

Abstract
Approximately 0.7 million tons of azo dyes are synthesized each year. Azo dyes are composed of one or more R1-N=N-R2 linkages. Studies have shown that both mammalian and microbial azoreductases cleave the azo bonds of the dyes to form compounds that are potentially genotoxic. The human gastrointestinal tract harbors a diverse microbiota comprised of at least several thousand species. Both water-soluble and water-insoluble azo dyes can be reduced by intestinal bacteria. Some of the metabolites produced by intestinal microbiota have been shown to be carcinogenic to humans although the parent azo dyes may not be classified as being carcinogenic. Azoreductase activity is commonly found in intestinal bacteria. Three types of azoreductases have been characterized in bacteria. They are flavin dependent NADH preferred azoreductase, flavin dependent NADPH preferred azoreductase, and flavin free NADPH preferred azoreductase. This review highlights how azo dyes are metabolized by intestinal bacteria, mechanisms of azo reduction, and the potential contribution in the carcinogenesis/mutagenesis of the reduction of the azo dyes by intestinal microbiota.
Keywords: Azo dye, Azoreductase, Degradation, Metabolite, Genotoxicity, Intestinal Microbiota, Review
2. INTRODUCTION
One of the remarkable abilities of the human eye is to recognize colors; therefore compounds like dyes are used to enhance the attractiveness of objects. Today, there are thousands of various types of dyes used widely in foods, pharmaceuticals, cosmetics, textiles and plastics (36, 90, 128), some of which may pose a risk to human and ecological health due to their toxic properties. Dyes have been used for thousands of years and approximately 0.7 million tons of dyes are synthesized annually worldwide (41, 110). Azo dyes are a group of compounds containing one or more R1-N=N-R2 bonds, some of the reduced products of the dyes are carcinogenic (59). Azo dyes represent the majority of all the dyes used in today’s society, because the azo bond facilitates π-electron delocalization, which gives rise to the absorption at visible spectrum wavelengths (78).
Microorganisms have evolved in a long-lasting mutual interactions with human beings to influence host physiology (67). Although the study of intestinal bacteria was hindered by the fact that most of the bacteria were unculturable ex vivo (67), recent molecular and genomic sequencing approaches have revealed a vast and dynamic consortium of bacteria, with an estimated 100 trillion bacteria inhabiting in the human gut (68). The function of intestinal bacteria includes enhancing host digestive efficiency, providing instructive signals for several aspects of intestinal development, and protecting their hosts against pathogenic infections (68). As a result of enhancing host digestive efficiency, germ-free rodents must consume about 30% more calories to sustain their body weight than that of conventionally raised animals (6). The relationship of host immune system and the intestinal microbiota has been reviewed recently (68). Colonization barrier disruption of the intestinal bacteria has been linked to several types of diseases to the host. For example, inflammatory bowel disease (IBD) was associated with activating inappropriately the host mucosal immune system by losing of tolerance to gut commensals (51, 82).
Human intestinal bacteria also play a role in degrading various xenobiotics to metabolites, some of which are involved in genotoxicity and carcinogenesis. Human exposure to xenobiotics including azo dyes occurs via ingestion, skin contact, or inhalation. Azo dyes can be biotransformed into colorless aromatic amines, some of these may become less toxic than the original dye, but others, such as arylamines and free radicals, are potentially carcinogenic (40). Sudan azo dyes such as Sudan I, II, III, IV and Para Red are often illegally used to enhance and/or maintain the appearance of food products and they have been found as food contaminants (28, 138). The initial step in the anaerobic metabolism of azo dyes by intestinal microbiota is reduction of the azo bond by the NAD(P)H dependent azoreductases. Three distinct groups of azoreductases have been found, flavin dependent NADH preferred azoreductase, flavin dependent NADPH preferred azoreductase, and flavin free NADPH preferred azoreductase (31, 87). The flavin dependent NADH preferred azoreductase has been purified from Escherichia coli (95), Enterococcus faecalis (33), and Pseudomonas aeruginosa (134). The flavin dependent NADPH preferred azoreductase has been purified from Bacillus cereus OY1–2 (129), and similar genes were found from Rhodobacter sphaaeroides (16), Staphylococcus aureus (32), and Saccharomyces cerevisiae (80). The flavin free NADPH preferred azoreductase has been purified from Xenophilus azovorans KF46F (18) and Pigmentiphaga kullae K24 (19, 31). There are also some studies on the characterization and crystal structure of azoreductases in B. subtilis (93), E. coli (69, 70), Ent. faecalis (34, 81, 83), P. aeruginosa (113, 134, 135), and S. cerevisiae (80). Azo dyes can be reduced by NAD(P)H catalyzed by azoreductases as shown in Figure 1
Figure 1.
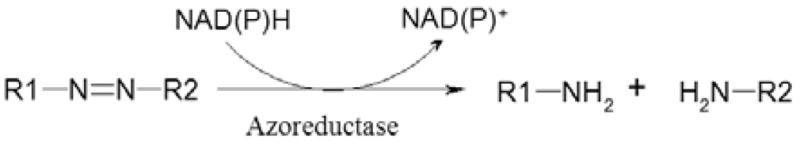
The reduction of azo dyes by NAD(P)H catalyzed by azoreductase.
The aim of this review is to summarize the mechanism of azo dye degradation by human intestinal bacteria and the potential toxicity of azo dyes and their metabolites. We also provide a brief discussion on azoreductases from intestinal bacteria and diversity of intestinal bacteria and their capability to reduce azo dyes.
4. HUMAN INTESTINAL MICROBIOTA
The human gastrointestinal tract contains about 1014 bacterial cells, 10 times the number of human cells present in our bodies. The largest diversity of the human associated microbes is found in the intestinal tract with about 1011 per gram in stool (121). It is reported that most of the intestinal bacteria are members of the Firmicutes and Bacteroidetes phyla (52). The Gram-positive Firmicutes phyla include Clostridia class and the families of Enterococcaceae and Lactobacillacea. The Gram-negative Bacteroidetes are comprised of several Bacteroides species. The other relatively few (less than 10%) intestinal bacteria belong to Proteobacteria, Actinobacteria, Fusobacteria, and Verrucomicrobia phyla (52).
With recent advances in genome sequencing, our understanding of role of commensal microbiota in causing disease and mutualistic relationship to host has been advancing (49). The human intestinal microbiota are essential to individual health. They play important roles in the digestion, vitamin synthesis, epithelial development, pathogen resistance, and immune responses (15). A classic example in nutrition is polysaccharide degradation. Most of the polymers are not degraded by the host, while, they are energy sources of the intestinal microbiota. Herbivores can get up to 70% of their energy intake from the digestion of polymers by intestinal bacteria (57). Germ-free mice need additional vitamin K and certain B vitamins (137), because the intestinal bacteria produce these vitamins in conventional animals (66). Moreover, the intestinal microbiota are able to change the metabolic potential of liver by influencing hepatic gene expression through altering its xennobiotic response to drugs (17).
An initial barrier in studying the microbial ecology of the human gastrointestinal tract was that most of the intestinal bacteria were difficult to culture in the laboratory. Recently, modern molecular methods including small-subunit rRNA (16S rRNA) gene sequencing (52), quantitative PCR coupled to denaturing gradient gel electrophoresis (DGGE) (94) have been applied to study the uncultured intestinal microbial community. Metagenomics, a culture independent genomic analysis method, provides a valuable tool for the study of gut microbial diversity. By employing high resolution nuclear magnetic resonance (NMR) spectroscopy and mass spectrometry (MS), metabolomics can simultaneously analyze hundreds to thousands of small molecule metabolites in the samples, therefore, it has been also used in the study of intestinal bacteria, especially in the study of host-microbe interactions (64).
4. AZO DYES
All azo dyes contain one or more azo bonds. Although azo dyes can supply a complete rainbow of colors in theory, most of commercial azo dyes supply yellow, orange, and red colors. Among all the dyes synthesized annually worldwide, 15% of the used dyes produced enter the environment as industrial effluents (110), which reduces sunlight penetration into the deeper layer of the water. Therefore, serious pollution occurs (119). Environmental release of azo dyes can also cause contamination of agricultural products. Some physical and chemical techniques including adsorption, coagulation and membrane processes have been developed to remove azo dyes from the wastewaters (127). However, the treatments are limited by the cost and the possibility of causing secondary pollution. Presently, bio-decolorization is being intensively investigated because of its low cost and environmental friendly nature (43, 60, 71, 74, 120, 122, 126).
4.1. Azo dyes containing amine group
Based on the structure, azo dyes can be divided into two groups, one group contains aromatic amine radical (R1-N=N-R2-NH2), such as 4-aminoazobenzene, and the other group can produce amine components only after reduction (R1-N=N-R2), for example, Methyl Red. Some azo dyes have been used safely in foods, drugs and cosmetics, while others are carcinogenic.
Most of the azo dyes containing amine group are carcinogenic. As is the case for 4-aminoazobenzene, the compound has high hepatocarcinogenicity to male mice when given as a single intraperitoneal injection, and a linearly relationship has been found between the hepatoma multiplicity induced by the dye and the dose of 0.017 to 0.15 μmol/g body weight. However, female mice were resistant to tumor induction under these conditions due to the insensitivity of them to 4-aminoazobenzene and other hepatic carcinogens (48). The dye also produces liver tumors in rats by oral administration and epidermal tumors by application to the skin (2).
o-Aminoazotoluene can produce tumors of urinary bladder, gall bladder and liver in dogs (96). The carcinogenicity of the dye was also found in mice, rats and hamsters producing tumors of the urinary bladder, gall bladder, lung and liver (2). It is strongly mutagenic to Salmonella Typhimurium TA98 and TA100 with S9 activation (47). The structures of this type of azo dye are included in Table 1.
Table 1.
Chemical structure of azo dyes
| Azo dye | Structure | Group(a) |
|---|---|---|
| 4-Aminoazobenzene |
|
2B(b) |
| o-Aminoazotoluene |
|
2B |
| Acid Orange 63 |

|
-(c) |
| Acid Orange 8 |

|
- |
| Acid Red 88 |

|
- |
| Allura Red |

|
- |
| Congo Red |
|
- |
| Methyl Orange |
|
- |
| Methyl Red |

|
3 (d) |
| Orange II |

|
- |
| Orange G |

|
3 |
| Para Red |

|
- |
| Ponceau BS |

|
- |
| Ponceau S |

|
- |
| 1-Phenylazo-2-naphthol |

|
3 |
| 1-[(2,4-Dimethylphenyl)azo]-2-nahpthalenol |

|
3 |
| 1-(4-Phenylazophenylazo)-2-nahpthalenol |

|
3 |
| 1-[[2-Methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol |

|
3 |
| Sunset Yellow |

|
3 |
| Tartrazine |
|
- |
Classified by the IARC monographs,
Possibly carcinogenic to humans,
Not on the list of classification by International Agency for Research on Cancer,
Not classifiable as to its carcinogenicity to humans
4.2. Azo dyes used in food
Azo dyes can provide a complete rainbow of colors for use in the food industry. Azo dyes are used as food additives to enhance color of the manufactured foods, which account for approximately 60–70% of all dyes used in the food process. Tartrazine (1H-pyrazole-3-carboxylic acid, 4,5-dihydro-5-oxo-1-(4-sulfophenyl)-4-((4-sulfophenyl)azo)-,tri-sodium salt, Table 1) is a yellow coloring agent, and commonly used in food and pharmaceutical products. Tartrazine has no mutagenic and carcinogenic potential with a daily intake of 7.5 mg/kg body weight (53). Allura Red (Table 1) is classified as nongenotoxic, and is permitted for use as a food coloring agent. Sunset Yellow (Table 1) is not mutagenic in Salmonella Typhimurium testing strain (26). The other non-food azo dyes further being discussed in this paper are all mutagenic/carcinogenic or have the potential to be genotoxic in mammals (Table 1).
4.3. Sudan dyes as food contaminants
Sudan dyes are non-ionic fat-soluble azo dyes used in plastics, printing inks, waxes, leather, fabrics, and floor polish (8, 139). The chemical name of Sudan I, II, III, IV, and Para Red are 1-phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-nahpthalenol, 1-(4-phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, and 1-(4-nitrophenylazo)-2-naphthol, respectively, and the structures of these dyes are shown in Table 1. These dyes are classified by the International Agency for Research on Cancer (IARC) as category 3 carcinogens to humans. Although Sudan dyes are banned for food usage in most countries, they are illegally used to maintain the color in food products because of their low cost, bright staining, and wide availability. In May 2003, some chili powder and a variety of foodstuffs were found contaminated by Sudan dyes throughout Europe and Asia (63). Sudan I was also found in China in chili sauce, meat and eggs (65). Therefore, accurate determination of the concentration of Sudan dyes in foodstuffs by regulatory authorities is very important. The analytical techniques for determination of Sudan dyes have been reviewed recently (108). Several methods based on liquid chromatography (LC), enzyme-linked immunosorbent assay (ELISA), and gas chromatography-mass spectrometry (GC-MS) have been developed to detect Sudan dyes. The LC methods are the main methods to analyze Sudan dyes. In most cases, C18 columns were used as the stationary phases, and the mixtures of acetonitrile with acetate or formic acid buffers used as mobile phases. The ultra-violet visible (UV-vis) absorbance and photodiode array (PDA) detectors are most widely used in the analysis of Sudan dyes. ELISA is a sensitive, selective, and rapid method for the determination of Sudan dyes. The ELISA method has been developed for Sudan dyes in chili powder, ketchup and egg yolk (8). GC-MS also has been used to determine Sudan dyes in chili powder (65).
4.4. Mechanism of the reduction of azo dyes
Azo dyes are degraded mainly via reductive reactions. When the azo dyes enter body through ingestion, both the intestinal microorganisms and the enzymes in liver are responsible for the reduction of the dyes. Some studies showed that the intestinal bacterial reductases may be more important than the liver enzymes in metabolism of the azo dyes (38, 39, 53). The azo dyes are reduced to aromatic amines by a type of reductase named azoreductases, which are widely distributed in intestinal bacteria (Figure 1). In some cases, the azo dyes can be degraded via oxidative reactions catalyzed by oxidases and peroxidases (30). The oxidative metabolites of azo dyes were also found in the bile of rat (86, 117, 131, 136).
Three groups of azoreductases have been found in bacteria (31). Two groups are flavin dependent, which are composed of NADH-preferred and NADPH-preferred azoreductase. Another group is flavin free azoreductase. The catalytic mode of flavin dependent enzymes, such as the azoreductases from E. coli (AzoR) and from Ent. faecalis (AzoA), is a ping-pong Bi-Bi mechanism, the electrons from NAD(P)H are transferred to substrate via FMN (33, 95). AzoA has two separate active sites, and FMN lies in one site, the other site provides room for both NAD(P)H and substrate binding (33, 34, 81). In the case of Methyl Red reduction by AzoA, NADH firstly transfers the electron to FMN and then releases from the enzyme as NAD+, after that, Methyl Red bind with the enzyme and the electron is transferred from FMNH2 to Methyl Red. As for reducing Methyl Red, two cycles of the ping-pong mechanism are required, as shown in Figure 2. In theory, a hydrazo intermediate should exist in the reduction process. However, only one research group has detected the intermediate from the reduction of Methyl Red using MS/MS technique. The authors also reported that the intermediate is not stable and can be reduced quickly to the amine (16). A different mechanism is found in the flavin free azoreductase. In the case of Orange I reduction by azoreductase from P. kullae K24 (AzoB), AzoB joins both substrates (NADPH and Orange I) together in the active site at the same time, and then a direct hydride transfer occurs in the reaction center of AzoB. This mechanism requires a sophisticated binding mode, therefore, this group of azoreductases shows narrow substrate specificity, only Orange I can be degraded by AzoB in the tested azo dyes (31). On the other hand, the flavin dependent azoreductase families have a broad substrate specificity. AzoA from Ent. faecalis can degrade Methyl Red, Orange II, Amaranth, Ponceau BS, Ponceau S, and Menadione (33).
Figure 2.

Proposed pathway of the reduction of Methyl Red by Ent. faecalis AzoA. EFMN, flavin dependent azoreductase; EFMNH2, reduced flavin dependent azoreductase.
The solubility of azo dyes and the distribution of azoreductase in the cell also affect the dye biodegradation. As for the water-soluble dyes, for example Methyl Red, the dye molecules were first dispersed in solution, then went through the bacterial cell membrane, and subsequently reduced in the cytoplasm by the azoreductase (55). As for the water-insoluble dyes, for example 1-[(2,4-dimethylphenyl)azo]-2-naphthalenol, the dye particles were quickly bound to the cells after being added to the culture, and then reduced on the cell membrane. After that, the cells bound other dye particles from the culture, therefore, the amount of dye bound to the cells kept constant until the dye disappeared in the supernatant of the culture (139). Because almost 100% of azoreductase activity of E. coli was found in the cytoplasm, E. coli lacks the ability to significantly reduce water-insoluble azo dyes. However, about 55% of the azoreductase activity of Ent. faecalis was bound in the membrane, therefore, some water-insoluble azo dyes including 1-phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-naphthalenol, 1-(4-phenylazophenylazo)-2-naphthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, and 1-(4-nitrophenylazo)-2-naphthol can be reduced by the bacterium (139). The mutants of Shewanella oneidensis MR-1 generated by random transposon also confirmed that the outer membrane, cytoplasmic membrane, and the periplasm all participated in the reduction of azo dyes (24).
5. TOXICITY OF AZO DYE METABOLITES PRODUCED BY HUMAN INTESTINAL BACTERIA
Azo dyes are degraded mainly via reductive reactions, therefore, we will briefly discuss the toxicity of some of the reduction products of the dyes in this review. Both mutagenicity and carcinogenicity assays are taken into account for the azo dyes and their metabolites. In addition, some azo dyes or metabolites showed marked selectively toxic/carcinogenic, for example, 4-aminoazobenzene demonstrates hepatocarcinogenicity to male mice, but not to female mice. Therefore the clinical data and the literature should be retrospectively analyzed (25).
5.1. Genotoxicity of metabolites of azo dyes
The reduction of Orange G, 1-phenylazo-2-naphthol, and 1-(4-phenylazophenylazo)-2-nahpthalenol can produce aniline (Table 2). Aniline is one of the most important compounds of industrial chemistry. About 2.3 million tons aniline were produced in 1996 by more than 40 companies around the world. Aniline is mainly used in the synthesis of diphenylmethane-4,4′-diisocyanate (MDI), rubber chemicals, dyes, pesticides, pharmaceuticals, and fibers (132). The genotoxic activities of aniline has been reviewed by Bomhard and Herbold (21). When Salmonella Typhimurium and E. coli systems were employed in the mutagenicity test, aniline was proved to be non-mutagenic with or without metabolic activation (by S9 mix). Aniline did not induce direct DNA damage to E. coli and Bacillus subtilis in rec assays and in the Umu test with S. Typhimurium (84, 88, 115). But at the high concentrations (156–624 μg/plate), aniline has a weakly positive role in the DNA repair assay without metabolic activation system (S9) (45, 46). At the HPRT locus of V79 cells, aniline did not induce mutations even at concentrations over 1,860 μg/ml (54). At a relatively high concentration aniline can induce the chromosome aberrations, which means a clastogenicity by aniline. Aniline can induce tumors in the spleen of rats (21), but does not induce bladder cancer in humans or animals (5). The data from a contact allergen surveillance network between 1992 and 2004 showed that aniline is not an independent sensitizer, but may elicit allergic reactions in its substituted compounds (132).
Table 2.
Chemical structure of metabolites from azo dyes
| Name | Structure | Substrate | Grou p(a) |
|---|---|---|---|
| Aniline |

|
Orange G, 1-phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-naphthalenol | 3(b) |
| 2,4-Dimethylaniline |

|
1-[(2,4-Dimethylphenyl)azo]-2-naphthalenol | 3 |
| 4-Aminobenzenesulfonic acid |

|
Methyl Orange, Orange II, Ponceau BS, Ponceau S | -(c) |
| p-Nitroaniline |
|
Para Red | - |
| o-Toluidine |

|
1-[[2-Methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol | 1(d) |
| N,N-Dimethyl-p-phenylenediamine |

|
Methyl Orange, Methyl Red | - |
| p-Phenylenediamine |

|
1-(4-Phenylazophenylazo)-2-naphthalenol | 3 |
| 2,5-Diminobenzene sulfonic acid |

|
Ponceau BS, Ponceau S | - |
| 1-Amino-2-naphthol |

|
Acid Orange 8, Acid Red 88, Orange II, Para Red, Ponceau BS, 1-phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-naphthalenol, 1-(4-phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol | - |
| Benzidine |
|
Congo Red | 1 |
| Benzidine-3-Sulfonic Acid |

|
Acid Orange 63 | - |
Classified by the IARC monographs,
Not classifiable as to its carcinogenicity to humans,
Not on the list of classification by International Agency for Research on Cancer,
Carcinogenic to human
2,4-Dimethylaniline can elicit positive DNA repair responses and possess mutagenicity and carcinogenicity (141). In the Vibrio test system, 2,4-dimethylaniline showed direct genotoxicity (97). In the Xenopus frog test, 2,4-dimethylaniline showed no mortality even at a concentration of 100 mg/L, but possessed teratogenic properties (98). One of the metabolites of 1-[(2,4-dimethylphenyl)azo]-2-naphthalenol is 2,4-dimethylaniline (Table 2).
It was shown that 4-aminobenzenesulfonic acid (4-ABS) did not possess a mutagenic effect with or without S9 (13, 14, 85). It should be noted that 4-ABS is a metabolite of food dyes of Sunset Yellow and Tartrazine. This compound can also be produced in the reduction of Methyl Orange, Orange II, Ponceau BS, and Ponceau S (Table 2).
p-Nitroaniline was first reported as very weakly mutagenic or non-mutagenic in the presence of S9 mix in S. Typhimurium strain TA98 (124). The mutagenicity of the compound was later confirmed in the test system of S. Typhimurium strain TA98 with or without S9 activation, but the results were negative for all other strains. Equivocal evidence of carcinogenic activity was found in mice (3). p-Nitroaniline is a metabolite of Para Red (Table 2).
o-Toluidine was first synthesized in 1844, and was mainly used as a chemical intermediate in dye, rubber, and pharmaceutical production (44, 123). o-Toluidine is a mutagen, and is carcinogenic to experimental animals (4). The aromatic amine o-toluidine has been recognized as carcinogenic (Group 1) to humans by IARC. The carcinogenicity of o-toluidine as a human bladder carcinogen has been reviewed (123). The recent research also supports the hypothesis that o-toluidine is a human bladder carcinogen (29). o-Toluidine also has genotoxic activity. o-Toluidine can produce DNA damage and induce aneuploidy in fungi and mammalian cultured cells. The genetic toxicology of o-toluidine has been reviewed (44). The p-toluidine-m-sulfonic acid did not show the mutagenicity, and the carcinogenicity of this compound is unknown (141). o-Toluidine is a metabolite of 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol (Table 2).
N,N-Dimethyl-p-phenylenediamine (DMPD) possesses strong mutagenicity with or without metabolic activation (13, 14, 37). The mutagenicity of DMPD did not depend on the dose, because the highest mutagenic effect was obtained with the lowest dose (10 μg/plate) (13). Since the nonmethylated compound p-phenylenediamine is not a mutagen, the dimethyl group contributes to the mutagenicity of DMPD, when located in the para position to the amino group on the benzene ring (13). A similar result was also reported in the mutagenicity of other dyes showing methyl substituents were more genotoxic than that of the nonmethylated dyes (118). DMPD is a metabolite of Methyl Orange and Methyl Red, which are used widely in laboratory (Table 2).
p-Phenylenediamine is one of the primary intermediates in the azo dyes. Poisoning with p-phenylenediamine causes angioneurotic edema, intravascular hemolysis, rhabdomyolysis with acute renal failure (9). It was reported that there was no convincing evidence of carcinogenicity in rats and mice by dietary administration of p-phenylenediamine dihydrochloride (1). In vitro, a weak genotoxic potential was found. In the S. Typhimurium test systems, p-phenylenediamine possessed weak genotoxicity (58). The carcinogenicity of hair dyes which contained p-phenylenediamine has been reviewed (20). In the Umu test using S. Typhimurium, 2,5-diaminotoluene showed positive genotoxicity with S9 mix (140). It is reported that 2,5-diaminobenzene sulfonic acid induced mutagenicity, while the carcinogenicity is unknown (141). The compounds of p-phenylenediamine and 2,5-diaminotoluene can be produced from the reduction of 1-(4-phenylazophenylazo)-2-naphthalenol and 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, respectively. The compound of 2,5-diaminobenzene sulfonic acid is a product of the reduction of Ponceau BS and Ponceau S (Table 2).
1-Amino-2-naphthol, which is one of the components of an important group of 1-amino-2-naphthol-based azo dyes, has been reported to be a carcinogen. Although there is a study reporting that it was not mutagenic when tested in the Salmonella/microsome test system (36, 37), the later studies showed that this compound is mutagenic (50, 61). In the standard protocols with S9, 1-amino-2-naphthol was mutagenic to S. Typhimurium strain TA100 but not to strain TA98 (50). When Orange II was reduced, the toxicity increased almost 100-fold. While, when the structurally similar compound Sunset Yellow was reduced, the toxicity slightly decreased. Therefore, the product of 1-amino-2-naphthol should contribute to the increased toxicity (61). This study also proved that the metabolite of Sunset Yellow 1-amino-2-naphthol-6-sulphonate was not mutagenic, which could be due to the decrease in membrane permeability of this compound (61). A study about the mutagenicity of 1-amino-2-naphthol-based azo dyes also indicated that sulphonated 1-amino-2-naphthol at strategic sites inhibits the mutagenicity (111). Acid Orange 8, Acid Red 88, Orange II, Para Red, Ponceau BS, 1-Phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-naphthalenol, 1-(4-phenylazophenylazo)-2-naphthalenol, and 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol can be reduced to produce 1-amino-2-naphthol; while, the food dyes of Allura Red and Sunset Yellow can produce 1-amino-2-naphthol-6-sulphonate (Table 2).
Benzidine is known as carcinogen for the human urinary bladder (62). In Japan, a recent study found that the concentration of benzidine in placemats made of cotton exceeds the EU regulation limits (72). In the testing systems with mouse liver enzyme preparations, benzidine was mutagenic to S. Typhimurium strain TA98 and TA100 (75). In the testing systems with FMN and hamster liver S9, benzidine was also mutagenic (102). The nitro derivative of benzidine was more mutagenic than that of parent compound in TA98 without S9 (142). While, the addition of a sulfonic acid group to benzidine reduced the mutagenicity (10). In addition, when both sides of the azo linkage of the azo dyes were sulfonated, the compounds were not carcinogenic in any animal species (27). The similar situation was also found with the mutagenicities of aniline and 4-aminobenzenesulfonic acid, 1-amino-2-naphthol, and 1-amino-2-naphthol-6-sulphonate as discussed before. In United States today, the production and use of benzidine has been banned (11). Benzidine is a product of the reduction of Congo Red, and the sulfonated benzidine is a product of the reduction of Acid Orange 6 (Table 2).
5.2. Capability of human intestinal microbiota to reduce azo dyes
The main intestinal bacteria and their roles in the reduction of azo dyes are shown in Table 3. Bacteroides vulgatus is one of the most abundant bacterial species in the human colon (92), and may play a role in the pathogenesis of Crohn’s disease (112). B. vulgatus can reduce 1-(4-phenylazophenylazo)-2-naphthalenol and 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol completely, and can partially reduce 1-phenylazo-2-naphthol and Para Red (139).
Table 3.
Some prevalent intestinal bacteria and their activities for the reduction of azo dyes
| Organism | Azo dyes | Azoreduction products | Reference |
|---|---|---|---|
| Bacteroides vulgatus | 1-Phenylazo-2-naphthol, 1-(4-phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, Para Red | Aniline, 1-amino-2-naphthol, p-phenylenediamine, o-toluidine, 2-methyl-1,4-benzenediamine, p-nitroaniline | (139) |
| Bacteroides ovatus | 1-Phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-nahpthalenol, 1-(4-phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, Para Red | Aniline, 1-amino-2-naphthol, 2,4-dimethylaniline, p-phenylenediamine, o-toluidine, 2-methyl-1,4-benzenediamine, p-nitroaniline | (139) |
| Bacteroides distasonis | 1-Phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-nahpthalenol, 1-(4-phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, Para Red | Aniline, 1-amino-2-naphthol, 2,4-dimethylaniline, p-phenylenediamine, o-toluidine, 2-methyl-1,4-benzenediamine, p-nitroaniline | (139) |
| Baceroides fragilis | 1-Phenylazo-2-naphthol, 1-(4-phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, Para Red | Aniline, 1-amino-2-naphthol, p-phenylenediamine, o-toluidine, 2-methyl-1,4-benzenediamine, p-nitroaniline | (139) |
| Bacteroides thetaiotaomicron | Amaranth, Sunset Yellow, Tartrazine, Ponceau SX, Methyl Orange, Orange II, Allura Red, 1-phenylazo-2-naphthol, 1-(4-phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, Para Red | 4-Animobenzenesulfonic acid, 4-amino-3-hydroxy-2,7-naphthalenedisulfonic acid, 4-aminobenzenesulfonic acid, 4-amino-3-hydroxy-7-naphthalenesulfonic acid, 5-hydroxy-1-(4-sulforatophenyl)-4-amino-1H-pyrazole-3-carboxylate, 1-amino-2,4-dimethyl-5-benzenesulfonic acid, 3-amino-4-hydroxy-1-naphthalenesulfonic acid, N,N-dimethyl-p-phenylenediamine, 1-amino-2naphthol, 1-amino-2-methoxy-5-methyl-4-benzenesulfonic acid, aniline, p-phenylenediamine, o-toluidine, 2-methyl-1,4-benzenediamine, p-nitroaniline | (38, 130, 139) |
| Bifidobacterium longum | 1-(4-Phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, Para Red | Aniline, 1-amino-2-naphthol, p-phenylenediamine, o-toluidine, 2-methyl-1,4-benzenediamine, p-nitroaniline | (139) |
| Bifidobacterium adolescentis | 1-(4-Phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol | Aniline, 1-amino-2-naphthol, p-phenylenediamine, o-toluidine, 2-methyl-1,4-benzenediamine | (139) |
| Bifidobacterium infantis | 1-Phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-nahpthalenol, 1-(4-phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, Para Red | Aniline, 1-amino-2-naphthol, 2,4-dimethylaniline, p-phenylenediamine, o-toluidine, 2-methyl-1,4-benzenediamine, p-nitroaniline | (139) |
| Clostridium perfringens | Amaranth, 1-phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-nahpthalenol, 1-(4-phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, Para Red | 4-Animobenzenesulfonic acid, 4-amino-3-hydroxy-2,7-naphthalenedisulfonic acid, aniline, 1-amino-2-naphthol, 2,4-dimethylaniline, p-phenylenediamine, o-toluidine, 2-methyl-1,4-benzenediamine, p-nitroaniline | (87, 139) |
| Clostridium botulinum | 1-Phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-nahpthalenol, 1-(4-phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, Para Red | Aniline, 1-amino-2-naphthol, 2,4-dimethylaniline, p-phenylenediamine, o-toluidine, 2-methyl-1,4-benzenediamine, p-nitroaniline | (139) |
| Clostridium difficile | 1-Phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-nahpthalenol, 1-(4-phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, Para Red | Aniline, 1-amino-2-naphthol, 2,4-dimethylaniline, p-phenylenediamine, o-toluidine, 2-methyl-1,4-benzenediamine, p-nitroaniline | (139) |
| Enterococcus faecalis | 1-Phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-nahpthalenol, 1-(4-phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, Para Red, Methyl Red, Orange II, Orange G, Amaranth, Ponceau BS, Ponceau S | Aniline, 1-amino-2-naphthol, 2,4-dimethylaniline, p-phenylenediamine, o-toluidine, 2-methyl-1,4-benzenediamine, p-nitroaniline, N,N-dimethyl-p-phenylenediamine, 4-animobenzenesulfonic acid, 4-amino-3-hydroxy-5,8-naphthalenedisulfonic acid, 4-amino-3-hydroxy-2,7-naphthalenedisulfonic acid, 2,5-diminobenzenesulfonic acid | (33, 139) |
| Enterococcus faecium | 1-Phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-nahpthalenol, 1-(4-phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, Para Red | Aniline, 1-amino-2-naphthol, 2,4-dimethylaniline, p-phenylenediamine, o-toluidine, 2-methyl-1,4-benzenediamine, p-nitroaniline | (139) |
| Escherichia coli | Methyl Red, Ponceau BS, Orange II | Aniline, N,N-dimethyl-p-phenylenediamine, 4-animobenzenesulfonic acid, 2,5-diminobenzenesulfonic acid, 1-amino-2-naphthol | (55, 95) |
| Lactobacillus paracasei | Methyl Orange, Acid Orange 8, Tartrazine, Acid Orange 63, 1-phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-nahpthalenol, 1-(4-phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, Para Red | N,N-Dimethyl-p-phenylenediamine, 4-animobenzenesulfonic acid, 4-amino-3-methylbenzenesulfonic acid, 1-amino-2-naphthol, 4-aminobenzenesulfonic acid, 5-hydroxy-1-(4-sulforatophenyl)-4-amino-1H-pyrazole-3-carboxylate, 3-methyl-4-amino-1-phenyl-1H-pyrzaol-5ol, benzidine-3-sulfonic acid, 4-methylphenyl-4-aminobenzenesulfonate, aniline, 2,4-dimethylaniline, p-phenylenediamine, o-toluidine, 2-methyl-1,4-benzenediamine, p-nitroaniline | (101, 139) |
Bacteroides ovatus is a Gram-negative intestinal bacterium and can causes serum antibody responses in IBD patients (114). Azoreductase activity was already found in B. ovatus (87). The strain B. ovatus ATCC 8483 can reduce 1-phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-naphthalenol, 1-(4-phenylazophenylazo)-2-naphthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol and Para Red (139).
Bacteroides distasonis is one of the most common species from human feces, and was reclassified as Parabacteroides distasonis (116). P. distasonis has the activity to reduce 1-phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-naphthalenol, 1-(4-phenylazophenylazo)-2-naphthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, and Para Red (139).
Baceroides fragilis is one of the most common species from the normal human colonic microbiota, however the bacterium can cause abscesses, soft-tissue infections and bacteremias to the host (91). The crude extract of B. fragilis has a strong fibrinogenolytic activity suggesting an important biological factor in Bacteroides infection (35). B. fragilis can reduce 1-phenylazo-2-naphthol, 1-(4-phenylazophenylazo)-2-naphthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol and can partially degrade Para Red (139).
Bacteroides thetaiotaomicron is an anaerobe and predominate in the intestinal microbiota, accounting for 6% of all bacteria in the human intestine (143). B. thetaiotaomicron has a central role in nutrient metabolism, especially in the starch utilization (42, 109) and has the activity to reduce Amaranth, Sunset Yellow (38, 130), Tartrazine, Ponceau SX, Methyl Orange, Orange II, Allura Red (38), 1-phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-naphthalenol, 1-(4-phenylazophenylazo)-2-naphthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, and Para Red (139).
Bifidobacterium longum is used as a probiotic in functional foods (12). The bacterium is acquired from the mother during delivery and habits in the newborn intestine (7). 1-(4-Phenylazophenylazo)-2-naphthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, and Para Red can be reduced by B. longum (139).
Bifidobacterium adolescentis is also a type of probiotic bacteria, and a butanol extract of the bacterium can increase the activity of host immune system (76). B. adolescentis was also studied as a gene delivery system to solid tumors (79). 1-(4-Phenylazophenylazo)-2-naphthalenol, and 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol can be reduced by B. adolescentis (139).
Bifidobacterium infantis as probiotic bacterium has the ability to reduce intestinal inflammation (22, 23). It is reported that B. infantis can decolorize 1-phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-naphthalenol, 1-(4-phenylazophenylazo)-2-nahpthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, and Para Red within 2 days (139).
It is reported that Clostridium species from human gastrointestinal tract have the azoreductase activity and these azoreductases share a similar enzyme structure (106, 107). The enzyme from Clostridium perfringens had the highest azoreductase activity in the test bacteria (105). The azoreductase was purified and addition of FAD enhanced the enzyme activity. The enzyme also reduced the nitro substituent of nitroaromatic compounds (103, 104). McBain and Macfarlane reported a similar finding that culture samples of C. perfringens can reduce amaranth and nitrobenzoic acid (87). C. perfringens also reduced 1-phenylazo-2-naphthol, 1-(4-phenylazophenylazo)-2-naphthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, and Para Red (139).
Clostridium botulinum known for the ability to form botulinum neurotoxin has the similar azoreductase activity to C. perfringens (99). C. botulinum can reduce 1-phenylazo-2-naphthol, 1-(4-phenylazophenylazo)-2-naphthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, and Para Red (139).
Clostridium difficile infection associated with some gastrointestinal diseases (125). C. difficile toxin A caused acute colitis in rats and can be inhibited by a compound contained azo bond, APAZA, which is a molecule of 5-aminosalicylic acid linked to another molecule of 4-aminophenylacetic acid via the azo bond. The reduced product of APAZA was responsible for protection against colitis (89). C. difficile can metabolize azo dyes similar to C. butyricum (139).
Ent. faecalis is commonly isolated from mammalian gastrointestinal tract (56). The azoreductase from Ent. faecalis has a higher specific activity compared to the enzyme from several other intestinal bacteria (133). The azoreductase from Ent. faecalis has been purified and characterized (33). The enzyme is FMN-depended and NADH is a preferred electron donor for its activity. The structure of the enzyme has been studied and the FMN binding site has been characterized (34, 81). The azoreductase from Ent. faecalis (AzoA) was overexpressed in E. coli and the results showed that cytoplasmic azoreductase is involved in the decolorization of dyes in vivo (55). Ent. faecalis has been shown to possess higher azoreductase activity than those of many other intestinal bacteria (133). The bacterium can degrade a broad spectrum of azo dyes, including 1-phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-naphthalenol, 1-(4-phenylazophenylazo)-2-naphthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, Para Red, Methyl Red, Orange II, Orange G, Amaranth, Ponceau BS, and Ponceau S (33, 138, 139).
Enterococcus faecium is an intestinal bacterium that also has been isolated from patients with severe infection in hospital and possesses a broad spectrum of antibiotic resistance (73). Interestingly, some strains of Ent. faecium are gaining biotechnological interest since they may produce bacteriocins (77). The azoreductase from Ent. faecium was purified, and shared 67% identity with the enzyme (AzoA) from Ent. faecalis (83). Like AzoA, the enzyme also possesses a broad substrate spectrum of azoreductase activity (83). Ent. faecium can metabolize 1-phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-naphthalenol, 1-(4-phenylazophenylazo)-2-naphthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, and Para Red (139).
The azoreductase gene of E. coli was annotated first as an acyl carrier protein phosphodiesterase (acpD) gene. The gene product was purified, and it was found that the enzyme can not convert holo-acyl carrier protein into the apo form, and instead the azoreductase activity was identified, therefore, the acpD gene was redesigned as azoR (95). AzoR is a FMN-dependent and NADH-preferred enzyme, and catalyzes the reduction of azo dyes by ping-pong mechanism. The crystal structure of AzoR with FMN has been reported (69). The E. coli strain contained the overexpressed AzoR has a similar decolorization activity at aerobic and anaerobic conditions (Unpublished data), which means the azoreductase from E. coli is oxygen tolerant. Methyl Red, Ponceau BS, and Orange II can be reduced by E. coli (55, 95).
Lactobacillus paracasei is a facultative anaerobic bacteria found in intestinal tracts. It is reported that this bacterium can metabolize azo dyes under anaerobic conditions (101). The bacterium can cause red-color spoilage in cucumber pickle products when the azo dye Tartrazine was used as yellow coloring (100). 1-Phenylazo-2-naphthol, 1-[(2,4-dimethylphenyl)azo]-2-naphthalenol, 1-(4-phenylazophenylazo)-2-naphthalenol, 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol, Para Red, Methyl Orange, Acid Orange 8, Tartrazine, and Acid Orange 63 can be reduced by the bacterium (101, 139).
6. SUMMARY
Azo dyes are used widely in consumer products. Human intestinal microbiota play a key role in the metabolism of azo dyes and azoreductase from intestinal bacteria can reduce the azo bond of the dyes via NAD(P)H. There are three groups of azoreductases: flavin dependent NADH preferred azoreductase, such as the enzymes from Ent. faecalis, Ent. faecium, and E. coli; flavin dependent NADPH preferred azoreductase, such as the enzymes from S. aureus, B. cereus, and B. subtilis; and flavin free NADPH preferred azoreductase, such as the enzymes from P. kullae and X. azovorans. A portion of the intestinal microbiota community can metabolize both water-soluble and water-insoluble azo dyes. Some of metabolites of the dyes produced by these microorganisms are carcinogenic, for example, o-toluidine and benzene are two of the metabolites of 1-[[2-methyl-4-[(2-methylphenyl)azo]phenyl]azo]-2-naphthalenol and Congo Red, respectively; and they are carcinogenic to humans and classed as group 1 by IARC. Recently, we demonstrated that human microbiota can reduce Sudan dyes to form toxic aromatic amines, which indicates the potential risk to consume food products contaminated with these dyes. Further studies on the enzymatic reduction mechanisms of both water-soluble and water-insoluble azo dyes by human intestinal microbiota along with the potential to produce genotoxic aromatic compounds are needed.
Acknowledgments
We thank Drs. Steven L. Foley and Huanli Liu for their critical review of the manuscript. This study was funded by the National Center for Toxicological Research, United States Food and Drug Administration, and supported in part by an appointment (J.F.) to the Postgraduate Research Fellowship Program by the Oak Ridge Institute for Science and Education through an interagency agreement between the U.S. Department of Energy and the U.S. Food and Drug Administration. The views presented in this article do not necessarily reflect those of the Food and Drug Administration.
References
- 1.Bioassay of p-phenylenediamine dihydrochloride for possible carcinogenicity. Natl Cancer Inst Carcinog Tech Rep Ser. 1979;174:1–107. [PubMed] [Google Scholar]
- 2.IARC monographs on the evaluation of the carcinogenic risk of chemicals to man: some aziridines, N-, S- & O-mustards and selenium. IARC Monogr Eval Carcinog Risk Chem Man. 1975;9:1–268. [PubMed] [Google Scholar]
- 3.NTP toxicology and carcinogenesis studies of p-nitroaniline (CAS No. 100-01-6) in B6C3F1 mice (gavage studies) Natl Toxicol Program Tech Rep Ser. 1993;418:1–203. [PubMed] [Google Scholar]
- 4.Ortho-toluidine. IARC Monogr Eval Carcinog Risks Hum. 2000;77:267–322. [PMC free article] [PubMed] [Google Scholar]
- 5.Overall evaluations of carcinogenicity: an updating of IARC Monographs volumes 1 to 42. IARC Monogr Eval Carcinog Risks Hum Suppl. 1987;7:1–440. [PubMed] [Google Scholar]
- 6.Ahmed R, Gray D. Immunological Memory and Protective Immunity: Understanding Their Relation. Science. 1996;272:54–60. doi: 10.1126/science.272.5258.54. [DOI] [PubMed] [Google Scholar]
- 7.Aires J, Anglade P, Baraige F, Zagorec M, Champomier-Verges MC, Butel MJ. Proteomic comparison of the cytosolic proteins of three Bifidobacterium longum human isolates and B. longum NCC2705. BMC Microbiol. 2010;10:29. doi: 10.1186/1471-2180-10-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Anfossi L, Baggiani C, Giovannoli C, Giraudi G. Development of enzyme-linked immunosorbent assays for Sudan dyes in chilli powder, ketchup and egg yolk. Food Addit Contam Part A Chem Anal Control Expo Risk Assess. 2009;26:800–7. doi: 10.1080/02652030902755283. [DOI] [PubMed] [Google Scholar]
- 9.Anuradha S, Arora S, Mehrotra S, Arora A, Kar P. Acute renal failure following para-phenylenediamine (PPD) poisoning: a case report and review. Ren Fail. 2004;26:329–32. doi: 10.1081/jdi-200026722. [DOI] [PubMed] [Google Scholar]
- 10.Ashby J, Paton D, Lefevre PA, Styles JA, Rose FL. Evaluation of two suggested methods of deactivating organic carcinogens by molecular modification. Carcinogenesis. 1982;3:1277–82. doi: 10.1093/carcin/3.11.1277. [DOI] [PubMed] [Google Scholar]
- 11.ATSDR. Toxicological Profile for Benzidine. 2001 [PubMed] [Google Scholar]
- 12.Audy J, Labrie S, Roy D, Lapointe G. Sugar source modulates exopolysaccharide biosynthesis in Bifidobacterium longum subsp. longum CRC 002. Microbiology. 2010;156:653–64. doi: 10.1099/mic.0.033720-0. [DOI] [PubMed] [Google Scholar]
- 13.Ben Mansour H, Barillier D, Corroler D, Ghedira K, Chekir-Ghedira L, Mosrati R. In vitro study of DNA damage induced by acid orange 52 and its biodegradation derivatives. Environ Toxicol Chem. 2009;28:489–95. doi: 10.1897/08-333.1. [DOI] [PubMed] [Google Scholar]
- 14.Ben Mansour H, Corroler D, Barillier D, Ghedira K, Chekir L, Mosrati R. Evaluation of genotoxicity and pro-oxidant effect of the azo dyes: acids yellow 17, violet 7 and orange 52, and of their degradation products by Pseudomonas putida mt-2. Food Chem Toxicol. 2007;45:1670–7. doi: 10.1016/j.fct.2007.02.033. [DOI] [PubMed] [Google Scholar]
- 15.Bik EM. Composition and function of the human-associated microbiota. Nutr Rev. 2009;67(Suppl 2):S164–71. doi: 10.1111/j.1753-4887.2009.00237.x. [DOI] [PubMed] [Google Scholar]
- 16.Bin Y, Jiti Z, Jing W, Cuihong D, Hongman H, Zhiyong S, Yongming B. Expression and characteristics of the gene encoding azoreductase from Rhodobacter sphaeroides AS1.1737. FEMS Microbiol Lett. 2004;236:129–36. doi: 10.1016/j.femsle.2004.05.034. [DOI] [PubMed] [Google Scholar]
- 17.Bjorkholm B, Bok CM, Lundin A, Rafter J, Hibberd ML, Pettersson S. Intestinal microbiota regulate xenobiotic metabolism in the liver. PLoS One. 2009;4:e6958. doi: 10.1371/journal.pone.0006958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Blumel S, Knackmuss HJ, Stolz A. Molecular cloning and characterization of the gene coding for the aerobic azoreductase from Xenophilus azovorans KF46F. Appl Environ Microbiol. 2002;68:3948–55. doi: 10.1128/AEM.68.8.3948-3955.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Blumel S, Stolz A. Cloning and characterization of the gene coding for the aerobic azoreductase from Pigmentiphaga kullae K24. Appl Microbiol Biotechnol. 2003;62:186–90. doi: 10.1007/s00253-003-1316-5. [DOI] [PubMed] [Google Scholar]
- 20.Bolt HM, Golka K. The debate on carcinogenicity of permanent hair dyes: new insights. Crit Rev Toxicol. 2007;37:521–36. doi: 10.1080/10408440701385671. [DOI] [PubMed] [Google Scholar]
- 21.Bomhard EM, Herbold BA. Genotoxic activities of aniline and its metabolites and their relationship to the carcinogenicity of aniline in the spleen of rats. Crit Rev Toxicol. 2005;35:783–835. doi: 10.1080/10408440500442384. [DOI] [PubMed] [Google Scholar]
- 22.Brenner DM, Chey WD. Bifidobacterium infantis 35624: a novel probiotic for the treatment of irritable bowel syndrome. Rev Gastroenterol Disord. 2009;9:7–15. [PubMed] [Google Scholar]
- 23.Brenner DM, Moeller MJ, Chey WD, Schoenfeld PS. The utility of probiotics in the treatment of irritable bowel syndrome: a systematic review. Am J Gastroenterol. 2009;104:1033–49. doi: 10.1038/ajg.2009.25. quiz 1050. [DOI] [PubMed] [Google Scholar]
- 24.Brigé A, Motte B, Borloo J, Buysschaert G, Devreese B, Van Beeumen JJ. Bacterial decolorization of textile dyes is an extracellular process requiring a multicomponent electron transfer pathway. Microbial Biotechnology. 2008;1:40–52. doi: 10.1111/j.1751-7915.2007.00005.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Brown JP, Dietrich PS. Mutagenicity of selected sulfonated azo dyes in the Salmonella/microsome assay: use of aerobic and anaerobic activation procedures. Mutat Res. 1983;116:305–15. doi: 10.1016/0165-1218(83)90068-x. [DOI] [PubMed] [Google Scholar]
- 26.Brown JP, Roehm GW, Brown RJ. Mutagenicity testing of certified food colors and related azo, xanthene and triphenylmethane dyes with the Salmonella/microsome system. Mutat Res. 1978;56:249–71. doi: 10.1016/0027-5107(78)90192-6. [DOI] [PubMed] [Google Scholar]
- 27.Brown MA, De Vito SC. Predicting azo dye toxicity. Critical Reviews in Environmental Science and Technology. 1993;23:249–324. [Google Scholar]
- 28.Calbiani F, Careri M, Elviri L, Mangia A, Zagnoni I. Accurate mass measurements for the confirmation of Sudan azo-dyes in hot chilli products by capillary liquid chromatography-electrospray tandem quadrupole orthogonal-acceleration time of flight mass spectrometry. J Chromatogr A. 2004;1058:127–35. [PubMed] [Google Scholar]
- 29.Carreon T, Hein MJ, Viet SM, Hanley KW, Ruder AM, Ward EM. Increased bladder cancer risk among workers exposed to o-toluidine and aniline: a reanalysis. Occup Environ Med. 2010;67:348–50. doi: 10.1136/oem.2009.051136. [DOI] [PubMed] [Google Scholar]
- 30.Chen H. Recent advances in azo dye degrading enzyme research. Curr Protein Pept Sci. 2006;7:101–11. doi: 10.2174/138920306776359786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Chen H, Feng J, Kweon O, Xu H, Cerniglia C. Identification and molecular characterization of a novel flavin-free NADPH preferred azoreductase encoded by azoB in Pigmentiphaga kullae K24. BMC Biochemistry. 2010;11:13. doi: 10.1186/1471-2091-11-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Chen H, Hopper SL, Cerniglia CE. Biochemical and molecular characterization of an azoreductase from Staphylococcus aureus, a tetrameric NADPH-dependent flavoprotein. Microbiology. 2005;151:1433–41. doi: 10.1099/mic.0.27805-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chen H, Wang RF, Cerniglia CE. Molecular cloning, overexpression, purification, and characterization of an aerobic FMN-dependent azoreductase fromEnterococcus faecalis. Protein Expr Purif. 2004;34:302–10. doi: 10.1016/j.pep.2003.12.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Chen H, Xu H, Kweon O, Chen S, Cerniglia CE. Functional role of Trp-105 of Enterococcus faecalis azoreductase (AzoA) as resolved by structural and mutational analysis. Microbiology. 2008;154:2659–67. doi: 10.1099/mic.0.2008/019877-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Chen Y, Kinouchi T, Kataoka K, Akimoto S, Ohnishi Y. Purification and characterization of a fibrinogen-degrading protease in Bacteroides fragilis strain YCH46. Microbiol Immunol. 1995;39:967–77. doi: 10.1111/j.1348-0421.1995.tb03300.x. [DOI] [PubMed] [Google Scholar]
- 36.Chung KT. The significance of azo-reduction in the mutagenesis and carcinogenesis of azo dyes. Mutat Res. 1983;114:269–81. doi: 10.1016/0165-1110(83)90035-0. [DOI] [PubMed] [Google Scholar]
- 37.Chung KT, Fulk GE, Andrews AW. Mutagenicity testing of some commonly used dyes. Appl Environ Microbiol. 1981;42:641–8. doi: 10.1128/aem.42.4.641-648.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Chung KT, Fulk GE, Egan M. Reduction of azo dyes by intestinal anaerobes. Appl Environ Microbiol. 1978;35:558–62. doi: 10.1128/aem.35.3.558-562.1978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Chung KT, Stevens SE, Jr, Cerniglia CE. The reduction of azo dyes by the intestinal microflora. Crit Rev Microbiol. 1992;18:175–90. doi: 10.3109/10408419209114557. [DOI] [PubMed] [Google Scholar]
- 40.Collier SW, Storm JE, Bronaugh RL. Reduction of azo dyes during in vitro percutaneous absorption. Toxicol Appl Pharmacol. 1993;118:73–9. doi: 10.1006/taap.1993.1011. [DOI] [PubMed] [Google Scholar]
- 41.Combes RD, Haveland-Smith RB. A review of the genotoxicity of food, drug and cosmetic colours and other azo, triphenylmethane and xanthene dyes. Mutat Res. 1982;98:101–248. doi: 10.1016/0165-1110(82)90015-x. [DOI] [PubMed] [Google Scholar]
- 42.D’Elia JN, Salyers AA. Effect of regulatory protein levels on utilization of starch byBacteroides thetaiotaomicron. J Bacteriol. 1996;178:7180–6. doi: 10.1128/jb.178.24.7180-7186.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Dafale N, Rao NN, Meshram SU, Wate SR. Decolorization of azo dyes and simulated dye bath wastewater using acclimatized microbial consortium--biostimulation and halo tolerance. Bioresour Technol. 2008;99:2552–8. doi: 10.1016/j.biortech.2007.04.044. [DOI] [PubMed] [Google Scholar]
- 44.Danford N. The genetic toxicology of ortho-toluidine. Mutat Res. 1991;258:207–36. doi: 10.1016/0165-1110(91)90010-s. [DOI] [PubMed] [Google Scholar]
- 45.De Flora S, Camoirano A, Zanacchi P, Bennicelli C. Mutagenicity testing with TA97 and TA102 of 30 DNA-damaging compounds, negative with other Salmonella strains. Mutat Res. 1984;134:159–65. doi: 10.1016/0165-1110(84)90009-5. [DOI] [PubMed] [Google Scholar]
- 46.De Flora S, Zanacchi P, Camoirano A, Bennicelli C, Badolati GS. Genotoxic activity and potency of 135 compounds in the Ames reversion test and in a bacterial DNA-repair test. Mutat Res. 1984;133:161–98. doi: 10.1016/0165-1110(84)90016-2. [DOI] [PubMed] [Google Scholar]
- 47.Degawa M, Kanazawa C, Hashimoto Y. In vitro metabolism of o-aminoazotoluene and mutagenesis of Salmonella by the metabolites. Carcinogenesis. 1982;3:1113–7. doi: 10.1093/carcin/3.10.1113. [DOI] [PubMed] [Google Scholar]
- 48.Delclos KB, Tarpley WG, Miller EC, Miller JA. 4-aminoazobenzene and N,N-dimethyl-4-aminoazobenzene as equipotent hepatic carcinogens in male C57BL/6 X C3H/He F1 mice and characterization of N-(Deoxyguanosin-8-yl)-4-aminoazobenzene as the major persistent hepatic DNA-bound dye in these mice. Cancer Res. 1984;44:2540–50. [PubMed] [Google Scholar]
- 49.Dethlefsen L, McFall-Ngai M, Relman DA. An ecological and evolutionary perspective on human-microbe mutualism and disease. Nature. 2007;449:811–8. doi: 10.1038/nature06245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Dillon D, Combes R, Zeiger E. Activation by caecal reduction of the azo dye D & C red no. 9 to a bacterial mutagen. Mutagenesis. 1994;9:295–9. doi: 10.1093/mutage/9.4.295. [DOI] [PubMed] [Google Scholar]
- 51.Duchmann R, Kaiser I, Hermann E, Mayet W, Ewe K, Meyer zum Buschenfelde KH. Tolerance exists towards resident intestinal flora but is broken in active inflammatory bowel disease (IBD) Clin Exp Immunol. 1995;102:448–55. doi: 10.1111/j.1365-2249.1995.tb03836.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Eckburg PB, Bik EM, Bernstein CN, Purdom E, Dethlefsen L, Sargent M, Gill SR, Nelson KE, Relman DA. Diversity of the human intestinal microbial flora. Science. 2005;308:1635–8. doi: 10.1126/science.1110591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Elhkim MO, Heraud F, Bemrah N, Gauchard F, Lorino T, Lambre C, Fremy JM, Poul JM. New considerations regarding the risk assessment on Tartrazine An update toxicological assessment, intolerance reactions and maximum theoretical daily intake in France. Regul Toxicol Pharmacol. 2007;47:308–16. doi: 10.1016/j.yrtph.2006.11.004. [DOI] [PubMed] [Google Scholar]
- 54.Fassina G, Abbondandolo A, Mariani L, Taningher M, Parodi S. Mutagenicity in V79 cells does not correlate with carcinogenity in small rodents for 12 aromatic amines. J Toxicol Environ Health. 1990;29:109–30. doi: 10.1080/15287399009531376. [DOI] [PubMed] [Google Scholar]
- 55.Feng J, Heinze TM, Xu H, Cerniglia CE, Chen H. Evidence for significantly enhancing reduction of Azo dyes in Escherichia coli by expressed cytoplasmic Azoreductase (AzoA) ofEnterococcus faecalis. Protein Pept Lett. 2010;17:578–84. doi: 10.2174/092986610791112701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Fisher K, Phillips C. The ecology, epidemiology and virulence ofEnterococcus. Microbiology. 2009;155:1749–57. doi: 10.1099/mic.0.026385-0. [DOI] [PubMed] [Google Scholar]
- 57.Flint HJ, Bayer EA, Rincon MT, Lamed R, White BA. Polysaccharide utilization by gut bacteria: potential for new insights from genomic analysis. Nat Rev Microbiol. 2008;6:121–31. doi: 10.1038/nrmicro1817. [DOI] [PubMed] [Google Scholar]
- 58.Garrigue JL, Ballantyne M, Kumaravel T, Lloyd M, Nohynek GJ, Kirkland D, Toutain H. In vitro genotoxicity of para-phenylenediamine and its N-monoacetyl or N,N′-diacetyl metabolites. Mutat Res. 2006;608:58–71. doi: 10.1016/j.mrgentox.2006.05.001. [DOI] [PubMed] [Google Scholar]
- 59.Golka K, Kopps S, Myslak ZW. Carcinogenicity of azo colorants: influence of solubility and bioavailability. Toxicol Lett. 2004;151:203–10. doi: 10.1016/j.toxlet.2003.11.016. [DOI] [PubMed] [Google Scholar]
- 60.Gopinath KP, Asan Meera Sahib H, Muthukumar K, Velan M. Improved biodegradation of Congored by using Bacillus sp. Bioresour Technol. 2009;100:670–675. doi: 10.1016/j.biortech.2008.07.048. [DOI] [PubMed] [Google Scholar]
- 61.Gottlieb A, Shaw C, Smith A, Wheatley A, Forsythe S. The toxicity of textile reactive azo dyes after hydrolysis and decolourisation. J Biotechnol. 2003;101:49–56. doi: 10.1016/s0168-1656(02)00302-4. [DOI] [PubMed] [Google Scholar]
- 62.Haley TJ. Benzidine revisited: a review of the literature and problems associated with the use of benzidine and its congeners. Clin Toxicol. 1975;8:13–42. doi: 10.3109/15563657508988044. [DOI] [PubMed] [Google Scholar]
- 63.Han D, Yu M, Knopp D, Niessner R, Wu M, Deng A. Development of a highly sensitive and specific enzyme-linked immunosorbent assay for detection of Sudan I in food samples. J Agric Food Chem. 2007;55:6424–30. doi: 10.1021/jf071005j. [DOI] [PubMed] [Google Scholar]
- 64.Han J, Antunes LC, Finlay BB, Borchers CH. Metabolomics: towards understanding host-microbe interactions. Future Microbiol. 2010;5:153–61. doi: 10.2217/fmb.09.132. [DOI] [PubMed] [Google Scholar]
- 65.He L, Su Y, Fang B, Shen X, Zeng Z, Liu Y. Determination of Sudan dye residues in eggs by liquid chromatography and gas chromatography-mass spectrometry. Anal Chim Acta. 2007;594:139–46. doi: 10.1016/j.aca.2007.05.021. [DOI] [PubMed] [Google Scholar]
- 66.Hill MJ. Intestinal flora and endogenous vitamin synthesis. Eur J Cancer Prev. 1997;6(Suppl 1):S43–5. doi: 10.1097/00008469-199703001-00009. [DOI] [PubMed] [Google Scholar]
- 67.Hooper LV, Gordon JI. Commensal Host-Bacterial Relationships in the Gut. Science. 2001;292:1115–1118. doi: 10.1126/science.1058709. [DOI] [PubMed] [Google Scholar]
- 68.Hooper LV, Macpherson AJ. Immune adaptations that maintain homeostasis with the intestinal microbiota. Nat Rev Immunol. 2010;10:159–169. doi: 10.1038/nri2710. [DOI] [PubMed] [Google Scholar]
- 69.Ito K, Nakanishi M, Lee WC, Sasaki H, Zenno S, Saigo K, Kitade Y, Tanokura M. Three-dimensional structure of AzoR from Escherichia coli. An oxidereductase conserved in microorganisms. J Biol Chem. 2006;281:20567–76. doi: 10.1074/jbc.M513345200. [DOI] [PubMed] [Google Scholar]
- 70.Ito K, Nakanishi M, Lee WC, Zhi Y, Sasaki H, Zenno S, Saigo K, Kitade Y, Tanokura M. Expansion of substrate specificity and catalytic mechanism of azoreductase by X-ray crystallography and site-directed mutagenesis. J Biol Chem. 2008;283:13889–96. doi: 10.1074/jbc.M710070200. [DOI] [PubMed] [Google Scholar]
- 71.Jadhav JP, Parshetti GK, Kalme SD, Govindwar SP. Decolourization of azo dye methyl red by Saccharomyces cerevisiae MTCC 463. Chemosphere. 2007;68:394–400. doi: 10.1016/j.chemosphere.2006.12.087. [DOI] [PubMed] [Google Scholar]
- 72.Kawakami T, Isama K, Nakashima H, Tsuchiya T, Matsuoka A. Analysis of primary aromatic amines originated from azo dyes in commercial textile products in Japan. J Environ Sci Health A Tox Hazard Subst Environ Eng. 2010;45:1281–95. doi: 10.1080/10934529.2010.493827. [DOI] [PubMed] [Google Scholar]
- 73.Klare I, Konstabel C, Badstubner D, Werner G, Witte W. Occurrence and spread of antibiotic resistances inEnterococcus faecium. Int J Food Microbiol. 2003;88:269–90. doi: 10.1016/s0168-1605(03)00190-9. [DOI] [PubMed] [Google Scholar]
- 74.Kolekar YM, Pawar SP, Gawai KR, Lokhande PD, Shouche YS, Kodam KM. Decolorization and degradation of Disperse Blue 79 and Acid Orange 10, by Bacillus fusiformis KMK5 isolated from the textile dye contaminated soil. Bioresour Technol. 2008;99:8999–9003. doi: 10.1016/j.biortech.2008.04.073. [DOI] [PubMed] [Google Scholar]
- 75.Lazear EJ, Louie SC. Mutagenicity of some congeners of benzidine in the Salmonella typhimurium assay system. Cancer Lett. 1978;4:21–5. doi: 10.1016/s0304-3835(78)93137-3. [DOI] [PubMed] [Google Scholar]
- 76.Lee do K, Jang S, Kim MJ, Kim JH, Chung MJ, Kim KJ, Ha NJ. Anti-proliferative effects of Bifidobacterium adolescentis SPM0212 extract on human colon cancer cell lines. BMC Cancer. 2008;8:310. doi: 10.1186/1471-2407-8-310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Leroy F, Foulquie Moreno MR, De Vuyst L. Enterococcus faecium RZS C5, an interesting bacteriocin producer to be used as a co-culture in food fermentation. Int J Food Microbiol. 2003;88:235–40. doi: 10.1016/s0168-1605(03)00185-5. [DOI] [PubMed] [Google Scholar]
- 78.Levine WG. Metabolism of azo dyes: implication for detoxication and activation. Drug Metab Rev. 1991;23:253–309. doi: 10.3109/03602539109029761. [DOI] [PubMed] [Google Scholar]
- 79.Li X, Fu GF, Fan YR, Liu WH, Liu XJ, Wang JJ, Xu GX. Bifidobacterium adolescentis as a delivery system of endostatin for cancer gene therapy: selective inhibitor of angiogenesis and hypoxic tumor growth. Cancer Gene Ther. 2003;10:105–11. doi: 10.1038/sj.cgt.7700530. [DOI] [PubMed] [Google Scholar]
- 80.Liger D, Graille M, Zhou CZ, Leulliot N, Quevillon-Cheruel S, Blondeau K, Janin J, van Tilbeurgh H. Crystal structure and functional characterization of yeast YLR011wp, an enzyme with NAD(P)H-FMN and ferric iron reductase activities. J Biol Chem. 2004;279:34890–7. doi: 10.1074/jbc.M405404200. [DOI] [PubMed] [Google Scholar]
- 81.Liu ZJ, Chen H, Shaw N, Hopper SL, Chen L, Chen S, Cerniglia CE, Wang BC. Crystal structure of an aerobic FMN-dependent azoreductase (AzoA) fromEnterococcus faecalis. Arch Biochem Biophys. 2007;463:68–77. doi: 10.1016/j.abb.2007.03.003. [DOI] [PubMed] [Google Scholar]
- 82.Macpherson A, Khoo UY, Forgacs I, Philpott-Howard J, Bjarnason I. Mucosal antibodies in inflammatory bowel disease are directed against intestinal bacteria. Gut. 1996;38:365–75. doi: 10.1136/gut.38.3.365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Macwana SR, Punj S, Cooper J, Schwenk E, John GH. Identification and Isolation of an Azoreductase fromEnterococcus faecium. Curr Issues Mol Biol. 2009;12:43–48. [PubMed] [Google Scholar]
- 84.Mamber SW, Bryson V, Katz SE. The Escherichia coli WP2/WP100 rec assay for detection of potential chemical carcinogens. Mutat Res. 1983;119:135–44. doi: 10.1016/0165-7992(83)90121-5. [DOI] [PubMed] [Google Scholar]
- 85.Mansour HB, Mosrati R, Corroler D, Ghedira K, Barillier D, Chekir-Ghedira L. Mutagenicity and genotoxicity of acid yellow 17 and its biodegradation products. Drug Chem Toxicol. 2009;32:222–9. doi: 10.1080/01480540902862269. [DOI] [PubMed] [Google Scholar]
- 86.Marhold J, Rambousek V, Pipalova J, Matrka M. Oxidation of carcinogenic azo-dyes. VII. Metabolites of some 4′-substituted derivatives of dimethyl-aminoazobenzene in the bile of rats. Neoplasma. 1969;16:191–4. [PubMed] [Google Scholar]
- 87.McBain AJ, Macfarlane GT. Ecological and physiological studies on large intestinal bacteria in relation to production of hydrolytic and reductive enzymes involved in formation of genotoxic metabolites. J Med Microbiol. 1998;47:407–16. doi: 10.1099/00222615-47-5-407. [DOI] [PubMed] [Google Scholar]
- 88.McCarroll NE, Piper CE, Keech BH. An E coli microsuspension assay for the detection of DNA damage induced by direct-acting agents and promutagens. Environ Mutagen. 1981;3:429–44. doi: 10.1002/em.2860030404. [DOI] [PubMed] [Google Scholar]
- 89.McVey DC, Liddle RA, Riggs-Sauthier J, Ekwuribe N, Vigna SR. Inhibition of Clostridium difficile toxin A-induced colitis in rats by APAZA. Dig Dis Sci. 2005;50:565–73. doi: 10.1007/s10620-005-2476-1. [DOI] [PubMed] [Google Scholar]
- 90.Moller P, Wallin H. Genotoxic hazards of azo pigments and other colorants related to 1-phenylazo-2-hydroxynaphthalene. Mutat Res. 2000;462:13–30. doi: 10.1016/s1383-5742(99)00090-3. [DOI] [PubMed] [Google Scholar]
- 91.Moncrief JS, Obiso R, Jr, Barroso LA, Kling JJ, Wright RL, Van Tassell RL, Lyerly DM, Wilkins TD. The enterotoxin of Bacteroides fragilis is a metalloprotease. Infect Immun. 1995;63:175–81. doi: 10.1128/iai.63.1.175-181.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Moore WE, Holdeman LV. Human fecal flora: the normal flora of 20 Japanese-Hawaiians. Appl Microbiol. 1974;27:961–79. doi: 10.1128/am.27.5.961-979.1974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Morokutti A, Lyskowski A, Sollner S, Pointner E, Fitzpatrick TB, Kratky C, Gruber K, Macheroux P. Structure and function of YcnD from Bacillus subtilis, a flavin-containing oxidoreductase. Biochemistry. 2005;44:13724–33. doi: 10.1021/bi0510835. [DOI] [PubMed] [Google Scholar]
- 94.Muyzer G, de Waal EC, Uitterlinden AG. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol. 1993;59:695–700. doi: 10.1128/aem.59.3.695-700.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Nakanishi M, Yatome C, Ishida N, Kitade Y. Putative ACP phosphodiesterase gene (acpD) encodes an azoreductase. J Biol Chem. 2001;276:46394–9. doi: 10.1074/jbc.M104483200. [DOI] [PubMed] [Google Scholar]
- 96.Nelson AA, Woodard G. Tumors of the urinary bladder, gall bladder, and liver in dogs fed o-aminoazotoluene or p-dimethylaminoazobenzene. J Natl Cancer Inst. 1953;13:1497–1509. [PubMed] [Google Scholar]
- 97.Osano O, Admiraal W, Klamer HJ, Pastor D, Bleeker EA. Comparative toxic and genotoxic effects of chloroacetanilides, formamidines and their degradation products on Vibrio fischeri andChironomus riparius. Environ Pollut. 2002;119:195–202. doi: 10.1016/s0269-7491(01)00334-7. [DOI] [PubMed] [Google Scholar]
- 98.Osano O, Oladimeji AA, Kraak MH, Admiraal W. Teratogenic effects of amitraz, 2,4-dimethylaniline, and paraquat on developing frog (Xenopus) embryos. Arch Environ Contam Toxicol. 2002;43:42–9. doi: 10.1007/s00244-002-1132-4. [DOI] [PubMed] [Google Scholar]
- 99.Peck MW. Biology and genomic analysis ofClostridium botulinum. Adv Microb Physiol. 2009;55:183–265. 320. doi: 10.1016/S0065-2911(09)05503-9. [DOI] [PubMed] [Google Scholar]
- 100.Perez-Diaz IM, Kelling RE, Hale S, Breidt F, McFeeters RF. Lactobacilli and tartrazine as causative agents of red-color spoilage in cucumber pickle products. J Food Sci. 2007;72:M240–5. doi: 10.1111/j.1750-3841.2007.00460.x. [DOI] [PubMed] [Google Scholar]
- 101.Perez-Diaz IM, McFeeters RF. Modification of azo dyes by lactic acid bacteria. J Appl Microbiol. 2009;107:584–9. doi: 10.1111/j.1365-2672.2009.04227.x. [DOI] [PubMed] [Google Scholar]
- 102.Prival MJ, Bell SJ, Mitchell VD, Peiperl MD, Vaughan VL. Mutagenicity of benzidine and benzidine-congener dyes and selected monoazo dyes in a modified Salmonella assay. Mutat Res. 1984;136:33–47. doi: 10.1016/0165-1218(84)90132-0. [DOI] [PubMed] [Google Scholar]
- 103.Rafii F, Cerniglia CE. Comparison of the azoreductase and nitroreductase fromClostridium perfringens. Appl Environ Microbiol. 1993;59:1731–4. doi: 10.1128/aem.59.6.1731-1734.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Rafii F, Cerniglia CE. Reduction of azo dyes and nitroaromatic compounds by bacterial enzymes from the human intestinal tract. Environ Health Perspect. 1995;103(Suppl 5):17–9. doi: 10.1289/ehp.95103s417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Rafii F, Franklin W, Cerniglia CE. Azoreductase activity of anaerobic bacteria isolated from human intestinal microflora. Appl Environ Microbiol. 1990;56:2146–51. doi: 10.1128/aem.56.7.2146-2151.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Rafii F, Hall JD, Cerniglia CE. Mutagenicity of azo dyes used in foods, drugs and cosmetics before and after reduction by Clostridium species from the human intestinal tract. Food Chem Toxicol. 1997;35:897–901. doi: 10.1016/s0278-6915(97)00060-4. [DOI] [PubMed] [Google Scholar]
- 107.Rafii F, Smith DB, Benson RW, Cerniglia CE. Immunological homology among azoreductases from Clostridium and Eubacterium strains isolated from human intestinal microflora. J Basic Microbiol. 1992;32:99–105. doi: 10.1002/jobm.3620320204. [DOI] [PubMed] [Google Scholar]
- 108.Rebane R, Leito I, Yurchenko S, Herodes K. A review of analytical techniques for determination of Sudan I–IV dyes in food matrixes. J Chromatogr A. 2010;1217:2747–57. doi: 10.1016/j.chroma.2010.02.038. [DOI] [PubMed] [Google Scholar]
- 109.Reeves AR, Wang GR, Salyers AA. Characterization of four outer membrane proteins that play a role in utilization of starch byBacteroides thetaiotaomicron. J Bacteriol. 1997;179:643–9. doi: 10.1128/jb.179.3.643-649.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Robinson T, McMullan G, Marchant R, Nigam P. Remediation of dyes in textile effluent: a critical review on current treatment technologies with a proposed alternative. Bioresour Technol. 2001;77:247–55. doi: 10.1016/s0960-8524(00)00080-8. [DOI] [PubMed] [Google Scholar]
- 111.Rosenkranz HS, Klopman G. Structural basis of the mutagenicity of 1-amino-2-naphthol-based azo dyes. Mutagenesis. 1990;5:137–46. doi: 10.1093/mutage/5.2.137. [DOI] [PubMed] [Google Scholar]
- 112.Ruseler-van Embden JG, van der Helm R, van Lieshout LM. Degradation of intestinal glycoproteins by Bacteroides vulgatus. FEMS Microbiol Lett. 1989;49:37–41. doi: 10.1016/0378-1097(89)90338-8. [DOI] [PubMed] [Google Scholar]
- 113.Ryan A, Laurieri N, Westwood I, Wang CJ, Lowe E, Sim E. A novel mechanism for azoreduction. J Mol Biol. 2010;400:24–37. doi: 10.1016/j.jmb.2010.04.023. [DOI] [PubMed] [Google Scholar]
- 114.Saitoh S, Noda S, Aiba Y, Takagi A, Sakamoto M, Benno Y, Koga Y. Bacteroides ovatus as the predominant commensal intestinal microbe causing a systemic antibody response in inflammatory bowel disease. Clin Diagn Lab Immunol. 2002;9:54–9. doi: 10.1128/CDLI.9.1.54-59.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Sakagami Y, Yamazaki H, Ogasawara N, Yokoyama H, Ose Y, Sato T. The evaluation of genotoxic activities of disinfectants and their metabolites by umu test. Mutat Res. 1988;209:155–60. doi: 10.1016/0165-7992(88)90034-6. [DOI] [PubMed] [Google Scholar]
- 116.Sakamoto M, Benno Y. Reclassification of Bacteroides distasonis, Bacteroides goldsteinii and Bacteroides merdae as Parabacteroides distasonis gen. nov., comb. nov., Parabacteroides goldsteinii comb. nov. and Parabacteroides merdae comb. nov. Int J Syst Evol Microbiol. 2006;56:1599–605. doi: 10.1099/ijs.0.64192-0. [DOI] [PubMed] [Google Scholar]
- 117.Samejima K, Tamura Z, Ishidate M. Metabolism of 4-dimethylaminoazobenzene and related compounds. IV. Metabolites of o-aminoazotoluene in rat bile. Chem Pharm Bull (Tokyo) 1967;15:964–75. doi: 10.1248/cpb.15.964. [DOI] [PubMed] [Google Scholar]
- 118.Sandhu P, Chipman JK. Bacterial mutagenesis and hepatocyte unscheduled DNA synthesis induced by chrysoidine azo-dye components. Mutat Res. 1990;240:227–36. doi: 10.1016/0165-1218(90)90062-7. [DOI] [PubMed] [Google Scholar]
- 119.Saratale RG, Saratale GD, Chang JS, Govindwar SP. Decolorization and biodegradation of textile dye Navy blue HER by Trichosporon beigelii NCIM-3326. J Hazard Mater. 2009;166:1421–8. doi: 10.1016/j.jhazmat.2008.12.068. [DOI] [PubMed] [Google Scholar]
- 120.Saratale RG, Saratale GD, Kalyani DC, Chang JS, Govindwar SP. Enhanced decolorization and biodegradation of textile azo dye Scarlet R by using developed microbial consortium-GR. Bioresour Technol. 2009;100:2493–2500. doi: 10.1016/j.biortech.2008.12.013. [DOI] [PubMed] [Google Scholar]
- 121.Savage DC. Microbial ecology of the gastrointestinal tract. Annu Rev Microbiol. 1977;31:107–33. doi: 10.1146/annurev.mi.31.100177.000543. [DOI] [PubMed] [Google Scholar]
- 122.Seesuriyachan P, Takenaka S, Kuntiya A, Klayraung S, Murakami S, Aoki K. Metabolism of azo dyes by Lactobacillus casei TISTR 1500 and effects of various factors on decolorization. Water Res. 2007;41:985–92. doi: 10.1016/j.watres.2006.12.001. [DOI] [PubMed] [Google Scholar]
- 123.Sellers C, Markowitz S. Reevaluating the carcinogenicity of ortho-toluidine: a new conclusion and its implications. Regul Toxicol Pharmacol. 1992;16:301–17. doi: 10.1016/0273-2300(92)90010-7. [DOI] [PubMed] [Google Scholar]
- 124.Shahin MM. Mutagenicity evaluation of nitroanilines and nitroaminophenols in Salmonella typhimurium. Int J Cosmet Sci. 1985;7:277–89. doi: 10.1111/j.1467-2494.1985.tb00422.x. [DOI] [PubMed] [Google Scholar]
- 125.Shannon-Lowe J, Matheson NJ, Cooke FJ, Aliyu SH. Prevention and medical management of Clostridium difficile infection. BMJ. 2010;340:c1296. doi: 10.1136/bmj.c1296. [DOI] [PubMed] [Google Scholar]
- 126.Sharma P, Singh L, Dilbaghi N. Optimization of process variables for decolorization of Disperse Yellow 211 by Bacillus subtilis using Box-Behnken design. J Hazard Mater. 2009;164:1024–9. doi: 10.1016/j.jhazmat.2008.08.104. [DOI] [PubMed] [Google Scholar]
- 127.Slokar YM, Majcen Le Marechal A. Methods of decoloration of textile wastewaters. Dyes and Pigments. 1998;37:335–356. [Google Scholar]
- 128.Stingley RL, Zou W, Heinze TM, Chen H, Cerniglia CE. Metabolism of azo dyes by human skin microbiota. J Med Microbiol. 2010;59:108–14. doi: 10.1099/jmm.0.012617-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Suzuki Y, Yoda T, Ruhul A, Sugiura W. Molecular cloning and characterization of the gene coding for azoreductase from Bacillus sp. OY1–2 isolated from soil. J Biol Chem. 2001;276:9059–65. doi: 10.1074/jbc.M008083200. [DOI] [PubMed] [Google Scholar]
- 130.Sweeney EA, Chipman JK, Forsythe SJ. Evidence for direct-acting oxidative genotoxicity by reduction products of azo dyes. Environ Health Perspect. 1994;102(Suppl 6):119–22. doi: 10.1289/ehp.94102s6119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Tamura Z, Semejima K. Metabolism of 4-dimethylaminoazobenzene and related compounds. 3. Metabolites of 4-dimethylaminoazobenzene in rat bile and influence of DAB feeding on their amounts. Chem Pharm Bull (Tokyo) 1963;11:1014–21. doi: 10.1248/cpb.11.1014. [DOI] [PubMed] [Google Scholar]
- 132.Uter W, Stropp G, Schnuch A, Lessmann H. Aniline--a ‘historical’ contact allergen? Current data from the IVDK and review of the literature. Ann Occup Hyg. 2007;51:219–26. doi: 10.1093/annhyg/mel076. [DOI] [PubMed] [Google Scholar]
- 133.Walker R, Gingell R, Murrells DF. Mechanisms of azo reduction by Streptococcus faecalis. I. Optimization of assay conditions. Xenobiotica. 1971;1:221–9. doi: 10.3109/00498257109033171. [DOI] [PubMed] [Google Scholar]
- 134.Wang CJ, Hagemeier C, Rahman N, Lowe E, Noble M, Coughtrie M, Sim E, Westwood I. Molecular cloning, characterisation and ligand-bound structure of an azoreductase fromPseudomonas aeruginosa. J Mol Biol. 2007;373:1213–28. doi: 10.1016/j.jmb.2007.08.048. [DOI] [PubMed] [Google Scholar]
- 135.Wang CJ, Laurieri N, Abuhammad A, Lowe E, Westwood I, Ryan A, Sim E. Role of tyrosine 131 in the active site of paAzoR1, an azoreductase with specificity for the inflammatory bowel disease prodrug balsalazide. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2010;66:2–7. doi: 10.1107/S1744309109044741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Watanabe M, Ishidate M. Metabolism of 4-dimethylaminoazobenzene and related compounds. V. Quantitative analysis of biliary and urinary metabolites of 4-dimethylamino-azobenzene in rat. Chem Pharm Bull (Tokyo) 1967;15:1461–9. doi: 10.1248/cpb.15.1461. [DOI] [PubMed] [Google Scholar]
- 137.Wostmann BS. The germfree animal in nutritional studies. Annu Rev Nutr. 1981;1:257–79. doi: 10.1146/annurev.nu.01.070181.001353. [DOI] [PubMed] [Google Scholar]
- 138.Xu H, Heinze TM, Chen S, Cerniglia CE, Chen H. Anaerobic metabolism of 1-amino-2-naphthol-based azo dyes (Sudan dyes) by human intestinal microflora. Appl Environ Microbiol. 2007;73:7759–62. doi: 10.1128/AEM.01410-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Xu H, Heinze TM, Paine DD, Cerniglia CE, Chen H. Sudan azo dyes and Para Red degradation by prevalent bacteria of the human gastrointestinal tract. Anaerobe. 2010;16:114–9. doi: 10.1016/j.anaerobe.2009.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Yasunaga K, Kiyonari A, Nakagawa M, Yoshikawa K. Different results of the Salmonella umu test between three isomers of phenylenediamine (PDA) derivatives. Drug Chem Toxicol. 2006;29:203–13. doi: 10.1080/01480540600566766. [DOI] [PubMed] [Google Scholar]
- 141.Yoshimi N, Sugie S, Iwata H, Niwa K, Mori H, Hashida C, Shimizu H. The genotoxicity of a variety of aniline derivatives in a DNA repair test with primary cultured rat hepatocytes. Mutat Res. 1988;206:183–91. doi: 10.1016/0165-1218(88)90159-0. [DOI] [PubMed] [Google Scholar]
- 142.You Z, Brezzell MD, Das SK, Hooberman BH, Sinsheimer JE. Substituent effects on the in vitro and in vivo genotoxicity of 4-aminobiphenyl and 4-aminostilbene derivatives. Mutat Res. 1994;320:45–58. doi: 10.1016/0165-1218(94)90058-2. [DOI] [PubMed] [Google Scholar]
- 143.Zocco MA, Ainora ME, Gasbarrini G, Gasbarrini A. Bacteroides thetaiotaomicron in the gut: Molecular aspects of their interaction. Digestive and Liver Disease. 2007;39:707–712. doi: 10.1016/j.dld.2007.04.003. [DOI] [PubMed] [Google Scholar]