

Abstract
Background
A key feature of joints in rheumatoid arthritis (RA) is the formation of hyperplastic destructive pannus tissue, which is orchestrated by activated fibroblast-like synoviocytes (FLS). We have demonstrated that the RA risk gene and tumor suppressor Limb bud and heart development (LBH) regulates cell cycle progression in FLS. Methotrexate (MTX) is the first-line treatment for RA, but its mechanisms of action remain incompletely understood. Here, we studied the effects of MTX on mitogen-induced FLS proliferation and expression of cell cycle regulators in vitro.
Methods
Primary FLS from patients with RA or osteoarthritis were stimulated with the mitogen platelet-derived growth factor (PDGF) and the cytokine interleukin-1β (IL-1β) in the presence or absence of MTX. Cells were then subjected to qPCR for gene expression and cell cycle analysis by flow cytometry.
Results
Stimulation with PDGF and IL-1β increased the percentage of FLS in the G2/M phase and shifted the cell morphology to a dendritic shape. These effects were inhibited by MTX. Furthermore, PDGF + IL-1β reduced LBH mRNA expression. However, MTX treatment yielded significantly higher transcript levels of LBH, and of CDKN1A (p21) and TP53 (p53), compared to untreated samples upon mitogen stimulation. The expression of DNA methyltransferase-1 (DNMT1) was also higher in the presence of MTX and there was strong correlation between DNMT1 and LBH expression.
Conclusions
Therapeutic concentrations of MTX abolish the effects of PDGF and IL-1β on tumor suppressor expression and inhibit mitogen-promoted FLS proliferation. These data demonstrate novel and important effects of MTX on pathogenic effector cells in the joint, which might involve epigenetic mechanisms.
Keywords: Rheumatoid arthritis, Fibroblast-like synoviocytes, Cell cycle, Platelet-derived growth factor, Interleukin-1beta, Methotrexate
Background
The rheumatoid arthritis (RA) joint pathology is characterized by persistent inflammation and synovial hyperplasia, leading to destruction of cartilage and bone and ultimately functional disability [1]. Key players in these processes are fibroblast-like synoviocytes (FLS), which develop an aggressive, tumor-like phenotype [2] with increased invasiveness and disturbed control of cell proliferation [3]. The local FLS expansion in the synovial lining layer correlates with clinical disease activity parameters in RA [4]. Also, treatment with cell cycle inhibitors targeting FLS ameliorates arthritis in mouse models [5, 6]. Thus, studies of pathways regulating the proliferation of FLS are of considerable interest.
FLS in RA are activated to an aggressive phenotype by repeated exposure to numerous pro-inflammatory cytokines and growth factors acting on top of predisposing genetic and epigenetic factors [7, 8]. Stimulating FLS in vitro with tumor necrosis factor (TNF) and interleukin-1β (IL-1β) induces a variable expression of many of the genes characterizing the aggressive RA FLS phenotype. Platelet-derived growth factor (PDGF) is a known fibroblast mitogen and both PDGF and IL-1β have been reported to increase FLS proliferation [9, 10]. Interestingly, the cytokine (TNF or IL-1β) potentiates the effects of growth factors (PDGF and transforming growth factor (TGF)-β) on cytokine and matrix metalloproteinase production in FLS [11].
Moreover, RA FLS display intrinsic dysregulation of proliferation and apoptosis pathways, such as abnormalities of the tumor suppressor p53 [12] and its downstream mediator p21 [13] which in the normal synoviocyte induces cell cycle arrest. The number of p21-positive FLS inversely correlates with the thickness synovial lining layer in RA [13]. Recently, the transcription regulator, Limb bud and heart development (LBH), was identified as a new candidate gene for the pathogenic FLS phenotype in RA [14]. Reduced expression of this tumor suppressor in FLS has been suggested to increase the risk of RA development. LBH is regulated by growth factors and epigenetic mechanisms [15] and modulates proliferation in primary FLS by regulating cell cycle progression at the G1 to S phase transition checkpoint [16, 17]. In particular, a regulatory DNA element in the LBH gene region is hypo-methylated in RA. This DNA element is believed to negatively regulate LBH transcription.
Methotrexate (MTX) is still the first-line treatment for RA, but its exact mechanisms of action and target cells remain incompletely understood. The anti-inflammatory effect of low-dose MTX used in rheumatic diseases has in part been attributed to release of adenosine [18]. Studies have also demonstrated that MTX treatment restores global DNA methylation of peripheral blood mononuclear cells [19] and increases expression of cell cycle checkpoint genes in the T cells in patients with RA [20]. However, the effects of MTX on FLS in RA are largely unknown.
In order to further understand the pharmacodynamics of MTX in RA and to identify novel drug targets and markers to predict drug response, this study aimed to investigate the effects of MTX on mitogen-induced FLS proliferation in vitro, and in particular on the expression of cell cycle regulators.
Methods
Synovial samples
Human synovial tissue specimens were obtained from patients with RA (n = 6) or osteoarthritis (OA) (n = 7) during joint replacement surgery at Sahlgrenska University Hospital in Sweden. All Patients with RA fulfilled the American College of Rheumatology 1987 revised criteria for the disease [21] and all had established seropositive and erosive disease. MTX was stopped ≥ 2 weeks before surgery. The procedures were approved by the Ethics Committee of Gothenburg, and all patients gave written informed consent.
Cell culture
Primary FLS were isolated as previously described [2] and cultured in Dulbecco’s modified Eagle’s medium (DMEM) GlutaMAX (Gibco, Carlsbad, CA, USA) supplemented with antibiotics (penicillin/streptomycin, gentamicin) and 10% heat-inactivated fetal bovine serum (FBS), in a humidified 5% CO2 atmosphere. Cells at passages three to nine were seeded into 6-well or 24-well plates for experiments. Stimulation with the human recombinant proteins PDGF-BB (Invitrogen, Carlsbad, CA, USA) and IL-1β (R&D Systems, Minneapolis, MN, USA) was performed in 1% FBS medium after overnight serum starvation.
MTX dose and treatment protocol
Therapeutic doses of MTX range from 10 to 25 mg weekly among patients with RA, resulting in peak serum concentrations at 1 μM [22, 23]. Concentrations are higher in synovial tissue than in blood [24]. In previous in vitro studies of FLS, concentrations of MTX up to 100 μM have been used [25]. Treatment of FLS cultures for 48 h with 0.01–10 μM MTX had no effect on viability or apoptosis of FLS in RA [26].
Here, FLS were pretreated for 24 h with MTX (Apoteket, Sweden) or control medium, then stimulated with PDGF + IL-1β in 1% FBS medium in the presence or absence of MTX. The cells were harvested 24–48 h after mitogen stimulation for gene expression or cell cycle analysis. Only FLS lines responding to mitogen stimulation (defined by a fold change in LBH mRNA below 0.3 compared to the unstimulated control) under the present experimental time frame were included.
Cell cycle analysis
FLS were detached by trypsin-EDTA and resuspended in PBS, fixed in 70% ethanol and stained with the DNA label 7-amino-actinomycin D (7-AAD) (BD Pharmingen, San Diego, CA, USA). DNA content was measured using a FACSVerse flow cytometer and FACSuite software (BD Biosciences) on at least 10,000 cells/sample. Data were analyzed using FlowJo software (Tree Star, Ashland, OR, USA) and the percentages of cells in G1 and G2/M phase, respectively, were calculated from histograms of DNA content after exclusion of debris and doublets. The gating strategy is presented in Fig. 1a.
Fig. 1.
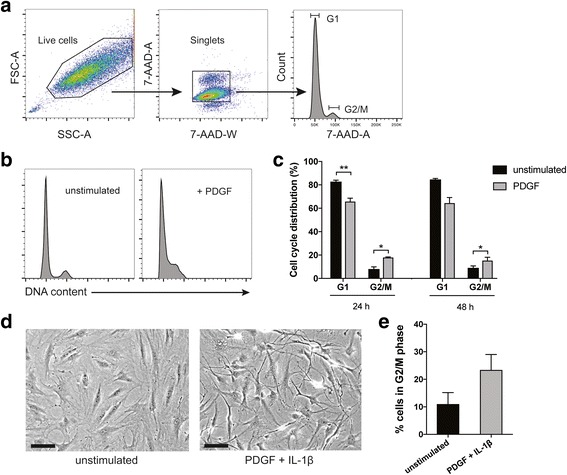
Effects of mitogens on the cell cycle and cell morphology of fibroblast-like synoviocytes (FLS). Primary rheumatoid arthritis (RA) FLS were serum-starved and stimulated with platelet-derived growth factor (PDGF) or a combination of PDGF and IL-1β. The cells were then harvested and DNA-stained followed by flow cytometry. a Gating strategy identifying live cells and singlets and histogram gating of cell cycle analysis by DNA content. b Representative histograms of FLS in an unstimulated state and upon PDGF stimulation. c Percentage of cells in the G1 and G2/M phase, respectively, in unstimulated FLS and upon stimulation with PDGF (20 ng/mL) for 24 and 48 h, respectively. d Morphology of FLS in 2D culture, before stimulation and after incubation for 48 h with PDGF (20 ng/mL) + IL-1β (2 ng/mL), as visualized by light microscopy. Scale bar 20 μM. e Percentage of cells in the G2/M phase upon PDGF + IL-1β stimulation compared to percentage of cells without stimulation. Values are the mean ± SEM of three different FLS lines. *p < 0.05, **p < 0.01. FSC-A, forward scatter-area; SSC-A, side scatter-area
RNA extraction and gene expression analysis
Total RNA was isolated from cells using RNeasy Mini Kit (Qiagen) and quantified with a NanoDrop 100 spectrophotometer (Thermo Scientific). Complementary DNA was synthesized using TaqMan reverse transcription agents (Applied Biosystems, Carlsbad, CA, USA) with 250 ng RNA per reaction. Quantitative polymerase chain reaction (qPCR) was performed on a ViiA Real-Time PCR System using TaqMan Gene Expression Assays (Applied Biosystems) for GAPDH (Hs99999905_m1), LBH (Hs00368853_m1), CDKN1A (Hs00355782_m1), CDKN2A (Hs00923894_m1), TP53 (Hs01034249_m1), CCND1 (Hs00765553_m1), CCNE1 (Hs01026536_m1), CDC25A (Hs00947994_m1), DNMT1 (Hs00154749_m1), and DNMT3A (Hs01027166_m1). Cycle threshold (Ct) values were normalized to GAPDH. Results were presented as fold changes calculated by the 2-ΔΔCt method.
Statistical analysis
Results are presented as the mean ± SEM. Differences were evaluated by paired Student’s t test for comparison of two groups, or one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test for comparison of multiple groups. Shapiro-Wilk’s normality test was used to assess sample distribution. Statistical analysis of differences in gene expression was performed on ΔCt values. Correlation between fold changes in gene expression was tested with Spearman’s rank correlation coefficient. The analyses were performed using GraphPad Prism software (version 7.0b; GraphPad Inc., La Jolla, CA, USA): p values less than 0.05 were considered significant.
Results
PDGF and IL-1β induce FLS proliferation
It has previously been reported that IL-1β enhances FLS proliferation in vitro as measured by 3H-thymidine incorporation [10], and that PDGF induces colony growth of primary FLS [9]. In this study, we used cell cycle analysis by flow cytometry to assay the effects of the mitogen on cultured RA FLS. Stimulation with PDGF 20 ng/mL alone promoted cell cycle progression as expected (Fig. 1b and c), increasing the percentage of cells in the G2/M phase (18 ± 1% versus 8 ± 2% in unstimulated controls, p = 0.03) while reducing the percentage of cells in the G1 phase (66 ± 3% versus 83 ± 1% in unstimulated controls, p = 0.009), after 24 h. Similar results were observed after 48 h (Fig. 1c). Upon stimulation with a combination of PDGF 20 ng/mL and IL-1β 2 ng/mL, a pronounced shift in FLS morphology to a more dendritic cell shape was seen, with cells extending long pseudopodia (Fig. 1d). The mean percentage of cells in the G2/M phase was also higher with this combination compared to unstimulated cells (Fig. 1e), although not statistically significant (23 ± 6% of cells in the G2/M phase versus 11 ± 4% in unstimulated controls, p = 0.14).
MTX inhibits the effects of mitogens on FLS proliferation
To investigate the effects of MTX on the proliferation and gene expression of RA FLS in an inflammatory setting, we developed an experimental set-up as overviewed in Fig. 2a. In cell cycle analysis by flow cytometry, MTX treatment alone did not affect the DNA content distribution seen in unstimulated cultures (Fig. 2b). Stimulation with PDGF + IL-1β could induce cell cycle progression, but this effect was abolished by the presence of MTX. Treatment with MTX resulted in only 5 ± 2% of the cell population in the G2/M phase upon mitogen stimulation, compared to 25 ± 4% on mitogen stimulation alone (p = 0.02) (Fig. 2c). Instead, cells were halted in the G1 phase when MTX was present upon mitogen stimulation (87 ± 3% versus 71 ± 6% in the absence of MTX, p = 0.04) (Fig. 2c).
Fig. 2.
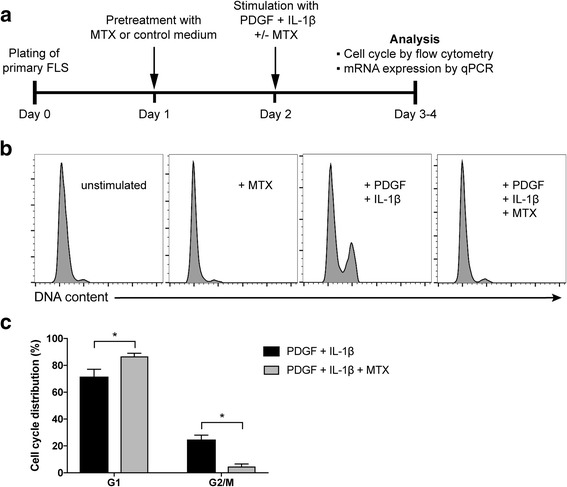
Effects of methotrexate (MTX) on mitogen-induced cell cycle progression in fibroblast-like synoviocytes (FLS). a Design and timeline of the current MTX experiments. b Rheumatoid arthritis (RA) FLS were stimulated with platelet-derived growth factor (PDGF, 20 ng/mL) + IL-1β (2 ng/mL) for 24 h in the presence or absence of 1 μM MTX, or treated with MTX only. The cells were then DNA-stained followed by cell cycle analysis by flow cytometry. Representative histograms of DNA content with indicated treatments are shown. c Percentage of cells in the G1 and G2/M phase, respectively, in three different FLS lines. The experiments were repeated more than three times. Values are mean ± SEM. *p < 0.05. OA, osteoarthritis
PDGF and IL-1β reduce LBH expression
It has recently been demonstrated that LBH is a modulator of FLS proliferation and that it is regulated by growth factors implicated in RA. PDGF has been shown to reduce LBH expression in a dose-dependent manner and with a peak effect at 12 h [16]. We found that stimulation with IL-1β also reduced LBH mRNA expression in FLS and to the same extent as with PDGF (Fig. 3a). Significant effects of IL-1β were obtained at 2 ng/mL and 20 ng/mL (Fig. 3b). The combination of PDGF + IL-1β yielded the strongest effect on LBH expression (Fig. 3a) and was therefore employed in subsequent experiments.
Fig. 3.
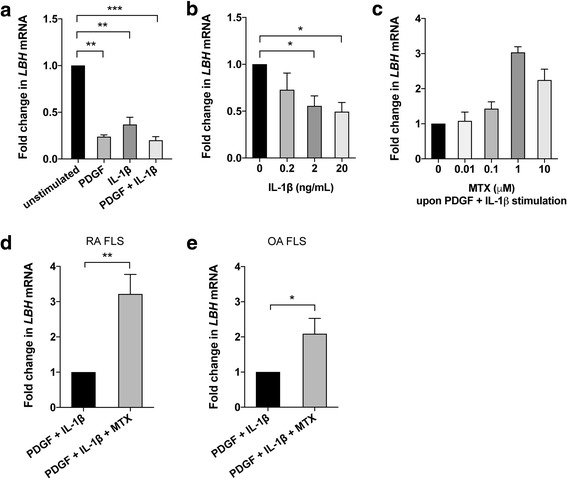
LBH expression in fibroblast-like synoviocytes (FLS) in response to platelet-derived growth factor (PDGF) and IL-1β, with or without methotrexate (MTX). a, b Fold change in mRNA levels of LBH compared to unstimulated was assessed by qPCR in serum-starved FLS after stimulation for 12 h with PDGF (10 ng/mL) and/or IL-1β (2 ng/mL) (a), or IL-1β at different concentrations (0.2–20 ng/mL) (b). c Fold change in LBH mRNA levels was measured after 48 h of mitogen stimulation (PDGF 20 ng/mL + IL-1β 2 ng/mL) in the presence or absence of MTX at various doses (0.01–10 μM; n = 2). d, e Treatment with 1 μM MTX upon PDGF 20 ng/mL + IL-1β 2 ng/mL stimulation was evaluated in rheumatoid arthritis (RA) FLS (d) and in osteoarthritis (OA) FLS (e). Values are the mean fold change ± SEM of three to six different FLS lines. *p < 0.05, **p < 0.01, ***p < 0.0001
MTX inhibits the effects of mitogens on LBH expression
Next, we examined if MTX inhibited the effects of PDGF + IL-1β on LBH expression in FLS. In the presence of MTX, there was a dose-dependent increase in LBH mRNA levels after mitogen stimulation compared to cultures without MTX. The optimal concentration was found to be 1 μM MTX (Fig. 3c), which corresponds to concentrations found in patients treated with therapeutic doses of the drug (see “Methods”). LBH expression was increased 3.2 ± 0.5 fold (p = 0.002) after MTX treatment in RA FLS (Fig. 3d). Since osteoarthritis (OA) FLS also can be “activated” to some extent under certain conditions in vitro [27], we repeated the experiment with OA samples. We found that stimulation with PDGF + IL-1β reduced LBH expression and MTX treatment increased the expression 2.2 ± 0.5 fold (p = 0.02) in OA FLS (Fig. 3e). The difference between RA and OA FLS was not statistically significant under the present conditions.
MTX abolishes the effects of mitogens on CDKN1A and TP53 expression
We continued to study the effects of MTX on other cell cycle regulator genes in RA FLS, as presented in Fig. 4a. CDKN1A encodes the protein p21 (CIP1/WAF1), which regulates cell cycle progression primarily by acting as a cyclin-dependent kinase inhibitor (CDKI) at the G1 to S phase transition. Strikingly, the CDKN1A mRNA expression was 14.3 ± 4.4 fold higher (p = 0.006) in the presence of MTX upon mitogen stimulation compared to without MTX. Another CDKI, p16 encoded by the CDKN2A gene, was also significantly increased (2.0 ± 0.1 fold, p = 0.002). One of the regulators of p21 is the transcription factor p53 (encoded by TP53), known to inhibit cell cycle activity and inflammation. The mRNA levels of TP53 were 1.8 ± 0.2 fold higher (p = 0.03) in the presence of MTX. Within the experimental time frame, we did not detect any significant effects on the transcription of CCND1 (cyclin D1), CCNE1 (cyclin E1) or CDC25A (M-phase inducer phosphatase 1).
Fig. 4.
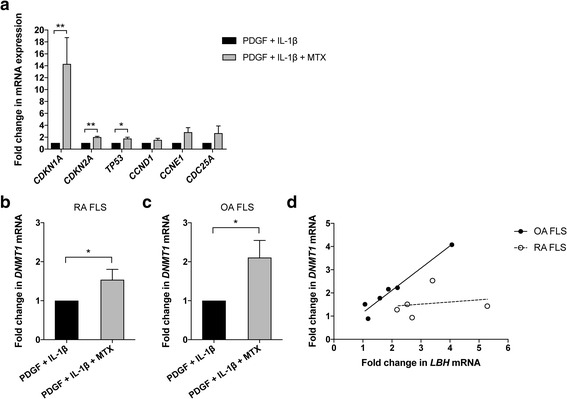
Expression of cell cycle regulator genes and DNMT1 in response to mitogens +/− methotrexate (MTX). Treatment of fibroblast-like synoviocytes (FLS) was performed according to the protocol in Fig. 2a (platelet-derived growth factor (PDGF) 20 ng/mL + IL-1β 2 ng/mL +/− MTX 1 μM). mRNA expression was analyzed by qPCR after 48 h of mitogen stimulation. a Fold change in mRNA levels of the indicated cell cycle regulator genes. b, c Fold change in DNMT1 expression in rheumatoid arthritis (RA) FLS (b) and in osteoarthritis (OA) FLS (c). Values are the mean fold change ± SEM in three to six different FLS lines. *p < 0.05, **p < 0.01. d Correlation between DNMT1 and LBH expression in OA FLS (solid circles; n = 6) and RA FLS (open circles; n = 5). Each dot represents an individual FLS line. Values are fold change in mRNA expression on PDGF + IL-1β + MTX treatment compared to PDGF + IL-1β alone. Lines show fitting of data to a linear regression
DNMT1 expression is increased after MTX treatment and correlates with LBH
It has been demonstrated that IL-1β reduces the expression of DNA methylating enzymes and global DNA methylation in FLS [28]. Since MTX has previously been reported to alter DNA methylation in lymphocytes and monocytes in patients with RA, we also assessed the expression of DNA methyltransferases (DNMTs) in FLS in our experimental setting. Stimulation with PDGF + IL-1β reduced DNMT1 mRNA expression, confirming earlier reports. However, in the presence of MTX the DNMT1 transcript levels were significantly higher compared to mitogen-stimulated FLS without MTX treatment, both in RA FLS (1.5 ± 0.3 fold, p = 0.04) (Fig. 4b) and OA FLS (2.1 ± 0.4 fold, p = 0.03) (Fig. 4c). There were no significant differences in DNMT3A expression (data not shown). Recent data suggest that the expression of LBH is in part regulated by DNA methylation in FLS [15]. We found strong correlation between fold change in DNMT1 and LBH mRNA expression in the OA samples (Spearman’s correlation coefficient, rs = 0.94, p = 0.017) but not in RA FLS (rs = 0.30, p = 0.68). Interestingly, the RA and OA samples clustered in distinct groups (Fig. 4d).
Discussion
A hallmark of the pathogenesis of RA is the formation of destructive pannus tissue in which activated FLS play a key role. This hyperplastic invasive tissue is overpopulated by FLS mediating persistent inflammation and joint destruction by production of pro-inflammatory cytokines, chemokines, growth factors and matrix degrading enzymes. Both increased resistance to apoptosis and increased proliferation is believed to cause the increased FLS number [8] and the predominant mechanism most probably varies by disease duration and between individuals. Targeting cytokines and immune cells in the treatment of RA has proven insufficient in a substantial number of patients and there is a need for more efficient treatment. Studies of the mechanisms regulating FLS proliferation are therefore of utter importance, as there is currently no therapy specifically targeting these pivotal effector cells in the joint in RA.
Low-dose MTX is the standard of care in the treatment of RA. About one third of the patients will reach low disease activity with MTX mono-therapy and another third will reach good clinical response in combination with a biologic agent [29]. The prevailing theory of the pharmacodynamic effects of MTX is that it targets the activity of immune cells. However, a recent study demonstrates the inhibitory effect of MTX on TNF-induced NF-κB activation in FLS in vitro [30].
We performed in vitro studies where the proliferative FLS phenotype was induced by PDGF + IL-1β. This is to our knowledge the only study investigating effects on cell cycle progression of FLS with this combination. IL-1β has been shown to reduce expression of DNA methylating enzymes (DNMTs) in FLS [28] and might thus act on a different regulatory level (epigenetic) than other cytokines and growth factors and could therefore potentiate or modulate their effects on gene expression. Interestingly, we found that MTX treatment inhibited cell cycle progression in PDGF + IL-1β-stimulated FLS, which suggests a new mechanism of action of this drug.
Furthermore, we studied the effects of MTX on the recently identified RA risk gene, LBH, which encodes a highly conserved transcription regulator and the expression of LBH in FLS is believed to be protective in RA [31]. LBH has been shown to regulate FLS growth by acting as a tumor suppressor [16]. A recent study demonstrating that LBH deficiency exacerbates arthritis in an in vivo model supports these findings [17]. In our experiments, the mitogens PDGF and IL-1β were found to reduce LBH mRNA expression in RA FLS, and MTX treatment abrogated these effects. A pronounced effect of MTX was also seen on the cell cycle checkpoint gene CDKN1A, encoding p21, and to some extent on CDKN2A (p16) and TP53 (p53). This is consistent with earlier studies of T cells in RA, where MTX treatment induces the expression of p21 and p53 [20].
In this and earlier studies, PDGF also reduced LBH expression in OA FLS [16] and we demonstrated that MTX can increase expression of LBH and other tumor suppressor genes after mitogen stimulation not only in RA FLS but also to some extent in OA FLS. The gene expression profiles of primary FLS from patients with OA converge with those of RA FLS under high serum conditions in vitro [27]. This finding suggests some common intrinsic response patterns of FLS, and synovitis is common in OA [32]. However, sustained exposure to growth factors and cytokines, as in the joints in RA, has been demonstrated to prime FLS to enhanced pathogenic responses [33]. This together with genetic and epigenetic factors of RA FLS [8] might explain the differences in the degree of synovial activation and inflammation in the two diseases. Interestingly, treatment with anti-TNF agents has failed to provide symptomatic benefits in OA trials [34] but a recent study also suggests analgesic efficacy of MTX in OA [35]. It is important to point out that the RA synovial samples in this study were obtained from patients with end-stage disease and it is possible that the differences in results between RA and OA FLS would have been larger in FLS from early RA.
We have earlier demonstrated that LBH expression in FLS is regulated by DNA methylation [15]. Since a suggested mechanism of action of MTX in lymphocytes is interference with DNA methylation, we also studied the effects of MTX on DNA methylating enzymes in FLS. Indeed, we found that the mRNA expression of DNMT1 was higher after MTX treatment compared to untreated PDGF + IL1β-stimulated FLS. Moreover, there was strong correlation between DNMT1 and LBH expression in MTX-treated OA FLS, suggesting that MTX might mediate the effects on cell cycle regulator gene expression in FLS by restoring epigenetic patterns. In RA FLS, the samples clustered with high fold change in LBH expression, but the correlation with DNMT1 expression was not present, indicating that other factors also influence gene expression. This is in agreement with our previous findings that LBH expression in RA FLS is regulated by growth factors as well as by DNA methylation and genetic polymorphism of a regulatory element [15].
Conclusion
We demonstrate that therapeutic concentrations of MTX abolish mitogen-promoted proliferation of RA FLS and inhibit the effects of mitogens on cell cycle regulators. This could be one hitherto unknown mode of action contributing to the efficacy of MTX in RA. Understanding the molecular mechanisms of the effects of MTX on FLS is of great importance for the development of novel, more FLS-specific drugs and for identifying biomarkers to predict response to MTX.
Acknowledgements
We gratefully acknowledge Dr Thomas Eisler and Dr Christian Anderberg for participating in the recruitment of patients and establishment of the synovial tissue biobank.
Funding
This work was supported by grants from the Swedish Society of Medicine, the King Gustaf V:s 80-year Foundation, the Professor Nanna Svartz Foundation, the Rune and Ulla Amlöv Foundation, the Gothenburg Region Foundation for Rheumatology Research, and the Swedish Rheumatism Foundation.
Availability of data and materials
The datasets analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- 7-AAD
7-Amino-actinomycin D
- ANOVA
Analysis of variance
- CDKI
Cyclin-dependent kinase inhibitor
- DNMT
DNA methyltransferase
- FBS
Fetal bovine serum
- FLS
Fibroblast-like synoviocytes
- IL
Interleukin
- LBH
Limb bud and heart development
- MTX
Methotrexate
- OA
Osteoarthritis
- PBS
Phosphate-buffered saline
- PDGF
Platelet-derived growth factor
- qPCR
Quantitative polymerase chain reaction
- RA
Rheumatoid arthritis
- TGF
Transforming growth factor
- TNF
Tumor necrosis factor
Authors’ contributions
AKE directed the research. AKE and BB collected the patient samples and established the biobank. AKE and BB designed the experiments. BB and AKE conducted the experiments and acquired the data. AKE, BB and HC analyzed the data. BB drafted the manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to participate
The procedures were approved by the Ethics Committee of Gothenburg in Sweden, Jan 2008. Informed consent was obtained from all patients.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Beatrice Bergström, Email: beatrice.bergstrom@gu.se.
Hans Carlsten, Email: hans.carlsten@rheuma.gu.se.
Anna-Karin Hultgård Ekwall, Phone: +46-31-3426411, Email: ak.ekwall@rheuma.gu.se.
References
- 1.Firestein GS, McInnes IB. Immunopathogenesis of rheumatoid arthritis. Immunity. 2017;46:183–196. doi: 10.1016/j.immuni.2017.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ekwall A-KH, Eisler T, Anderberg C, Jin C, Karlsson N, Brisslert M, et al. The tumour-associated glycoprotein podoplanin is expressed in fibroblast-like synoviocytes of the hyperplastic synovial lining layer in rheumatoid arthritis. Arthritis Res Ther. 2011;13:R40. doi: 10.1186/ar3274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bartok B, Firestein GS. Fibroblast-like synoviocytes: key effector cells in rheumatoid arthritis. Immunol Rev. 2010;233:233–255. doi: 10.1111/j.0105-2896.2009.00859.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Izquierdo E, Cañete JD, Celis R, Del Rey MJ, Usategui A, Marsal S, et al. Synovial fibroblast hyperplasia in rheumatoid arthritis: clinicopathologic correlations and partial reversal by anti-tumor necrosis factor therapy. Arthritis Rheum. 2011;63:2575–2583. doi: 10.1002/art.30433. [DOI] [PubMed] [Google Scholar]
- 5.Sekine C, Sugihara T, Miyake S, Hirai H, Yoshida M, Miyasaka N, et al. Successful treatment of animal models of rheumatoid arthritis with small-molecule cyclin-dependent kinase inhibitors. J Immunol. 2008;180:1954–1961. doi: 10.4049/jimmunol.180.3.1954. [DOI] [PubMed] [Google Scholar]
- 6.Hosoya T, Iwai H, Yamaguchi Y, Kawahata K, Miyasaka N, Kohsaka H. Cell cycle regulation therapy combined with cytokine blockade enhances antiarthritic effects without increasing immune suppression. Ann Rheum Dis. 2016;75:253–259. doi: 10.1136/annrheumdis-2014-205566. [DOI] [PubMed] [Google Scholar]
- 7.Croft AP, Naylor A, Zimmermann B, Desanti G, Hardie D, Muller-Ladner U, et al. Synovial fibroblasts differentiate into distinct functional subsets in the presence of cytokines and cartilage. Arthritis Res Ther. 2016;18:270. doi: 10.1186/s13075-016-1156-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bottini N, Firestein GS. Duality of fibroblast-like synoviocytes in RA: passive responders and imprinted aggressors. Nat Rev Rheumatol. 2013;9:24–33. doi: 10.1038/nrrheum.2012.190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lafyatis R, Remmers EF, Roberts AB, Yocum DE, Sporn MB, Wilder RL. Anchorage-independent growth of synoviocytes from arthritic and normal joints. Stimulation by exogenous platelet-derived growth factor and inhibition by transforming growth factor-beta and retinoids. J Clin Invest. 1989;83:1267–1276. doi: 10.1172/JCI114011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lacey D, Sampey A, Mitchell R, Bucala R, Santos L, Leech M, et al. Control of fibroblast-like synoviocyte proliferation by macrophage migration inhibitory factor. Arthritis Rheum. 2003;48:103–109. doi: 10.1002/art.10733. [DOI] [PubMed] [Google Scholar]
- 11.Rosengren S, Corr M, Boyle DL. Platelet-derived growth factor and transforming growth factor beta synergistically potentiate inflammatory mediator synthesis by fibroblast-like synoviocytes. Arthritis Res Ther. 2010;12:R65. doi: 10.1186/ar2981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Han Z, Boyle DL, Shi Y, Green DR, Firestein GS. Dominant-negative p53 mutations in rheumatoid arthritis. Arthritis Rheum. 1999;42:1088–1092. doi: 10.1002/1529-0131(199906)42:6<1088::AID-ANR4>3.0.CO;2-E. [DOI] [PubMed] [Google Scholar]
- 13.Perlman H, Bradley K, Liu H, Cole S, Shamiyeh E, Smith RC, et al. IL-6 and matrix metalloproteinase-1 are regulated by the cyclin-dependent kinase inhibitor p21 in synovial fibroblasts. J Immunol. 2003;170:838–845. doi: 10.4049/jimmunol.170.2.838. [DOI] [PubMed] [Google Scholar]
- 14.Whitaker JW, Boyle DL, Bartok B, Ball ST, Gay S, Wang W, et al. Integrative omics analysis of rheumatoid arthritis identifies non-obvious therapeutic targets. PLoS One. 2015;10:e0124254. doi: 10.1371/journal.pone.0124254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hammaker D, Whitaker JW, Maeshima K, Boyle DL, Ekwall A-KH, Wang W, et al. LBH Gene transcription regulation by the interplay of an enhancer risk allele and DNA methylation in rheumatoid arthritis. Arthritis Rheumatol. 2016;68:2637–2645. doi: 10.1002/art.39746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ekwall AKH, Whitaker JW, Hammaker D, Bugbee WD, Wang W, Firestein GS. The rheumatoid arthritis risk gene LBH regulates growth in fibroblast-like synoviocytes. Arthritis Rheumatol. 2015;67:1193–1202. doi: 10.1002/art.39060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Matsuda S, Hammaker D, Topolewski K, Briegel KJ, Boyle DL, Dowdy S, et al. Regulation of the cell cycle and inflammatory arthritis by the transcription cofactor LBH gene. J Immunol. 2017;199:2316–2322. doi: 10.4049/jimmunol.1700719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cronstein BN, Naime D, Ostad E. The antiinflammatory mechanism of methotrexate. Increased adenosine release at inflamed sites diminishes leukocyte accumulation in an in vivo model of inflammation. J Clin Invest. 1993;92:2675–2682. doi: 10.1172/JCI116884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.de Andres MC, Perez-Pampin E, Calaza M, Santaclara FJ, Ortea I, Gomez-Reino JJ, et al. Assessment of global DNA methylation in peripheral blood cell subpopulations of early rheumatoid arthritis before and after methotrexate. Arthritis Res Ther. 2015;17:233. doi: 10.1186/s13075-015-0748-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Spurlock CF, Tossberg JT, Fuchs HA, Olsen NJ, Aune TM, Aune TM. Methotrexate increases expression of cell cycle checkpoint genes via JNK activation. Arthritis Rheum. 2012;64:1780–1789. doi: 10.1002/art.34342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Arnett FC, Edworthy SM, Bloch DA, McShane DJ, Fries JF, Cooper NS, et al. The American Rheumatism Association 1987 revised criteria for the classification of rheumatoid arthritis. Arthritis Rheum. 1988;31:315–324. doi: 10.1002/art.1780310302. [DOI] [PubMed] [Google Scholar]
- 22.Edelman J, Biggs DF, Jamali F, Russell AS. Low-dose methotrexate kinetics in arthritis. Clin Pharmacol Ther. 1984;35:382–386. doi: 10.1038/clpt.1984.47. [DOI] [PubMed] [Google Scholar]
- 23.Combe B, Edno L, Lafforgue P, Bologna C, Bernard JC, Acquaviva P, et al. Total and free methotrexate pharmacokinetics, with and without piroxicam, in rheumatoid arthritis patients. Br J Rheumatol. 1995;34:421–428. doi: 10.1093/rheumatology/34.5.421. [DOI] [PubMed] [Google Scholar]
- 24.Bologna C, Edno L, Anaya J-M, Canovas F, Vanden BM, Jorgensen C, et al. Methotrexate concentrations in synovial membrane and trabecular and cortical bone in rheumatoid arthritis patients. Arthritis Rheum. 1994;37:1770–1773. doi: 10.1002/art.1780371210. [DOI] [PubMed] [Google Scholar]
- 25.Revu S, Neregård P, af Klint E, Korotkova M, Catrina AI. Synovial membrane immunohistology in early-untreated rheumatoid arthritis reveals high expression of catabolic bone markers that is modulated by methotrexate. Arthritis Res Ther. 2013;15:R205. doi: 10.1186/ar4398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Xu K, Cai Y, Lu S-M, Li X-L, Liu L, Li Z, et al. Autophagy induction contributes to the resistance to methotrexate treatment in rheumatoid arthritis fibroblast-like synovial cells through high mobility group box chromosomal protein 1. Arthritis Res Ther. 2015;17:374. doi: 10.1186/s13075-015-0892-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Filer A, Antczak P, Parsonage GN, Legault HM, O’Toole M, Pearson MJ, et al. Stromal transcriptional profiles reveal hierarchies of anatomical site, serum response and disease and identify disease specific pathways. PLoS One. 2015;10:e0120917. doi: 10.1371/journal.pone.0120917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Nakano K, Boyle DL, Firestein GS. Regulation of DNA methylation in rheumatoid arthritis synoviocytes. J Immunol. 2013;190:1297–1303. doi: 10.4049/jimmunol.1202572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Smolen JS, Aletaha D. Rheumatoid arthritis therapy reappraisal: strategies, opportunities and challenges. Nat Rev Rheumatol. 2015;11:276–289. doi: 10.1038/nrrheum.2015.8. [DOI] [PubMed] [Google Scholar]
- 30.Spurlock CF, Gass HM, Bryant CJ, Wells BC, Olsen NJ, Aune TM. Methotrexate-mediated inhibition of nuclear factor κB activation by distinct pathways in T cells and fibroblast-like synoviocytes. Rheumatology (Oxford) 2015;54:178–187. doi: 10.1093/rheumatology/keu279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kim K, Bang S-Y, Lee H-S, Cho S-K, Choi C-B, Sung Y-K, et al. High-density genotyping of immune loci in Koreans and Europeans identifies eight new rheumatoid arthritis risk loci. Ann Rheum Dis. 2015;74:e13. doi: 10.1136/annrheumdis-2013-204749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Mathiessen A, Conaghan PG. Synovitis in osteoarthritis: current understanding with therapeutic implications. Arthritis Res Ther. 2017;19:18. doi: 10.1186/s13075-017-1229-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Crowley T, O’Neil JD, Adams H, Thomas AM, Filer A, Buckley CD, et al. Priming in response to pro-inflammatory cytokines is a feature of adult synovial but not dermal fibroblasts. Arthritis Res Ther. 2017;19:35. doi: 10.1186/s13075-017-1248-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Dimitroulas T, Lambe T, Klocke R, Kitas GD, Duarte RV. Biologic drugs as analgesics for the management of osteoarthritis. Semin Arthritis Rheum. 2017;46:687–691. doi: 10.1016/j.semarthrit.2016.12.001. [DOI] [PubMed] [Google Scholar]
- 35.Wenham CYJ, Grainger AJ, Hensor EMA, Caperon AR, Ash ZR, Conaghan PG. Methotrexate for pain relief in knee osteoarthritis: an open-label study. Rheumatology. 2013;52:888–892. doi: 10.1093/rheumatology/kes386. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets analyzed during the current study are available from the corresponding author on reasonable request.