

Abstract
Background
The envelope protein of lentiviruses are type I transmembrane proteins, and their transmembrane domain contains conserved potentially charged residues. This highly unusual feature would be expected to cause endoplasmic reticulum (ER) localization. The aim of this study was to determine by which means the HIV-1 Env protein is transported to the cell surface although its transmembrane domain contains a conserved arginine residue.
Results
We expressed various chimeric proteins and analyzed the influence of their transmembrane domain on their intracellular localization. The transmembrane domain of the HIV-1 Env protein does not cause ER retention. This is not due to the presence of conserved glycine residues, or to the position of the arginine residue, but to the length of the transmembrane domain. A shortened version of the Env transmembrane domain causes arginine-dependent ER targeting. Remarkably, the transmembrane domain of the HIV-1 Env protein, although it does not confer ER retention, interacts efficiently with negatively charged residues in the membrane.
Conclusion
These results suggest that the intrinsic properties of the HIV-1 Env transmembrane domain allow the protein to escape ER-retention mechanisms, while maintaining its ability to interact with cellular proteins and to influence cellular physiology.
Keywords: Secretory pathway, Transmembrane domain, Envelope protein, gp160, HIV-1, Endoplasmic reticulum
Background
Transmembrane proteins present at the surface of eukaryotic cells are initially inserted in the membrane of the endoplasmic reticulum (ER), from where they are transported to the Golgi apparatus and ultimately to the cell surface. Intracellular transport along the secretory pathway is coupled with sorting of proteins and lipids. As a consequence, each individual protein can eventually be found in the ER, in the Golgi apparatus, or at the cell surface. This ensures the proper localization of individual proteins in the compartment where their function is required (e.g. the ER for the signal peptidase, or the surface for the transferrin receptor). It also avoids the transport to the cell surface of proteins that are misfolded or incompletely assembled, and participates in the quality control of secreted proteins (reviewed in [1, 2]).
To ensure its proper sorting, each protein inserted in the ER exhibits specific motifs that can be recognized by the cellular transport and sorting machinery. These sorting motifs can be found in luminal domains (e.g. a KDEL ER-localization sequence), in cytosolic domains (e.g. a C-terminal KKXX ER-localization sequence) or in transmembrane domains (TMDs) (reviewed in [3]). The best-characterized ER localization motifs found in TMDs are potentially charged residues found in a number of type I transmembrane proteins. Charged residues are for example found in the TMDs of the various subunits of the T-cell receptor, and of a collection of receptors associated with DAP10, DAP12 or the FcRγ chain [4]. Typically a single charged residue in a TMD is sufficient to cause ER retention unless it is masked by the assembly and folding of protein complexes [5]. The molecular machinery that ensures the recognition and ER localization of sorting motifs in TMDs is still largely unknown. To date, the best-characterized mechanism proposes that the Rer1 protein acts as a receptor recognizing specific features of TMDs and ensuring their localization in the ER [6].
The envelope protein (Env) of HIV-1 (human immunodeficiency virus type 1) exhibits a conserved arginine residue in the TMD of its gp41subunit. The role of this potentially charged residue is poorly understood. It has notably been proposed that the arginine and/or several conserved glycines may drive interactions with other cellular proteins [7]. In particular, peptides mimicking a portion of the HIV-1 Env TMD were shown to interact with subunits of the T-cell receptor and to modulate T-cell activation [8, 9]. Similar experiments suggested an interaction of the Env TMD with TLR2 in macrophages [10]. In addition, mutations in the Env TMD may influence its intracellular transport [11] or alter its ability to induce membrane fusion [12, 13].
The assembly of HIV-1 virions requires the presence of the processed Env protein at the cell surface of infected cells [14]. Indeed, it has been amply demonstrated that the Env protein is transported to the surface of a variety of cells [15, 16], although the presence of a charged residue in its TMD would be expected to ensure its localization in the ER. The aim of the current study is to study this apparent paradox and to determine to what extent the TMD of the HIV-1 Env protein influences its intracellular transport.
Results
Potentially charged residues in the TMD of lentiviral envelope proteins
In almost every sequenced isolate of HIV-1, an arginine residue is positioned in the TMD of the Env protein (Fig. 1). In a few isolates (e.g. isolate 622,166-KT1247896.1) it is replaced, remarkably, with a lysine residue. This suggests that a positively charged residue at this position plays a critical role in the infectious cycle of HIV-1. An arginine or a lysine residue is also found in the TMD of the envelope protein of most other lentiviruses, notably HIV-2, simian (SIV) and bovine (BIV) immunodeficiency viruses, caprine arthritis/encephalitis virus (CAEV), Maedi visna ovine pneumonia virus (MVV) and equine infectious anemia virus (EIAV) (Fig. 1). In addition to a charged residue, many of these TMDs exhibit conserved glycine residues. In Feline Immunodeficiency virus (FIV), the TMD of the envelope protein contains no potentially charged residues, but exhibits six glycine residues (Fig. 1). The functional significance of these observations remains mostly unclear to date.
Fig. 1.

TMDs of lentiviral envelope proteins exhibit conserved charged residues. The sequence of the TMDs and surrounding region of several lentiviral envelope proteins are represented. Amino acid positions for each sequence are numbered in italic. The approximate position of the predicted TMDs is indicated. Potentially charged amino-acid residues are in red and glycine residues in italic. The Uniprot reference number of each protein is indicated
The TMD of the HIV-1 Env protein does not confer ER retention
The Env protein of HIV-1 is efficiently transported to the surface of infected cells despite the presence of a putatively charged arginine residue in its TMD which would be expected to cause ER retention [5]. In order to test if the Env TMD can affect its intracellular transport, we expressed fusion proteins composed of the extracellular domain of the Tac antigen fused to various TMDs (Fig. 2a and Table 1) and determined their intracellular localization. The Tac protein comprises an essentially hydrophobic 21 residues TMD (T-H0) and is readily transported to the cell surface, as revealed by successive labeling of surface and total Tac antigen in transfected cells (Fig. 2b). In the T-E14 protein, the Tac extracellular domain was fused to the Env TMD and cytoplasmic domain (Fig. 2a and Table 1). The Env extracellular juxtamembrane segment is very conserved in many lentiviral envelope proteins (Fig. 1) and it was also included in T-E14. The T-E14 protein was also abundantly present at the cell surface although a fraction of the protein was seen in intracellular compartments, most likely due to endocytic motifs in its cytosolic domain that have been shown to drive internalization from the cell surface to endosomes [17]. To quantify these observations, the surface level of Tac chimeric proteins was determined relative to the total expression level in individual cells. The T-E14 protein was indeed slightly less abundant at the cell surface than the T-H0 protein (Fig. 2c). In agreement with this interpretation the T-E15 protein, identical to T-E14 except for the deletion of its cytosolic domain, was almost exclusively localized at the cell surface (Fig. 2b and c). Similarly, the T-E26 protein where the Tac extracellular domain was fused to the TMD of Env without preserving the Env juxtamembrane region was mostly found at the cell surface (Fig. 2b and c). Together, these results indicate that despite the presence of a potentially charged arginine residue, the HIV-1 Env TMD does not target membrane proteins for efficient retention in the ER. This lack of ER retention is not due to an effect of the cytosolic or of the luminal juxtamembrane domains.
Fig. 2.
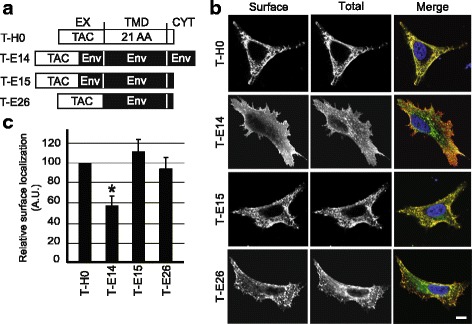
The HIV-1 Env TMD allows efficient cell surface localization. a The T-H0 chimera is composed of the extracellular domain of the Tac protein, a 21-hydrophobic residue transmembrane domain (TMD), and a short cytosolic domain (CYT). In T-E14, the Tac extracellular domain is fused to the Env extracellular juxtamembrane segment followed by the Env TMD and its complete cytoplasmic domain. The T-E15 protein exhibits a short truncated cytosolic domain. In T-E26, the Tac extracellular domain is fused directly to the Env TMD and a short cytoplasmic tail. Detailed amino acid sequences are described in Table 1. b Hela cell expressing the indicated Tac chimeric protein were labeled by immunofluorescence before (Surface) and after (Total) permeabilization, using antibodies specific for the Tac extracellular domain. All pictures were taken with the same settings with a confocal microscope (LSM700, Zeiss). Scale bar: 10 μm. c The amount of each Tac chimeric protein present at the cell surface was determined by dividing the surface fluorescence by the total fluorescence. For calibration, this ratio is set to 100 arbitrary units (A.U.) for T-H0. T-E14 is significantly less localized at the cell surface than T-H0 (n = 4; one-way analysis of variance: p < 0.01; *: post-hoc Tukey-Kramer p < 0.05)
Table 1.
Amino acids sequence of the transmembrane and cytosolic domains of the Tac chimeric proteins
| Name | Luminal | Transmembrane | Cytoplasmic |
|---|---|---|---|
| T-H0 | EY | QVAVAACVFLLIAVLLLSGLTWQ | RRQRKSRRTI |
| T-R8 | EY | QVAVAACVRLLIAVLLLSGLTWQ | RRQRKSRRTI |
| T-H0KKxx | EY | QVAVAACVFLLIAVLLLSGLTWQ | RRQRLETFKKTN |
| T-E14 | DL WLWYIK | IFIMIVGGLVGLRIVFAVLSIV | NRVR---LERILL |
| T-E15 | DL WLWYIK | IFIMIVGGLVGLRIVFAVLSIV | NRVR |
| T-E26 | DL | IFIMIVGGLVGLRIVFAVLSIV | NRVR |
| T-E28 | DL | IMIVGGLVGLRIVFAVLSIV | NRVR |
| T-E30 | DL | IVGGLVGLRIVFAVLSIV | NRVR |
| T-E34 | DL | IVGGLVGLLIVFAVLSIV | NRVR |
| T-E39 | DL | IFIMIVGGLVGLLIVFAVLSIV | NRVR |
| T-E41 | DL | IFIMIVGGLVGLLIVFAVRSIV | NRVR |
| T-E42 | DL | IFIMIVGGLVGLLIVFRVLSIV | NRVR |
| T-E43 | DL | IFIMIVGGLVGLLIRFAVLSIV | NRVR |
| T-E44 | DL | IFIMIVGGLVRLLIVFAVLSIV | NRVR |
| T-E45 | DL | IFIMIVGGRVGLLIVFAVLSIV | NRVR |
| T-E46 | DL | IFIMIVRGLVGLLIVFAVLSIV | NRVR |
| T-E59 | DL | IFIMIVLLLVLLRIVFAVLSIV | NRVR |
| T-E60 | DL | IFIMIVLLLVLLLIVFAVLSIV | NRVR |
The length of the HIV-1 Env TMD allows its exit from the ER
We next tried to determine why the arginine residue present in the Env TMD failed to confer ER localization. We tested specifically three possibilities: first, efficient transport to the cell surface may be due to an effect of conserved glycine residues in the Env TMD; second, the position of the charge in the Env TMD may be inadequate to cause ER retention; third the length of the Env TMD may cancel ER targeting by the arginine.
The glycine residues present in the Env TMD are very conserved in many HIV-1 isolates, as well as in other lentiviruses (Fig. 1). They have been proposed to play a role in the trimerization of the HIV-1 Env TMD [7, 18], and this may in principle modify the recognition of the Env TMD by the cellular sorting machinery. To test this hypothesis, we compared the surface expression of T-E26, which comprises the Env TMD with that of T-E39 (where the arginine was mutated to leucine), of T-E59 (where the three glycine residues were mutated to leucines) and to T-E60 (where both the arginine and the glycine residues were mutated to leucines) (Fig. 3a). All of these chimeric proteins were present at similar levels at the cell surface (Fig. 3b), demonstrating that the arginine residue present in the Env TMD does not confer ER retention, even in the absence of glycine residues.
Fig. 3.
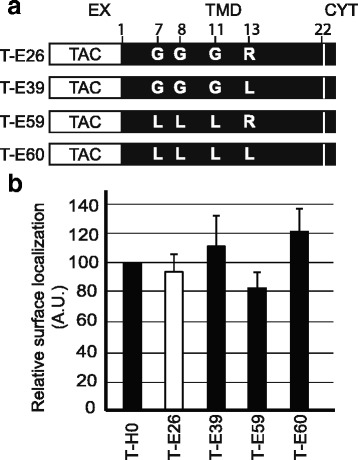
Glycine residues in the HIV-1 Env TMD do not affect its intracellular targeting. a Fusion proteins composed of the extracellular domain of the Tac protein, the transmembrane domain of Env protein (TMD) and a short cytosolic tail (CYT) are derived from the T-E26 chimera. In T-E39 the arginine present in the Env TMD is mutated to leucine. In T-E59 glycine residues in the Env TMD are mutated to leucines. Both mutations are combined in T-E60 (see also Table 1). b The fraction of each chimeric protein present at the cell surface was determined as described in the legend to Fig. 2. Statistical analysis revealed no significant differences from T-H0 (n = 3; one-way analysis of variance: p = 0.246)
Since the position of potentially charged residues in a TMD can affect their ability to cause ER retention [5], we then studied the intracellular localization of fusion proteins where the arginine was placed at positions 7, 9, 11, 13, 15, 17 or 19 in the Env TMD (Fig. 4a and Table 1). This corresponded to a variety of positions at different levels in the lipid bilayer, as well as on various sides of the TM helix. For example positions 11 and 13 correspond to approximately the two opposite sides of the TM helix. All of the corresponding fusion proteins were efficiently expressed at the cell surface (Fig. 4b), indicating that the TMD of the Env protein does not cause ER retention independent of the position where an arginine residue is placed.
Fig. 4.
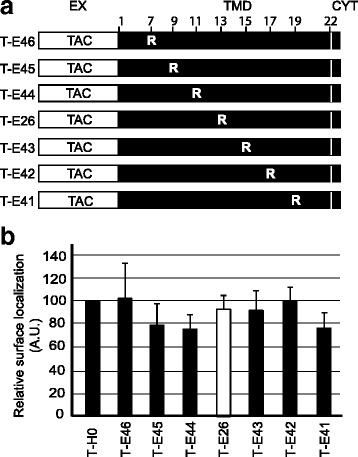
The position of the arginine residue in the HIV-1 Env TMD does not affect its intracellular targeting. a In the T-E26 chimera, an arginine is present in the Env TMD at position 14. A series of chimeric proteins where the arginine was moved to other positions were constructed (T-E41 to T-E46) (see also Table 1). Positions within the sequence are numbered from the luminal to the cytoplasmic end of the TMD. b The fraction of each chimeric protein present at the cell surface was determined as described in the legend to Fig. 2. Statistical analysis revealed no significant differences from T-H0 (n = 3; one-way analysis of variance: p = 0.77)
Previous studies have shown that the length of a TMD can also be an important element in its ability to confer specific intracellular location. For example, lengthening the TMD of a Golgi-targeted protein relocates it to the cell surface [19, 20], and similarly lengthening an ER-targeted TMD allows it to reach the cell surface [21]. There is however no algorithm capable of predicting with absolute precision the length of a TMD, and to define if a given TMD will be detected as unusually long by the intracellular sorting machinery. In order to test if the length of the Env TMD accounts for its ability to reach the cell surface, we expressed two chimeric proteins with a shortened TMD: the Env TMD of T-E26 was shortened by two residues in T-E28, and by four residues in T-E30 (Fig. 5a and Table 1). T-E28 was localized at the cell surface like T-E26 (Fig. 5b and c). T-E30 however was virtually depleted from the cell surface and localized almost exclusively in a reticular compartment akin to the ER (Fig. 5b and c). In order to verify if the intracellular retention of T-E30 was caused by the presence of a charged residue in its TMD, the arginine residue was replaced with a leucine in the fusion protein T-E34 (Fig. 5a and Table 1). T-E34 localized readily to the cell surface (Fig. 5b and c). These observations are also in agreement with a previous study showing that the unusually long TMD of the ε chain of the TCR does not cause retention in the ER despite the presence of a negatively charged residue [21].
Fig. 5.
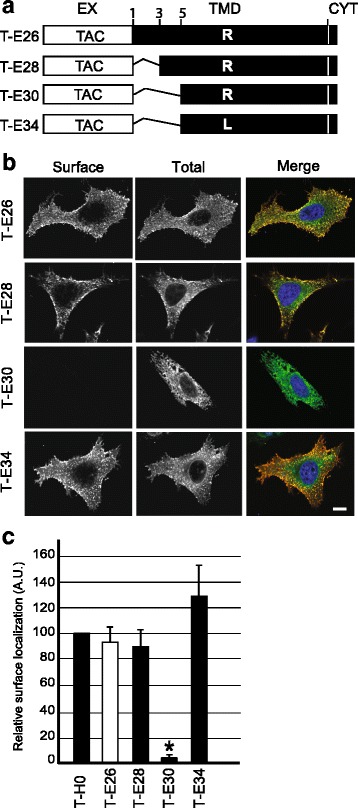
The intracellular targeting of the HIV-1 Env TMD is influenced by its length. a Compared to T-E26, two amino-acids were deleted from the Env TMD in T-E28, and four in T-E30. In addition, in T-E34 the arginine present in the TMD was mutated to a leucine residue (see also Table1). b Intracellular localization of each Tac chimeric protein was determined as described in the legend to Fig. 2. Scale bar: 10 μm. c The fraction of each chimeric protein present at the cell surface was determined as described in the legend to Fig. 2. T-E30 is significantly less localized at the cell surface than T-H0 (n = 4; one-way analysis of variance: p < 0.01; *: post-hoc Tukey-Kramer p < 0.05)
In order to ascertain that the intracellular T-E30 was localized in the ER, we coexpressed it with an ER-targeted fluorescent GFP protein. While T-H0 and T-E26 did not colocalize with the ER marker, T-H0KKxx which exhibits a cytosolic ER localization was mostly present in the ER, as well as T-E30 (Fig. 6a). This result was confirmed by calculating the Pearson correlation coefficient: the correlation coefficient between Tac fusion proteins and the ER marker was negative for T-H0 and T-E26, and positive for T-H0KKxx and T-E30 (Fig. 6b).
Fig. 6.
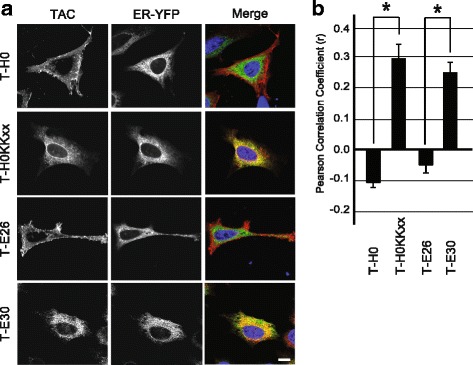
A shortened HIV-1 Env TMD acts as an ER-targeting motif. a Immunofluorescence microscopy of HeLa cells co-expressing various Tac fusion proteins (stained with an anti-Tac antibody) and a marker of the endoplasmic reticulum (ER-YFP). All pictures were taken with a confocal microscope (LSM700, Zeiss). Scale bar: 10 μm. b The colocalization of Tac proteins with the ER was quantified by measuring the Pearson’s correlation coefficient with Imaris software. T-E30 and T-H0KKxx are significantly localized in the ER (n = 4; *: Student’s t-test p < 0.01)
Together these results demonstrate that the length of the Env TMD is a key element allowing it to tolerate the presence of a charged residue while still escaping retention in the ER.
The HIV-1 Env intramembrane arginine can interact with other proteins
One mechanism by which the Env TMD could exit the ER despite the presence of an arginine would be if the length of the TMD somehow masked the arginine residue, making it invisible to the sorting machinery. To test this possibility, we coexpressed various Tac chimeric proteins and a protein (the δ subunit of the T-cell receptor) exhibiting a TMD containing an aspartic acid, and fused to β-galactosidase (Fig. 7a). We then immunoprecipitated the Tac antigen and determined the percentage of the β-galactosidase activity associated with it. As described previously [22], a TMD containing a charged residue (such as T-R8; Table 1) associated efficiently with a TMD containing a negative charge, whereas a hydrophobic TMD (T-H0) showed much less association (Fig. 7b). The Env TMD (T-E26) associated efficiently with its proposed partner, and this association was lost when the arginine was mutated to a leucine (T-E39 and T-E60), but did not decrease significantly when the glycine residues were mutated to leucines (T-E59)(Fig. 7b). These observations indicate that the arginine present in the Env TMD is readily available for interactions within the membrane.
Fig. 7.
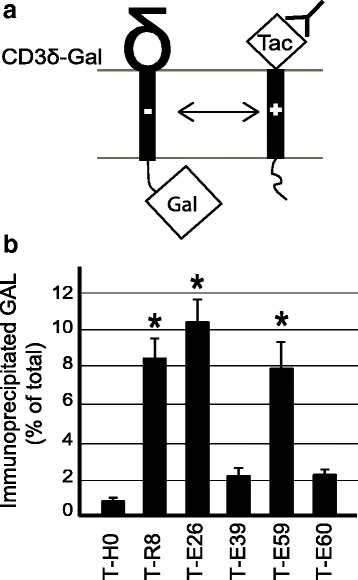
The HIV-1 Env TMD associates efficiently with a TMD containing a negative charge. a To reveal a putative interaction between TMDS, HeLa cells were co-transfected with plasmids encoding the δ chain of the CD3 receptor fused to the β-galactosidase (Gal) and various Tac fusion proteins. b Tac fusion proteins were immunoprecipitated and the amount of co-precipitated β-galactosidase activity assessed to reveal the degree of association with the δ chain of the CD3 receptor. The mean ± SEM of at least five independent experiments are indicated (one-way analysis of variance: p < 0.01; *: post-hoc Tukey-Kramer p < 0.05)
Discussion
This study was aimed at clarifying the potential role of the HIV-1 Env TMD in its intracellular transport. Our observations indicate that the charged residue present in the TMD of HIV-1 does not confer retention of the protein in the ER. According to our results, this is due to the fact that the TMD of HIV-1 is long enough to tolerate a charge and not be retained in the ER. Notwithstanding, the arginine residue is still capable of engaging into interactions within the membrane.
Our results shed new light on the molecular mechanisms that ensure recognition and ER retention of TMDs containing charges. Indeed, based on our current knowledge, two mechanisms can be envisaged to account for ER targeting by TMDs: TMDs containing a charged residue may be recognized by membrane receptors such as Rer1 that would ensure their proper intracellular sorting. Alternatively, structural and biophysical properties of a TMD, or interaction with various membrane lipids, may control its tendency to form aggregates, or its sorting to membrane subdomains enriched in specific lipids [3, 23]. According to this second model, a long TMD containing a charged residue may have biophysical properties akin to a shorter, hydrophobic TMD, while still exhibiting a charged residue capable of engaging into interactions in the membrane. In agreement with this model, our observations suggest that it is not the mere presence in the membrane of a charged residue capable of engaging into interactions that causes retention of a protein in the ER. Our observations rather suggest that the sorting machinery recognizes TMDs by a more complex mechanism, not based on simple interactions between charged residues within the membrane.
Importantly, throughout this text, we have made the choice of interpreting results based on the implicit assumption that localization of a protein in the ER reflects the presence of an ER retention or ER retrieval motif, while the absence of a retention motif would allow surface localization. An alternative interpretation would be that proteins are localized at the cell surface when they exhibit an ER-exit motif, and localized in the ER when they do not contain such a motif. Accordingly, a hydrophobic TMD, or a long TMD containing an arginine would represent an ER exit motif, while a short TMD would not direct for ER exit. For example, association with Erv14 may be favored with a long TMD but impossible with a short TMD containing an arginine [3, 24]. The two interpretations are not mutually exclusive: a long TMD may be transported efficiently at the cell surface due to both its association with Erv14 (facilitating ER exit) and its non-association with Rer1 (allowing escape from ER retrieval).
Our results bring two new elements to the discussion concerning the role of the charged residue in the HIV-1 Env TMD: first, despite the presence of an arginine residue the HIV-1 Env TMD does not per se cause retention in the ER; second, the arginine is available for interactions in the membrane. Our results are notably compatible with the notion that the Env TMD may interact with subunits of the T-cell receptor bearing TMDs with negatively charged residues, i.e. CD3 γ, δ, ε, and ζ. Indeed, in this study, the interaction partner that was proposed to the Env TMD was the CD3 δ subunit of the T-cell receptor, and our observations indicate that the HIV-1 Env TMD is capable of interacting specifically with this protein via its intramembrane arginine. Further experiments are needed to establish if, in the context of a full-length Env protein, the Env TMD is still available for efficient interactions with CD3 subunits. It remains also to be seen if in an HIV-infected T cell, the concentrations of the various proteins do allow such an interaction to effectively take place, and what its physiological consequences would be.
Interestingly, a recent study demonstrates that when an HIV-infected T cell is apposed to a non-infected T-cell, the Env protein at the surface of the infected cell interacts with the CD4 molecule on the uninfected cell [25]. Remarkably, this interaction results in the clustering of CD3 at the sites of contact, an observation compatible with the notion that the Env protein can directly interact with some elements of the T-cell receptor in an infected cell. Concomitant with this clustering, the T-cell receptor is activated, stimulating virus production, and facilitating virus spread to uninfected cells. We speculate that evolution may have favored an HIV-1 Env TMD that interacts with the T-cell receptor while still being able to escape ER retention and to reach the cell surface.
Conclusions
The TMD of the HIV-1 Env protein contains a conserved charged residue. Contrary to several TMDs exhibiting the same feature, this TMD does not confer localization of the protein in the ER. Our results indicate that this is an intrinsic property of the HIV-1 Env TMD, which is long enough to tolerate a charge and not be retained in the ER. Despite the fact that it is not recognized by the ER targeting machinery, the Env TMD is still capable of interacting with other proteins within the membrane, and may by this means influence the physiology of infected cells.
Methods
Cell culture and media
HeLa cells were grown at 37 °C and 5% CO2 in Dulbecco’s modified Eagle’s medium (Gibco) containing 10% fetal bovine serum (Gibco) and 100 μg/ml of penicillin-streptomycin (Sigma). To express various fusion proteins, cells were transfected two days before the experiment using polyethylenimine (PEI) as previously described [26].
Intracellular transport of tac chimeric proteins
We used a pCDM8-based vector containing the coding sequence of the α chain of the interleukin-2 receptor (Tac) as previously described [22]. In this study, all Tac fusion proteins were obtained by inserting the sequence coding for the TMD of interest (see Table 1) in the vector digested with BglII and XbaI.
Tac fusion proteins present at the cell surface were labeled with a mouse anti-Tac antibody (7G7) [27] for 15 min at 4 °C. Cells were then fixed for 10 min at room temperature in Phosphate Buffered Saline (PBS) containing 4% paraformaldehyde, washed with PBS containing 20 mM NH4Cl and incubated for 30 min at room temperature with an Alexa-Fluor-647-coupled anti-mouse-IgG antibody (Life Technologies, A21235) in PBS containing 0.2% Bovine Serum Albumine (PBS-BSA). Cells were then permeabilized for 10 min in PBS containing 0.2% saponin and labeled with 7G7 in PBS-BSA for 30 min. Finally, cells were incubated for 30 min with the Alexa-Fluor-488-coupled anti-mouse-IgG antibody (Life Technologies, A11029) before being mounted in Möwiol. Cells with a comparable total expression level were analyzed using a confocal microscope (LSM700, Zeiss).
When indicated, the ER was labeled by expressing a YFP-KDEL (ER-YFP, a kind gift of Nicolas Demaurex, University of Geneva, Switzerland).
Surface labeling was quantified with ImageJ software (http://rsb.info.nih.gov/ij/). The surface/total ratio was calculated for each individual cell using the surface fluorescence intensity and the total fluorescence intensity (in arbitrary units, using T-H0 as 100 a.u. for normalization). In each independent experiment, at least 15 cells were quantified.
Association assays
Experiments were performed as previously described [28]. HeLa cells were co-transfected with Tac chimeric constructs and δ chain of the CD3 receptor fused to the β-galactosidase. Tac protein from the lysate was immunoprecipitated using protein A-agarose beads coated with 7G7 antibodies. The galactosidase activity is revealed upon addition of Chlorophenol Red-β-D-galactopyranoside and quantified by measuring absorbance at 600 nm. The percentage of β-galactosidase immunoprecipitated was determined.
Acknowledgments
The students of pratical classes who participated in the elaboration of Tac chimeric proteins. The Bioimaging Core Facility at the University of Geneva Medical School provided access to confocal microscopy equipment.
Funding
This research was supported by Swiss National Science Foundation Grant 31003A-172951 (to P.C.).
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- ER
Endoplasmic reticulum.
- HIV-1
Humain immunodeficiency virus type I.
- TCR
T-cell receptor.
- TMD
Transmembrane domain.
Authors’ contributions
JP, AB, AV and PC performed experiments. PC and JP wrote the manuscript. All authors read and corrected the manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Pelham HR. The dynamic organisation of the secretory pathway. Cell Struct Funct. 1996;21(5):413–419. doi: 10.1247/csf.21.413. [DOI] [PubMed] [Google Scholar]
- 2.Teasdale RD, Jackson MR. Signal-mediated sorting of membrane proteins between the endoplasmic reticulum and the golgi apparatus. Annu Rev Cell Dev Biol. 1996;12:27–54. doi: 10.1146/annurev.cellbio.12.1.27. [DOI] [PubMed] [Google Scholar]
- 3.Cosson P, Perrin J, Bonifacino JS. Anchors aweigh: protein localization and transport mediated by transmembrane domains. Trends Cell Biol. 2013;23(10):511–517. doi: 10.1016/j.tcb.2013.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lanier LL. DAP10- and DAP12-associated receptors in innate immunity. Immunol Rev. 2009;227(1):150–160. doi: 10.1111/j.1600-065X.2008.00720.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bonifacino JS, Cosson P, Shah N, Klausner RD. Role of potentially charged transmembrane residues in targeting proteins for retention and degradation within the endoplasmic reticulum. EMBO J. 1991;10(10):2783–2793. doi: 10.1002/j.1460-2075.1991.tb07827.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sato K, Sato M, Nakano A. Rer1p, a retrieval receptor for endoplasmic reticulum membrane proteins, is dynamically localized to the Golgi apparatus by coatomer. J Cell Biol. 2001;152(5):935–944. doi: 10.1083/jcb.152.5.935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Abad C, Martinez-Gil L, Tamborero S, Mingarro I. Membrane topology of gp41 and amyloid precursor protein: interfering transmembrane interactions as potential targets for HIV and Alzheimer treatment. Biochim Biophys Acta. 2009;1788(10):2132–2141. doi: 10.1016/j.bbamem.2009.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Cohen T, Cohen SJ, Antonovsky N, Cohen IR, Shai Y. HIV-1 gp41 and TCRalpha trans-membrane domains share a motif exploited by the HIV virus to modulate T-cell proliferation. PLoS Pathog. 2010;6(9):e1001085. doi: 10.1371/journal.ppat.1001085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Rotem E, Reuven EM, Klug YA, Shai Y. The transmembrane domain of HIV-1 gp41 inhibits T-cell activation by targeting multiple T-cell receptor complex components through its GxxxG motif. Biochemistry. 2016;55(7):1049–1057. doi: 10.1021/acs.biochem.5b01307. [DOI] [PubMed] [Google Scholar]
- 10.Reuven EM, Ali M, Rotem E, Schwarzer R, Gramatica A, Futerman AH, Shai Y. The HIV-1 envelope transmembrane domain binds TLR2 through a distinct dimerization motif and inhibits TLR2-mediated responses. PLoS Pathog. 2014;10(8):e1004248. doi: 10.1371/journal.ppat.1004248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Miyauchi K, Curran AR, Long Y, Kondo N, Iwamoto A, Engelman DM, Matsuda Z. The membrane-spanning domain of gp41 plays a critical role in intracellular trafficking of the HIV envelope protein. Retrovirology. 2010;7:95. doi: 10.1186/1742-4690-7-95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Owens RJ, Burke C, Rose JK. Mutations in the membrane-spanning domain of the human immunodeficiency virus envelope glycoprotein that affect fusion activity. J Virol. 1994;68(1):570–574. doi: 10.1128/jvi.68.1.570-574.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Apellaniz B, Rujas E, Serrano S, Morante K, Tsumoto K, Caaveiro JM, Jimenez MA, Nieva JL. The atomic structure of the HIV-1 gp41 transmembrane domain and its connection to the immunogenic membrane-proximal external region. J Biol Chem. 2015;290(21):12999–13015. doi: 10.1074/jbc.M115.644351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Freed EO, Martin MA. The role of human immunodeficiency virus type 1 envelope glycoproteins in virus infection. J Biol Chem. 1995;270(41):23883–23886. doi: 10.1074/jbc.270.41.23883. [DOI] [PubMed] [Google Scholar]
- 15.Dubay JW, Roberts SJ, Brody B, Hunter E. Mutations in the leucine zipper of the human immunodeficiency virus type 1 transmembrane glycoprotein affect fusion and infectivity. J Virol. 1992;66(8):4748–4756. doi: 10.1128/jvi.66.8.4748-4756.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Yu X, Yuan X, McLane MF, Lee TH, Essex M. Mutations in the cytoplasmic domain of human immunodeficiency virus type 1 transmembrane protein impair the incorporation of Env proteins into mature virions. J Virol. 1993;67(1):213–221. doi: 10.1128/jvi.67.1.213-221.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ohno H, Aguilar RC, Fournier MC, Hennecke S, Cosson P, Bonifacino JS. Interaction of endocytic signals from the HIV-1 envelope glycoprotein complex with members of the adaptor medium chain family. Virology. 1997;238(2):305–315. doi: 10.1006/viro.1997.8839. [DOI] [PubMed] [Google Scholar]
- 18.Chen B, Chou JJ. Structure of the transmembrane domain of HIV-1 envelope glycoprotein. FEBS J. 2016; [DOI] [PMC free article] [PubMed]
- 19.Masibay AS, Balaji PV, Boeggeman EE, Qasba PK. Mutational analysis of the Golgi retention signal of bovine beta-1,4-galactosyltransferase. J Biol Chem. 1993;268(13):9908–9916. [PubMed] [Google Scholar]
- 20.Banfield DK. Mechanisms of protein retention in the Golgi. Cold Spring Harb Perspect Biol. 2011;3(8):a005264. doi: 10.1101/cshperspect.a005264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lankford SP, Cosson P, Bonifacino JS, Klausner RD. Transmembrane domain length affects charge-mediated retention and degradation of proteins within the endoplasmic reticulum. J Biol Chem. 1993;268(7):4814–4820. [PubMed] [Google Scholar]
- 22.Cosson P, Lankford SP, Bonifacino JS, Klausner RD. Membrane protein association by potential intramembrane charge pairs. Nature. 1991;351(6325):414–416. doi: 10.1038/351414a0. [DOI] [PubMed] [Google Scholar]
- 23.Kaiser HJ, Orlowski A, Rog T, Nyholm TK, Chai W, Feizi T, Lingwood D, Vattulainen I, Simons K. Lateral sorting in model membranes by cholesterol-mediated hydrophobic matching. Proc Natl Acad Sci U S A. 2011;108(40):16628–16633. doi: 10.1073/pnas.1103742108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Herzig Y, Sharpe HJ, Elbaz Y, Munro S, Schuldiner M. A systematic approach to pair secretory cargo receptors with their cargo suggests a mechanism for cargo selection by Erv14. PLoS Biol. 2012;10(5):e1001329. doi: 10.1371/journal.pbio.1001329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Len AC, Starling S, Shivkumar M, Jolly C. HIV-1 activates T cell signaling independently of antigen to drive viral spread. Cell Rep. 2017;18(4):1062–1074. doi: 10.1016/j.celrep.2016.12.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Longo PA, Kavran JM, Kim MS, Leahy DJ. Transient mammalian cell transfection with polyethylenimine (PEI) Methods Enzymol. 2013;529:227–240. doi: 10.1016/B978-0-12-418687-3.00018-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Rubin LA, Kurman CC, Biddison WE, Goldman ND, Nelson DL. A monoclonal antibody 7G7/B6, binds to an epitope on the human interleukin-2 (IL-2) receptor that is distinct from that recognized by IL-2 or anti-tac. Hybridoma. 1985;4(2):91–102. doi: 10.1089/hyb.1985.4.91. [DOI] [PubMed] [Google Scholar]
- 28.Perrin J, Le Coadic M, Vernay A, Dias M, Gopaldass N, Ouertatani-Sakouhi H, Cosson P. TM9 family proteins control surface targeting of glycine-rich transmembrane domains. J Cell Sci. 2015;128(13):2269–2277. doi: 10.1242/jcs.164848. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.