

Abstract
The ability to solubilize fixed inorganic phosphorus (P) for plant growth is important for increasing crop yield. More P can be released by inoculating soil with inorganic-phosphate-solubilizing bacteria (iPSBs). We used 96-well microplates instead of traditional 200-mm petri dishes to rapidly screen iPSB strains for their solubilizing ability. We simultaneously obtained 76 iPSB isolates from 576 wells containing two agricultural soils. This method conveniently identified positive iPSB strains and effectively prevented fungal cross-contamination. Maximum-likelihood phylogenetic trees of the isolated strains showed that Bacillus megaterium was the most dominant iPSB, and strains Y99, Y95, Y924 and Y1412 were selected as representatives for the analysis of P solubilization. Succinic acid was the main organic acid of B. megaterium for releasing P. It was strongly correlated with the increase in soluble P concentration during 168 h of incubation of these four strains. pH was negatively exponentially correlated with the amount of soluble P in the medium, and the amount of succinic acid was strongly linearly correlated with the amount of P released (P < 0.001), suggesting that organic acid may mobilize microbial P. Our study provides an efficient and effective method for identifying and analyzing the growth of iPSB strains able to solubilize inorganic P and gives a better understanding of the mechanism of P solubilization.
Keywords: Phosphorus, Inorganic phosphate solubilizing bacteria, Isolation, Characterization
Introduction
Phosphorus (P), a non-renewable macronutrient, plays an essential role in plants (Elser et al. 2007). Inorganic P is mined to produce chemical P fertilizers that are extensively applied to cropland (Elser and Bennett 2011; Penuelas et al. 2013). The majority of soluble inorganic P, however, is rapidly immobilized by soil fixation and becomes unavailable for plant uptake, leading to low P-use efficiency and potentially excess P (Kochian 2012). Soil P must thus be managed to minimize its loss and increase its use efficiency.
Microorganisms are actively involved in many biogeochemical processes, including the mineralization, solubilization and transformation of soil P (van der Heijden et al. 2008). Inorganic-phosphate-solubilizing bacteria (iPSBs) are particularly effective in releasing P from pools of inorganic P. iPSBs can also prevent the liberated P from being fixed again (Richardson et al. 2009; Richardson and Simpson 2011). Screening highly efficient iPSBs as soil inoculum is a useful method for improving plant growth and yield (Richardson et al. 2009). Many methods for screening iPSBs have been reported (Chen et al. 2006; Chung et al. 2005; Mehta and Nautiyal 2001; Nautiyal 1999), but isolating iPSBs on separate petri dishes can be time-consuming and labor-intensive.
The mechanism of inorganic-P microbial mobilization is generally associated with extrusion of low-molecular-weight organic acids (Goldstein 1995), which can competitively chelate the cations bound to P via hydroxyl and carboxyl groups and convert them into soluble forms (Jones and Oburger 2011; Richardson and Simpson 2011). The solubilization of inorganic P, however, is complex and depends on numerous factors such as soil properties, plant nutritional requirements and physiological and growth conditions. Studies of the factors affecting solubilization are thus still needed.
We developed a rapid method for screening iPSBs and evaluating their effectiveness at solubilizing inorganic P, with an emphasis on potential highly efficient iPSBs for agricultural use. The isolated iPSB strains were identified and characterized. The types of organic acids secreted by the iPSB strains with high P-solubilizing abilities and the relationships between the organic acids, pH and P solubilization were analyzed.
Materials and methods
Soil sampling and characterization
Soil samples were collected from agricultural fields near Hailun in Heilongjiang Province (47′26″N, 126′38″E) and Yingtan in Jiangxi Province (28′14″N 116′54″E), China (Table 1). Approximately 500 g of surface soil (0–15 cm) was collected after crop harvests in June 2014. The soils were then air-dried, sieved (0.2 mm) and stored at 4 °C until analysis.
Table 1.
Basic information and soil properties of the two soil samples
| Sample | Location | pH | Total P (g kg−1) | Olsen P (mg kg−1) | Organic P (mg kg−1) | Inorganic P (mg kg−1) |
|---|---|---|---|---|---|---|
| Hailun | 47′26″N, 126′38″E | 5.70 ± 0.08 | 779.35 ± 44.33 | 58.80 ± 2.64 | 351.15 ± 6.37 | 428.21 ± 37.96 |
| Yingtan | 28′14″N 116′54″E | 5.01 ± 0.03 | 522.60 ± 7.54 | 23.56 ± 1.53 | 370.57 ± 5.50 | 152.04 ± 2.05 |
Soil pH was measured using a 1:2.5 (w/v) suspension of dry soil: water and a XL60 pH meter (Fisher Scientific, USA) (Shen et al. 2008). The amounts of total P and available P (Olsen P) were determined using the molybdate-blue method (Murphy and Riley 1962) and sodium bicarbonate extraction (Olsen et al. 1954) after acid digestion (Parkinson and Allen 1975), respectively. The inorganic-P concentration was measured by shaking 0.2 g of soil in 20 mL of 1 M HCl at 200 rpm for 30 min, followed by centrifugation at 4200g for 10 min. The amount of inorganic P in the supernatant was then measured by the molybdate-blue method. The amount of organic P was calculated by subtracting the inorganic-P concentration from the amount of total P.
Rapid screening of iPSB strains
The iPSBs were rapidly screened using 96-well microplates (Fig. 1). A modified Pikovskaya medium (PVK) without yeast extract (Nautiyal 1999) was used as the culture medium and was added to each well in advance. The PVK was supplemented with 10 μM bromocresol purple as an indicator. For each microplate, 1 g of soil was homogenized with 100 mL of sterilized water, and this suspension was then serially diluted (10–105). One microliter of diluted soil suspension was added to each well and then incubated at 30 °C for at least 72 h. An uninoculated well served as a control. Wells with no bacterial growth were considered negative. Wells with bacterial growth but no obvious color change were classified as containing uncertain strains, and wells with obvious bacterial growth and a yellow color were classified as positive. Three replicates of each dilution were tested. The effectiveness of this method was verified by streaking both uncertain and positive strains on plates containing solid modified PVK.
Fig. 1.
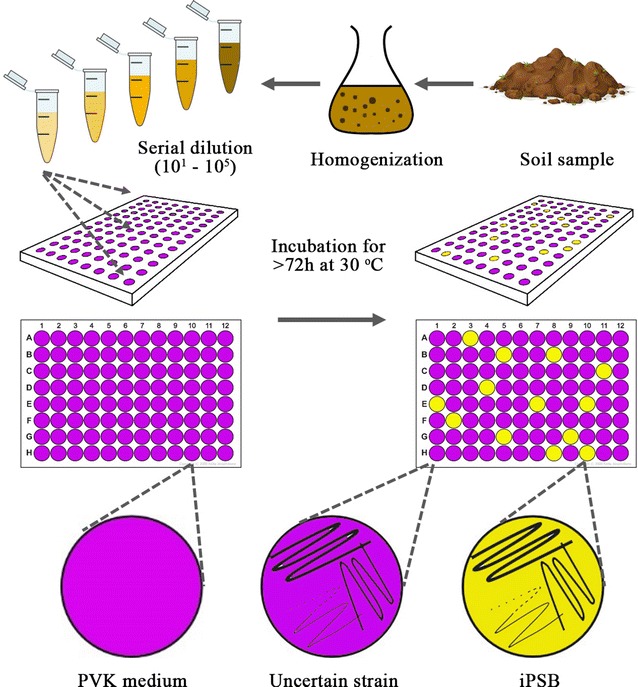
Flowchart of the 96-well iPSB screening method. The sampled soil was homogenized in autoclaved water and serially diluted for incubation at 30 °C for > 72 h in 96-well microplates containing PVK with bromocresol purple as an indicator. Wells with no color change or strain growth, or with strain growth but no color change, were classified as containing unknown strains without P-solubilizing ability. Wells that produced a yellow color were classified as containing iPSB strains
Biochemical characterization of the iPSB strains
The pH and soluble-P concentration of the medium after incubation were used as indices for iPSB screening. The indices were measured by incubating all strains in 50 mL of liquid modified PVK (without agar and indicator) at 30 °C for 72 h. The supernatants obtained after centrifugation (4200g for 10 min) were used to measure pH with a XL60 pH meter (Fisher Scientific, USA) and phosphate concentration using the molybdate-blue method (Murphy and Riley 1962).
Phylogenetic identification with 16S rRNA sequencing
All positive strains were incubated in liquid modified PVK (without indicator) at 30 °C for 24 h. The full lengths (~ 1500 bp) of 16S rRNA genes were obtained by PCR using universal bacterial primers (24F: AGAGTTTGATCCTGGCTCAG and 1492R: TACGGYTACCTTGTTACGACTT) (Farris and Olson 2007). Each 50-μL PCR reaction contained 1 μL of Premix Ex Taq Hot Start Version (TAKARA, Dalian, China), 0.2 μM each primer and 1 μL of bacterial culture suspension as DNA template. The amplification protocol was an initial denaturation at 95 °C for 4 min, 30 cycles of 95 °C for 30 s, 58 °C for 90 s and 72 °C for 30 s and a final 5-min extension at 72 °C. The amplicons were purified with a Universal DNA Purification Kit (TIANGEN, Beijing, China) and submitted for sequencing (Invitrogen, Shanghai, China). The sequences were aligned with those from bacterial lineages in GenBank at the National Center for Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov/) with the BLAST program. The 16S rRNA sequences of all strains were uploaded to the NCBI Sequence Read Archive with Accession Numbers KU647195-KU647270 (Table 2).
Table 2.
Accession numbers, medium pH and soluble-P concentrations of the iPSB strains after incubation for 72 h, and the closest reference strains
| Strain | Accession number | pH of medium | Soluble-P concentration (μg mL−1) | Closest reference strain | ||
|---|---|---|---|---|---|---|
| Affiliation | Accession number | Similarity (%) | ||||
| Bacillus megaterium 01-A3 | KU647195 | 4.80 | 85.57 | Bacillus sp. BS3(2015) | KR063183 | 99 |
| Bacillus megaterium 02-A7 | KU647196 | 4.59 | 89.08 | Bacillus sp. KU6 | JF895481 | 99 |
| Pseudomonas frederiksbergensis 03-D2 | KU647197 | 5.21 | 64.28 | Pseudomonas sp. WS06 | JN210901 | 99 |
| Rhodococcus opacus 04-OD7 | KU647198 | 5.17 | 28.06 | Rhodococcus opacus DSM 43205 | LN827919 | 99 |
| Arthrobacter phenanthrenivorans 05-OD11 | KU647199 | 5.89 | 12.24 | Arthrobacter phenanthrenivorans L43 | LN890039 | 99 |
| Arthrobacter defluvii 06-OD12 | KU647200 | 8.34 | 59.11 | Uncultured bacterium D1-57 | KC554872 | 99 |
| Arthrobacter chlorophenolicus 07-OD13 | KU647201 | 5.58 | 20.84 | Arthrobacter sp. M29 | KF430812 | 99 |
| Arthrobacter oxydans 08-OY2 | KU647202 | 6.64 | 3.85 | Uncultured bacterium D1-57 | KC554872 | 99 |
| Arthrobacter sp. 09-OY5 | KU647203 | 5.11 | 43.00 | Arthrobacter sp. WS03 | JN210899 | 99 |
| Bacillus megaterium 10-Y11 | KU647204 | 4.77 | 106.46 | Bacillus megaterium HNS88 | KF933685 | 99 |
| Pseudomonas frederiksbergensis 11-D3 | KU647205 | 5.25 | 81.76 | Pseudomonas sp. B3039 | KC236870 | 99 |
| Massilia putida 12-OD1 | KU647206 | 4.63 | 97.29 | Uncultured bacterium clone HF31 | KR188907 | 99 |
| Duganella sp. 13-D4 | KU647207 | 5.69 | 10.78 | Duganella sp. ZLP-XI | KF896136 | 99 |
| Bacillus megaterium 14-Y2 | KU647208 | 4.75 | 101.58 | Bacillus megaterium Y20 | JQ798391 | 99 |
| Pseudoduganella sp. 15-Y6 | KU647209 | 5.29 | 49.64 | Pseudoduganella sp. NI28 | KM087999 | 99 |
| Bacillus megaterium 16-Y9 | KU647210 | 4.66 | 80.20 | Bacillus sp. RBB1 | GU979225 | 99 |
| Bacillus megaterium 17-Y5 | KU647211 | 4.85 | 80.39 | Uncultured Bacillus sp. clone T7F50d237 | JN187411 | 99 |
| Variovorax paradoxus 19-D4 | KU647212 | 5.42 | 55.69 | Variovorax paradoxus EPS | NR_074646 | 99 |
| Rhizobium leguminosarum 20-OD2 | KU647213 | 5.69 | 10.78 | Rhizobium sp. SG6 | LC042447 | 99 |
| Rhodanobacter sp. 21-Y7 | KU647214 | 7.72 | 2.58 | Rhodanobacter sp. GR14-4 | FJ821729 | 99 |
| Bacillus megaterium 22-A1 | KU647215 | 5.00 | 100.51 | Bacillus sp. B2(2010b) | HM104462 | 99 |
| Pseudomonas frederiksbergensis 23-D2 | KU647216 | 5.20 | 63.41 | Pseudomonas sp. WS06 | JN210901 | 99 |
| Bacillus megaterium 24-Y916 | KU647217 | 4.79 | 109.39 | Bacillus sp. BDH23 | KF933618 | 99 |
| Rhodanobacter sp. 25-Y8 | KU647218 | 4.82 | 18.20 | Rhodanobacter sp. GR14-4 | FJ821729 | 99 |
| Bacillus megaterium 26-Y91 | KU647219 | 4.63 | 46.61 | Bacillus megaterium HNS79 | KF933676 | 99 |
| Bacillus megaterium 27-Y93 | KU647220 | 4.37 | 117.30 | Bacillus sp. NyZ44 | HQ231223 | 99 |
| Bacillus megaterium 28-Y911 | KU647221 | 4.54 | 126.48 | Bacillus megaterium Bacteria I | KT427436 | 99 |
| Bacillus megaterium 29-Y924 | KU647222 | 4.55 | 136.83 | Bacillus sp. BDH4 | KF933626 | 99 |
| Bacillus megaterium 30-Y1411 | KU647223 | 4.48 | 134.39 | Bacillus sp. WXGRY7 | KJ184905 | 99 |
| Bacillus megaterium 31-Y142 | KU647224 | 4.71 | 97.29 | Bacillus megaterium Bacteria I | KT427436 | 99 |
| Arthrobacter sp. 32-OD9 | KU647225 | 5.31 | 43.19 | Arthrobacter sp. WS20 | JN899573 | 99 |
| Streptomyces tumescens 33-X1 | KU647226 | 8.02 | 2.77 | Streptomyces tumescens OTP-4-2 | AF346485 | 99 |
| Streptomyces prasinopilosus 34-Y1 | KU647227 | 7.75 | 3.07 | Streptomyces sp. GS15 | JX679244 | 99 |
| Streptomyces rishiriensis 35-Y3 | KU647228 | 5.76 | 44.37 | Streptomyces sp. YRA147 | JX430828 | 99 |
| Kurthia zopfii 36-Y7 | KU647229 | 4.52 | 81.57 | Bacillus thermophilus SgZ-10 | NR_109677 | 97 |
| Rhodanobacter sp. 37-Y8 | KU647230 | 4.88 | 32.55 | Rhodanobacter sp. GR14-4 | FJ821729 | 99 |
| Bacillus megaterium 38-Y92 | KU647231 | 4.51 | 91.04 | Bacillus megaterium HNS88 | KF933685 | 99 |
| Bacillus megaterium 39-Y94 | KU647232 | 4.43 | 91.62 | Bacillus megaterium HNS79 | KF933676 | 99 |
| Bacillus megaterium 40-Y95 | KU647233 | 4.44 | 134.49 | Bacillus sp. S10 | HE662645 | 99 |
| Bacillus megaterium 41-Y99 | KU647234 | 4.41 | 159.48 | Bacillus megaterium BS17 | KR063197 | 99 |
| Bacillus megaterium 42-Y910 | KU647235 | 4.58 | 75.22 | Bacillus megaterium B2 | KT307979 | 99 |
| Bacillus megaterium 43-Y912 | KU647236 | 4.58 | 72.39 | Bacillus sp. BDH23 | KF933618 | 99 |
| Bacillus megaterium 44-Y913 | KU647237 | 4.50 | 46.51 | Bacillus megaterium HNS88 | KF933685 | 99 |
| Bacillus megaterium 45-Y914 | KU647238 | 4.65 | 94.26 | Bacillus megaterium ML482 | KC692173 | 99 |
| Bacillus megaterium 46-Y923 | KU647239 | 4.62 | 81.57 | Bacillus sp. BDH4 | KF933626 | 99 |
| Bacillus megaterium 47-Y141 | KU647240 | 4.62 | 70.73 | Bacillus sp. BSp-2 | KF835394 | 99 |
| Rhizobium sp. 48-Y930 | KU647241 | 7.86 | 3.75 | Rhizobium sp. CC-SKC2 | HQ113369 | 99 |
| Bacillus megaterium 49-Y1412 | KU647242 | 4.60 | 138.68 | Bacillus megaterium HNS88 | KF933685 | 99 |
| Rhizobium sp. 50-Y1414 | KU647243 | 8.02 | 5.60 | Rhizobium sp. CC-SKC2 | HQ113369 | 99 |
| Burkholderia cepacia 51-Y1415 | KU647244 | 5.00 | 2.03 | Burkholderia sp. xin-1 | KF059269 | 99 |
| Arthrobacter defluvii 52-OD12 | KU647245 | 4.59 | 76.10 | Uncultured bacterium D1-57 | KC554872 | 99 |
| Bacillus acidiceler 53-Q11 | KU647246 | 4.39 | 127.07 | Bacillus sp. S21001 | D84560 | 99 |
| Streptomyces prasinopilosus 54-Y1 | KU647247 | 5.29 | 49.64 | Streptomyces sp. GS15 | JX679244 | 99 |
| Pseudomonas frederiksbergensis 55-D3 | KU647248 | 4.96 | 35.87 | Pseudomonas sp. B3039 | KC236870 | 99 |
| Burkholderia phytofirmans 56-OY3 | KU647249 | 8.20 | 3.85 | Burkholderia sp. C2-14 | JF900054 | 99 |
| Variovorax paradoxus 57-Y925 | KU647250 | 5.30 | 10.88 | Variovorax sp. LZA10 | GQ861460 | 99 |
| Telluria mixta 58-Y97 | KU647251 | 4.62 | 106.85 | Uncultured bacterium SuR5 | AB608684 | 99 |
| Sphingomonas koreensis 59-Y96 | KU647252 | 7.31 | 2.77 | Uncultured Sphingomonas sp. Plot4-G09 | EU449628 | 99 |
| Streptomyces flaveolus 60-OD3 | KU647253 | 7.95 | 2.19 | Streptomyces flaveolus NRRL B-1334 | NR_116094 | 99 |
| Rhodanobacter sp. 61-Y8 | KU647254 | 4.49 | 62.23 | Rhodanobacter sp. GR14-4 | FJ821729 | 99 |
| Streptomyces sp. 62-Y930 | KU647255 | 6.50 | 3.46 | Streptomyces sp. N4-145 | EF063495 | 99 |
| Rhodococcus cercidiphylli 63-OD5 | KU647256 | 6.77 | 3.07 | Rhodococcus sp. AB73 | KC019201 | 98 |
| Bacillus megaterium 64-Y98 | KU647257 | 4.53 | 107.44 | Bacillus sp. BDH23 | KF933618 | 100 |
| Bacillus megaterium 65-Y918 | KU647258 | 4.71 | 69.75 | Bacillus megaterium B2 | KT307979 | 99 |
| Bacillus megaterium 66-Y143 | KU647259 | 4.55 | 82.84 | Bacillus megaterium Bacteria I | KT427436 | 99 |
| Rhodococcus sp. 67-OD10 | KU647260 | 5.45 | 52.67 | Uncultured bacterium clone Md-133 | KT905708 | 99 |
| Arthrobacter oxydans 68-OY1 | KU647261 | 6.15 | 16.44 | Arthrobacter oxydans BGSLP35 | KP192013 | 99 |
| Pseudomonas sp. 69-Y94 | KU647262 | 4.87 | 71.51 | Pseudomonas sp. B3042 | KC236872 | 99 |
| Bacillus megaterium 70-Y917 | KU647263 | 4.43 | 76.10 | Bacillus sp. S10 | HE662645 | 99 |
| Pseudomonas sp. 71-Y928 | KU647264 | 5.41 | 37.82 | Pseudomonas sp. B3042 | KC236872 | 99 |
| Bacillus megaterium 72-Y13 | KU647265 | 4.61 | 112.03 | Bacillus megaterium HNS79 | KF933676 | 99 |
| Bacillus megaterium 73-Y142 | KU647266 | 4.77 | 106.46 | Bacillus megaterium HNS88 | KF933685 | 99 |
| Streptomyces sp. 74-Y144 | KU647267 | 5.00 | 22.50 | Streptomyces sp. N4-145 | EF063495 | 99 |
| Leifsonia shinshuensis 75-Y145 | KU647268 | 4.54 | 27.08 | Leifsonia shinshuensis DB 102 | NR_043663 | 99 |
| Bacillus megaterium 76-Y149 | KU647269 | 4.78 | 59.70 | Bacillus sp. BSp-2 | KF835394 | 99 |
| Streptomyces sp. 77-Y1410 | KU647270 | 5.25 | 34.41 | Streptomyces sp. N4-145 | EF063495 | 99 |
Growth analysis of representative iPSB strains
The four iPSB strains with the highest P-solubilizing capacity were used as representatives for further analysis (Bacillus megaterium Y99 was stored in China Center for Type Culture Collection, CCTCC, No. CCTCC AB 2017149). The representative strains were inoculated into 100 mL of liquid modified PVK and incubated at 30 °C for 168 h. The solubilized-P concentration and organic acid production were measured at 12, 24, 48, 96 and 168 h. The solubilized-P concentration was determined by the molybdate-blue method as described above. The P solubilizing percentage (%) was calculated as follows,
Pfree indicated the free solubilizing phosphate concentration (µg mL−1) in the supernatant of liquid medium, Ptotal indicated the total P concentration (µg mL−1) in the liquid medium. The amounts of the organic acids, including lactic, acetic, propionic, gluconic, succinic, oxalic and citric acids, were determined using ion chromatography (ICS-3000, Dionex, USA) as previously described (Hu et al. 2009). The reference standards of the corresponding sodium salts of these organic acids were chromatographically pure (Sigma-Aldrich, Shanghai, China).
Statistical analyses
Figures were generated using Microsoft Office 365. The sequences were aligned and the phylogenetic trees were constructed and annotated using Clustal X 2.0 (Larkin et al. 2007), MEGA 6.0 (Tamura et al. 2013) and iTOL v3 (Letunic and Bork 2016), respectively. The correlations and variance analyses (ANOVAs) used IBM SPSS Statistics 21.
Results
Rapid screening of the iPSB strains
Six 96-well microplates (576 wells in total) with modified PVK were used for screening the two soil samples for iPSBs (Fig. 2a). We isolated 39 and 35 iPSB strains from the Hailun and Yingtan soils, respectively, and two uncertain bacterial strains were obtained from the Hailun soil. The uncertain strains were transferred to 200-mm PVK medium plates (Fig. 2b), where they survived and produced slight color changes and clearance zones. We thus classified these two strains as iPSB strains. The other iPSB strains produced obvious color changes after inoculation (Fig. 2c). Hence, a total of 76 iPSB strains were obtained with an average screening efficiency of 13.19 ± 1.47%. Only one bacterial strain survived in each microplate well, which was confirmed by streaking onto 200-mm PVK medium plates. Fungal contamination was well confined to single wells (Fig. 2a, purple circle).
Fig. 2.
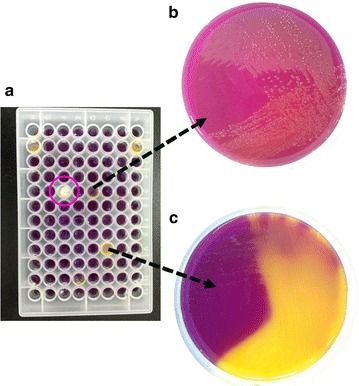
Photographs of a a 96-well iPSB screening microplate containing PVK media, b an uncertain strain and c an iPSB strain cultivated on a PVK medium plate with color change. The purple circle indicates that fungal growth was confined to a single well
Phylogenetic identification of iPSB strains
The maximum-likelihood phylogenetic trees based on 16S rRNA sequences from the 76 iPSB strains are presented in Fig. 3. Five phyla were identified: Actinobacteria, Firmicutes and α-, β- and γ-Proteobacteria. Bacillus megaterium was the most dominant species (32 isolates). The other 44 isolates were eight Arthrobacter sp., seven Streptomyces sp., seven Pseudomonas sp., four Rhodanobacter sp., three Rhizobium sp., three Rhodococcus sp., two Burkholderia sp., two Variovorax paradoxus and single isolate of B. acidceler, Duganella sp., Kurthia zopfii, Leifsonia shinshuensis, Massilia sp., Pseudoduganella sp., Sphingomonas koreensis and Tellura mixta. The closest phylogenic reference strains with their similarities are listed in Table 2. Interestingly, K. zopfii 36-Y7 was only 97% similar to the closest strain based on the reference sequence from the GenBank database.
Fig. 3.
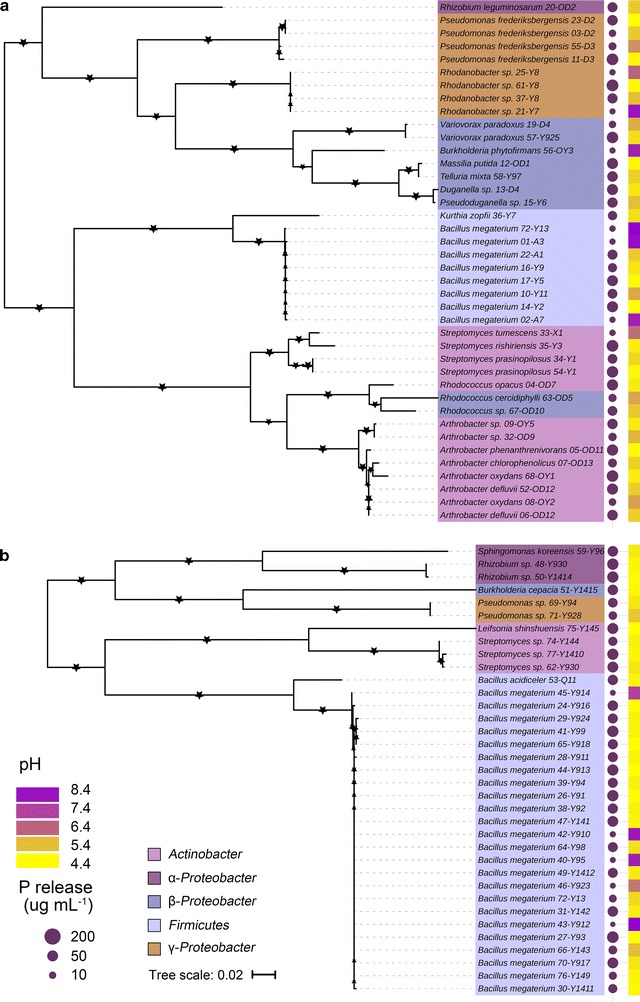
Phylogenic trees of 76 iPSB strains from a Hailun and b Yingtan soil based on their 16S rRNA sequences. Maximum likelihood was used to construct the trees with bootstrapping (1000 replicates). Bootstrap percentages > 50% are marked with stars. The scale bar indicates 0.02 accumulated changes per nucleotide position
Biochemical characterization of the iPSB strains
Details of the medium pH and soluble-P concentration after 72 h incubation are shown in Fig. 3 and Table 2. The pH decreased as the soluble-P concentration increased in the liquid medium. The pH for B. megaterium 27-Y93 decreased from an initial 7.0 to 4.37 after 72 h. Interestingly, the pH for Arthrobacter defluvii 06-OD12, Streptomyces tumescens 33-X1, Rhizobium sp. 48-Y930, Rhizobium sp. 50-Y1414, Burkholderia phytofirmans 56-OY3 and Streptomyces flaveolus 60-OD3 increased to 8.34, 8.02, 7.86, 8.02, 8.20 and 7.95, respectively. The soluble-P concentration ranged from 2.03 to 159.48 μg mL−1. Neither soluble P nor a decrease in pH was detected in the control treatment. The soluble-P concentration was highest for B. megaterium Y99 (159.48 μg mL−1), followed by B. megaterium Y1412 (138.68 μg mL−1), B. megaterium Y924 (136.83 μg mL−1) and B. megaterium Y95 (134.49 μg mL−1), and their medium pH decreased to about 4.5. These four strains were further analyzed as representative iPSB strains (Fig. 4a–d). Medium pH was significantly negatively correlated with soluble-P concentration (P < 000.1) (Fig. 5a).
Fig. 4.
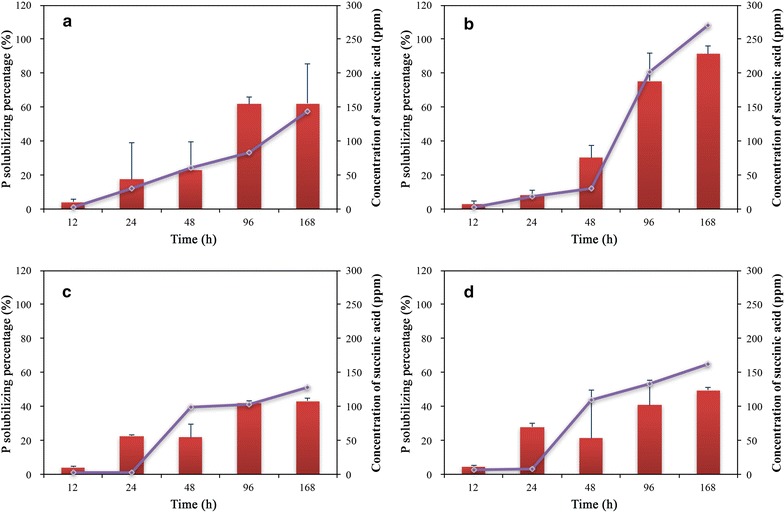
The P-solubilizing percentages (red bars) and succinic acid production (purple lines) for the four representative iPSB strains over 168 h of growth. a B. megaterium Y1412, b B. megaterium Y99, c B. megaterium Y95 and d B. megaterium Y924. Each value represents the mean of three replicates ± standard deviation
Fig. 5.
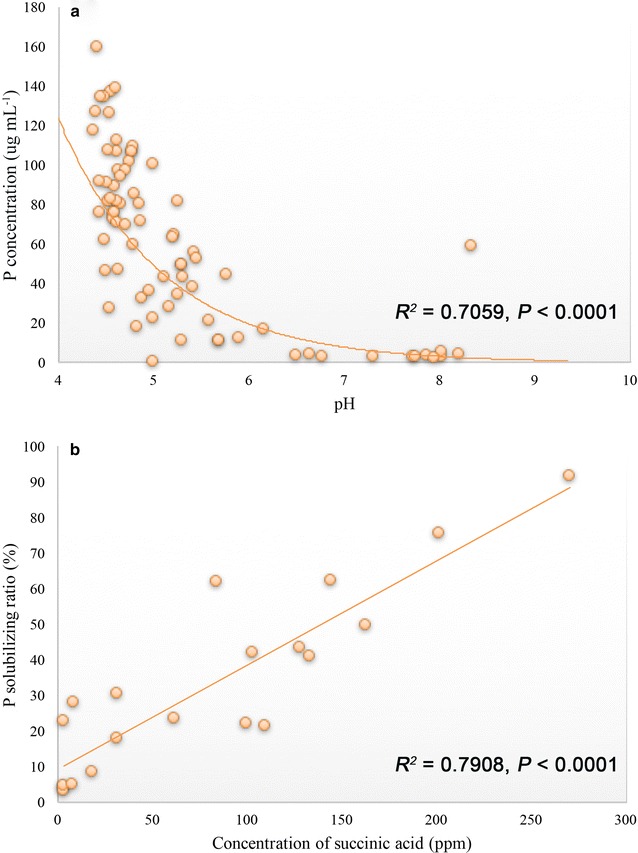
Regression analysis of a the correlation between pH and P release for all iPSB strains (regression function: y = 4926.1e−0.922x) and b the correlation between the concentration of succinic acid and the P-solubilizing percentage for the four representative iPSB strains (regression function: y = 0.2925x + 9.3694)
Growth analysis of the four representative iPSB strains
The four representative iPSB strains were incubated with PVK at 30 °C for 168 h. The organic acids were identified by ion chromatography (Table 3). Only three of the seven organic acids (succinic, oxalic and citric) were detected. Among these, the concentration of succinic acid peaked at 209 ppm, which was significantly higher than the concentrations of the other organic acids (P < 0.05).
Table 3.
Amounts of organic acids secreted by the four reference iPSB strains after incubation for 168 h
| iPSB strain | Organic acid (ppm) | ||||||
|---|---|---|---|---|---|---|---|
| Lactic | Acetic | Propionic | Gluconic | Succinic | Oxalic | Citric | |
| B. megaterium Y95 | ND | ND | ND | ND | 202.30 ± 105.27 | 6.66 ± 1.22 | 2.33 ± 0.89 |
| B. megaterium Y99 | ND | ND | ND | ND | 196.60 ± 99.77 | 6.53 ± 0.06 | 0.24 ± 0.02 |
| B. megaterium Y924 | ND | ND | ND | ND | 208.88 ± 91.90 | 5.21 ± 2.10 | 4.12 ± 0.51 |
| B. megaterium Y1412 | ND | ND | ND | ND | 142.21 ± 27.42 | 6.23 ± 0.83 | 2.37 ± 1.67 |
ND not detected
We further analyzed the relationship between the soluble-P and succinic acid concentrations for these four strains (Fig. 4). Both concentrations increased with bacterial growth. A linear regression analysis indicated that the amount of soluble P was significantly positively correlated with the release of succinic acid (P < 0.001) (Fig. 5b).
Discussion
Current agricultural practices tend to use sustainable technology, including the use of biofertilizers with multiple functions, to achieve high crop yields, which requires high soluble-P concentrations. iPSBs thus play an essential role in releasing P from soil-fixation systems and in preserving enough free phosphate in the rhizosphere for plant uptake and growth. iPSB inoculation can promote plant growth (Kaur and Reddy 2015; Yu et al. 2012), so identifying highly efficient iPSB strains thus becomes important. Our rapid screening using 96-well microplates provided an efficient method for simultaneously isolating numerous iPSB strains. The method isolated 76 positive iPSB strains within 72 h from two soils with three replicates. Each well has limited space, so only a single bacterial strain may survive if the sample is sufficiently diluted. Each well in our study contained a sole iPSB strain with a 105 dilution of soil sample from two remote sites with different chemical properties, which was verified by incubation on 200-mm medium plates (Fig. 2), suggesting that further purification by a second or third colony transfer may not be needed. Positive iPSB strains were also easy to observe and count using bromocresol purple as an indicator.
Wells without bacterial growth and a color change were deemed to be negative, and yellow wells (definitely with bacterial growth) were deemed to be positive. Only the uncertain or ambiguous wells may need further confirmation. Two slightly yellow wells in our study were ultimately demonstrated to be positive iPSB strains with P-solubilizing ability. Our method can also effectively prevent the spread of fungal contamination. Most screening media contain glucose or other carbohydrates as carbon sources, so heterologous fungal spores can easily grow or be initially introduced in the diluted sample. Once a single fungal colony appears, it can promptly spread throughout the medium plate and interfere with screening. The first screening of iPSBs from soil samples also may need more than 72 h of incubation, so fungal survival is likely. Fungal spores in our method (Fig. 2a, purple circle), however, were confined to only one well instead of spreading and affecting surrounding wells. The rapid screening with 96-well microplates was demonstrated to be an efficient and effective way to isolate iPSB strains.
The identification of iPSB strains based on 16S rRNA sequences illustrated the phylogenetic structure of the potential iPSB communities in the two soils. B. megaterium contributed > 30% to the total iPSB population, which was also most frequently found in the Yingtan soil with an acidic pH (Fig. 3). Bacillus is abundant in various types of soil and has a strong P-solubilizing ability in cropland (Chen et al. 2006; Karagöz et al. 2012; Oliveira et al. 2009; Xuan et al. 2011). The next two most common genera, Arthrobacter and Streptomyces, which belong to Actinobacteria, are common in alkaline soils where considerable P is released (Aislabie et al. 2006; Gopalakrishnan et al. 2011; Xiong et al. 2012). Pseudomonas is a known iPSB genus and has been well studied due to genetic evidence of inorganic-P solubilization (Babu-Khan et al. 1995; Kwak et al. 2015; Umezawa et al. 2015). Other strains rarely act as iPSBs but can promote plant growth in various types of soils (Richardson et al. 2009; Rodríguez and Fraga 1999; Zhao et al. 2014).
In our study, B. megaterium was observed to solubilize more P than the other genera, including Streptomyces, Arthrobacter and Pseudomonas. The presence of B. megaterium in the rhizosphere could thus benefit plant P assimilation. B. megaterium isolates Y95, Y99, Y924 and Y1412 release > 130 μg mL−1 soluble P when cultured in liquid PVK. Lactic and propionic acids were reported to be the two most common organic acids secreted by B. megaterium in a previous study (Chen et al. 2006), but we did not observe any ion-chromatographic peaks in the corresponding retention times of these two organic acids (Table 3). Gluconic acid has also been considered an important organic acid for P release (Rodríguez et al. 2006), but we did not detect this acid for any of the four representative iPSB strains. Of the three secreted organic acids, citric and oxalic acids are efficient P-solubilizing acids (Bolan et al. 1994), but their amounts released by these four strains were relatively low and may not be sufficient to liberate much P. The dominant succinic acid was not a common organic acid for P release, but the amounts secreted were strongly correlated with P release for the four representative strains over time (Fig. 4). Succinic acid concentration was strongly, positively and linearly correlated with P release (Fig. 5a) (R2 = 0.7908, P < 0.001), suggesting that succinic acid was the main solubilizing acid secreted by B. megaterium.
Environmental pH and organic acid secretion were two common factors accounting for the mobilization of available P. pH is usually negatively correlated with the release of P (Chen et al. 2006; Rodríguez and Fraga 1999). The pH of the medium for some of the iPSBs in our study, however, was alkaline (Table 2), and the iPSBs likely had the ability to release P because they survived well at a high cell density (data not shown). The regression analysis of medium pH and P release (Fig. 5a) indicated a negative but not linear correlation. The pH may only sharply decrease with an increase in P release at acidic pHs, suggesting that soil pH may not be a universal factor to account for microbial P solubilization by iPSB stains.
We demonstrated the efficiency and effectiveness of this rapid iPSB screening method using 96-well microplates. This method is rapid and easy to manipulate and observe and can prevent the spread of fungal growth. B. megaterium was the main iPSB strain but released mostly succinic acid rather than other common organic acids for P solubilization. Our study may be useful for mechanistic study of microbial inorganic-P solubilization.
Authors’ contributions
Y-GZ and X-RY conceived the project. D-PZ and Q-FB conducted the soil samplings. B-XZ and MI conceived the rapid screening method. H-ZL and G-WZ conducted the bacterial isolation and identification. B-XZ, MI and KD conducted the experiment. B-XZ, Y-GZ and JP analyzed the results and wrote the manuscript. All authors read and approved the final manuscript.
Acknowledgements
The authors wish to acknowledge Dr. Jian-Qiang Su and Ms. Yu Yan (Institute of Urban Environment, Chinese Academy of Sciences) for their helpful suggestions and assistance with figure production.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The 16S rRNA sequences were uploaded to the NCBI Sequence Read Archive (KU647195-KU647270). The biochemical properties and other analyzed data has been fully stated in this study.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Funding
This study was financially supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB15020402), the Natural Science Foundation of China (41771285, 41430858), the National Key Research and Development Program of China (2017YFD0200201) and the European Research Council Synergy Grant ERC-SyG-2013-610028 IMBALANCE-P.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- P
phosphorus
- iPSBs
inorganic phosphate solubilizing bacteria
- Olsen P
available P
- PVK
Pikovskaya medium
- NCBI
National Center for Biotechnology Information
- ANOVA
correlations and variance analyses
Footnotes
Bang-Xiao Zheng and Muhammad Ibrahim contributed equally to this work
Contributor Information
Bang-Xiao Zheng, Email: bxzheng@iue.ac.cn, Email: b.zheng@creaf.uab.cat.
Muhammad Ibrahim, Email: muhammad@iue.ac.cn.
Ding-Peng Zhang, Email: zdp1994@foxmail.com.
Qing-Fang Bi, Email: qfbi@iue.ac.cn.
Hong-Zhe Li, Email: hzli@iue.ac.cn.
Guo-Wei Zhou, Email: gwzhou@iue.ac.cn.
Kai Ding, Email: kding@iue.ac.cn.
Josep Peñuelas, Email: josep.penuelas@uab.cat.
Yong-Guan Zhu, Email: ygzhu@iue.ac.cn.
Xiao-Ru Yang, Phone: (+86) 592 6190560, Email: xryang@iue.ac.cn.
References
- Aislabie JM, Chhour K-L, Saul DJ, Miyauchi S, Ayton J, Paetzold RF, Balks MR. Dominant bacteria in soils of Marble point and Wright valley, Victoria land, Antarctica. Soil Biol Biochem. 2006;38(10):3041–3056. doi: 10.1016/j.soilbio.2006.02.018. [DOI] [Google Scholar]
- Babu-Khan S, Yeo TC, Martin WL, Duron MR, Rogers RD, Goldstein AH. Cloning of a mineral phosphate-solubilizing gene from Pseudomonas cepacia. Appl Environ Microbiol. 1995;61(3):972–978. doi: 10.1128/aem.61.3.972-978.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolan NS, Naidu R, Mahimairaja S, Baskaran S. Influence of low-molecular-weight organic acids on the solubilization of phosphates. Biol Fertil Soils. 1994;18(4):311–319. doi: 10.1007/BF00570634. [DOI] [Google Scholar]
- Chen YP, Rekha PD, Arun AB, Shen FT, Lai WA, Young CC. Phosphate solubilizing bacteria from subtropical soil and their tricalcium phosphate solubilizing abilities. Appl Soil Ecol. 2006;34(1):33–41. doi: 10.1016/j.apsoil.2005.12.002. [DOI] [Google Scholar]
- Chung H, Park M, Madhaiyan M, Seshadri S, Song J, Cho H, Sa T. Isolation and characterization of phosphate solubilizing bacteria from the rhizosphere of crop plants of Korea. Soil Biol Biochem. 2005;37(10):1970–1974. doi: 10.1016/j.soilbio.2005.02.025. [DOI] [Google Scholar]
- Elser J, Bennett E. Phosphorus cycle: a broken biogeochemical cycle. Nature. 2011;478(7367):29–31. doi: 10.1038/478029a. [DOI] [PubMed] [Google Scholar]
- Elser JJ, Bracken ME, Cleland EE, Gruner DS, Harpole WS, Hillebrand H, Ngai JT, Seabloom EW, Shurin JB, Smith JE. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol Lett. 2007;10(12):1135–1142. doi: 10.1111/j.1461-0248.2007.01113.x. [DOI] [PubMed] [Google Scholar]
- Farris MH, Olson JB. Detection of Actinobacteria cultivated from environmental samples reveals bias in universal primers. Lett Appl Microbiol. 2007;45(4):376–381. doi: 10.1111/j.1472-765X.2007.02198.x. [DOI] [PubMed] [Google Scholar]
- Goldstein AH. Recent progress in understanding the molecular genetics and biochemistry of calcium phosphate solubilization by gram negative bacteria. Biol Agric Hortic. 1995;12(2):185–193. doi: 10.1080/01448765.1995.9754736. [DOI] [Google Scholar]
- Gopalakrishnan S, Humayun P, Kiran BK, Kannan IGK, Vidya MS, Deepthi K, Rupela O. Evaluation of bacteria isolated from rice rhizosphere for biological control of charcoal rot of sorghum caused by Macrophomina phaseolina (Tassi) Goid. World J Microbiol Biotechnol. 2011;27(6):1313–1321. doi: 10.1007/s11274-010-0579-0. [DOI] [PubMed] [Google Scholar]
- Hu JL, Lin XG, Wang JH, Chu HY, Yin R, Zhang JB. Population size and specific potential of P-mineralizing and-solubilizing bacteria under long-term P-deficiency fertilization in a sandy loam soil. Pedobiologia. 2009;53(1):49–58. doi: 10.1016/j.pedobi.2009.02.002. [DOI] [Google Scholar]
- Jones DL, Oburger E. Solubilization of phosphorus by soil microorganisms. In: Bünemann EK, Oberson A, Frossard E, editors. Phosphorus in action. 1. New York: Springer; 2011. [Google Scholar]
- Karagöz K, Ateş F, Karagöz H, Kotan R, Çakmakçı R. Characterization of plant growth-promoting traits of bacteria isolated from the rhizosphere of grapevine grown in alkaline and acidic soils. Eur J Soil Biol. 2012;50:144–150. doi: 10.1016/j.ejsobi.2012.01.007. [DOI] [Google Scholar]
- Kaur G, Reddy MS. Effects of phosphate-solubilizing bacteria, rock phosphate and chemical fertilizers on maize–wheat cropping cycle and economics. Pedosphere. 2015;25(3):428–437. doi: 10.1016/S1002-0160(15)30010-2. [DOI] [Google Scholar]
- Kochian LV. Rooting for more phosphorus. Nature. 2012;488(7412):466–467. doi: 10.1038/488466a. [DOI] [PubMed] [Google Scholar]
- Kwak Y, Jung BK, Shin JH. Complete genome sequence of Pseudomonas rhizosphaerae IH5T (= DSM 16299T), a phosphate-solubilizing rhizobacterium for bacterial biofertilizer. J Biotechnol. 2015;193:137–138. doi: 10.1016/j.jbiotec.2014.11.031. [DOI] [PubMed] [Google Scholar]
- Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG. Clustal W and Clustal X version 2.0. Bioinformatics. 2007;23(21):2947–2948. doi: 10.1093/bioinformatics/btm404. [DOI] [PubMed] [Google Scholar]
- Letunic I, Bork P. Interactive tree of life (iTOL) v3: an online tool for the display and annotation of phylogenetic and other trees. Nucleic Acid Res. 2016;44(W1):W242–W245. doi: 10.1093/nar/gkw290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mehta S, Nautiyal CS. An efficient method for qualitative screening of phosphate-solubilizing bacteria. Curr Microbiol. 2001;43(1):51–56. doi: 10.1007/s002840010259. [DOI] [PubMed] [Google Scholar]
- Murphy J, Riley JP. A modified single solution method for the determination of phosphate in natural waters. Anal Chim Acta. 1962;27:31–36. doi: 10.1016/S0003-2670(00)88444-5. [DOI] [Google Scholar]
- Nautiyal CS. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol Lett. 1999;170(1):265–270. doi: 10.1111/j.1574-6968.1999.tb13383.x. [DOI] [PubMed] [Google Scholar]
- Oliveira CA, Alves VMC, Marriel IE, Gomes EA, Scotti MR, Carneiro NP, Guimarães CT, Schaffert RE, Sá NMH. Phosphate solubilizing microorganisms isolated from rhizosphere of maize cultivated in an oxisol of the Brazilian Cerrado Biome. Soil Biol Biochem. 2009;41(9):1782–1787. doi: 10.1016/j.soilbio.2008.01.012. [DOI] [Google Scholar]
- Olsen SR, Cole CV, Wantanabe FS, Dean LA. Estimation of available phosphorus in soils by extraction with sodium bicarbonate. Washington: USDA; 1954. [Google Scholar]
- Parkinson JA, Allen SE. A wet oxidation procedure suitable for the determination of nitrogen and mineral nutrients in biological material. Commun Soil Sci Plant Anal. 1975;6(1):1–11. doi: 10.1080/00103627509366539. [DOI] [Google Scholar]
- Penuelas J, Poulter B, Sardans J, Ciais P, van der Velde M, Bopp L, Boucher O, Godderis Y, Hinsinger P, Llusia J, Nardin E, Vicca S, Obersteiner M, Janssens IA. Human-induced nitrogen–phosphorus imbalances alter natural and managed ecosystems across the globe. Nat Commun. 2013;4:2934. doi: 10.1038/ncomms3934. [DOI] [PubMed] [Google Scholar]
- Richardson AE, Simpson RJ. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiol. 2011;156(3):989–996. doi: 10.1104/pp.111.175448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richardson AE, Barea JM, McNeill AM, Prigent-Combaret C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil. 2009;321(1–2):305–339. doi: 10.1007/s11104-009-9895-2. [DOI] [Google Scholar]
- Rodríguez H, Fraga R. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Biotechnol Adv. 1999;17(4):319–339. doi: 10.1016/S0734-9750(99)00014-2. [DOI] [PubMed] [Google Scholar]
- Rodríguez H, Fraga R, Gonzalez T, Bashan Y. Genetics of phosphate solubilization and its potential applications for improving plant growth-promoting bacteria. Plant Soil. 2006;287(1–2):15–21. doi: 10.1007/s11104-006-9056-9. [DOI] [Google Scholar]
- Shen JP, Zhang LM, Zhu YG, Zhang JB, He JZ. Abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea communities of an alkaline sandy loam. Environ Microbiol. 2008;10(6):1601–1611. doi: 10.1111/j.1462-2920.2008.01578.x. [DOI] [PubMed] [Google Scholar]
- Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013;30(12):2725–2729. doi: 10.1093/molbev/mst197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umezawa K, Takeda K, Ishida T, Sunagawa N, Makabe A, Isobe K, Koba K, Ohno H, Samejima M, Nakamura N, Igarashi K, Yoshida M. A novel pyrroloquinoline quinone-dependent 2-keto-d-glucose dehydrogenase from Pseudomonas aureofaciens. J Bacteriol. 2015;197(8):1322–1329. doi: 10.1128/JB.02376-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Heijden MG, Bardgett RD, van Straalen NM. The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol Lett. 2008;11(3):296–310. doi: 10.1111/j.1461-0248.2007.01139.x. [DOI] [PubMed] [Google Scholar]
- Xiong J, Liu Y, Lin X, Zhang H, Zeng J, Hou J, Yang Y, Yao T, Knight R, Chu H. Geographic distance and pH drive bacterial distribution in alkaline lake sediments across Tibetan Plateau. Environ Microbiol. 2012;14(9):2457–2466. doi: 10.1111/j.1462-2920.2012.02799.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xuan Y, Xu L, Tian HZ, Liu GH, Cui M. Isolation and characterization of phosphate-solubilizing bacteria from walnut and their effect on growth and phosphorus mobilization. Biol Fertil Soils. 2011;47(4):437–446. doi: 10.1007/s00374-011-0548-2. [DOI] [Google Scholar]
- Yu X, Liu X, Zhu TH, Liu GH, Mao C. Co-inoculation with phosphate-solubilzing and nitrogen-fixing bacteria on solubilization of rock phosphate and their effect on growth promotion and nutrient uptake by walnut. Eur J Soil Biol. 2012;50:112–117. doi: 10.1016/j.ejsobi.2012.01.004. [DOI] [Google Scholar]
- Zhao K, Penttinen P, Zhang XP, Ao XL, Liu MK, Yu XM, Chen Q. Maize rhizosphere in Sichuan, China, hosts plant growth promoting Burkholderia cepacia with phosphate solubilizing and antifungal abilities. Microbiol Res. 2014;169(1):76–82. doi: 10.1016/j.micres.2013.07.003. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The 16S rRNA sequences were uploaded to the NCBI Sequence Read Archive (KU647195-KU647270). The biochemical properties and other analyzed data has been fully stated in this study.