

Abstract
Background
Genetically enhanced Metarhizium pingshaense are being developed for malaria vector control in Burkina Faso. However, not much is known about the local prevalence and pathogenicity of this fungus, so we prospected mosquitoes and plant roots (a common habitat for Metarhizium spp.) for entomopathogenic fungi.
Results
Our investigations showed that Metarhizium spp. represented between 29–74% of fungi isolated from plant root rhizospheres in diverse collection sites. At low spore dosages (1 × 106 conidia/ml), two mosquito-derived M. pingshaense isolates (Met_S26 and Met_S10) showed greater virulence against Anopheles coluzzii (LT80 of ~7 days) than isolates tested in previous studies (LT80 of ~10 days). In addition, the local isolates did not cause disease in non-target insects (honeybees and cockroaches).
Conclusions
Our work provides promising findings for isolating local Metarhizium strains for application in mosquito biological control and for future transgenic biocontrol strategies in Burkina Faso.
Electronic supplementary material
The online version of this article (10.1186/s13071-018-2796-6) contains supplementary material, which is available to authorized users.
Keywords: Metarhizium, Entomopathogenic fungi, Mosquitoes, Vector control, Honeybees, Cockroaches, Malaria, Burkina Faso
Background
Unlike mosquitocidal bacteria and viruses, ascomycete fungi can infect and kill insects without being ingested. As with chemical insecticides, tarsal contact alone is sufficient to kill mosquitoes [1]. Despite intensive efforts to develop entomopathogenic fungi as biocontrol agents against malaria vectors, the strains under investigation have not met expectations due to their poorer efficacy relative to cheaper chemical insecticides [2]. The United States Department of Agriculture (USDA) ARSEF collection (the world’s largest collection of entomopathogenic fungi) has more than 12,000 isolates of insect pathogenic fungi. Of these, only 156 are from sub-Saharan Africa (South Africa and Benin are the source of 40 and 36 isolates, respectively), with none from Burkina Faso. The mosquitocidal activity of Metarhizium has been enhanced by engineering them to express insect-selective neurotoxins [3–5], and a transgenic strain of Metarhizium pingshaense is being evaluated in semi-field trials in Burkina Faso [5]. We speculate that future development of transgenic fungi worldwide will preferentially use local isolates as these may be better adapted to kill local mosquitoes and survive harsh local conditions (i.e. rainy season heat, sunlight and humidity) than exotic strains. However, the distribution and properties of indigenous Burkinabe Metarhizium spp. have not been characterized. The first objective of this study was to prospect for the presence and distribution of local Metarhizium strains. As well as prospecting mosquitoes, we also sampled rhizosphere soils (i.e. the soil in the vicinity of plant roots that is influenced by root secretions), as some Metarhizium spp. are abundant in the rhizosphere and may function as symbionts promoting plant growth. The plant-beneficial effects of Metarhizium species correlate with their association with roots and are mediated via plant hormones [6]. The second objective was to evaluate the pathogenicity of local Metarhizium isolates against wild-caught, insecticide-resistant Anopheles coluzzii. Finally, we also assessed the pathogenicity of the local isolates against American cockroaches and honeybees as representative non-target or beneficial species.
Methods
Fungal collection, isolation and morphological identification
Collections were carried out on a monthly basis during the 2015 rainy season (from July to September) from plant roots and wild-caught mosquitoes. Our three collection sites were the Kou Valley (11°23'N, 4°24'W), a rice crop area; Bana (11°9'41"N, 4°10'30"W), a savanna and forested area; and Soumousso (11°04'N, 4°03'W), a savanna and corn crop area (Fig. 1). One hundred and fifty-five plants were sampled from these three different agro-ecological sites. We followed the protocol described in [7] to collect rhizosphere soil and isolate fungi. The fungal selective medium contained 42 g potato-dextrose agar, 0.5 g chloramphenicol and 0.6 g cetyl trimethylammonium bromide per liter.
Fig. 1.
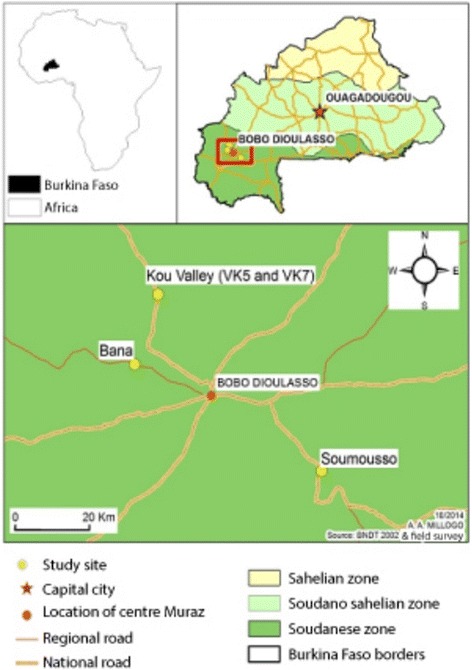
Rhizosphere and mosquito collection sites
Overall, 300 mosquitoes were collected from 3 types of resting sites (inhabited houses, abandoned houses and outdoor piles of wood). Mosquitoes were brought to the IRSS/Centre Muraz insectary, where they were fed on 6% sterile glucose ad libitum. Approximately 22% of collected mosquitoes (67 mosquitoes) died within 2 weeks and were plated on selective medium for fungal isolations.
Fungal isolates from rhizospheres or mosquitoes were identified using macro-morphological characters, such as conidiogenesis, estimation of radial growth, spore color and mycelia texture of the isolates on PDA media according to Humber [8]. In addition, we used microscopic morphology to identify Metarhizium spp. spores as described by Fernandes et al. [9]. Met_S10 and Met_S26 were confirmed as Metarhizium pingshaense through amplification and Sanger sequencing of the intron-rich region of translation elongation factor 1-α [10].
Fungal virulence on mosquitoes, honeybees and cockroaches
Initial screens on mosquitoes revealed two promising isolates (Met_S10 and Met_S26) isolated from mosquito cadavers from Soumousso and Bana, respectively, that readily grew on PDA and were highly virulent (Additional file 1: Table S1): these strains were therefore chosen for further characterization.
Bioassay on mosquitoes
For bioassays, we used An. coluzzii adult mosquitoes reared from larval collections at the Kou Valley, Burkina Faso. Mosquitoes from this area are known to be highly resistant to multiple insecticides [5, 11]. We carried out bioassays with local M. pingshaense isolates Met_S10 and Met_S26. A M. pingshaense strain that has been used as the foundation for development of transgenic mosquito control technologies was used as a positive control; this strain was engineered to constitutively express red fluorescent protein (RFP) [5]. Expression of RFP provides a fluorescent tag for following infection processes without altering virulence. We used an atomizer protocol for infections, as described previously [12]. Three serial concentrations were used: 1 × 108; 1 × 107; and 1 × 106 conidia/ml. We confirmed that this inoculation technique was able to deliver a repeatable inoculating dose (mean ± SE): 276 ± 16 spores per mosquito with 1 × 108; 211 ± 13 spores per mosquito with 1 × 107 spores/ml; and 44 ± 3 spores per mosquito with 1 × 106. Mortality was counted twice daily over two weeks.
Bioassay on non-target insects
We bioassayed Met_S10, Met_S26 and Met_RFP against a breeding line of honeybees, Apis mellifera adansonii (Latreille, 1804), as well as American cockroaches, Periplaneta americana (Linnaeus, 1758) caught in households from Soumousso. Spore doses were 1 × 108, 1 × 107 or 1 × 106 conidia/ml, as described previously [5]. Following treatment, insects were kept in our insectarium at 25.3 ± 1 °C and 70 ± 10% relative humidity. Mortality was counted twice daily over two weeks.
Results and discussion
Metarhizium spp. were isolated from rhizosphere soil samples across 3 sample sites: the Kou Valley, Bana and Soumousso. From the Kou Valley and Bana, we isolated 362 and 306 soil samples, respectively. Metarhizium spp. comprised 28.71% (n = 56) of the isolates from Bana and 30.72% (n = 94) of the total isolated fungi from the Kou Valley. We isolated 152 fungal strains from Soumousso; of these, 113 (74.34%) were Metarhizium, with a mean of 1.18 isolates/gram of soil (Additional file 2: Table S2). Soumousso is a savanna and corn crop area, and the higher proportion of Metarhizium fungi is consistent with previous studies that reported a strong association between Metarhizium spp. and soils from cultivated habitats, particularly field crops [13–15].
Isolates of Metarhizium spp. represented ~1% (8/801; 3 isolates from Culex spp. and 5 isolates from Anopheles gambiae (sensu lato) of the fungi isolated from mosquitoes. Fifteen colonies of Beauveria spp. were isolated on mosquitoes (5 isolates from Aedes aegypti and 10 isolates from Anopheles gambiae (s.l.) at Soumousso). Trichoderma was the predominant genus at all sites being isolated from 56% (Vallée du Kou) to 79% (Soumousso) of mosquitoes (Additional file 2: Table S2). However, two Metarhizium isolates (Met_S10 and Met_S26), collected from Anopheles gambiae (s.l.), in an inhabited house in Soumousso and in a woodpile in Bana, respectively, were more virulent against mosquitoes than other isolates, including those from rhizospheres (Additional file 1: Table S1). At 1 × 108 and 1 × 107 conidia/ml, both strains achieved lower LT50 than Met_RFP (LT50 of ~6 days) [16]. At the highest concentration (1 × 108 conidia/ml), the LT80 of Met_S10 (5.67 ± 0.17 days) was significantly lower than both Met_26 (LT80 = 7.50 ± 0.29 days; Welch t = -5.5, df = 3.2, P = 0.01) and Met_RFP (7.17 ± 0.17 days; Welch t = -6.364, df = 4, P = 0.003). At the lowest concentration (1 × 106 conidia/ml), Met_S10 still had a significantly (Welch t = -5.1962, df = 3.2, P = 0.011) lower LT80 (7.00 ± 0.29 days) compared to Met S26 and Met_RFP, which both had LT80’s of 10 days (Fig. 2, Table 1). At intermediate concentrations, all strains achieved 80% mortality, which is the threshold value from the World Health Organization Pesticide Evaluation Scheme (WHOPES) for successful control with insecticides [17]. Thus, our results revealed higher virulence for the native isolate Met_S10, against wild-caught, insecticide-resistant Anopheles coluzzii. The virulence of these isolates to mosquitoes is also higher than isolates from Benin and in Kenya where Metarhizium anisoplae strains were originally isolated from a white fly, Trialeurodes vaporariorum [16, 18].
Fig. 2.
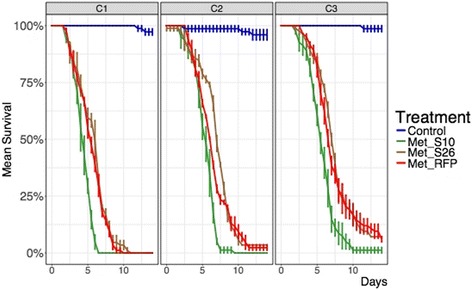
Survival curves of mosquitoes infected with Burkina Faso Metarhizium pingshaense isolates at different concentrations: C1, 1 × 108 conidia/ml; C2, 1 × 107 conidia/ml; C3, 1 × 106 conidia/ml
Table 1.
LT80s and grouping LT80 values for Anopheles coluzzii adults treated with Burkina Faso local Metarhizium pingshaense strains (Met_10 and Met_26) compared with wild type Metarhizium pingshaense expressing red fluorescent protein (Met_RFP) at three different concentrations
| Concentration (conidia/ml)a | Treatment | LT80 + SE (days) | Grouping LT80b |
|---|---|---|---|
| C1 (1 × 108) | Met_S10 | 5.67 ± 0.167 | a |
| Met_S26 | 7.50 ± 0.289 | b | |
| Met_RFP | 7.18 ± 0.167 | b | |
| C2 (1 × 107) | Met_S10 | 6.67 ± 0.167 | a |
| Met_S26 | 8.67 ± 0.167 | b | |
| Met_RFP | 8.83 ± 0.167 | b | |
| C3 (1 × 106) | Met_S10 | 7.00 ± 0.289 | a |
| Met_S26 | 10.00 ± 0.500 | b | |
| Met_RFP | 10.00 ± 1.041 | b |
Abbreviation: SE standard error of the mean
aIn 0.01% Tween80
bPairwise comparison of LT80 values per spraying conidia suspension concentrations; treatments with no letters in common differ significantly at P < 0.05
We bioassayed honeybees and cockroaches with the local strains and Met_RFP. However, even at the highest spore dosage (1 × 108 conidia/ml), these fungi did not significantly increase mortality compared to controls containing no conidia (Table 2). Fewer than 5% of honeybees and cockroaches died during the bioassays, and no mycosis was observed on any cadavers. This is in agreement with previous studies that report Met_RFP is a specialist to Culicidae [5]. The host ranges of different Metarhizium strains are chiefly controlled by recognition events on the cuticle [19], and the cuticles of honeybees, cockroaches and mosquitoes would likely have many topographical and chemical differences.
Table 2.
Two week-survival and grouping survival values for non-target insects (Honeybees and Cockroachs) treated with Burkina Faso local Metarhizium pingshaense strains (Met_10 and Met_26) compared with wild type Metarhizium pingshaense expressing red fluorescent protein (Met_RFP) at three different concentrations and a control (0.01% Tween)
| Non-target insect | Concentration (conidia/ml)a | Treatment | Survival + SE (%) | Grouping survivalb |
|---|---|---|---|---|
| Honeybee | C1 (1 × 108) | Control | 93.8 ± 1 | a |
| Met_RFP | 98.2 ± 1 | a | ||
| Met_S10 | 98.1 ± 2 | a | ||
| Met_S26 | 94.6 ± 2 | a | ||
| C2 (1 × 107) | Control | 94.6 ± 1 | a | |
| Met_RFP | 97.3 ± 2 | a | ||
| Met_S10 | 98.3 ± 1 | a | ||
| Met_S26 | 97.3 ± 0 | a | ||
| C3 (1 × 106) | Control | 95.3 ± 1 | a | |
| Met_RFP | 99.1 ± 1 | a | ||
| Met_S10 | 99.0 ± 0 | a | ||
| Met_S26 | 95.1 ± 2 | a | ||
| Cockroach | C1 (1 × 108) | Control | 95.7 ± 2 | a |
| Met_RFP | 97.8 ± 2 | a | ||
| Met_S10 | 98.8 ± 1 | a | ||
| Met_S26 | 97.5 ± 1 | a | ||
| C2 (1 × 107) | Control | 96.1 ± 2 | a | |
| Met_RFP | 97.5 ± 1 | a | ||
| Met_S10 | 98.7 ± 1 | a | ||
| Met_S26 | 97.7 ± 1 | a | ||
| C3 (1 × 106) | Control | 96.0 ± 1 | a | |
| Met_RFP | 97.0 ± 1 | a | ||
| Met_S10 | 97.0 ± 1 | a | ||
| Met_S26 | 96.0 ± 1 | a |
Abbreviation: SE standard error of the mean
aIn 0.01% Tween80
bPairwise comparison of survival mean values per spraying conidia suspension concentrations; treatments with no letters in common differ significantly at P < 0.05
Despite being more virulent than other WT Metarhizium strains, the Burkinabe Anopheles-derived isolates are still significantly less effective than transgenic strains expressing arthropod toxins [5]. However, our results suggest that these native Burkinabe Metarhizium strains would make attractive candidates for transgenic virulence enhancement and subsequent use as transgenic biocontrol agents.
Conclusion
Native fungal isolates may offer a superior alternative to introducing a foreign biocontrol strain, as they may be better adapted to both kill local mosquitoes and survive local conditions. There are also regulatory and ecological advantages to using strains already present in the country or in the ecosystem. This study provides a promising precedent for isolating local Metarhizium strains for application in mosquito biological control, and it lays a foundation for future transgenic biocontrol projects in Burkina Faso.
Additional files
Table S1. Preliminary infections data on mosquitoes. (XLSX 34 kb)
Table S2. List of fungal strains isolated from rhizosphere and mosquitoes. (XLSX 72 kb)
Acknowledgements
We are very grateful to Dr Amélie Vantaux for critical reading of the manuscript and to Dr Ali Drabo, Gnada Kobo Daniel, Eli Kabré, Athur D. Djibougou, Jacques E. Gnambani and Issiaka Saré for the technical assistance.
Funding
This work was supported by the US National Institute of Allergy and Infectious Diseases of the National Institutes of Health (Grant RO1 AI106998 to Raymond St. Leger) under Subaward Z047801/Centre Muraz to Dr Abdoulaye Diabate.
Availability of data and materials
The datasets generated during the current study are available from the corresponding author upon request.
Abbreviations
- Met_S10
Metarhizium pingshaense strain No. 10
- Met_S26
Metarhizium pingshaense strain No. 26
- Met_RFP
Metarhizium pingshaense expressing red fluorescent protein (RFP)
- LT50
Median (50%) lethal time after exposure to fungal infections
- LT80
80% lethal time after exposure to fungal infections
- SE
Standard error
Authors’ contributions
AD, BL, RKD, AS and EB designed the experiments. EB and BL performed the experiments and analysed the data. EB, BL, RJSL and AD wrote the manuscript. All authors read and approved the final manuscript.
Ethics approval
Ethical permissions were obtained through the Institutional Review of Institut de Recherche en Science de la Santé (IRSS) and Centre Muraz ethics committee (A012-2014/CE-CM). Prior authorization was granted from the Burkina Faso National Biosecurity Agency for sampling native fungi from rhizosphere of plants and mosquitoes (Ministerial Ordinance No. 2012-059/MRSI/SG/ANB). In addition, authorization was granted for importing and using both wild types and transgenic Metarhizium fungi for semi-field and lab work (Ministerial Ordinance No.2012-061/MRSI/SG/ANB). All bioassays with mosquitoes and non-target insects were carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. In addition, the protocols followed the IRSS Animal Welfare Assurance A5926-01.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Footnotes
Electronic supplementary material
The online version of this article (10.1186/s13071-018-2796-6) contains supplementary material, which is available to authorized users.
Contributor Information
Etienne Bilgo, Email: bilgo02@yahoo.fr.
Brian Lovett, Email: lovettbr@umd.edu.
Raymond J. St. Leger, Email: stleger@umd.edu
Antoine Sanon, Email: sanonant@yahoo.fr.
Roch K. Dabiré, Email: dabireroch@gmail.com
Abdoulaye Diabaté, Email: npiediab@gmail.com.
References
- 1.Farenhorst M, Knols BGJ. Fungal entomopathogens for the control of adult mosquitoes: a look at the issues. Proc Netherlands Entomol Soc Meet. 2007;18:51–9.
- 2.Thomas MB. Biological control of human disease vectors: a perspective on challenges and opportunities. BioControl. 2018;63:61–69. doi: 10.1007/s10526-017-9815-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Wang C, St Leger RJ. A scorpion neurotoxin increases the potency of a fungal insecticide. Nat Biotechnol. 2007;25:1455–1456. doi: 10.1038/nbt1357. [DOI] [PubMed] [Google Scholar]
- 4.Fang W, Lu H-L, King GF, St. Leger RJ. Construction of a hypervirulent and specific mycoinsecticide for locust control. Sci Rep. 2014;4:7345. doi: 10.1038/srep07345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bilgo E, Lovett B, Fang W, Bende N, King GF, Diabate A, et al. Improved efficacy of an arthropod toxin expressing fungus against insecticide-resistant malaria-vector mosquitoes. Sci Rep. 2017;7:3433. doi: 10.1038/s41598-017-03399-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Liao X, Lovett B, Fang W, St Leger RJ. Metarhizium robertsii produces indole-3-acetic acid, which promotes root growth in Arabidopsis and enhances virulence to insects. Microbiology. 2017;163:980–991. doi: 10.1099/mic.0.000494. [DOI] [PubMed] [Google Scholar]
- 7.Wyrebek M, Huber C, Sasan RK, Bidochka MJ. Three sympatrically occurring species of Metarhizium show plant rhizosphere specificity. Microbiology. 2011;157:2904–2911. doi: 10.1099/mic.0.051102-0. [DOI] [PubMed] [Google Scholar]
- 8.Humber RA. Entomopathogenic fungal identification. 2005. Available at https://www.ars.usda.gov/ARSUserFiles/80620520/apswkshoprev.pdf. Accessed 17 Nov 2017.
- 9.Fernandes ÉKK, Keyser CA, Rangel DEN, Foster RN, Roberts DW. CTC medium: a novel dodine-free selective medium for isolating entomopathogenic fungi, especially Metarhizium acridum, from soil. Biol Control. 2010;54:197–205. doi: 10.1016/j.biocontrol.2010.05.009. [DOI] [Google Scholar]
- 10.Bischoff JF, Rehner SA, Humber RA. A multilocus phylogeny of the Metarhizium anisopliae lineage. Mycologia. 2009;101:512–530. doi: 10.3852/07-202. [DOI] [PubMed] [Google Scholar]
- 11.Namountougou M, Simard F, Baldet T, Diabaté A, Ouédraogo JB, Martin T, et al. Multiple insecticide resistance in Anopheles gambiae s.l. populations from Burkina Faso, West Africa. PLoS One. 2012;7(11):e48412. doi: 10.1371/journal.pone.0048412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Fang W, Vega-Rodriquez J, Ghose AK, Jacobs-Lorena M, Kang A, St Leger RJ. Development of transgenic fungi that kill human malaria parasites in mosquitoes. Science. 2011;331:1074–1077. doi: 10.1126/science.1199115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Keyser CA, De Fine Licht HH, Steinwender BM, Meyling NV. Diversity within the entomopathogenic fungal species Metarhizium flavoviride associated with agricultural crops in Denmark. BMC Microbiol. 2015;15:249. doi: 10.1186/s12866-015-0589-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sánchez-Peña SR, Lara JS-J, Medina RF. Occurrence of entomopathogenic fungi from agricultural and natural ecosystems in Saltillo, México, and their virulence towards thrips and whiteflies. J Insect Sci. 2011;11:1. doi: 10.1673/031.011.0101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Quesada-Moraga E, Navas-Cortés JA, Maranhao EAA, Ortiz-Urquiza A, Santiago-Álvarez C. Factors affecting the occurrence and distribution of entomopathogenic fungi in natural and cultivated soils. Mycol Res. 2007;111:947–966. doi: 10.1016/j.mycres.2007.06.006. [DOI] [PubMed] [Google Scholar]
- 16.Scholte EJ, Takken W, Knols BGJ. Pathogenicity of five east African entomopathogenic fungi against adult Anopheles gambiae s.s. mosquitoes (Diptera: Culicidae) Proc Exp Appl Entomol Soc. 2003;14:25–29. [Google Scholar]
- 17.World Health Organization. Guidelines for laboratory and field-testing of long-lasting insecticidal nets. Who/Htm/Ntd/Whopes/20131; 2013. www.who.int/about/licensing/copyright_form/en/index.html. Accessed 26 Nov 2017.
- 18.Howard AFV, Koenraadt CJM, Farenhorst M, Knols BGJ, Takken W. Pyrethroid resistance in Anopheles gambiae leads to increased susceptibility to the entomopathogenic fungi Metarhizium anisopliae and Beauveria bassiana. Malar J. 2010;9:168. [DOI] [PMC free article] [PubMed]
- 19.Wang C, St Leger RJ. Developmental and transcriptional responses to host and nonhost cuticles by the specific locust pathogen. Society. 2005;4:937–947. doi: 10.1128/EC.4.5.937-947.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1. Preliminary infections data on mosquitoes. (XLSX 34 kb)
Table S2. List of fungal strains isolated from rhizosphere and mosquitoes. (XLSX 72 kb)
Data Availability Statement
The datasets generated during the current study are available from the corresponding author upon request.