

Abstract
In their seminal paper published in 1979, Gould and Lewontin argued that some traits arise as by-products of the development of other structures and not for direct utility in themselves. We show here that this applies to the trident, a pigmentation pattern observed on the thorax of Drosophila melanogaster. Using reporter constructs, we show that the expression domain of several genes encoding pigmentation enzymes follows the trident shape. This domain is complementary to the expression pattern of stripe (sr), which encodes an essential transcription factor specifying flight muscle attachment sites. We demonstrate that sr limits the expression of these pigmentation enzyme genes to the trident by repressing them in its own expression domain, i.e. at the flight muscle attachment sites. We give evidence that repression of not only yellow but also other pigmentation genes, notably tan, is involved in the trident shape. The flight muscle attachment sites and sr expression patterns are remarkably conserved in dipterans reflecting the essential role of sr. Our data suggest that the trident is a by-product of flight muscle attachment site patterning that arose when sr was co-opted for the regulation of pigmentation enzyme coding genes.
Introduction
Body pigmentation is at the interface between the organisms and their environment and fulfils many ecologically relevant functions. Indeed, the adaptive role of pigmentation seems often so obvious – i.e. in crypsis, mimicry, aposematism, mate recognition, UV protection or thermoregulation - that it is generally assumed that the observed patterns have been selected. Direct selection on pigmentation has been demonstrated in many cases, such as the famous industrial melanism of the peppered moth, Biston betularia, whose molecular basis has recently been identified1,2. Pigmentation patterns can also be the object of a trade-off between opposite selection forces. For example, in guppy Poecilia reticulata males, pigmentation patterns result from a balance between selection for crypsis, an anti-predator strategy, and selection for conspicuousness to attract females3. Further evidence for selection is the convergent loss of pigmentation when selection is relaxed in organisms living in the absence of light, such as cave animals4,5. However, since Gould and Lewontin’s seminal paper in 19796, it is widely acknowledged that adaptationist explanations should sometime be used with caution and developmental constraints also taken into account. Gould and Lewontin used the example of the spandrels decorated with splendid mosaics between the arches supporting the dome of the basilica of Saint-Mark in Venice. Although the mosaics fit remarkably well on the spandrels, the spandrels were not designed for them but result from architectural constraints imposed by the structure supporting the dome. Thus, Gould and Lewontin argued that some biological traits arose as by-products of developmental constraints on a crucial trait and were not selected for their direct utility6,7. Hence, developmental constraints favour particular patterns or morphologies whereas they forbid others. A few studies on pigmentation have addressed this question by exploring morphospaces with artificial selection experiments8,9. The size of the eyespots on butterfly Bicyclus anynana wings responds to selection but their colour is more constrained as only coordinated changes of pigments are possible8. Consequently, only some area of the morphospace can be occupied. Furthermore, the eyespot pattern through the genus Bicyclus follows a similar path of diversification, suggesting that evolution is partly constrained and that selection (or drift) can operate only in particular directions8.
Drosophila pigmentation is an interesting model to study the impact of developmental constraints10. This highly evolvable trait has been the focus of many studies analysing the genetic bases of morphological variation within and between species11,12. Body or wing pigmentation relies on the coordinated action of trans-regulatory factors on genes encoding pigmentation enzymes13–20. Most of these factors, for example Engrailed or Abdominal-B, are spatially restricted components of a deeply conserved regulatory landscape involved in development of many essential traits. Hence, pigmentation patterns can be interpreted as targets of indirect selection due to their association with another trait21.
The trident is a melanic pattern observed on the dorsal thorax of Drosophila melanogaster22. In natural populations, the intensity of the trident shows clinal genetic variation, with darker tridents observed at higher latitudes in Europe, India and Australia and higher altitudes in India and Africa23–26. The variable intensity of the trident is therefore thought to be an adaptation to temperature and/or UV24,25. Differences in the intensity of the trident were shown to be linked to genetic variation in the pigmentation gene ebony (e), which is less expressed in flies with a darker trident26,27. Besides, the trident is more clearly visible in an e mutant background28. Furthermore, the intensity of the trident is sensitive to developmental temperature24. This trait is therefore an example of phenotypic plasticity, “the ability of a given genotype to produce different phenotypes in response to distinct environmental conditions”29. Interestingly, in Drosophila busckii, a pattern similar to the trident is very clearly delineated on the thorax, being dark brown on a yellow background22,30. Thus, the intensity of the trident is variable, plastic and highly evolvable. By contrast, the developmental bases of its shape have attracted little attention. Regulatory genes, whose mutation modifies it, are the most promising candidates to address the developmental constraints exerted. Mutation in the stripe gene (sr1 allele) affects the shape of the trident31. This gene encodes an Egr-like zinc-finger transcription factor specifying epidermal tendon cells, to which muscles attach32–35. Therefore, sr expression labels the flight muscle attachment sites on the pupal thorax32,35. The expression pattern of sr on the pupal thorax36 seems complementary to the trident, which suggests that sr might delimit the trident by repressing melanin production on the thorax. Using reporter constructs made with the regulatory sequences of pigmentation enzyme coding genes, we investigate here the role of sr in the establishment of the trident’s pattern and its possible relationship with the positioning of the flight muscles. Using mutants, we show that sr represses the expression of several pigmentation enzyme-coding genes in the thorax epidermis, thus shaping the trident. The flight muscle attachment sites and sr expression pattern are remarkably conserved in dipterans, reflecting the essential role of sr36,37. Our data suggest that the shape of the trident is a by-product of flight muscle attachment site patterning that arose when sr was co-opted for the regulation of pigmentation enzyme-coding genes.
Results and Discussion
stripe represses melanin production on the thorax
The pattern of the trident varies among Drosophila melanogaster lines, being absent in some of them (w1118) whereas clearly visible in others (41Jd) (Fig. 1a,b). In ebony mutant (e1), the trident is very visible (Fig. 1c) as previously reported28. The trident is not limited to Drosophila melanogaster as it is reminiscent of the pigmentation pattern observed in another Drosophila species, Drosophila busckii (Fig. 1d).
Figure 1.

The trident pattern in Drosophila. (a) Absence of trident in w1118. (b) Trident clearly visible in 41Jd. (c) Dark trident of an ebony e1 mutant. (d) Distinctive trident in Drosophila busckii. (e) Trident of an ebony es mutant. (f) Dark longitunal stripe in an es sr1 double mutant. (g) Trident still visible in an y1, es mutant background. (g) Longitudinal dark band in a y1; es sr1 mutant.
Apparent complementarity between sr expression pattern and the trident in the pupal thorax36 suggests that sr might be involved in trident patterning. To address this question, we took advantage of the sr1 allele of sr. This is a very old allele, as it is reported to have been isolated by Calvin Bridges (1889–1938)31. In sr1 flies, the trident is replaced by a broad longitudinal stripe, hence the name of the gene31. It is only thanks to the much more recent description of the sr expression pattern and to the characterization of the sr1 allele that this phenotype can be interpreted. It was shown that sr1 is a regulatory mutant, which loses the most dorsal domains of sr expression on the thorax36. To render the dark longitudinal stripe more visible, we combined sr1 with the es hypomorphic allele31. The dark dorsal longitudinal stripe induced by sr1 was perfectly visible in the es background (compare Fig. 1e,f). These data suggest that the dark longitudinal stripe is a modified trident, in which spaces between teeth are filled with melanin.
These spaces correspond to the dorsal domains of sr expression that are missing in sr1, which suggests that sr shapes the trident by repressing melanin production. This repression could occur through different mechanisms implicating the expression, the stability or/and the activity of one or several pigmentation enzymes. Production of cuticle pigments involves many enzyme-coding genes arranged into a pathway38,39. Interestingly, among them, ple (encoding the Tyrosine Hydroxylase), Ddc (encoding the Dopa decarboxylase), yellow (y), and e were shown previously to be expressed in the trident28,40,41. Over-expression of y in the dorso-medial domain in an e mutant background is sufficient to generate a homogenous black pigmentation28. Thus, restriction of y expression to the trident is sufficient to explain its delimitation. However, in a y1, es double mutant, the trident is still visible (Fig. 1g) as previously reported28 and in a y1; sr1, es triple mutant, the longitudinal pattern typical of sr1 is clearly visible (Fig. 1h). This implies that other pigmentation enzymes than y and e are involved in the patterning of the trident downstream of sr.
Expression of stripe and tan in the thorax are complementary
Most pigmentation enzyme genes were previously shown to be expressed in the trident28,40,41. However, expression of tan (t), which encodes an enzyme involved in melanin synthesis, was never analysed in the thorax. Using a transgene, in which the expression of nuclear enhanced green fluorescent protein (nEGFP) was driven by the t abdominal enhancer t_MSE42,43, we observed nEGFP expression in the trident (Fig. 2a), showing than t_MSE was also activated in this motif. To reveal sr expression, we used the enhancer trap line srmd710 (sr-Gal4)36 and the UAS-mCherry-NLS transgene. mCherry expression was visible in the notum of pupae, where it precisely labelled the flight muscle attachment sites36 (Fig. 2b). By combining (srmd710, UAS-mCherry-NLS) with the t_MSE-nEGFP transgene, we observed a remarkable complementarity between the patterns of mCherry and nEGFP (Fig. 2c). All flight muscle attachment sites expressing mCherry corresponded to regions where nEGFP was absent, and a high nEGFP level was observed outside of the flight muscle attachment sites.
Figure 2.
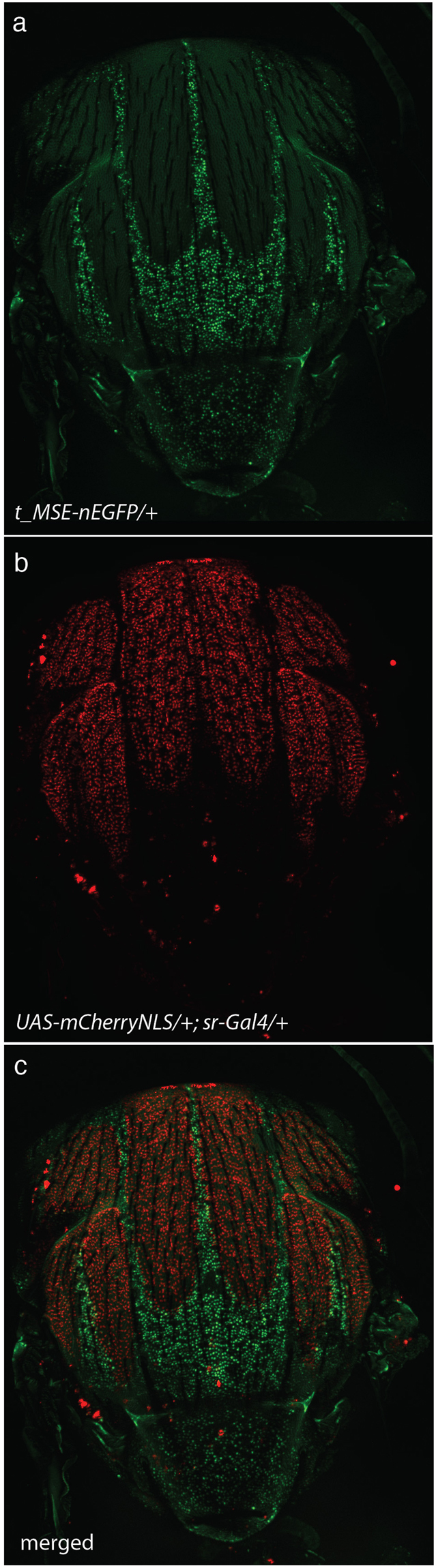
Complementary expression of stripe and tan in the thorax. Thorax of a UAS-mCherry-NLS/t_MSE-nEGFP; sr-Gal4/+ freshly eclosed fly. (a) nEGFP showing the activity of t regulatory sequences; (b) mCherry showing sr expression; (c) Merge.
stripe delimits the shape of the trident by repressing multiple pigmentation genes
Complementarity between sr and t expression domains on the thorax suggests that sr might repress the expression of t. This could also be the case for all pigmentation enzyme genes. Then, to draw up a precise and complete analysis of the expression domains of pigmentation enzymes in the thorax, as compared to the expression domain of sr, we used Ddc-Gal444 and ple-Gal445 associated with the UAS-mCherry-NLS transgene as well as transgenes expressing nEGFP under the control of t, e or y regulatory sequences13,41,42. In the control background, all reporters were expressed in the thorax with patterns that resembled the trident (Fig. 3a,c,e,g,i) suggesting that the expression of pigmentation enzyme genes was constrained to this motif. By contrast, in the sr1 background, expression of the reporters was extended dorsally and the trident motif disappeared (Fig. 3b,d,f,h,j as compared to a, c, e, g, i, respectively). These data indicate that sr represses Ddc, ple, t, e and y in the thorax. Hence, the shape of the trident likely reflects the spatial regulation of pigmentation enzyme coding genes by Stripe.
Figure 3.

Effect of the sr1 mutation on the expression of pigmentation enzyme coding genes. (a) Ddc expression in thorax of pharates visualized using Ddc-Gal4 and UAS-mCherry-NLS transgenes (UAS-mCherry-NLS/+; Ddc-Gal4). (b) Ddc expression in a sr1 background (UAS-mCherry-NLS/+; Ddc-Gal4, sr1/sr1). (c) ple expression visualized using ple-Gal4 and UAS-mCherry-NLS transgenes (UAS-mCherry-NLS/+; ple-Gal4/+). (d) ple expression in a sr1 background (UAS-mCherry-NLS/+; ple-Gal4, sr1/sr1). (e) t expression followed using the t_MSE-nEGFP transgene. (f) t expression in a sr1 background (t-MSE-nEGFP; sr1). (g) e expression visualized using the e-nEGFP transgene. (h) t expression in a sr1 background (e-nEGFP, sr1/sr1). (i) y expression followed using the y-wb-nEGFP transgene. (j) y expression in a sr1 background (y-wb-nEGFP; sr1).
Conclusion
We show here that sr regulates several pigmentation enzyme-coding genes on the thorax, although we do not know whether this regulation is direct or not. The expression of sr is conserved in Calliphora vicina, a species that diverged from Drosophila lineage about 100 million years ago, suggesting that this gene is a member of a deeply conserved regulatory landscape37. sr plays an essential role in the establishment of flight muscle attachment sites, and conservation of its expression is mirrored by a remarkable conservation of the flight muscle apparatus in dipterans46. In contrast, a clear thoracic trident complementary to the flight muscle attachment sites is observed in only a few species of flies, notably in Drosophila melanogaster22. Our results suggest that the co-option of sr for the regulation of pigmentation enzyme coding genes has led to the generation of the trident, a pigmentation pattern complementary to the flight muscle attachment sites. Therefore, the shape of the trident primarily results from a developmental constraint imposed by the flight muscle pattern. It is typically a “spandrel” in the sense of Gould and Lewontin (1979), a by-product of the development of another structure. The same applies to the position of large bristles (macrochaetes) on the thorax. Indeed, macrochaetes are excluded from the flight muscle attachment sites, and it was shown that the development of macrochaetes and tendon cells on the thorax are mutually exclusive36.
The fact that the trident was originally an indirect target of selection does not exclude that it has later become a direct target of selection. Indeed, clinal variation in the intensity of the trident strongly suggests an adaptive role in thermoregulation23,24,26. However, natural selection has targeted variation in the intensity of the trident, rather than in its shape that is highly constrained by fixed muscle attachment sites. Furthermore, it is possible that the expression of pigmentation enzymes in the trident confers new properties to the thorax cuticle that are important for flight, such as flexibility or mechanical endurance.
Methods
Fly stocks
w1118 is an inbred line used as control. The line 41Jb was established by Jean-Michel Gibert from a female caught in Marsais (France). The Drosophila busckii line was established from a female caught in Niort (France) and kindly provided by Dr Laure Teysset. The following stocks were obtained from the Bloomington Drosophila stock centre: e1 (BL-1658), es (BL-498), Ddc-Gal4 (BL-7009), ple-Gal4 aka TH-Gal4 (BL-8848), sr-Gal4 aka srmd710 (BL-26663) and UAS-mCherry-NLS (BL-38425). The lines y-wing-body-nEGFP, e-nEGFP (containing the regulatory regions ABC + intron) and t_MSE-nEGFP were kindly provided by Dr. Sean Carroll’s laboratory. Flies were grown on standard medium at 25 °C.
Image acquisitions
Thoracic cuticles of flies immerged in 75% ethanol were imaged with a binocular equipped with Leica DC480 digital camera, using the Leica IM50 Image Manager software. Stacks of 4–10 images were generated for each thorax. They were merged using Photoshop. Identical settings were used for all acquisitions.
Fluorescent images were acquired with a Macro-Apotome (Zeiss) with a 63 × objective on freshly decapitated flies immerged in PBS on an agarose substrate. Stacks were composed of around 75–115 pictures. Maximum intensity projections were created. Brightness and contrast were slightly adjusted in Photoshop.
Data availability
Data and materials used in this work are available on request.
Acknowledgements
Flybase provided information useful for this study. We thank the Bloomington Drosophila Stock Centre and the Sean Carroll laboratory for fly stocks and Dr Laure Teysset for the Drosophila busckii line. We thank other members of the lab for fruitful discussions.
Author Contributions
J.M.G. designed the study, performed the experiments and analysed the data. J.M.G., E.M.V. and F.P. wrote the paper.
Competing Interests
The authors declare no competing interests.
Footnotes
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Van’t Hof AE, et al. The industrial melanism mutation in British peppered moths is a transposable element. Nature. 2016;534:102–105. doi: 10.1038/nature17951. [DOI] [PubMed] [Google Scholar]
- 2.Nadeau NJ, et al. The gene cortex controls mimicry and crypsis in butterflies and moths. Nature. 2016;534:106–110. doi: 10.1038/nature17961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Endler JA. Natural selection on color patterns in Poecilia reticulata. Evolution. 1980;34:76–91. doi: 10.1111/j.1558-5646.1980.tb04790.x. [DOI] [PubMed] [Google Scholar]
- 4.Casane D, Rétaux S. Evolutionary Genetics of the Cavefish Astyanax mexicanus. Adv. Genet. 2016;95:117–159. doi: 10.1016/bs.adgen.2016.03.001. [DOI] [PubMed] [Google Scholar]
- 5.Bilandžija H, Cetković H, Jeffery WR. Evolution of albinism in cave planthoppers by a convergent defect in the first step of melanin biosynthesis. Evol. Dev. 2012;14:196–203. doi: 10.1111/j.1525-142X.2012.00535.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gould SJ, Lewontin RC. The spandrels of San Marco and the Panglossian paradigm: a critique of the adaptationist programme. Proc. R. Soc. Lond. B Biol. Sci. 1979;205:581–598. doi: 10.1098/rspb.1979.0086. [DOI] [PubMed] [Google Scholar]
- 7.Gould SJ. The exaptive excellence of spandrels as a term and prototype. Proc. Natl. Acad. Sci. USA. 1997;94:10750–10755. doi: 10.1073/pnas.94.20.10750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Allen CE, Beldade P, Zwaan BJ, Brakefield PM. Differences in the selection response of serially repeated color pattern characters: standing variation, development, and evolution. BMC Evol. Biol. 2008;8:94. doi: 10.1186/1471-2148-8-94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Beldade P, Koops K, Brakefield PM. Developmental constraints versus flexibility in morphological evolution. Nature. 2002;416:844–847. doi: 10.1038/416844a. [DOI] [PubMed] [Google Scholar]
- 10.Gompel N, Carroll SB. Genetic mechanisms and constraints governing the evolution of correlated traits in drosophilid flies. Nature. 2003;424:931–5. doi: 10.1038/nature01787. [DOI] [PubMed] [Google Scholar]
- 11.Wittkopp PJ, Carroll SB, Kopp A. Evolution in black and white: genetic control of pigment patterns in Drosophila. Trends Genet. 2003;19:495–504. doi: 10.1016/S0168-9525(03)00194-X. [DOI] [PubMed] [Google Scholar]
- 12.Massey JH, Wittkopp PJ. The Genetic Basis of Pigmentation Differences Within and Between Drosophila Species. Curr. Top. Dev. Biol. 2016;119:27–61. doi: 10.1016/bs.ctdb.2016.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Jeong S, Rokas A, Carroll SB. Regulation of body pigmentation by the Abdominal-B Hox protein and its gain and loss in Drosophila evolution. Cell. 2006;125:1387–99. doi: 10.1016/j.cell.2006.04.043. [DOI] [PubMed] [Google Scholar]
- 14.Arnoult L, et al. Emergence and diversification of fly pigmentation through evolution of a gene regulatory module. Science. 2013;339:1423–6. doi: 10.1126/science.1233749. [DOI] [PubMed] [Google Scholar]
- 15.Gompel N, Prud’homme B, Wittkopp PJ, Kassner VA, Carroll SB. Chance caught on the wing: cis-regulatory evolution and the origin of pigment patterns in Drosophila. Nature. 2005;433:481–7. doi: 10.1038/nature03235. [DOI] [PubMed] [Google Scholar]
- 16.Prud’homme B, et al. Repeated morphological evolution through cis-regulatory changes in a pleiotropic gene. Nature. 2006;440:1050–3. doi: 10.1038/nature04597. [DOI] [PubMed] [Google Scholar]
- 17.Koshikawa S, et al. Gain of cis-regulatory activities underlies novel domains of wingless gene expression in. Drosophila. Proc. Natl. Acad. Sci. USA. 2015;112:7524–7529. doi: 10.1073/pnas.1509022112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Werner T, Koshikawa S, Williams TM, Carroll SB. Generation of a novel wing colour pattern by the Wingless morphogen. Nature. 2010;464:1143–8. doi: 10.1038/nature08896. [DOI] [PubMed] [Google Scholar]
- 19.Williams TM, et al. The regulation and evolution of a genetic switch controlling sexually dimorphic traits in Drosophila. Cell. 2008;134:610–23. doi: 10.1016/j.cell.2008.06.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bray MJ, Werner T, Dyer KA. Two genomic regions together cause dark abdominal pigmentation in Drosophila tenebrosa. Heredity. 2014;112:454–462. doi: 10.1038/hdy.2013.124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Pigliucci, null & Kaplan, null. The fall and rise of Dr Pangloss: adaptationism andthe Spandrels paper 20 years later. Trends Ecol. Evol. 15, 66–70 (2000). [DOI] [PubMed]
- 22.Werner, T. & Jaenike, J. Drosophilids of the Midwest and Northeast. (River Campus Libraries, University of Rochester).
- 23.Munjal AK, et al. Thoracic trident pigmentation in Drosophila melanogaster: latitudinal and altitudinal clines in Indian populations. Genet Sel Evol. 1997;29:601–10. doi: 10.1186/1297-9686-29-5-601. [DOI] [Google Scholar]
- 24.David JR, Capy P, Payant V, Tsakas S. Thoracic trident pigmentation in Drosophila melanogaster: Differentiation of geographical populations. Genet Sel Evol. 1985;17:211–24. doi: 10.1186/1297-9686-17-2-211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bastide H, Yassin A, Johanning EJ, Pool JE. Pigmentation in Drosophila melanogaster reaches its maximum in Ethiopia and correlates most strongly with ultra-violet radiation in sub-Saharan Africa. BMC Evol Biol. 2014;14:179. doi: 10.1186/s12862-014-0179-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Telonis-Scott M, Hoffmann AA, Sgro CM. The molecular genetics of clinal variation: a case study of ebony and thoracic trident pigmentation in Drosophila melanogaster from eastern Australia. Mol Ecol. 2011;20:2100–10. doi: 10.1111/j.1365-294X.2011.05089.x. [DOI] [PubMed] [Google Scholar]
- 27.Takahashi A, Takahashi K, Ueda R, Takano-Shimizu T. Natural variation of ebony gene controlling thoracic pigmentation in Drosophila melanogaster. Genetics. 2007;177:1233–7. doi: 10.1534/genetics.107.075283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wittkopp PJ, True JR, Carroll SB. Reciprocal functions of the Drosophila yellow and ebony proteins in the development and evolution of pigment patterns. Development. 2002;129:1849–58. doi: 10.1242/dev.129.8.1849. [DOI] [PubMed] [Google Scholar]
- 29.Pigliucci, M. Phenotypic Plasticity, Beyond Nature and Nurture. (2001).
- 30.Shorrocks, B. Drosophila. (Ginn & company limited, 1972).
- 31.Lindsley, D. L. & Zimm, G. G. The genome of Drosophila melanogaster. (Academic Press Limited, 1992).
- 32.Fernandes JJ, Celniker SE, VijayRaghavan K. Development of the indirect flight muscle attachment sites in Drosophila: role of the PS integrins and the stripe gene. Dev Biol. 1996;176:166–84. doi: 10.1006/dbio.1996.0125. [DOI] [PubMed] [Google Scholar]
- 33.Frommer G, Vorbrüggen G, Pasca G, Jäckle H, Volk T. Epidermal egr-like zinc finger protein of Drosophila participates in myotube guidance. EMBO J. 1996;15:1642–1649. [PMC free article] [PubMed] [Google Scholar]
- 34.Lee JC, VijayRaghavan K, Celniker SE, Tanouye MA. Identification of a Drosophila muscle development gene with structural homology to mammalian early growth response transcription factors. Proc. Natl. Acad. Sci. USA. 1995;92:10344–10348. doi: 10.1073/pnas.92.22.10344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ghazi A, Paul L, VijayRaghavan K. Prepattern genes and signaling molecules regulate stripe expression to specify Drosophila flight muscle attachment sites. Mech. Dev. 2003;120:519–528. doi: 10.1016/S0925-4773(03)00042-X. [DOI] [PubMed] [Google Scholar]
- 36.Usui K, Pistillo D, Simpson P. Mutual exclusion of sensory bristles and tendons on the notum of dipteran flies. Curr Biol. 2004;14:1047–55. doi: 10.1016/j.cub.2004.06.026. [DOI] [PubMed] [Google Scholar]
- 37.Richardson J, Simpson P. A conserved trans-regulatory landscape for scute expression on the notum of cyclorraphous Diptera. Dev. Genes Evol. 2006;216:29–38. doi: 10.1007/s00427-005-0028-5. [DOI] [PubMed] [Google Scholar]
- 38.Wright TR. The genetics of biogenic amine metabolism, sclerotization, and melanization in Drosophila melanogaster. Adv. Genet. 1987;24:127–222. [PubMed] [Google Scholar]
- 39.Gibert J-M, Mouchel-Vielh E, Peronnet F. Modulation of yellow expression contributes to thermal plasticity of female abdominal pigmentation in Drosophila melanogaster. Sci. Rep. 2017;7:43370. doi: 10.1038/srep43370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Gibert JM, Peronnet F, Schlotterer C. Phenotypic Plasticity in Drosophila Pigmentation Caused by Temperature Sensitivity of a Chromatin Regulator Network. PLoS Genet. 2007;3:e30. doi: 10.1371/journal.pgen.0030030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Rebeiz M, Pool JE, Kassner VA, Aquadro CF, Carroll SB. Stepwise modification of a modular enhancer underlies adaptation in a Drosophila population. Science. 2009;326:1663–7. doi: 10.1126/science.1178357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Jeong S, et al. The evolution of gene regulation underlies a morphological difference between two Drosophila sister species. Cell. 2008;132:783–93. doi: 10.1016/j.cell.2008.01.014. [DOI] [PubMed] [Google Scholar]
- 43.Gibert J-M, Mouchel-Vielh E, De Castro S, Peronnet F. Phenotypic Plasticity through Transcriptional Regulation of the Evolutionary Hotspot Gene tan in Drosophila melanogaster. PLoS Genet. 2016;12:e1006218. doi: 10.1371/journal.pgen.1006218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Li H, Chaney S, Roberts IJ, Forte M, Hirsh J. Ectopic G-protein expression in dopamine and serotonin neurons blocks cocaine sensitization in Drosophila melanogaster. Curr. Biol. CB. 2000;10:211–214. doi: 10.1016/S0960-9822(00)00340-7. [DOI] [PubMed] [Google Scholar]
- 45.Friggi-Grelin F, et al. Targeted gene expression in Drosophila dopaminergic cells using regulatory sequences from tyrosine hydroxylase. J Neurobiol. 2003;54:618–27. doi: 10.1002/neu.10185. [DOI] [PubMed] [Google Scholar]
- 46.Costa M, Calleja M, Alonso CR, Simpson P. The bristle patterning genes hairy and extramacrochaetae regulate the development of structures required for flight in Diptera. Dev. Biol. 2014;388:205–215. doi: 10.1016/j.ydbio.2013.12.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data and materials used in this work are available on request.