

Abstract
Chemotherapy and hematopoietic stem cell transplantation are effective treatments for most Hodgkin lymphoma patients, however there remains a need for better tumor-specific target therapy in Hodgkin lymphoma patients with refractory or relapsed disease. Herein, we demonstrate that membrane CD83 is a diagnostic and therapeutic target, highly expressed in Hodgkin lymphoma cell lines and Hodgkin and Reed-Sternberg cells in 29/35 (82.9%) Hodgkin lymphoma patient lymph node biopsies. CD83 from Hodgkin lymphoma tumor cells was able to trogocytose to surrounding T cells and, interestingly, the trogocytosing CD83+T cells expressed significantly more programmed death-1 compared to CD83−T cells. Hodgkin lymphoma tumor cells secreted soluble CD83 that inhibited T-cell proliferation, and anti-CD83 antibody partially reversed the inhibitory effect. High levels of soluble CD83 were detected in Hodgkin lymphoma patient sera, which returned to normal in patients who had good clinical responses to chemotherapy confirmed by positron emission tomography scans. We generated a human anti-human CD83 antibody, 3C12C, and its toxin monomethyl auristatin E conjugate, that killed CD83 positive Hodgkin lymphoma cells but not CD83 negative cells. The 3C12C antibody was tested in dose escalation studies in non-human primates. No toxicity was observed, but there was evidence of CD83 positive target cell depletion. These data establish CD83 as a potential biomarker and therapeutic target in Hodgkin lymphoma.
Introduction
Hodgkin lymphoma (HL) is a B-cell neoplasm that is defined by the presence of Hodgkin Reed-Sternberg cells (HRS). During recent decades, the long-term survival of HL patients has increased, and most patients can be cured through multi-agent chemotherapy, radiotherapy and/or hematopoietic stem cell transplantation.1 Despite this, 25–30% of patients experience either disease relapse or are refractory to chemotherapy and their survival is substantially reduced, especially for elderly patients who do not tolerate intensive therapy.2,3 New targeted therapies for HL are warranted, especially for refractory/relapsed patients and elderly patients where limiting treatment toxicity is essential. Recent studies have focused on the development of therapeutic agents that target HL-specific antigens or regulate the natural immune response in patients. Antibodies targeting HL surface antigens such as CD25 (daclizumab),4 CD20 (rituximab, tositumomab)5,6 or CD30 (brentuximab)7–10 have shown promising results. The programmed death-1(PD-1)/PD-ligand 1 (PD-L1) checkpoint inhibitors (nivolumab, pembrolizumab), that reverse the suppres sive communication between the tumor and immune system in tumor microenvironments have also been effective in HL patients.11–13
To date, the main utility of identifying membrane-bound CD83 has been to define activated dendritic cells (DC), but CD83 is also expressed on the surface of some activated B cells, T cells, macrophages and neutrophils.14–18 In addition to a membrane-bound form, there is a membrane cleaved soluble (s) form of CD83. We reported that lymphoma tumor cells (HL and non-Hodgkin lymphoma [NHL]) expressed CD83 and released sCD83 into serum.19,20 Recombinant sCD83 protein has immune inhibitory function in mice and humans.21,22 Recently, CD83 was identified as one of the four classifiers to distinguish HL with anaplastic lymphoma kinase (ALK)-anaplastic large cell lymphoma.23 Despite its potential as a relatively specific target, CD83 has not been investigated as a therapeutic target on either HL or NHL. We generated a human anti-human CD83 antibody, 3C12C, which prevents graft-versus-host disease (GvHD) but preserves anti-tumor T-cell function in mice after transplantation with human peripheral blood mononuclear cell (PBMC).24,25 The availability of this potential therapeutic anti-CD83 antibody prompted us to investigate CD83 biology in HL. We show herein that an antibody that detects CD83 in paraffin sections stains Hodgkin and Reed-Sternberg (HRS) cells in most HL lymph node biopsy samples, that HL tumor cells secrete sCD83, and the serum sCD83 level in HL patients correlates with the clinical response. The 3C12C antibody, and its toxin conjugate, killed HL lines and 3C12C depleted CD83 target cells in non-human primate studies without any evidence of toxicity.
Methods
HL tissue section and serum samples
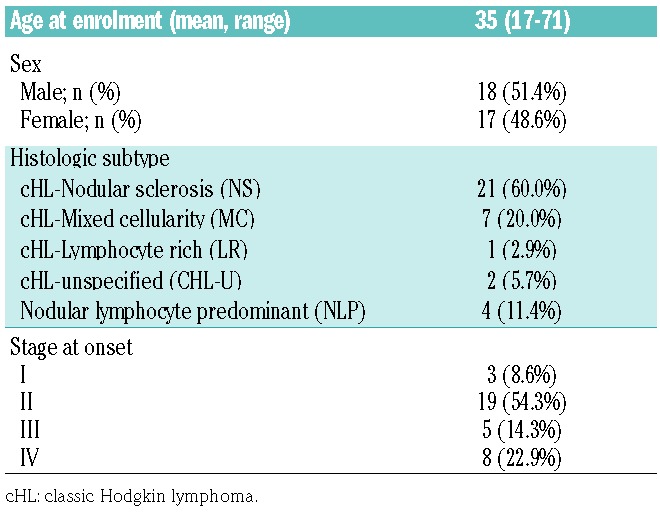
Lymph node biopsies and serum of HL patients were collected after approval by the Sydney Local Health District (SLHD) Human Research Ethics Committee, consistent with the Declaration of Helsinki. Archival paraffin embedded lymph node biopsies were obtained from 35 HL patients at initial diagnosis (Table 1), while serum samples were collected from six HL patients at diagnosis and during chemotherapy.
Table 1.
Characteristics of 35 Hodgkin lymphoma patients.
Immunohistochemistry
Immunohistochemical staining was performed on 3μm sections of formalin fixed paraffin embedded lymph node biopsies of HL patients or non-human primates. The primary antibodies used were mouse anti-human CD20 (L26, Dako), CD83 monoclonal antibodies (mAb; F5, Santa Cruz Biotechnology), CD30 (Ber-H2, Dako), and staining was performed on a Leica Bond III Autostainer (Leica Biosystems) using a Bond Polymer Refine Detection kit for visualization with 3, 3′-diaminobenzidine (DAB). Images were taken with an Olympus BX51 microscopy with an Olympus PP71 camera using Olympus labSens software.
sCD83 analysis
For the analysis of sCD83 levels, KM-H2, L428 and HDLM2 cells were cultured at concentrations of 106 cells/ml in complete roswell park memorial institute (RPMI) medium containing 10% fetal calf serum, 2mM glutaMAX™, 100U/ml penicillin, 100μg/ml streptomycin (Thermo Fisher Scientific) at 37°C, in 5% CO2. Cell culture supernatant were collected 24 hours after fresh medium change. Human sCD83 was analyzed by a sCD83 ELISA kit (Sino Biological Inc.).
Antibody Dependent Cell Cytotoxicity (ADCC) Assays
Target HL cells labeled with 25μM Calcein-AM (Life Technologies) were co-cultured at a ratio of 1:25 with human PBMC of a healthy donor used as effector cells. Supernatants were collected after three hours to measure released calcein using an enzyme-linked immunosorbent assay (ELISA) Reader (Perkin Elmer). The percentage of specific cytolysis was calculated as described.25
3C12C conjugation with monomethyl auristatin E (3C12C-MMAE) and cytotoxicity on CD83+ cell lines
3C12C is a human immunoglobulin G1 (IgG1) anti-human CD83 mAb selected from a phage display library26 and further engineered to improve affinity.25,27 To produce 3C12C-MMAE, a lysosomal cathepsin B-cleavable, self-emolative dipeptide (ValCit) maleimide linker was prepared from MMAE for conjugation to partially reduced 3C12C using a similar method to brentuximab vedotin.28 The cytotoxic activity was assessed by 7-amino-actinomycin D (7AAD, Thermo Fisher Scientific) staining using flow cytometry.
3C12C trials in non-human primates
The SLHD Animal Research Ethics Committee approved the study of five non-human primates (Papio Hamadryas baboon), which received intravenous human-IgG (Intragam, CSL) (10 mg/kg) or 3C12C mAb (1, 5, 10, 10 mg/kg) at days 0, 7, 14 and 21. PBMC were analyzed for immune cell populations including Dendritic cells (DC), T cells and B cells on a Fortessa X20 flow cytometer (BD Biosciences). Liver and kidney function were assessed by measuring alkaline phosphatase (ALP), aspartate transaminase (AST) and creatinine in serum samples using the Cobas 8000 (Roche). Lymph nodes were taken from 3C12C (10mg/kg) or human IgG (10mg/kg) treated animals at day 28 for immunohistological staining.
Statistical Analysis
Mean values with standard error of mean (SEM) bars are shown in graphs. Statistical analyses were performed using Prism 6.0 (GraphPad Software). A Mann-Whitney or one-way analysis of variance (ANOVA) test with Greenhouse-Geisser correction for multiple comparisons were used. Differences with P<0.05 were considered significant.
Results
CD83 is expressed on HL cell lines and HRS cells in lymph node biopsies of HL patients
Expression of CD83 was analyzed using the mouse anti-human antibodies HB15a, HB15e and potential therapeutic human anti-human CD83 antibody 3C12C.25 KM-H2 cells expressed the most expressive CD38 cell surface, stained as it was with all three anti-CD83 antibodies, whilst the L428 and HDLM2 lines expressed less CD83. All three lines expressed CD30 (Figure 1A). This data was confirmed by confocal CD83 staining on KM-H2 cells (Figure 1B), detection of CD83 messenger ribonucleic acid (mRNA) transcripts by reverse transcription polymerase chain reaction (RT-PCR) and intracellular CD83 expression in the three HL lines (Online Supplementary Figure S1).
Figure 1.

CD83 is expressed on Hodgkin lymphoma cell lines. (A) Expression of CD83 was analyzed by flow cytometry on KM-H2, L428 and HDLM2 cell lines, which were stained with HB15a-fluorescein isothiocyanate (FITC), HB15e-FITC or 3C12C-FITC anti-CD83 mAbs, respectively. Gray histograms represent isotype control, while open histograms represent anti-CD83 antibodies. CD30 staining was used as a positive control. These data are representative of three independent experiments with comparable results. (B) CD83 expression (red) on KM-H2 cells with HB15a, HB15e or 3C12C mAb were imaged by confocal microscopy. Nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI; blue). Human IgG1 was used as control for 3C12C mAb. Scale bar: 5μm.
Next, CD83 expression was analyzed on the paraffin-embedded lymph node biopsies of 35 HL patients (Table 1). The HRS cells were identified as CD30+ (Figure 2A). Of note, 8/35 (22.9%) biopsies expressed high levels of CD83 on the HRS cells (>90% positive), 21/35 (60%) expressed middle levels (10–90% positive), and 6/35 (17.1%) expressed low levels of CD83 (<10% positive) (Figure 2B,C). Of the 29 biopsies with high or middle expression, 21 (72.4%) had strong or moderate intensity, while 8/29 (27.6%) showed weak intensity of CD83 on HRS cells (Online Supplementary Table S1). The subtype analysis showed that 16/21 (79.2%) of HRS cells in nodular sclerosis (NS) HL were CD83 high or middle, and 85.7% were CD83 high or middle in mixed cellularity (MC) HL. Most (20/22, 90.9%) of stage I-II HL were CD83 high or middle, and 9/13 (69.2%) HL in stage III-IV were CD83 high or middle. Interestingly, strong CD83 expression on HRS cells was found in two out of three relapsed HL (Online Supplementary Table S1). Epstein-Barr virus (EBV) infection is associated with an increasing risk of developing EBV-positive HL. A number of viral products, including EBV nuclear antigens (EBNA), EBV latent membrane proteins (LMP1 and LMP2) and EBV encoding small ribonucleic acids (RNA; EBER) have been implicated. LMP1 induced CD83 in EBV-infected human B cells by activation of NF-kB.29 CD83/LMP1 has been reported to be correlated in MC HL, but not for NS HL.23 By in situ staining of EBER of 35 HL samples, we found that seven HL were EBER positive, including 2/7(28.6%) MC and 3/22 (13.6%) NS HL (Figure 2D). On six out of seven EBER positive HL samples, CD83 staining of HRS were strong or moderate (Online Supplementary Table S1).
Figure 2.

CD83 is expressed on Hodgkin and Reed-Sternberg (HRS) cells in Hodgkin lymphoma patients. (A) CD83 and CD30 expression (brown) on paraffin-embedded lymph node biopsy samples of HL was imaged by microscopy with ×200 magnification. One representative sample of 35 biopsies shown. (B) Pie chart analysis of CD83 expression level in HRS cells of HL patients (n=35). High: CD83 positive in >90% HRS cells; middle: 10–90% CD83+in HRS cells; low: 10% CD83+ in HRS cells. One representative sample of each group is shown in (C), upper panel: original magnification ×40, lower panel shown with high amplification (×200). Arrows indicate HRS cells expressing CD83. (D) Epstein-Barr virus encoding small ribonucleic acids (RNA; EBER) in 35 HL biopsies were detected by in situ hybridization; one of the seven EBER positive samples is shown.
CD83 is trogocytosed from HL cells to T cells
We found previously that CD83 was able to transfer from the membrane of DC to T cells via trogocytosis.15 Similar trogocytosis was observed to occur between HL cell lines and T cells. When these two cell types were co-cultured for four hours, CD83 surface expression was detected on 5–15% of T cells (Figure 3A,B), whereas no CD83 was detected on T cells in the absence of KM-H2 cells. Furthermore, separating the T and KM-H2 cells during culture by a 0.4μm transwell filter prevented trogocytosis (Online Supplementary Figure S2). To confirm the trogocytosis involved membrane transfer, KM-H2 cells were labeled with fluorescent dye (CellVue Claret) and co-cultured with CD3+ T cells. Cell membrane transfer from KM-H2 cells to T cells was confirmed by flow cytometry and confocal microscopy (Figure 3C,D, and Online Supplementary Figure S2). No differences were observed in the CD4+ and CD8+ T-cell ratio during the co-culture of KM-H2 and T cells within four hours (data not shown). However, the CD83+ T cells expressed significantly higher levels of PD-1 than CD83− T cells (P=0.048) and T cells cultured without KM-H2 (P=0.005) (Figure 3E). The increase in PD-1 was significantly higher on the trogocytosed CD83+CD4+ T cells than non-trogocytosed CD83− T cells (P=0.049). In contrast, no difference in PD-1 expression was seen between the CD83+ and CD83− CD8+ T cells, (P=0.185) although both KM-H2 co-cultured CD4+ and CD8+ T cells had higher PD-1 expression than T cells cultured alone (Figure 3F,G). The CD83+CD4+ T cells had the same proportion of regulatory T cells (Treg) as non-trogocytosed CD4+ T cells (Online Supplementary Figure S2).
Figure 3.

Trogocytosis of CD83 molecule from Hodgkin and Reed-Sternberg cells to T cells. (A) T cells from healthy donor PBMCs were co-cultured with KM-H2 cells for four hours at a ratio of 1:5. CD83 and CD30 expression on CD3+T cells was analyzed by flow cytometry, data were from one of seven experiments and summarized data (mean± SEM and P-value) are shown in (B). (C) KM-H2 cells were labeled with CellVue Claret (red) and co-cultured with purified T cells (green) at a ratio of 5:1 for four hours. CellVue Claret and CD83 expression on T cells was analyzed by flow cytometry. (D) Confocal microscopy image of Claret labeled KMH2 cells co-cultured with T cells that stained with biotinylated mouse anti-human CD3 mAb and Stepdavidin-AF488. Nuclei were stained with DAPI. Scale bar: 5μm. Upper insert: trogocytosed T cells, lower insert: non-trogocytosed T cells. Data representative of three experiments. (E) PD-1 expression on CD83+ trogocytosed T cells co-cultured with KM-H2 cells for four hours was determined by flow cytometry (n=4). P-value of one-way ANOVA analysis shown. (F) PD-1 expression on trogocytosed CD4+T or CD8+ T cells after co-culture with KM-H2 cells for four hours was analyzed (n=4). P-value of one-way ANOVA analysis shown. A representative experiment shown in (G). FSC: forward scatter; SSC: side scatter; PD-1: programmed death-1; FMO: fluorescence minus one.
Supernatant from HL cell lines inhibits T-cell proliferation
Surface CD83 can be cleaved into sCD83.15,20 We detected it in the supernatants of activated DC and B lymphocytes,20 as well as the serum of NHL and chronic lymphocytic leukemia patients.30 High levels of sCD83 were found in the supernatant of KM-H2 (460.6±11.8 pg/ml) and L428 (200.8±53.2 pg/ml), but low in HDLM2 (21.67±1.45 pg/ml) (Figure 4A). HL patients had significantly higher serum sCD83 (360.5±54.82 pg/ml, n=10) at diagnosis than healthy donors (52.6±9.5 pg/ml. Figure 4A).
Figure 4.

Soluble CD83 (sCD83) from Hodgkin lymphoma (HL) cell lines inhibits T-cell proliferation which is abolished by binding to 3C12C. (A) sCD83 was detected in the supernatant of KM-H2, L428, and HDLM2 lines that were cultured for 24 hours at 1×106/ml after changing fresh complete Roswell Park Memorial Institute (RPMI) medium and diagnostic sera of HL patients by ELISA. The P-value of A Mann-Whitney test is shown. (B) Carboxyfluorescein N-hydroxysuccinimidyl ester (CFSE) labeled purified T cells were stimulated with CD2/CD3/CD28 beads (3:1) in the presence of 25% supernatant (SN) of KM-H2 or plus 3C12C (5 μg/ml) for five days. Cells were analyzed by flow cytometry and the proliferation index (PI), that is defined as the total number of divisions divided by the number of cells that went into division, were calculated for total CD3+, CD4+ and CD8+ T cells using Flow Jo (n=6). The P-value of one-way ANOVA analysis is shown. (C) Different volumes (v/v) of KM-H2 supernatant were added to CD2/CD3/CD28 microbead-stimulated CFSE-labeled human T cells. T cells were collected and CFSE was analyzed by flow cytometry at day five. The PI and division index (DI), that is the average number of cell divisions that a cell in the original population has undergone, were calculated as indicators for proliferation. Representative data from one of three similar experiments shown. (D) CFSE-labeled T cells were stimulated with CD2/CD3/CD28 microbeads. T cells were then cultured in 25% (v/v) KM-H2 SN with 3C12C (5 and 10 μg/ml). T-cell proliferation was analyzed on day five. (E) The effect of different concentrations of 3C12C on proliferation of CFSE-labeled T cells was determined after CD2/CD3/CD28 microbead stimulation.
We then tested the effect of KM-H2 cell supernatant on T-cell function. KM-H2 supernatant containing sCD83 inhibited CD2/CD3/CD28 bead stimulated T-cell proliferation (Figure 4B) in a dose-dependent manner (Figure 4C). Only proliferation of CD8+ T cells seemed inhibited by KM-H2 supernatant (P=0.09), and not CD4+ T-cell proliferation (P=0.732). Administration of the anti-CD83 antibody, 3C12C, partially abolished the inhibitory effect of KM-H2 supernatant (Figure 4D). 3C12C alone had no effect on T-cell proliferation (Figure 4E).
HL patient serum sCD83 declined to normal levels correlated with a complete or partial response by PET-CT scan
We monitored changes in circulating sCD83 in six HL patients during sequential chemotherapy. All assessments of response were made by positron emission tomography – computed tomography (PET-CT) scan using the Lugano classification system. All patients received 3–6 cycles of chemotherapy; five achieved a complete response (CR) and one patient a partial response (PR) by PET-CT scan (Figure 5, Online Supplementary Table S2). Serum sCD83 decreased, returning to normal levels when the patients had a CR to chemotherapy, as documented by PET-CT scan in patients #1 and #2. In patients #3 and #6, the serum sCD83 level was still elevated when the PET-CT scan showed CR but normalized after one further cycle of chemotherapy. Patient #4 showed a PR prior to cycle 5 by PET-CT-scan, however the serum sCD83 level only started to decrease during cycle #5 reaching a normal range in cycle 6, coinciding with CR. PET-CT scans in patient #5 showed progressive disease (PD) after cycle 2, but a PR after another two cycles of chemotherapy, when the corresponding sCD83 reduced to normal level.
Figure 5.

Time course of soluble CD83 (sCD83) in Hodgkin lymphoma patients during chemotherapy. The sCD83 level in the sera of six HL patients during different cycles of chemotherapy was examined by ELISA. Arrows indicate when PET-CT scans were performed and the results of complete response (CR), partial response (PR) or progressive disease (PD) are noted.
3C12C and 3C12C-MMAE kills HL cell lines
The ADCC activity of the anti-CD83 mAb, 3C12C, was tested on the three HL lines: KM-H2, L428 and HDLM2. Whilst 3C12C killed KM-H2 and L428 efficiently, HDLM2 was relatively resistant to it (Figure 6A). To elucidate this, the stability of 3C12C binding on the HL cell surface were tested. HL lines were cultured in saturating concentration of 3C12C (10 μg/ml) on ice followed by washing off unbound antibody. Cells were then cultured without 3C12C for up to two hours. The remaining 3C12C bound on the cell surface were detected by a secondary anti-human antibody. Though L428 and HDLM2 have a lower level of surface CD83 expression compared to KM-H2, our analysis showed that the 3C12C level on the surface of HDLM2 reduced much faster than on L428, while the 3C12C bound to L428 were far more stable (Figure 6B). This suggested 3C12C was rapidly internalized in KM-H2 and HDLM2, while 3C12C was internalized slower in L428. To investigate further potential therapeutic applications, we generated a 3C12C toxin-conjugate (3C12C-MMAE). In vitro, 3C12C-MMAE killed CD83+ KM-H2 cells most efficiently, followed by HDLM2 and L428, while CD83− HL-60 cells were the least sensitive to 3C12C-MMAE (Figure 6C and Online Supplementary Figure S3). In addition, the intracellular CD83 level in HDLM2 was much higher than L428, lending to more sensitivity of the HDLM2 to the killing of 3C12C-MMAE (Online Supplementary Figure S1).
Figure 6.

3C12C and 3C12C conjugation with monomethyl auristatin E (3C12C-MMAE) kill Hodgkin lymphoma (HL) cell lines in vitro. (A) Target cells KM-H2, L428 or HDLM2, labeled with Calcein-AM were co-cultured with effector cells (human PBMC) at effector: target ratio of 25:1 with increasing 3C12C concentration from 0 μg/ml to 1 μg/ml at 37°C for three hours. Supernatant was collected for fluorescence reading (excitation 485nm, emission 538nm) of released Calcein. Antibody (Ab)-dependent cell cytotoxicity was calculated (n=3). (B) HL cells were cultured in 3C12C saturation concentration (10 μg/ml) on ice followed by intensive washing and culture without 3C12C from 0–2 hours. The remaining levels of 3C12C bound on the cell surface were detected by a secondary anti-human antibody with flow cytometry. The remaining surface level of 3C12C on KM-H2, L428 and HDLM2 was normalized to the level of time 0. (n=3). (C) CD83+ KM-H2, L428, HDLM2 or CD83−HL-60 cells were cultured with different concentrations of 3C12C-MMAE for three days before determining viable cells by 7-amino-actinomycin D (7AAD) staining with flow cytometry. The half maximal inhibitory concentration (IC50) is shown. Data was from one of four representative experiments.
Administration of 3C12C is safe in non-human primates
To “de-risk” the antibodies before advancing 3C12C into a clinical trial, we performed pre-clinical dose-escalation studies of 3C12C in non-human primates. Five baboons were injected intravenously with 3C12C (1, 5, 10 mg/kg on d0, 7, 14, and 21). No adverse clinical events were recorded during follow up for 84 days. We assessed blood counts and biochemistry weekly, and monitored different immune cell populations by flow cytometry or immune histology. Administration of 3C12C did not affect blood cell counts (white blood cells [WBC], red blood cells [RBC], and platelets), liver (ALP and AST) or kidney (creatinine) function (Online Supplementary Figure S4). The total T-cell number, and ratio of CD4+/CD8+ T cells all remained normal up to day 84 (data not shown). However, there was evidence of 3C12C efficacy in that CD1c+DC counts were reduced. We found that baboon blood B cells expressed CD83 as human B cells (data not shown), and reductions in blood B cells were noted by flow cytometry (Figure 7A). In addition, B-cell areas in lymph nodes were reduced in the 3C12C-treated animals (10mg/kg) compared to the control animals (human IgG 10mg/kg) (Figure 7B).
Figure 7.

3C12C reduced B cells in non-human primates. Five nonhuman primates were injected with 3C12C (1, 5, 10, 10 mg/kg, n=4) or human Immunoglobulin G (IgG; 10mg/kg, n=1) at days 0, 7, 14 and 21. Blood and serum samples were collected for cell counts (red cells, white cells and platelets), liver function (ALP and AST levels) and kidney function (creatinine level) analysis. (A) CD19+ B cells were enumerated from PBMC of five animals by flow cytometry. Dashed lines indicate the base cell number at day 0. *indicates one time point when WBC was extremely high on that animal. (B) A lymph node biopsy was taken at day 28 from 3C12C (10mg/kg) and control-treated animals. B cells stained with anti-human CD20 mAb on paraffin-embedded lymph node biopsy samples are shown. One of the two similar results for the two animals receiving 10 mg/kg 3C12C showing reduced B-cell areas compared to the human IgG control animal.
Discussion
HL is driven by the malignant HRS cell, which are of B lineage origin.31,32 A significant number of patients experience relapsed/refractory disease following first-line chemotherapy.33 Less toxic treatments for relapsed/refractory HL would be highly desirable, as exemplified by the introduction of the anti-CD30 antibody drug conjugate (brentuximab).10,34 In the study herein, we were able to identify sCD83 as a new potential biomarker for HL, and CD83 as a target for a therapeutic mAb and derivatives.
CD83 was first described on activated B cells and we originally detected CD83 on HL using frozen sections.19 Our ability to stain paraffin embedded lymph node biopsy samples of HL patients encouraged this study and allows for the assessment of CD83 expression in routine clinical practice. CD83 was highly expressed on HRS cells with a different staining pattern to CD30. Thus, CD83 is potentially another diagnostic marker of HL. More importantly, this work suggests that the majority of HL patients might be suitable for a therapeutic mAb targeting CD83. An anti-CD83 mAb may also work synergistically with chemotherapy, which is similar to the treatment of stage III or IV HL with anti-CD30 antibody drug conjugate (brentuximab).10 As CD83 is inducible, it is possible that either drug induced or inflammatory activation would induce greater CD83 expression on HRS cells. We also identified serum sCD83 as a potential disease marker. Its immunosuppressive effect was reversed by anti-CD83 mAb at levels readily obtained in vivo. We predict that 3C12C would target HL cells directly through ADCC, but it has the additional therapeutic effect of reversing the inherent immunosuppressive effect of CD83. Such a synergistic response has the potential to have a significant clinical effect with limited toxicity.
Recent studies revealed the impact of tumor microenvironment on tumor progression and therapy. HL is a leading example. The low frequency malignant HRS cells secrete several factors and generate a surrounding infiltrate of immune cells that contribute to the pathogenesis of the disease.35–37 CD83 appears to be involved in this process. We previously demonstrated that the transfer of membrane proteins on myeloma cells to T cells disrupted the immune response and was associated with poor prognosis.38 We found that HL tumor cells express CD83 and can transfer surface CD83 molecules by trogocytosis. CD83 transfer from KM-H2 to T cells in vitro was consistent with the finding that some lymphocytes in the lymph node biopsy samples, especially in CD83 high expression patients, expressed CD83. The proportion of Treg in the trogocytosed CD83+CD4+ T cells was not increased, but CD83+ T cells, especially CD4+ T cells, expressed a higher level of PD-1 than CD83−T cells. PD-1 and PD-1L interaction contributes to the immunosuppressive microenvironment of HL.39 Such PD-1high CD83+ T cells might become unresponsive in the tumor microenvironment.40 A CD83 target therapy might be combined with brentuximab and PD-1 blockage to enhance the clinical response.
The serum of some hematopoietic malignancies have increased levels of sCD83.20,30 The supernatant of HL cells inhibited T-cell proliferation, but this inhibitory effect was not related to Treg induction (data not shown). The anti-CD83 mAb, 3C12C, partially abolished the inhibition by KM-H2 supernatant. Thus, sCD83 from the supernatant plays a key inhibitory role on T cells. We also found that HL cell line supernatant inhibited normal B-cell proliferation (data not shown). Other cytokines or soluble factors from the KM-H2 supernatant may also contribute to the inhibitory effect, e.g., sCD30. An 85kDa soluble form of the CD30 molecule (sCD30) has been shown to be released by CD30+ cells in vitro and in vivo. sCD30 was elevated in the serum of HL41,42 and other CD30-expressing tumors, as well as inflammatory conditions with strong T-or B-cell activation. The CD30-Fc fusion protein inhibits T-cell proliferation,43 whilst sCD30 is also involved in the pathogenesis of renal, islet transplant rejection.44,45 The effect of brentuximab on sCD30 has not been investigated. The thymus and activation related chemokine (TARC; CCL17) is expressed by HRS cells.46
TARC is confirmed as a biomarker of HL, since elevated serum TARC reflected the disease activity and correlated with clinical response.47,48 Herein, as well as confirming elevation of serum sCD83 in active HL, we monitored sCD83 on six HL patients who underwent sequential cycles of chemotherapy. A complete response shown by PET-CT scan correlated with the decreased sCD83 levels. Thus sCD83 may be another biomarker candidate for monitoring the potential clinical response. A much larger cohort of HL patients will be monitored prospectively in order to explore this further. Additional investigations regarding the effect of sCD83 on HL biology may well assist therapeutic development in HL. Natural sCD83 has proved difficult to obtain for functional studies, suggesting the sCD83 structure and/or function is sensitive to in vitro manipulation.49
We have developed a human anti-human CD83 mAb, 3C12C, to investigate in clinical trials. It kills HL cells through ADCC, but in order to enhance its activity, we developed a 3C12C toxin conjugate (3C12C-MMAE). The unconjugated parent anti-CD30 antibody SGN-30 had no effect on HL in a study of 38 patients.50 Brentuximab vedotin SGN35, which is a drug conjugate of SGN-30 with MMAE, has, however, proven to be a highly promising drug with CD30+ lymphoma.10 We found 3C12C-MMAE kills CD83+ HL cell lines KM-H2 and HDLM2 very efficiently. Although HDLM2 cells express less surface CD83 and are resistant to ADCC killing, the high killing efficiency of HDLM2 with 3C12C-MMAE is likely related to the rapid antibody internalization and high intracellular CD83 turnover. 3C12C binding on L428 is relatively stable, rendering it sensitive to killing by ADCC, but less likely to be killed with 3C12C-MMAE, which is mediated via antibody internalization. Further improvements in the 3C12C-MMAE conjugate preparation are planned.
Finally, we tested the safety of 3C12C in non-human primates. We saw no clinical toxicity, abnormalities of blood count, liver or renal function or a decrease in the target CD1c+DC population (data not shown). By monitoring B cells, we saw depletion in the blood and lymph nodes. The depletion of activated CD83+ B cells led to a significant reduction in the B-cell area of lymph nodes. This early evidence of CD83 target cell depletion in non-human primates is most encouraging, suggesting an ADCC effect that should translate readily to the clinic.
Taken together, these data demonstrate that CD83 is a new potential diagnostic HL marker and serum sCD83 levels are likely to reflect HL disease load. CD83 is a target for therapeutic mAb development as well as CD83 target derivatives. The potential therapeutic human anti-CD83 antibody, 3C12C, kills HL cells efficiently in vitro. It is safe in non-human primates, and depletes CD83+ target cells. Further development of 3C12C in human studies merits serious consideration.
Supplementary Material
Footnotes
Check the online version for the most updated information on this article, online supplements, and information on authorship & disclosures: www.haematologica.org/content/103/4/655
References
- 1.Ansell SM. Hodgkin lymphoma: 2016 update on diagnosis, risk-stratification, and management. Am J Hematol. 2016;91(4):434–442. [DOI] [PubMed] [Google Scholar]
- 2.Castagna L, Carlo-Stella C, Mazza R, Santoro A. Current role of autologous and allogeneic stem cell transplantation for relapsed and refractory hodgkin lymphoma. Mediterr J Hematol Infect Dis. 2015;7(1):e2015015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Reddy NM, Perales MA. Stem cell transplantation in Hodgkin lymphoma. Hematol Oncol Clin North Am. 2014;28(6):1097–1112. [DOI] [PubMed] [Google Scholar]
- 4.Janik JE, Morris JC, O’Mahony D, et al. 90Y-daclizumab, an anti-CD25 monoclonal antibody, provided responses in 50% of patients with relapsed Hodgkin’s lymphoma. Proc natl Acad Sci USA. 2015;112(42):13045–13050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Rehwald U, Schulz H, Reiser M, et al. Treatment of relapsed CD20+ Hodgkin lymphoma with the monoclonal antibody rituximab is effective and well tolerated: results of a phase 2 trial of the German Hodgkin Lymphoma Study Group. Blood. 2003;101(2):420–424. [DOI] [PubMed] [Google Scholar]
- 6.Jacene H, Crandall J, Kasamon YL, et al. Initial experience with tositumomab and I-131-labeled tositumomab for treatment of relapsed/refractory Hodgkin lymphoma. Mol Imaging Biol. 2017;19(3):429–436. [DOI] [PubMed] [Google Scholar]
- 7.Younes A, Bartlett NL, Leonard JP, et al. Brentuximab vedotin (SGN-35) for relapsed CD30-positive lymphomas. N Engl J Med. 2010;363(19):1812–1821. [DOI] [PubMed] [Google Scholar]
- 8.Deng C, Pan B, O’Connor OA. Brentuximab vedotin. Clin Cancer Res. 2013;19(1):22–27. [DOI] [PubMed] [Google Scholar]
- 9.Gravanis I, Tzogani K, van Hennik P, et al. The European Medicines Agency review of brentuximab vedotin (Adcetris) for the treatment of adult patients with relapsed or fefractory CD30+ Hodgkin lymphoma or systemic anaplastic large cell lymphoma: summary of the scientific assessment of the Committee for Medicinal Products for Human Use. Oncologist 2016;21(1):102–109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Connors JM, Jurczak W, Straus DJ, et al. ‘Brentuximab vedotin with chemotherapy for stage III or IV Hodgkin’s lymphoma. N Engl J Med. 2017. December 12 [Epub ahead of print] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Villasboas JC, Ansell SM. Nivolumab for the treatment of classical Hodgkin lymphoma after failure of autologous stem cell transplant and brentuximab. Expert Rev Anticancer Ther. 2016;16(1):5–12. [DOI] [PubMed] [Google Scholar]
- 12.Ansell SM, Lesokhin AM, Borrello I, et al. PD-1 blockade with nivolumab in relapsed or refractory Hodgkin’s lymphoma. N Engl J Med. 2015;372(4):311–319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kasamon YL, de Claro RA, Wang Y, Shen YL, Farrell AT, Pazdur R. FDA approval summary: nivolumab for the treatment of relapsed or progressive classical Hodgkin lymphoma. Oncologist. 2017;22(5):585–591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zhou LJ, Tedder TF. Human blood dendritic cells selectively express CD83, a member of the immunoglobulin superfamily. J Immunol. 1995;154(8):3821–3835. [PubMed] [Google Scholar]
- 15.Ju X, Silveira PA, Hsu WH, et al. The analysis of CD83 expression on human immune cells identifies a unique CD83+-activated T cell population. J Immunol. 2016;197(12):4613–4625. [DOI] [PubMed] [Google Scholar]
- 16.Kretschmer B, Luthje K, Guse AH, et al. CD83 modulates B cell function in vitro: increased IL-10 and reduced Ig secretion by CD83Tg B cells. PloS One. 2007;2(8):e755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cao W, Lee SH, Lu J. CD83 is preformed inside monocytes, macrophages and dendritic cells, but it is only stably expressed on activated dendritic cells. Biochem J. 2005;385(Pt 1):85–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Yamashiro S, Wang JM, Yang D, Gong WH, Kamohara H, Yoshimura T. Expression of CCR6 and CD83 by cytokine-activated human neutrophils. Blood. 2000;96(12):3958–3963. [PubMed] [Google Scholar]
- 19.Sorg UR, Morse TM, Patton WN, et al. Hodgkin’s cells express CD83, a dendritic cell lineage associated antigen. Pathology. 1997;29(3):294–299. [DOI] [PubMed] [Google Scholar]
- 20.Hock BD, Kato M, McKenzie JL, Hart DN. A soluble form of CD83 is released from activated dendritic cells and B lymphocytes, and is detectable in normal human sera. Int Immunol. 2001;13(7):959–967. [DOI] [PubMed] [Google Scholar]
- 21.Dudziak D, Nimmerjahn F, Bornkamm GW, Laux G. Alternative splicing generates putative soluble CD83 proteins that inhibit T cell proliferation. J Immunol. 2005;174(11):6672–6676. [DOI] [PubMed] [Google Scholar]
- 22.Horvatinovich JM, Grogan EW, Norris M, et al. Soluble CD83 inhibits T cell activation by binding to the TLR4/MD-2 complex on CD14+ monocytes. J Immunol. 2017;198(6):2286–2301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Doring C, Hansmann ML, Agostinelli C, et al. A novel immunohistochemical classifier to distinguish Hodgkin lymphoma from ALK anaplastic large cell lymphoma. Mod Pathol. 2014;27(10):1345–1354. [DOI] [PubMed] [Google Scholar]
- 24.Wilson J, Cullup H, Lourie R, et al. Antibody to the dendritic cell surface activation antigen CD83 prevents acute graft-versus-host disease. J Exp Med. 2009;206(2):387–398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Seldon TA, Pryor R, Palkova A, et al. Immunosuppressive human anti-CD83 monoclonal antibody depletion of activated dendritic cells in transplantation. Leukemia. 2016;30(3):692–700. [DOI] [PubMed] [Google Scholar]
- 26.Sheets MD, Amersdorfer P, Finnern R, et al. Efficient construction of a large nonimmune phage antibody library: the production of high-affinity human single-chain antibodies to protein antigens. Proc Natl Acad Sci U S A. 1998;95(11):6157–6162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Jones ML, Seldon T, Smede M, et al. A method for rapid, ligation-independent reformatting of recombinant monoclonal antibodies. J Immunol Methods. 2010;354(1–2):85–90. [DOI] [PubMed] [Google Scholar]
- 28.Francisco JA, Cerveny CG, Meyer DL, et al. cAC10-vcMMAE, an anti-CD30-monomethyl auristatin E conjugate with potent and selective antitumor activity. Blood. 2003;102(4):1458–1465. [DOI] [PubMed] [Google Scholar]
- 29.Dudziak D, Kieser A, Dirmeier U, et al. Latent membrane protein 1 of Epstein-Barr virus induces CD83 by the NF-kappaB signaling pathway. J Virol. 2003;77(15):8290–8298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hock BD, Haring LF, Steinkasserer A, Taylor KG, Patton WN, McKenzie JL. The soluble form of CD83 is present at elevated levels in a number of hematological malignancies. Leuk Res. 2004;28(3):237–241. [DOI] [PubMed] [Google Scholar]
- 31.Kuppers R. The biology of Hodgkin’s lymphoma. NaR Rev Cancer. 2009;9(1):15–27. [DOI] [PubMed] [Google Scholar]
- 32.Scott DW, Gascoyne RD. The tumour microenvironment in B cell lymphomas. Nat Rev Cancer. 2014;14(8):517–534. [DOI] [PubMed] [Google Scholar]
- 33.Adams HJ, Nievelstein RA, Kwee TC. Systematic review and meta-analysis on the prognostic value of complete remission status at FDG-PET in Hodgkin lymphoma after completion of first-line therapy. Ann Hematol. 2016;95(1):1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Forero-Torres A, Holkova B, Goldschmidt J, et al. Phase 2 study of frontline brentuximab vedotin monotherapy in Hodgkin lymphoma patients aged 60 years and older. Blood. 2015;126(26):2798–2804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Aldinucci D, Gloghini A, Pinto A, De Filippi R, Carbone A. The classical Hodgkin’s lymphoma microenvironment and its role in promoting tumour growth and immune escape. J Pathol. 2010;221(3):248–263. [DOI] [PubMed] [Google Scholar]
- 36.Ma Y, Visser L, Roelofsen H, et al. Proteomics analysis of Hodgkin lymphoma: identification of new players involved in the cross-talk between HRS cells and infiltrating lymphocytes. Blood. 2008;111(4):2339–2346. [DOI] [PubMed] [Google Scholar]
- 37.Vardhana S, Younes A. The immune microenvironment in Hodgkin lymphoma: T cells, B cells, and immune checkpoints. Haematologica. 2016;101(7):794–802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Brown R, Kabani K, Favaloro J, et al. CD86+ or HLA-G+ can be transferred via trogocytosis from myeloma cells to T cells and are associated with poor prognosis. Blood. 2012;120(10):2055–2063. [DOI] [PubMed] [Google Scholar]
- 39.Yamamoto R, Nishikori M, Kitawaki T, et al. PD-1-PD-1 ligand interaction contributes to immunosuppressive microenvironment of Hodgkin lymphoma. Blood. 2008;111(6):3220–3224. [DOI] [PubMed] [Google Scholar]
- 40.Jiang Y, Li Y, Zhu B. T-cell exhaustion in the tumor microenvironment. Cell Death Dis. 2015;6:e1792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Gause A, Pohl C, Tschiersch A, et al. Clinical significance of soluble CD30 antigen in the sera of patients with untreated Hodgkin’s disease. Blood. 1991;77(9):1983–1988. [PubMed] [Google Scholar]
- 42.Nadali G, Vinante F, Ambrosetti A, et al. Serum levels of soluble CD30 are elevated in the majority of untreated patients with Hodgkin’s disease and correlate with clinical features and prognosis. J Clin Oncol. 1994;12(4):793–797. [DOI] [PubMed] [Google Scholar]
- 43.Su CC, Chiu HH, Chang CC, Chen JC, Hsu SM. CD30 is involved in inhibition of T-cell proliferation by Hodgkin’s Reed-Sternberg cells. Cancer Res. 2004;64(6):2148–2152. [DOI] [PubMed] [Google Scholar]
- 44.Billing H, Sander A, Susal C, et al. Soluble CD30 and ELISA-detected human leukocyte antigen antibodies for the prediction of acute rejection in pediatric renal transplant recipients. Transpl Int. 2013;26(3):331–338. [DOI] [PubMed] [Google Scholar]
- 45.Saini D, Ramachandran S, Nataraju A, et al. Activated effector and memory T cells contribute to circulating sCD30: potential marker for islet allograft rejection. Am J Transplant. 2008;8(9):1798–1808. [DOI] [PubMed] [Google Scholar]
- 46.van den Berg A, Visser L, Poppema S. High expression of the CC chemokine TARC in Reed-Sternberg cells. A possible explanation for the characteristic T-cell infiltratein Hodgkin’s lymphoma. Am J Pathol. 1999;154(6):1685–1691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Weihrauch MR, Manzke O, Beyer M, et al. Elevated serum levels of CC thymus and activation-related chemokine (TARC) in primary Hodgkin’s disease: potential for a prognostic factor. Cancer Res. 2005;65(13):5516–5519. [DOI] [PubMed] [Google Scholar]
- 48.Plattel WJ, van den Berg A, Visser L, et al. Plasma thymus and activation-regulated chemokine as an early response marker in classical Hodgkin’s lymphoma. Haematologica. 2012;97(3):410–415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Hock BD, Fernyhough LJ, Gough SM, Steinkasserer A, Cox AG, McKenzie JL. Release and clinical significance of soluble CD83 in chronic lymphocytic leukemia. Leuk Res. 2009;33(8):1089–1095. [DOI] [PubMed] [Google Scholar]
- 50.Forero-Torres A, Leonard JP, Younes A, et al. A Phase II study of SGN-30 (anti-CD30 mAb) in Hodgkin lymphoma or systemic anaplastic large cell lymphoma. Br J Haematol. 2009;146(2):171–179. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.