

Abstract
The protective effect of sevoflurane on apoptosis of rat H9c2 cardiomyocytes induced by H2O2 and the effect on the expression of glucose-regulated protein 78 (GRP78) were investigated. H9c2 cells were routinely cultured and divided into the control, model and sevoflurane groups. Cells in the model group were treated with 400 µM H2O2, and cells in the sevoflurane group were pretreated with sevoflurane prior to treatment with 400 µM H2O2. MTT assay was used to assess cell viability. Annexin V-propidium iodide (AV-PI) double staining flow cytometry was used to detect apoptosis. The intracellular free Ca2+ concentration was measured by the fluorescence-based assay using Fluo-3 AM as a calcium ion fluorescence probe. The mRNA expression level of GRP78 and protein expression levels of GRP78, CHOP and caspase-12 were measured using reverse transcription quantitative polymerase chain reaction (RT-qPCR) and western blot analysis, respectively. The assays showed that after sevoflurane pretreatment the H9c2 cell viability was significantly increased, whereas the H2O2-induced apoptosis, intracellular Ca2+ concentration, mRNA expression of GRP78, and the protein expression of GRP78, CHOP and caspase-12 were all reduced. The results show that pretreatment with sevoflurane inhibited H2O2-induced apoptosis in H9c2 cells. The mechanism may be related to inhibition of the stress-related protein GRP78 expression in endoplasmic reticulum, resulting in decreased intracellular Ca2+ concentration and the downregulation of CHOP and caspase-12 expression levels.
Keywords: sevoflurane, H9c2 cells, H2O2, glucose-regulated protein 78, apoptosis
Introduction
Oxidative stress injury refers to tissue and cell damage due to an imbalance between oxidation and anti-oxidation in the body when exposed to a variety of internal and external stimuli (1). Oxidative stress is considered the main cause of various cardiovascular diseases such as myocardial reperfusion injury, myocardial infarction and atherosclerosis (2). A large amount of reactive oxygen species (ROS) is generated in the process of myocardial ischemia-reperfusion, resulting in the apoptosis of myocardial cells due to oxidative damage. Therefore, oxidative stress is the main culprit of myocardial cell death in myocardial ischemia-reperfusion (2,3).
Hydrogen peroxide (H2O2) is a molecule belonging to ROS that can easily pass through the cell membrane. It converts into other more reactive ROS in the body, which directly oxidize the lipids and proteins on the cell membrane, resulting in apoptosis through multiple pathways (4). At present, H2O2-induced oxidative stress injury of myocardial cells has become a commonly used in vitro experimental model of myocardial ischemia-reperfusion injury (5). Endoplasmic reticulum stress is an imbalance stress process in cell endoplasmic reticulum, which leads to physiological dysfunction (6). It was found that endoplasmic reticulum stress plays a key role in multiple disease processes such as myocardial ischemia-reperfusion injury and myocardial infarction (7).
In clinical applications, sevoflurane was found to exert protective effect on myocardial cells. For example, sevoflurane treatment played a role in enhancing myocardial contractility (6), slowing heart rate (7), lowering left ventricular diastolic pressure (8), and reducing ischemia-reperfusion injury to myocardial cells (9). It was reported that the mechanism underlying inhibition of myocardial cell apoptosis through sevoflurane treatment may be due to the upregulated expression of anti-apoptotic Bcl-2 protein, downregulated expression of pro-apoptotic Bax protein, and resulted inhibition of the mitochondrial apoptotic signaling pathways (10). At present there is no study on whether sevoflurane can reduce endoplasmic reticulum stress to inhibit oxidative stress-induced cardiomyocyte apoptosis. In this study, rat H9c2 cardiomyocytes were used to establish an in vitro experimental model of H2O2-induced oxidative stress to examine the inhibitory effect of sevoflurane pretreatment on H2O2-induced cardiomyocyte injury and apoptosis. In particular, the expression levels of glucose-regulated protein 78 (GRP78) mRNA and protein as stress indicators in endoplasmic reticulum, the intracellular free Ca2+ concentration, and the downstream protein expression levels of CHOP and caspase-12 in endoplasmic reticulum stress were measured, in order to investigate whether sevoflurane-mediated inhibition of H2O2-induced cardiomyocyte apoptosis was associated with the reduction of endoplasmic reticulum stress.
Materials and methods
Materials
The following materials were purchased from commercial sources: Rat embryonic ventricular myocardium H9c2 from the Chinese Academy of Sciences Cell Bank (Beijing, China); Dulbeccos modified Eagles medium (DMEM) medium and fetal bovine serum (FBS) from HyClone Laboratories (Logan, UT, USA); GRP78, CHOP, caspase-12, glyceraldehyde 3-phosphate dehydrogenase (GAPDH) primary antibody, and HRP secondary antibody from Proteintech Group, Inc. (Wuhan, China); BCA protein concentration assay kit, Annexin V-FITC apoptosis detection kit, and Fluo-3 AM calcium ion detection kit from Beyotime Biotechnology, Inc. (Nantong, China); TRIzol kit, reverse transcription kit, and RT-PCR kit from Invitrogen (Carlsbad, CA, USA); primer synthesis from Takara (Dalian, China); hydrogen peroxide, sevoflurane, and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) from Sigma (St. Louis, MO, USA).
Cell culture
H9c2 cells were cultured in DMEM supplemented with 10% fetal bovine serum at 37°C in a humidified incubator containing 5% CO2. When cells covered 80% of the bottom of the culture container, the cells were treated with trypsin to obtain a single cell suspension, and then seeded into different culture plates for group experiments according to the experimental requirement.
Viability of H9c2 cells determined by MTT assay
H9c2 cells in the logarithmic growth phase were collected and seeded into 96-well plates. After 24 h incubation, the cells were divided into the control, model and sevoflurane groups. The cells in the control group received no treatment, cells in the model group were treated with 400 µM H2O2, and cells in the sevoflurane group were pretreated with sevoflurane followed by treatment with 400 µM H2O2. The sevoflurane pretreatment was performed as previously mentioned (11). H9c2 cells were placed in a sterile sealed container connected with the breathing circuit of an anesthesia machine. The sevoflurane vaporizer was filled with sevoflurane and the vaporizer dial was set at 2.5%. The concentration of sevoflurane in the outlet of the closed container was monitored. When it reached 2.5%, the sevoflurane flow was maintained for 20 min, after which the cells were returned to a normal incubator for another 10 min (11). The hydrogen peroxide treatment was carried out by incubation of the cells with a final concentration of 400 µM H2O2 to generate the H2O2 stress injury model (12). After 4 h incubation with H2O2, the culture medium was discarded, and the wells were washed three times with phosphate-buffered saline (PBS), followed by the addition of 100 µl MTT solution (5 mg/ml) per well and incubation for 4 h. Then, 100 µl dimethyl sulfoxide (DMSO) was added to each well, followed by agitation for 10 min and measurement of absorbance value at 570 nm using a microplate reader (model 680; Bio-Rad Laboratories, Hercules, CA, USA). The cell survival rate was calculated using the formula: Survival rate (%) = 100 × (OD value of experimental group/OD value of control group).
Apoptosis of H9c2 cells determined by Annexin V-propidium iodide (AV-PI) double staining assay
H9c2 cells in the logarithmic phase were collected and seeded into 6-well plates. The cells were grouped and processed following the protocol described above. After H2O2 treatment, the cells were treated with 0.25% trypsin, followed by three washes with PBS. The cell concentration was then adjusted to 5×105 cells/ml by the addition of 300 µl binding buffer. Then, 10 µl Annexin V-FITC and 5 µl PI were added to each sample well. After incubation for 15 min at room temperature in the dark, the samples were diluted with another 300 µl binding buffer, followed by filtration through a 300 mesh cell strainer. The apoptotic rate of each sample was measured by flow cytometry (Becton-Dickinson and Company, Franklin Lakes, NJ, USA).
Intracellular Ca2+ levels of H9c2 cells measured by fluorescence-based calcium ion probe using Fluo-3 AM
H9c2 cells in the logarithmic phase were collected and seeded into 6-well plates. The cells were grouped and processed following the protocol described above. After treatment with H2O2, the cells were digested with trypsin followed by centrifugation at 8,000 × g for 10 min. The remaining steps were performed following the instructions of the calcium assay kit. The collected cells were re-suspended in 500 µl staining buffer containing 2.5 µM Fluo-3 AM. After incubation at 37°C for 40 min, the samples were analyzed by flow cytometry (Becton-Dickinson and Company).
Expression levels of GRP78 mRNA in H9c2 cells measured by RT-qPCR
H9c2 cells in the logarithmic phase were collected and seeded into 6-well plates. The cells were grouped and processed following the protocol described above. After treatment with H2O2, the cells were digested with trypsin, followed by centrifugation 8,000 × g. Total RNA was extracted from the collected cells in each group with TRIzol reagent. Then cDNA was synthesized from the RNA template via reverse transcription using oligo(dT) primers. PCR amplification was carried out on the obtained cDNA using the primers listed in Table I. PCR conditions were as follows: 95°C for 10 min; then 40 cycles of 95°C for 15 sec and 60°C for 1 min. The GAPDH gene was used as the reference gene. Cq values were obtained from the instrument outputs. Relative quantification of gene expression between different groups was estimated by calculation of 2−ΔCq, where ΔCqtarget gene = Cqtarget gene-Cqreference gene.
Table I.
RT-qPCR primer sequences.
| Genes | Primer sequences |
|---|---|
| GRP78 | F: 5′-CTGGGTACATTTGATCTGACTGG-3′ |
| R: 5′-GCATCCTGGTGGCTTTCCAGCCATTC-3′ | |
| GAPDH | F: 5′-GCACCGTCAAGGCTGAGAAC-3′ |
| R: 5′-TGGTGAAGACGCCAGTGGA-3′ |
qRT-PCR, quantitative real-time polymerase chain reaction; GRP78, glucose-regulated protein 78; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; F, forward; R, reverse.
Expression levels of proteins GRP78, CHOP and caspase-12 measured by western blot analysis
H9c2 cells in the logarithmic phase were collected and seeded into 6-well plates. The cells were grouped and processed following the protocol described above. After treatment with H2O2, the cells were digested with trypsin, followed by centrifugation. The collected cells were lysed using RIPA cell lysis buffer. Proteins were extracted, and the concentrations were measured using BCA assay. A sample containing 40 µg of proteins from each group was loaded onto a 12% gel for sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) analysis. The protein bands were transferred to PVDF membrane using a wet system. After the membrane was blocked with 5% skimmed milk powder for 2 h, the primary antibodies of GRP78, CHOP, caspase-12 and GAPDH (dilution, 1:1,000; cat. nos. 11587-1-AP, 15204-1-AP, 55238-1-AP and 10494-1-AP) were added, respectively, followed by incubation overnight at 4°C. After the membrane was washed three times with TBST buffer, the HRP-labeled secondary antibody (1:1,000 dilution) was added, followed by incubation at room temperature for 2 h. The ECL reagent was applied to the membrane in the dark after it was washed three times with TBST. The image of the membrane was scanned with a digital imager and was used in statistical analysis.
Statistical analysis. The experimental data were expressed as mean ± standard deviation, and were processed using SPSS 13.0 software (International Business Machines Corp., New York, NY, USA). Data were analyzed with single factor analysis of variance, and t-test was used in paired comparison. Differences were statistically significant at P<0.05.
Results
Effect of sevoflurane on viabilities of H9c2 cells treated with H2O2
Viabilities of H2O2-treated H9c2 cells determined using MTT assay are shown in Fig. 1. Viabilities of H9c2 cells in the model and sevoflurane groups were significantly lower than that in the control group (P<0.01). Moreover, the viability in the sevoflurane-pretreated group was higher than that in the model group (P<0.01).
Figure 1.

Viability of H2O2-treated H9c2 cells compared with untreated cells (**P<0.01 and ##P<0.01).
Effect of sevoflurane on H2O2-induced apoptosis of H9c2 cells
Apoptotic rates of H2O2-treated H9c2 cells determined by AV-PI double staining assay are shown in Fig. 2. Evidently, after H9c2 cells were treated with H2O2, the apoptotic rate in the model group was significantly increased as compared with that in the control group (P<0.01). By contrast, cells pretreated with sevoflurane prior to H2O2 challenge, showed an increased apoptotic rate (sevoflurane group), but which was much lower than that in the model group (P<0.01).
Figure 2.
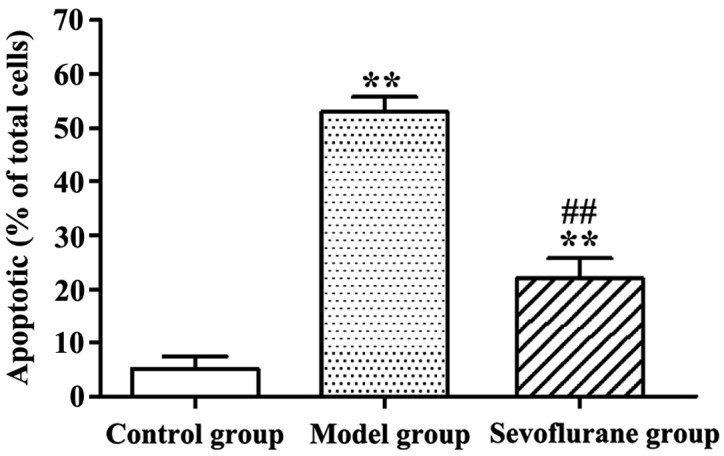
Apoptotic rates of H9c2 cells treated with H2O2 (**P<0.01 and ##P<0.01).
Effect of sevoflurane on intracellular Ca2+ concentration in H2O2-treated H9c2 cells
As shown in Fig. 3, after H9c2 cells were treated with H2O2, the intracellular Ca2+ concentration in the model group was significantly increased (P<0.01), compared to the normal value in the control group. By contrast, when cells were pretreated with sevoflurane prior to H2O2 challenge, the intracellular Ca2+ concentration as seen in the sevoflurane group, was higher than that in the control group, but much lower than that in the model group (P<0.01). This result suggested that sevoflurane pretreatment could balance the increase of the intracellular Ca2+ concentration induced by H2O2.
Figure 3.
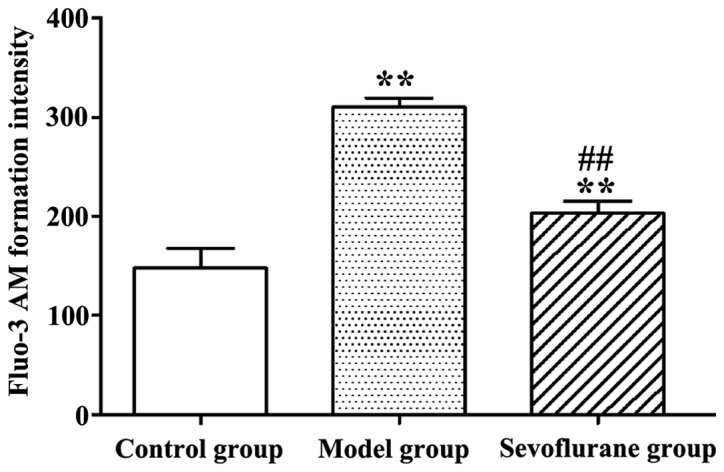
Intracellular Ca2+ concentrations in H2O2-treated H9c2 cells measured by fluo-3 AM fluorescent probe (**P<0.01 and ##P<0.01).
Effect of sevoflurane on the expression levels of GRP78 mRNA in H2O2-treated H9c2 cells
RT-qPCR results show that following H2O2-treatment, the expression level of intracellular GRP78 mRNA in the model group was significantly increased (P<0.01) (Fig. 4). By contrast then the cells were pretreated with sevoflurane prior to the H2O2 challenge, the expression level of GRP78 mRNA as shown in the sevoflurane group was increased, but much lower than that in the model group (P<0.01). This result suggested that pretreatment with sevoflurane could balance the increase of intracellular GRP78 mRNA expression induced by H2O2.
Figure 4.
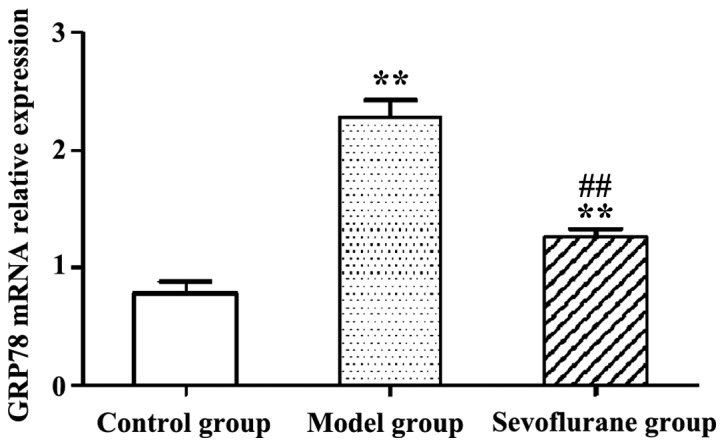
Glucose-regulated protein 78 (GRP78) mRNA relative expression levels in H2O2-treated H9c2 cells measured by qRT-PCR (**P<0.01 and ##P<0.01).
Effect of sevoflurane on the protein expression levels of GRP78, CHOP and caspase-12 in H2O2-induced H9c2 cells
Protein expression levels of GRP78, CHOP and caspase-12 measured by western blot analysis are shown in Fig. 5. The results showed that, after H2O2-treatment the intracellular protein expression levels of GRP78, CHOP and caspase-12 in the model group were all substantially increased (P<0.01). By contrast, when the cells were pretreated with sevoflurane prior to H2O2 challenge, the three protein levels were increased as shown in the sevoflurane group, but much lower than those in the model group (P<0.01).
Figure 5.

Protein expression levels of glucose-regulated protein 78 (GRP78), CHOP and caspase-12 in H2O2-treated H9c2 cells measured by western blot analysis (*P<0.05, **P<0.01 and ##P<0.01).
Discussion
Recent findings have shown that oxidative stress was closely associated with the onset and progression of cardiovascular diseases. In the process of myocardial ischemia-reperfusion, a large amount of ROS were produced, and the antioxidant capacity of tissues and cells became weak as well, leading to oxidative stress response that caused myocardial apoptosis (13,14). Hydrogen peroxide (H2O2) belongs to ROS and is a common molecule. It modulates the activity of transcription factor and related gene expression in the body. Excessive H2O2 can induce apoptosis (15–17). According to the literature, currently known mechanisms underlying myocardial apoptosis mainly include the death receptor, mitochondrial and endoplasmic reticulum pathways. Due to rich endoplasmic reticulum in cardiomyocytes, the endoplasmic reticulum pathway may play a key role in cardiomyocyte apoptosis (3).
GRP78 is an important molecular chaperone in endoplasmic reticulum, which modulates endoplasmic reticulum stress by maintaining correct synthesis and folding of endoplasmic reticulum proteins and stabilizing intracellular calcium concentration (18). It was found that the GRP78 protein expression level in the stress state can be increased several times to cope with modulation of the accumulation and release of calcium in the endoplasmic reticulum, thus maintaining calcium balance in the endoplasmic reticulum (19). Intracellular calcium overload is one of the major causes of myocardial ischemia-reperfusion injury. Prevention of intracellular calcium overload has become an important target for the treatment of myocardial ischemia-reperfusion injury (20). CHOP is induced by endoplasmic reticulum stress and mediates apoptosis. It plays a key role in cell proliferation and apoptosis (21). CHOP is downstream of the apoptotic pathway induced by endoplasmic reticulum stress. Therefore, the expression level of CHOP can reflect the degree of apoptosis induced by endoplasmic reticulum stress (22). In addition, caspase-12 located in the endoplasmic reticulum membrane plays an important role in endoplasmic reticulum stress-induced apoptosis (23).
In this study, rat H9c2 cardiomyocytes were employed to establish a model of oxidative stress injury induced by H2O2. The model was used to investigate the effect of sevoflurane pretreatment on myocardial injury and apoptosis induced by H2O2. Viabilities of H2O2-treated H9c2 cells were determined using MTT assay. The results indicated that viability of H9c2 cells in the model and sevoflurane groups was significantly lower than that in the control group. Moreover, the viability in the sevoflurane-pretreated group was higher than that in the model group. This suggested that sevoflurane had a protective effect on H9c2 cardiomyocytes from H2O2-induced injury. The results of AV-PI double staining assay showed that pretreatment with sevoflurane could reverse H2O2-induced apoptosis of H9c2 cells. After H9c2 cells were treated with H2O2, the intracellular Ca2+ concentration in the model group was significantly increased, compared to the normal value in the control group. By contrast when cells were pretreated with sevoflurane prior to H2O2 challenge, the intracellular Ca2+ concentration as seen in the sevoflurane group was higher than that in the control group, but much lower than that in the model group. This result suggests that sevoflurane pretreatment can balance the increase of the intracellular Ca2+ concentration induced by H2O2. The RT-qPCR results suggest that sevoflurane pretreatment can balance the increase of intracellular GRP78 mRNA expression induced by H2O2. Western blot analysis revealed that H2O2 induced the expression of GRP78 and the downstream proteins CHOP and caspase-12 in the endoplasmic reticulum stress. By contrast, when the cells were pretreated with sevoflurane prior to H2O2 challenge, expression levels of the three proteins were increased as compared with the control group, but much lower than those in the model group.
It was reported that sevoflurane pretreatment reduced myocardial injury caused by myocardial ischemia-reperfusion. This effect of sevoflurane was probably associated with the inhibition of cardiomyocyte apoptosis, but the mechanism was not clear (11,24). Myocardial ischemia-reperfusion induced overexpression of intracellular GRP78 protein, triggering the endoplasmic reticulum stress response and increase the expression of CHOP protein, the phosphorylation of JNK and activation of caspase-12. The three proteins were involved in endoplasmic reticulum stress-induced myocardial apoptosis (25). Thus, the results not only confirmed that sevoflurane reduced the injury and apoptosis of H9c2 cardiomyocytes induced by H2O2, but also confirmed that sevoflurane downregulated the expression of GRP78 mRNA and that of proteins GRP78, CHOP and caspase-12 as well.
In conclusion, results of this study have shown that sevoflurane can reduce H2O2-induced injury and apoptosis of H9c2 cardiomyocytes. The mechanism may be related to inhibition of the expression of GRP78, a regulating protein for endoplasmic reticulum stress, thereby reducing intracellular Ca2+ concentration, and the downregulation of CHOP and caspase-12 expression levels.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present study are available from the corresponding author on reasonable request.
Authors' contributions
ZW analyzed and interpreted the patient data, and was a major contributor in writing the manuscript. RC performed the experiment and participated in the design of the study. KW was a major contributor in designing the methods.
Ethics approval and consent to participate
The study was approved by the Ethics Committee of People's Hospital of Rizhao.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.Delbosc S, Paizanis E, Magous R, Araiz C, Dimo T, Cristol JP, Cros G, Azay J. Involvement of oxidative stress and NADPH oxidase activation in the development of cardiovascular complications in a model of insulin resistance, the fructose-fed rat. Atherosclerosis. 2005;179:43–49. doi: 10.1016/j.atherosclerosis.2004.10.018. [DOI] [PubMed] [Google Scholar]
- 2.Zhang GG, Teng X, Liu Y, Cai Y, Zhou YB, Duan XH, Song JQ, Shi Y, Tang CS, Yin XH, et al. Inhibition of endoplasm reticulum stress by ghrelin protects against ischemia/reperfusion injury in rat heart. Peptides. 2009;30:1109–1116. doi: 10.1016/j.peptides.2009.03.024. [DOI] [PubMed] [Google Scholar]
- 3.Gustafsson AB, Gottlieb RA. Mechanisms of apoptosis in the heart. J Clin Immunol. 2003;23:447–459. doi: 10.1023/B:JOCI.0000010421.56035.60. [DOI] [PubMed] [Google Scholar]
- 4.Sharikabad MN, Østbye KM, Brørs O. Effect of hydrogen peroxide on reoxygenation-induced Ca2+ accumulation in rat cardiomyocytes. Free Radic Biol Med. 2004;37:531–538. doi: 10.1016/j.freeradbiomed.2004.05.002. [DOI] [PubMed] [Google Scholar]
- 5.Benvenga S, Vicchio T, Di Bari F, Vita R, Fallahi P, Ferrari SM, Catania S, Costa C, Antonelli A. Favorable effects of myo-inositol, selenomethionine or their combination on the hydrogen peroxide-induced oxidative stress of peripheral mononuclear cells from patients with Hashimoto's thyroiditis: Preliminary in vitro studies. Eur Rev Med Pharmacol Sci. 2017;21(Suppl 2):89–101. [PubMed] [Google Scholar]
- 6.Schröder M. Endoplasmic reticulum stress responses. Cell Mol Life Sci. 2008;65:862–894. doi: 10.1007/s00018-007-7383-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Glembotski CC. Endoplasmic reticulum stress in the heart. Circ Res. 2007;101:975–984. doi: 10.1161/CIRCRESAHA.107.161273. [DOI] [PubMed] [Google Scholar]
- 8.Bouwman RA, Salic K, Padding FG, Eringa EC, van Beek-Harmsen BJ, Matsuda T, Baba A, Musters RJ, Paulus WJ, de Lange JJ, et al. Cardioprotection via activation of protein kinase C-delta depends on modulation of the reverse mode of the Na+/Ca2+ exchanger. Circulation. 2006;114(Suppl 1):I226–I232. doi: 10.1161/CIRCULATIONAHA.105.000570. [DOI] [PubMed] [Google Scholar]
- 9.Yildirim V, Doganci S, Aydin A, Bolcal C, Demirkilic U, Cosar A. Cardioprotective effects of sevoflurane, isoflurane, and propofol in coronary surgery patients: A randomized controlled study. Heart Surg Forum. 2009;12:E1–E9. doi: 10.1532/HSF98.20081137. [DOI] [PubMed] [Google Scholar]
- 10.Inamura Y, Miyamae M, Sugioka S, Domae N, Kotani J. Sevoflurane postconditioning prevents activation of caspase 3 and 9 through antiapoptotic signaling after myocardial ischemia-reperfusion. J Anesth. 2010;24:215–224. doi: 10.1007/s00540-010-0877-6. [DOI] [PubMed] [Google Scholar]
- 11.Allaouchiche B, Debon R, Goudable J, Chassard D, Duflo F. Oxidative stress status during exposure to propofol, sevoflurane and desflurane. Anesth Analg. 2001;93:981–985. doi: 10.1097/00000539-200110000-00036. [DOI] [PubMed] [Google Scholar]
- 12.Eguchi M, Liu Y, Shin EJ, Sweeney G. Leptin protects H9c2 rat cardiomyocytes from H2O2-induced apoptosis. FEBS J. 2008;275:3136–3144. doi: 10.1111/j.1742-4658.2008.06465.x. [DOI] [PubMed] [Google Scholar]
- 13.Lee Y, Gustafsson AB. Role of apoptosis in cardiovascular disease. Apoptosis. 2009;14:536–548. doi: 10.1007/s10495-008-0302-x. [DOI] [PubMed] [Google Scholar]
- 14.von Harsdorf R, Li PF, Dietz R. Signaling pathways in reactive oxygen species-induced cardiomyocyte apoptosis. Circulation. 1999;99:2934–2941. doi: 10.1161/01.CIR.99.22.2934. [DOI] [PubMed] [Google Scholar]
- 15.Ricci F, Masini F, Fossati B, Frascione P, De Waure C, Capizzi R, Guerriero C. Combination therapy with hydrogen peroxide (4%), salicylic acid (0.5%) and D-panthenol (4%): Efficacy and skin tolerability in common acne vulgaris during sun exposure period. Eur Rev Med Pharmacol Sci. 2016;20:232–236. [PubMed] [Google Scholar]
- 16.Then SM, Sanfeliu C, Top GM, Wan Ngah WZ, Mazlan M. γ-Tocotrienol does not substantially protect DS neurons from hydrogen peroxide-induced oxidative injury. Nutr Metab (Lond) 2012;9:1. doi: 10.1186/1743-7075-9-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Fu J, Huang H, Liu J, Pi R, Chen J, Liu P. Tanshinone IIA protects cardiac myocytes against oxidative stress-triggered damage and apoptosis. Eur J Pharmacol. 2007;568:213–221. doi: 10.1016/j.ejphar.2007.04.031. [DOI] [PubMed] [Google Scholar]
- 18.Shu CW, Sun FC, Cho JH, Lin CC, Liu PF, Chen PY, Chang MD, Fu HW, Lai YK. GRP78 and Raf-1 cooperatively confer resistance to endoplasmic reticulum stress-induced apoptosis. J Cell Physiol. 2008;215:627–635. doi: 10.1002/jcp.21340. [DOI] [PubMed] [Google Scholar]
- 19.Yang YM, Yang Y, Dai WW, Li XM, Ma JQ, Tang LP. Genistein-induced apoptosis is mediated by endoplasmic reticulum stress in cervical cancer cells. Eur Rev Med Pharmacol Sci. 2016;20:3292–3296. [PubMed] [Google Scholar]
- 20.Oyadomari S, Mori M. Roles of CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ. 2004;11:381–389. doi: 10.1038/sj.cdd.4401373. [DOI] [PubMed] [Google Scholar]
- 21.Okada K, Minamino T, Kitakaze M. Role of endoplasmic reticulum stress in hypertrophic and failing hearts. Nihon Yakurigaku Zasshi. 2005;126:385–389. doi: 10.1254/fpj.126.385. (In Japanese) [DOI] [PubMed] [Google Scholar]
- 22.Szegezdi E, Logue SE, Gorman AM, Samali A. Mediators of endoplasmic reticulum stress-induced apoptosis. EMBO Rep. 2006;7:880–885. doi: 10.1038/sj.embor.7400779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Nakagawa T, Zhu H, Morishima N, Li E, Xu J, Yankner BA, Yuan J. Caspase-12 mediates endoplasmic-reticulum-specific apoptosis and cytotoxicity by amyloid-beta. Nature. 2000;403:98–103. doi: 10.1038/47513. [DOI] [PubMed] [Google Scholar]
- 24.Lai E, Teodoro T, Volchuk A. Endoplasmic reticulum stress: Signaling the unfolded protein response. Physiology (Bethesda) 2007;22:193–201. doi: 10.1152/physiol.00050.2006. [DOI] [PubMed] [Google Scholar]
- 25.Yang Y, Zhang Y, Liu X, Zuo J, Wang K, Liu W, Ge J. Exogenous taurine attenuates mitochondrial oxidative stress and endoplasmic reticulum stress in rat cardiomyocytes. Acta Biochim Biophys Sin (Shanghai) 2013;45:359–367. doi: 10.1093/abbs/gmt034. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the present study are available from the corresponding author on reasonable request.