T. Intramembrane-sensing histidine kinases: a new family of cell envelope stress sensors in Firmicutes bacteria. FEMS Microbiol Lett. 2006;264: 133–44. doi: 10.1111/j.1574-6968.2006.00444.x [DOI] [PubMed] [Google Scholar]
- 46.Grundling A, Missiakas DM, Schneewind O. Staphylococcus aureus mutants with increased lysostaphin resistance. J Bacteriol. 2006;188: 6286–97. doi: 10.1128/JB.00457-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Dubrac S, Boneca IG, Poupel O, Msadek T. New insights into the WalK/WalR (YycG/YycF) essential signal transduction pathway reveal a major role in controlling cell wall metabolism and biofilm formation in Staphylococcus aureus. J Bacteriol. 2007;189: 8257–69. doi: 10.1128/JB.00645-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Fukushima T, Szurmant H, Kim EJ, Perego M, Hoch JA. A sensor histidine kinase co-ordinates cell wall architecture with cell division in Bacillus subtilis. Mol Microbiol. 2008;69: 621–32. doi: 10.1111/j.1365-2958.2008.06308.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Pinho MG, Errington J. Dispersed mode of Staphylococcus aureus cell wall synthesis in the absence of the division machinery. Mol Microbiol. 2003;50: 871–81. [DOI] [PubMed] [Google Scholar]
- 50.Wayne KJ, Li S, Kazmierczak KM, Tsui HC, Winkler ME. Involvement of WalK (VicK) phosphatase activity in setting WalR (VicR) response regulator phosphorylation level and limiting cross-talk in Streptococcus pneumoniae D39 cells. Mol Microbiol. 2012;86: 645–60. doi: 10.1111/mmi.12006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Gutu AD, Wayne KJ, Sham LT, Winkler ME. Kinetic characterization of the WalRKSpn (VicRK) two-component system of Streptococcus pneumoniae: dependence of WalKSpn (VicK) phosphatase activity on its PAS domain. J Bacteriol. 2010;192: 2346–58. doi: 10.1128/JB.01690-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Santa Maria JP Jr., Sadaka A, Moussa SH, Brown S, Zhang YJ, Rubin EJ, et al. Compound-gene interaction mapping reveals distinct roles for Staphylococcus aureus teichoic acids. Proc Natl Acad Sci USA. 2014;111: 12510–5. doi: 10.1073/pnas.1404099111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Schlag M, Biswas R, Krismer B, Kohler T, Zoll S, Yu W, et al. Role of staphylococcal wall teichoic acid in targeting the major autolysin Atl. Mol Microbiol. 2010;75: 864–73. doi: 10.1111/j.1365-2958.2009.07007.x [DOI] [PubMed] [Google Scholar]
- 54.Pei J, Mitchell DA, Dixon JE, Grishin NV. Expansion of type II CAAX proteases reveals evolutionary origin of gamma-secretase subunit APH-1. J Mol Biol. 2011;410: 18–26. doi: 10.1016/j.jmb.2011.04.066 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Wang M, Casey PJ. Protein prenylation: unique fats make their mark on biology. Nat Rev Mol Cell Biol. 2016;17: 110–22. doi: 10.1038/nrm.2015.11 [DOI] [PubMed] [Google Scholar]
- 56.Krute CN, Carroll RK, Rivera FE, Weiss A, Young RM, Shilling A, et al. The disruption of prenylation leads to pleiotropic rearrangements in cellular behavior in Staphylococcus aureus. Mol Microbiol. 2015;95: 819–32. doi: 10.1111/mmi.12900 [DOI] [PubMed] [Google Scholar]
- 57.Herbert S, Ziebandt AK, Ohlsen K, Schafer T, Hecker M, Albrecht D, et al. Repair of global regulators in Staphylococcus aureus 8325 and comparative analysis with other clinical isolates. Infect Immun. 2010;78: 2877–89. doi: 10.1128/IAI.00088-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual, second edition Cold Spring Harbor, N. Y: Cold Spring Harbor Laboratory; 1989. [Google Scholar]
- 59.Arnaud M, Chastanet A, Debarbouille M. New vector for efficient allelic replacement in naturally nontransformable, low-GC-content, gram-positive bacteria. Appl Environ Microbiol. 2004;70: 6887–91. doi: 10.1128/AEM.70.11.6887-6891.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Archambaud C, Gouin E, Pizarro-Cerda J, Cossart P, Dussurget O. Translation elongation factor EF-Tu is a target for Stp, a serine-threonine phosphatase involved in virulence of Listeria monocytogenes. Mol Microbiol. 2005;56: 383–96. doi: 10.1111/j.1365-2958.2005.04551.x [DOI] [PubMed] [Google Scholar]
- 61.Krausz KL, Bose JL. Bacteriophage transduction in Staphylococcus aureus: broth-based method In: Bose JL, editor. The Genetic Manipulation of Staphylococci: Methods and Protocols. New York, NY: Springer New York; 2016. p. 63–8. [DOI] [PubMed] [Google Scholar]
- 62.Falord M, Mader U, Hiron A, Debarbouille M, Msadek T. Investigation of the Staphylococcus aureus GraSR regulon reveals novel links to virulence, stress response and cell wall signal transduction pathways. PLoS One. 2011;6: e21323 doi: 10.1371/journal.pone.0021323 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Sullivan MA, Yasbin RE, Young FE. New shuttle vectors for Bacillus subtilis and Escherichia coli which allow rapid detection of inserted fragments. Gene. 1984;29: 21–6. [DOI] [PubMed] [Google Scholar]
- 64.Miller JH. Assay of β-Galactosidase. Experiments in molecular genetics. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory; 1972. p. 352–5. [Google Scholar]
- 65.Bradford M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72: 248–54. [DOI] [PubMed] [Google Scholar]
- 66.Even S, Burguiere P, Auger S, Soutourina O, Danchin A, Martin-Verstraete I. Global control of cysteine metabolism by CymR in Bacillus subtilis. J Bacteriol. 2006;188: 2184–97. doi: 10.1128/JB.188.6.2184-2197.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Langmead B, Trapnell C, Pop M, Salzberg SL. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009;10: R25 doi: 10.1186/gb-2009-10-3-r25 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Anders S, Pyl PT, Huber W. HTSeq—a Python framework to work with high-throughput sequencing data. Bioinformatics. 2015;31: 166–9. doi: 10.1093/bioinformatics/btu638 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria: http://wwwR-projectorg/. 2014. [Google Scholar]
- 70.Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014;15: 550 doi: 10.1186/s13059-014-0550-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Benjamini Y, Hiochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Statist Soc B. 1995;57: 289–300. [Google Scholar]
- 72.Dautin N, Karimova G, Ullmann A, Ladant D. Sensitive genetic screen for protease activity based on a cyclic AMP signaling cascade in Escherichia coli. J Bacteriol. 2000;182: 7060–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Schneider CA, Rasband WS, Eliceiri KW. NIH Image to ImageJ: 25 years of image analysis. Nat Meth. 2012;9: 671–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Delauné A, Poupel O, Mallet A, Coic YM, Msadek T, Dubrac S. Peptidoglycan crosslinking relaxation plays an important role in Staphylococcus aureus WalKR-dependent cell viability. PLoS One. 2011;6: e17054 doi: 10.1371/journal.pone.0017054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Kreiswirth BN, Lofdahl S, Betley MJ, O'Reilly M, Schlievert PM, Bergdoll MS, et al. The toxic shock syndrome exotoxin structural gene is not detectably transmitted by a prophage. Nature. 1983;305: 709–12. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Bacterial cultures were grown overnight, inoculated in TSB at a calculated OD600nm of 0.05 and incubated at 37°C with aeration. Optical densities were followed over a 6-hour period. Results are shown as the mean and standard deviation of three independent growth curves. Doubling times (http://www.doubling-time.com/compute.php) were calculated during the exponential growth phase (between 90 min and 214 min) and gave identical values of 32 min for each strain. Strains: HG001 (); ΔspdC (○).
(TIFF)
Staphylococcus aureus strain ST1386 cells carrying plasmid pSD3-41 (spdC’-lacZ fusion) were grown in TSB at 37°C and harvested at the indicated OD600 nm values by centrifuging 2 ml culture samples (2 min; 5,400 x g). β-galactosidase assays were then performed as previously described [21] and β-galactosidase specific activities expressed as Miller units mg−1 protein [64]. Protein concentrations were determined using the Bio-Rad protein assay (BioRad, Hercules, CA) [65]. Results are shown as the mean and standard deviation of three independent experiments. The background value for the strain carrying the pSA14 plasmid with the promoterless lacZ gene was less than two Miller units mg−1 protein under the same conditions.
(TIFF)
Bacteria were grown in TSB at 37°C with aeration until OD600 nm ≈ 1, pelleted (10 min; 5,400 x g), resuspended in phosphate buffered saline (PBS) with Triton X-100 (0.1%), and incubated at 37°C with aeration. Lysis was determined as the decrease in OD600 nm over time and indicated as a percentage of the initial OD (measured OD600 nm / initial OD600 nm). Results are shown as the mean and standard deviation of three independent experiments. Strains: HG001 (■); ΔspdC (▲).
(TIFF)
Biofilm assays were performed by growing cells in PVC microtiter plates (200 μl per well) in TSB with 0.75% glucose and 3.5% NaCl. After 24 h static growth at 37°C, biofilm cultures were treated with DNAseI (5 μg/ml), proteinase K (1 μg/ml), or sodium metaperiodate (25 mM) for two hours before washing and staining of biofilms. The remaining adherent biomass was rinsed twice with PBS, air dried, stained with 0.1% crystal violet for 15 min, resuspended in ethanol-acetone (80:20) and quantified by measuring OD595nm, normalized to the OD600nm of each culture. The values represent the mean of two biological replicates and seven technical replicates and standard deviations are indicated.
(TIFF)
Data Availability Statement
The complete RNA-Seq dataset was deposited in the EMBL European Nucleotide Archive (accession number PRJEB11849) and is accessible at the following URL: http://www.ebi.ac.uk/ena/data/view/PRJEB11849.