

Abstract
Fast‐scan cyclic voltammetry (FSCV) is an established method for measuring dopamine (DA) levels in the brain in real time. However, it is difficult to discriminate DA from other monoamines such as serotonin (5‐hydroxytryptamine, 5‐HT) and norepinephrine (NE). We report a novel DA‐specific biosensor consisting of a carbon‐fiber electrode coated with an ion‐exchange membrane, a layer containing monoamine oxidase B, and a cellulose membrane. We performed FSCV using the probe to monitor the amount of DA in vitro and in vivo. First, we measured currents in vitro in phosphate‐buffered saline as we added one micromole each of DA, 5‐HT, and NE. The results confirmed that the biosensor selectively detected DA. Next, we implanted the probe in the striatum of male rats to investigate whether it could selectively detect changes in the DA content in vivo. The probe detected both the tonic change induced by methamphetamine administration and the phasic change induced by electrical stimulation of the medial forebrain bundle. In contrast, the electrode in the 6‐hydroxydopamine–lesioned striatum did not respond to systemic selective serotonin or serotonin/norepinephrine reuptake inhibitors, confirming its selectivity. Furthermore, the probe in the striatum could still detect changes in the DA level 1 week after electrode implantation. The results suggest that the novel biosensor can measure real‐time changes in DA levels in vivo with a relatively high signal‐to‐noise ratio.
Keywords: electrode, fast‐scan cyclic voltammetry, monoamine oxidase
1. INTRODUCTION
Monoamine neurotransmitters such as dopamine (DA) play important roles in multiple central nervous system functions associated with various forms of behavior. Mesolimbic dopaminergic nuclei project axons to several brain regions, including the neocortex and basal ganglia, to modulate their neuronal activity (Alexander, DeLong, & Strick, 1986; Bourdy & Barrot, 2012; Fallon, 1981; Lindvall, Bjorklund, Moore, & Stenevi, 1974). These dopaminergic circuits are involved in various central functions such as motor output, emotion, and cognition (Bromberg‐Martin, Matsumoto, & Hikosaka, 2010; Floresco & Magyar, 2006; Hosp, Pekanovic, Rioult‐Pedotti, & Luft, 2011; McNamara, Tejero‐Cantero, Trouche, Campo‐Urriza, & Dupret, 2014). The importance of DA in central nervous system functioning and pathology has led to the development of high‐precision, real‐time DA measurement techniques.
Fast‐scan cyclic voltammetry (FSCV) is one of the primary techniques for measuring monoamine neurotransmitter regulation in vitro and in vivo. It can detect submicromolar levels of DA with a millisecond temporal resolution (Robinson, Venton, Heien, & Wightman, 2003). Dopaminergic neurons have two major firing patterns: a slow, “tonic” mode and a bursting, “phasic” mode (Grace & Bunney, 1983). The millisecond temporal resolution of voltammetry allows researchers to resolve these different modes and to measure DA in situ in real time. FSCV has been used to measure DA efflux in vitro and in vivo and to associate the release of DA with central nervous system functions (Garris, Walker, & Wightman, 1997; Hafizi, Kruk, & Stamford, 1990; Wu, Reith, Wightman, Kawagoe, & Garris, 2001). Importantly, using chronically indwelling electrodes, FSCV permits the correlation of DA fluctuations with behavior in awake animals (Fortin, Cone, Ng‐Evans, McCutcheon, & Roitman, 2015; Phillips, Robinson, Stuber, Carelli, & Wightman, 2003).
A major problem with FSCV is its poor selectivity. Catecholamines such as norepinephrine (NE) and indoleamines such as serotonin (5‐hydroxytryptamine; 5‐HT) have a similar structure to DA. These three monoamines show very similar FSCV current responses and oxidation potentials, making it very difficult to discriminate between them (Heien, Phillips, Stuber, Seipel, & Wightman, 2003; Kovach, Ewing, Wilson, & Wightman, 1984). Similar to dopaminergic neurons, other monoaminergic neurons project their axons broadly to central structures (Jung, Shi, Borland, Michael, & Weber, 2006). It is therefore difficult to discriminate the release of monoamines in vivo using spatial criteria. To aid the interpretation of results, it is preferable to minimize the difference between the response obtained by FSCV and the actual dopaminergic neural activity. The establishment of a method to discriminate the level of DA from that of other monoamines is a challenge that needs to be addressed.
Various improvements have been designed to increase the sensitivity and selectivity of FSCV systems (Anastassiou et al., 2006; Crespi, England, & Trist, 1995; Ramsson et al., 2015; Zachek, Hermans, Wightman, & McCarty, 2008). Because of the electrochemical similarity among monoamines, however, it is difficult to differentiate them using FSCV. To solve this problem, we aimed to produce a novel electrode that can discriminate the DA signal from that of other monoamines. We first developed a novel electrode coated with a double membrane comprising the ion exchanger Nafion and monoamine oxidase (MAO)‐B–diluted cellulose. We then evaluated the discriminative capacity of the electrode for DA against other monoamines in vitro and in vivo. Finally, the electrode was implanted into rat striata and used in chronic mode for in vivo experiments.
We successfully developed a novel biosensor with a relatively high signal‐to‐noise ratio, which can be implanted into the striatum to measure real‐time changes in DA level in vivo for up to a week.
2. MATERIALS AND METHODS
2.1. Voltammetric instrument and carbon‐fiber microelectrode
All data were obtained using an IMEC‐701 electrochemistry amplifier (Intermedical Co., Ltd., Nagoya, Japan). Voltammetric recordings were made every 100 ms by applying a triangular waveform (−0.4 to + 1.0 V, 300 V/s). Data were digitized using the NI USB‐6361 (Intermedical Co., Ltd.) and stored on a computer.
Carbon‐fiber electrodes were prepared by aspirating two carbon fibers (diameter 7–20 μm) into a glass capillary (length 8 mm; inner diameter 1 mm; Harvard Apparatus Ltd., Kent, UK, GC100‐10) (Figure 1). The glass tube was tapered and sealed using epoxy resin (Araldite, Huntsman Inc., Kobe, Japan). The exposed length (100 µm) of the carbon fiber extending from the tip of the glass capillary was soaked in a 20% MAO‐B/5% cellulose solution. MAO‐B was purchased from Sigma‐Aldrich (human recombinant, prepared from insect cells infected with recombinant baculovirus, M7441; St. Louis, MO). After drying for ∼30 to 60 min at room temperature, the MAO‐B/cellulose‐coated tip of the electrode was cross‐linked for 30 min by glutaraldehyde vapor, which was made of 25% glutaraldehyde solution in boiling water. Then, Nafion was applied to the tip. An electrical connection was made with the carbon fiber within the glass tube by applying silver paint to one end of a stainless‐steel tube and inserting it into the glass tube to reach the fiber. The distal end of the stainless tube was inserted into an electrical connector to make a convenient temporary connection to the preamp for the experiments. Ag/AgCl was applied to the surface of the glass capillary to form the reference electrode.
Figure 1.

(a,b) Photographs of the electrode (a) and its tip (b). (c) Schematic of the recording electrode. A carbon microfiber is sealed in a glass capillary with 100 μm of its length exposed, and attached to the implantation connector. The exposed tip is coated with a double membrane comprising Nafion and monoamine oxidase (MAO)‐B–diluted cellulose
2.2. Chemicals and drugs
DA, 5‐HT, NE, 3,4‐Dihydroxyphenylacetic acid (DOPAC), epinephrine (EPI), and acetylcholine (ACh) were purchased from Sigma‐Aldrich. Methamphetamine was obtained from Sumitomo Dainippon Pharma (Osaka, Japan). Tyrosine (TYR) was purchased from Katayama Chemicals (Osaka, Japan). L‐3,4‐Dihydroxyphenylalanine (L‐DOPA) and 4‐aminobutanoic acid (GABA) were obtained from Wako Pure Chemicals (Osaka, Japan). Methamphetamine powder was dissolved in 0.9% sterile saline at a concentration of 3 mg/ml. Fluoxetine and duloxetine were purchased from Sigma‐Aldrich. Both fluoxetine and duloxetine were dissolved in 0.9% saline (fluoxetine, 20 mg/ml; duloxetine, 16 mg/ml).
2.3. In vitro experiments
The electrodes were connected to the IMEC‐701 and immersed in 10 ml of the blank solution, phosphate‐buffered saline (PBS) at pH 7.4, at room temperature. Voltammetric analyses were carried out with 2 μl of 5 mM stock solutions of DA, 5‐HT, and NE added to the stirred PBS (1 μM) at 20‐s intervals.
To test the selectivity of the MAO‐B–containing probe, the responses to other neurotransmitters, DA precursors, and metabolites—TYR, DOPAC, L‐DOPA, EPI, Ach, and GABA—were assessed in the same manner as the three monoamines. To test the stability of the probe that was kept in sterile Dulbecco's Modified Eagle's Medium at 37 °C, the measurements were repeated 7 days after the initial measurements.
To test the need for both Nafion and cellulose in the electrode, the MAO‐B–free double membrane (Nafion/cross‐linked cellulose)–coated electrodes and the non–cross‐linked, MAO‐B–impregnated double membrane–coated electrodes were prepared and used for the experiments.
2.4. In vivo experiments
2.4.1. Animals
The in vivo responses of the coated electrodes were evaluated in adult male Wistar rats 8–9 months old, weighing 250–300 g (n = 15, Japan SLC Inc., Hamamatsu, Japan). The rats were housed in temperature‐controlled conditions (2) on a 12‐hr light/dark cycle with free access to food and water. We made every effort to minimize suffering and the number of animals used. All experimental procedures were approved by the committee on animal experimentation of Nagoya City University Medical School and were in accordance with the animal care guidelines of Nagoya University.
2.4.2. Electrode implantation
Rats were anesthetized with ketamine hydrochloride (70 mg/kg) and xylazine (10 mg/kg) and fixed in a stereotaxic frame. Dexamethasone (0.1 mg/kg; Banyu, Tokyo, Japan) was also administered intramuscularly to minimize brain edema. The scalp was cut at the midline and widely exposed, and six steel screws 0.5 mm in diameter were attached to the skull as anchors. The exposed skull was trepanated with a drill 1.2 mm rostral to the bregma and 2.0 mm lateral from the midline. The connector/electrode assembly was attached to a stereotaxic arm and lowered into the dorsal striatum, 5.5 mm below the surface. The connector and screws were cemented with dental acrylic to fix the electrode. After implantation, the rats were returned to their home cage to recover, with only one rat per cage to prevent damage to the implanted electrode. Positive‐ and negative‐control experiments were conducted 1 to 2 days after implantation. Chronic measurements were performed 7 and 14 days after surgery.
2.4.3. Positive‐control experiments
To evaluate the capacity of the novel electrode to measure the level of DA in vivo, DA release in the striatum was pharmacologically induced by methamphetamine injection to produce a tonic release, and electrically induced by stimulating the medial forebrain bundle to produce a phasic release (each n = 4). The medial forebrain bundle is a tract that includes axons from dopaminergic neurons in the ventral tegmental area.
Administration of methamphetamine
The electrode‐implanted rats were anesthetized with ketamine hydrochloride (70 mg/kg) and xylazine (10 mg/kg) and fixed in a stereotaxic frame. Carbon‐fiber and reference electrodes were connected to a voltammetric amplifier (IMEC‐701). A reference electrode was placed in the neck muscle of the rats. Recordings were initiated after allowing at least 30 min for stabilization of the animals and apparatus. We then administered a methamphetamine injection (3 mg/kg body weight, intraperitoneal) and attempted to measure the resultant DA upregulation in the striatum.
For chronic measurements, FSCV assays were conducted in the same manner at 7 and 14 days after electrode implantation.
Electrical stimulation of medial forebrain bundle
Anesthetized rats were fixed in a stereotaxic frame, and the skull was trepanated by drill 4.2 mm caudal to the bregma and 1.6 mm lateral from the midline. A glass‐insulated, tapered tungsten electrode (0.5–1 MΩ impedance at 1 kHz, diameter 310 mm; Alpha Omega Engineering, Nazareth, Israel) was lowered gently into the medial forebrain bundle, 8.5 mm below the brain surface. A series of 60‐ms, 60‐Hz bipolar pulses 2 ms in duration were applied, with the current controlled at 200 to 300 μA. The FSCV signals from the electrode were recorded before, during, and after stimulation.
2.4.4. Negative‐control experiments
A selective serotonin reuptake inhibitor (SSRI) or a serotonin and norepinephrine reuptake inhibitor (SNRI) was administered to the rats to increase 5‐HT and/or NE (n = 7). Because both of these drugs have potential DA‐releasing effects, the rats used in this section were prepared for FSCV testing by chemical lesioning of dopaminergic neurons in the substantia nigra, as follows (Hida et al., 2007).
Animals were anesthetized with ketamine hydrochloride (70 mg/kg) and xylazine (10 mg/kg) and fixed in a stereotaxic apparatus. The scalp was cut along the midline, and the muscle layers were retracted. A small hole was drilled in the skull, 3.8 mm rostral from the interaural line and 1.8 mm lateral from the midline. A 10‐μl 6‐hydroxydopamine solution, prepared by diluting the drug to 2 mg/ml in 0.9% sterile saline containing 0.5 mg/ml ascorbic acid, was injected into the substantia nigra, 7.8 mm below the brain surface. A glass micropipette connected to a Hamilton syringe was used to inject the solution at a rate of 1 μl/min over 5 min. The pipette was left in place for 5 min and was then pulled out slowly.
Two weeks after the 6‐hydroxydopamine injection, the rats were implanted with the novel electrode and connector. The electrode was implanted into the striatum ipsilateral to the 6‐hydroxydopamine injection site. Two days after surgery, either the SSRI fluoxetine (16 mg/kg) or the SNRI duloxetine (20 mg/kg) was injected intraperitoneally, and the FSCV measurements were conducted.
2.5. Statistical analysis
Statistical analyses were performed using JMP v.10 (JMP statistical software; SAS Institute, Cary, NC; http://www.jmp.com/en_us/software/jmp.html; RRID: SCR_014242). A significance level of p < .05 was chosen. Data were analyzed using 1‐way analysis of variance with post hoc Tukey test and Student t test. Paired t tests were used for in vitro repeated measurements. Data are presented as mean and standard error of the mean.
3. RESULTS
3.1. In vitro experiments
3.1.1. Sensitivity of the bare and double membrane–coated electrode to DA
To evaluate the DA selectivity of the novel electrode, we conducted in vitro and in vivo tests. First, we measured the sensitivity of the electrode to different doses of DA (0, 1, 2 μM) with or without the membrane coating. Cyclic voltammograms were collected in vitro by ramping the potential applied to the carbon‐fiber electrode from a holding potential of −0.4 V versus Ag/AgCl to 1.0 V and back every 100 ms, at 300 V/s. The current of bare electrode was 0.167 ± 0.000 μA (n = 3) at 1 μM and 0.318 ± 0.005 μA (n = 3) at 2 μM: The slope of dose–response curve was 0.159, and the linearity was 0.999. The current of the coated electrode was 0.141 ± 0.002 μA (n = 3) at 1 μM and 0.258 ± 0.004 μA (n = 3) at 2 μM: The slope was 0.129, and the linearity was 0.997.
3.1.2. Discrimination capacity versus other monoamines in DA measurement in vitro
Next, we compared the sensitivities of the electrodes to other monoamines (5‐HT, NE). We recorded the electrodes’ responses to the sequential addition of 1 μM DA, 5‐HT, and NE (Figure 2a–d). The noncoated carbon‐fiber electrodes showed clear responses to the addition of DA at an applied potential of 0.5 V, and small but substantial currents were detected with the addition of 5‐HT and NE (Figure 2a). The MAO‐B–impregnated double membrane–coated electrodes also detected the addition of 1 μM DA but barely responded to the addition of 5‐HT or NE (Figure 2b).
Figure 2.
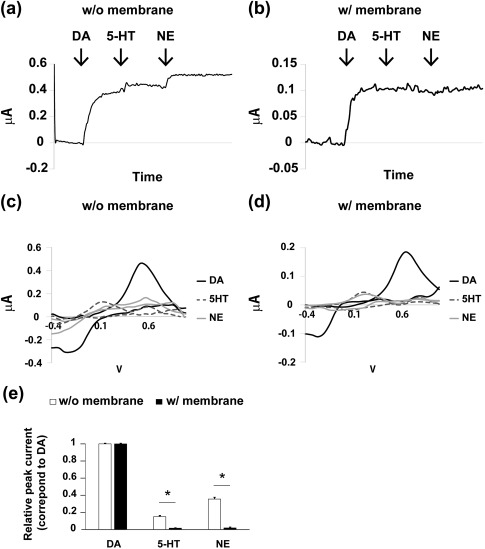
Discriminative capacity of the novel electrode with and without the double membrane in vitro. (a,b) I‐T plots for the addition of dopamine (DA), serotonin (5‐HT), and norepinephrine (NE) using (a) the bare electrode and (b) the coated electrode. (c,d) Representative, background‐subtracted cyclic voltammograms of DA, 5‐HT, and NE by (c) the noncoated and (d) the coated electrode. (e) Relative amplitude of the currents for DA, 5‐HT, and NE at the peak for DA. Student t test. * p < .001. Values are shown as mean ± standard error of the mean (SEM)
The background‐subtracted cyclic voltammogram showed that the peak current of DA was 0.456 μA at 0.521 V with the noncoated electrodes (Figure 2c). The voltammogram of DA with the MAO‐B–impregnated double membrane–coated electrode revealed that the oxidation peak current was 0.183 μA at 0.642 V (Figure 2d). In response to the addition of 5‐HT or NE, the bare noncoated electrodes detected considerable current at the oxidation potential of DA (5‐HT, 0.064 μA; NE, 0.137 μA; Figure 2c). In contrast, the double membrane–coated electrode responded much more weakly to 5‐HT and NE at 0.642 V (at the peak current of DA) than the bare electrodes (5‐HT, 0.003 μA; NE, 0.011 μA; Figure 2d).
The relative amplitudes of the currents in response to 5‐HT and NE at the oxidation potential of DA were significantly lower for the coated electrodes (5‐HT, 0.017% ± 0.002% of DA peak; NE, 0.020% ± 0.007%; Figure 2e) than for the bare electrodes (5‐HT, 0.151% ± 0.011%; NE, 0.356% ± 0.019%; each n = 5, p < .001, t test). These results indicate that the novel membrane‐coated electrode improved the signal‐to‐noise ratio of the DA measurements in the presence of other monoamines using FSCV in vitro.
3.1.3. Confirmation of the effects of the MAO‐B coating on DA selectivity
To increase the selectivity of the DA measurements, the probe was coated with MAO‐B–impregnated cellulose and Nafion membranes. As the MAO‐B enzyme has very low thermal stability (usually stored at −70 °C), and the membranes might affect DA permeability by low diffusion through membranes, we aimed to confirm the effectiveness and stability of our MAO‐B–coated probe.
We prepared two other types of electrodes: a glutaraldehyde–cross‐linked MAO‐B–free electrode double coated with cellulose and Nafion, and a non–cross‐linked MAO‐B–impregnated electrode double coated with cellulose and Nafion. We compared the sensitivities of these electrodes versus the monoamines, other neurotransmitters, and precursors and metabolites of DA (Figure 3).
Figure 3.

Confirmation of the efficiency and stability of the monoamine oxidase (MAO)‐B coating on the probe in vitro. (a–c) I‐T plots for the addition of dopamine (DA), serotonin (5‐HT), and norepinephrine (NE) using (a) the MAO‐B–free double membrane–coated electrodes; (b) the MAO‐B–impregnated, non–cross‐linked membrane–coated electrodes; and (c) the MAO‐B–impregnated, cross‐linked membrane–coated electrodes. (d–f) Representative, background‐subtracted cyclic voltammograms of DA, 5‐HT, and NE by (d) the MAO‐B–free membrane–coated electrodes, (e) the non–cross‐linked MAO‐B–coated electrodes, and (f) the cross‐linked MAO‐B–coated electrode. (g) Relative amplitudes of the currents for monoamines, other neurotransmitters, precursors, and metabolites of DA, corresponding to the peak for DA. TYR = tyrosine; DOPAC = 3,4‐dihydroxyphenylacetic acid; L‐DOPA = L‐3,4‐dihydroxyphenylalanine; EPI = epinephrine; Ach = acetylcholine; GABA = 4‐aminobutanoic acid. One‐way ANOVA with post hoc Tukey test. * p < .05. ** p < .001. Values are shown as mean ± standard error of the mean (SEM)
The double membrane–coated electrodes without MAO‐B showed clear responses to the addition of 5‐HT and NE at the oxidation potential of DA (Figure 3a). Background‐subtracted FSCV revealed that the peak currents were 0.485 ± 0.024 μA in DA (n = 5), 0.131 ± 0.018 μA in 5‐HT (n = 5), and 0.148 ± 0.013 μA in NE (n = 5) (Figure 3d). The non–cross‐linked MAO‐B–impregnated double‐coated electrodes showed substantial responses to the addition of 5‐HT and NE (Figure 3b). The FSCV peak currents were 0.424 ± 0.054 μA for DA (n = 5), 0.056 ± 0.016 μA for 5‐HT (n = 5), and 0.102 ± 0.008 μA for NE (n = 5) (Figure 3e). Compared with these electrodes, the cross‐linked MAO‐B–impregnated cellulose and Nafion double‐coated electrodes successfully suppressed the responses to 5‐HT and NE (Figure 3c). The FSCV peak currents were 0.416 ± 0.007 μA in DA (n = 5), 0.008 ± 0.003 μA in 5‐HT (n = 5), and 0.002 ± 0.001 μA in NE (n = 5) (Figure 3f).
The amplitudes of the currents in response to 5‐HT and NE relative to those to DA were significantly lower in the case of the cross‐linked MAO‐B–impregnated electrodes (5‐HT: 1.21% ± 0.32%, n = 5; NE: 0.91% ± 0.32%, n = 5) than in the case of the electrodes without MAO‐B (5‐HT: 25.03% ± 3.93%, n = 5, p < .001; NE: 28.85% ± 2.69%, n = 5, p < .001) or the non–cross‐linked MAO‐B–impregnated electrodes (5‐HT: 11.24% ± 2.15%, n = 5, p < .05; NE: 20.06% ± 1.75%, n = 5, p < .001) (Figure 3g). Moreover, the probes without MAO‐B and the cross‐linked MAO‐B–impregnated probes responded significantly differently to L‐DOPA (9.08% ± 2.42% vs. 1.00% ± 0.45%, p < .01; each n = 5) (Figure 3g).
Responses to other neurotransmitters such as ACh and GABA, the DA precursor TYR, and metabolites such as DOPAC and EPI were very small in the cross‐linked MAO‐B–impregnated cellulose and Nafion membrane electrode (Figure 3g). These results indicate that the improvement in the signal‐to‐noise ratio of the DA measurements was due to the cross‐linked MAO‐B coating on the electrodes.
3.1.4. Stability of the MAO‐B–coated electrode in vitro
We performed a series of measurements to confirm the stability of the cross‐linked MAO‐B–impregnated cellulose and Nafion membranes. After the first in vitro experiment, the electrode was kept in sterile culture medium for 7 days at 37 °C to mimic in vivo conditions, and the serial in vitro experiments were then performed using the same electrode.
The electrode responded to the addition of DA but showed minimal reaction to the addition of either 5‐HT or NE 7 days after the initial measurements (Figure 4a). Background‐subtracted FSCV showed that the peak currents were 0.422 ± 0.086 μA in DA (n = 4), 0.005 ± 0.000 μA in 5‐HT (n = 4), and 0.006 ± 0.000 μA in NE (n = 4) (Figure 4b). The amplitudes of the currents of 5‐HT and NE were quite small compared with those of DA (5‐HT: 1.05% ± 0.14%, n = 4; NE: 1.35% ± 0.23%, n = 4), showing that the DA selectivity of the electrode was maintained for at least 7 days (Figure 4c). There were no significant differences in the responses to monoamines, other neurotransmitters, DA precursors, or metabolites between day 1 and day 7 (each n = 4, p < .01).
Figure 4.

Chronic measurements in vitro. (a) I‐T and (b) I‐V representative plots for the addition of dopamine (DA), serotonin (5‐HT), and norepinephrine (NE) using the cross‐linked, monoamine oxidase (MAO)‐B–impregnated membrane–coated electrodes 7 days after the initial measurements. (c) The absolute amplitudes of the currents for monoamines, other neurotransmitters, DA precursors, and metabolites at the peak for DA. Values are shown as mean ± standard error of the mean (SEM)
These results indicate that the cross‐linked MAO‐B–impregnated cellulose membrane remained stable at 37 °C for at least 7 days in vitro.
3.2. In vivo experiments
3.2.1. Positive‐control experiments: Administration of a DA‐releasing drug and stimulation of a dopaminergic tract
To investigate whether the probe could selectively detect changes in the DA content in vivo, the electrode was implanted in the striatum to detect both the tonic change in the DA level induced by methamphetamine administration and the phasic change induced by electrical stimulation of the medial forebrain bundle (Figure 5).
Figure 5.

Positive‐control experiments in vivo. (a,b) Fast‐scan cyclic voltammetry (FSCV) in the rat striatum following administration of methamphetamine: (a) the I‐T plot and (b) the I‐V plot at 1,800 s post administration. (c,d) FSCV analysis during stimulation of the medial forebrain bundle: (c) the I‐T plot and (d) the I‐V plot at current peak
About 5 to 7 min after methamphetamine administration, the DA oxidative current started to increase (Figure 5a). The timing of the increase in the DA current was in line with published reports of methamphetamine‐induced behavioral changes (Cadet et al., 2010; Morisset et al., 2002). The background‐subtracted FSCV current gradually increased for at least 30 min post administration (Figure 5a) and returned to the baseline over the next 3 hr. The background‐subtracted peak current was 0.026 μA at 0.581 V (Figure 5b).
Next, we stimulated the medial forebrain bundle. The DA current rapidly increased with the stimulation, and then decreased to the prestimulation level within a few seconds (Figure 5c). The background‐subtracted peak FSCV current was 0.058 μA at 0.566 V and occurred after 2.3 ± 0.7 s (Figure 5d).
Taken together, the results show that the coated electrode successfully detected the upregulation of extracellular DA in the striatum of rats. The electrode responded to both tonic and phasic alterations of DA levels.
3.2.2. Negative‐control experiments: administration of 5‐HT/NE RIs
To evaluate the capacity of the coated electrode to discriminate non‐DA monoamines in vivo, the electrode was implanted into the 6‐hydroxydopamine–lesioned striatum, followed by systemic injections of SSRIs or SNRIs (Figure 6).
Figure 6.

Negative‐control experiments in vivo. The rats were prepared by lesioning the substantia nigra. Fast‐scan cyclic voltammetry (FSCV) analyses were carried out following the administration of (a,b) a serotonin selective reuptake inhibitor or (c,d) a serotonin/norepinephrine reuptake inhibitor
Neither the SSRI nor the SNRI produced any effect (Figure 6a,c). The background‐subtracted FSCV revealed no current in response to 5‐HT or NE (Figure 6b,d). After the experiments, methamphetamine was administered to confirm the deletion of dopaminergic neurons in the rats. We found no obvious current detected up to at least 30 min after methamphetamine treatment (data not shown).
These results demonstrate that the coated electrode was much less sensitive to 5‐HT and NE than to DA. Therefore, the newly designed electrode has high selectivity for DA.
3.3. Chronic DA measurement in vivo
To investigate the suitability of the electrode for chronic studies, the implanted rats were rested for 7 or 14 days following implantation, and the methamphetamine‐induced augmentation of DA was then recorded (Figure 7).
Figure 7.

Chronic measurements in vivo. (a) I‐T and (b) I‐V plots obtained in rat striatum by fast‐scan cyclic voltammetry (FSCV) assay with the novel biosensor, shortly after administration of methamphetamine at 7 days post implantation
At 7 days, the FSCV current measured with the novel electrode increased gradually for at least 30 min post administration, indicating successful detection of the methamphetamine‐induced continuous release of DA (Figure 7a). The current–voltage plot was very similar to that at 1 day after implantation: The peak current was 0.06 μA at 0.581 V (Figure 7b). At 14 days, however, the baseline became irregular, and atypical voltammograms were recorded (data not shown).
These results indicate that the coated electrode could detect the DA signal at least 1 week after implantation, but not after 2 weeks.
4. DISCUSSION
In the present study, we developed a novel electrode for use with the FSCV assay to improve its chemical selectivity to DA and thus improve its measurement. We coated a carbon‐fiber electrode with a double membrane consisting of Nafion and MAO‐B–diluted cellulose, with the aim of discriminating DA from other monoamines such as 5‐HT and NE. FSCV using the coated electrode demonstrated satisfactory sensitivity and selectivity for DA both in vitro and in vivo. The present system responded to both slow and rapid increases in DA but not to increases in 5‐HT or NE. In addition, the coated electrode was able to measure the DA signal in vivo after at least 1 week of implantation. These results indicate the advantage of the novel MAO‐B–impregnated electrode in the selective detection of DA in the presence of other monoamine neurotransmitters.
Dopaminergic neurons are found mainly in the substantia nigra pars compacta and in the ventral tegmental area, and these nuclei broadly project their axons to the prefrontal cortex, sensorimotor cortex, striatum, and limbic system, where they modulate neural activity (Fallon, 1981; Grace, Floresco, Goto, & Lodge, 2007; Lindvall et al., 1974). DA is an important monoamine neurotransmitter involved in various forms of behavior, including motor control, the reward system, and motivation and learning. Alteration of the firing pattern of dopaminergic neurons and of DA release could thus have wide‐ranging effects on behavior (Bromberg‐Martin et al., 2010; Floresco & Magyar, 2006; Hosp et al., 2011; McNamara et al., 2014). Therefore, there is an urgent need for highly accurate measurement of DA. FSCV is a promising measurement method because of its high temporal resolution (Robinson et al., 2003). However, the capacity of FSCV to discriminate between different monoamines is a major issue. The difficulty is that DA and 5‐HT possess quite similar current responses and oxidation potentials, making them difficult to discriminate (Kovach et al., 1984). Discriminating between DA and NE seems to be even more difficult (Anastassiou et al., 2006). DA is a direct precursor of NE, and their chemical properties are very similar, which means that their voltammograms are almost identical (Yoshimi, Kumada, Weitemier, Jo, & Inoue, 2015). DA, 5‐HT, and NE are therefore difficult to discriminate in electrochemical assays.
To overcome the selectivity problem, we devised a solution that discriminated DA from 5‐HT and NE by means of cross‐linked MAO‐B–impregnated cellulose and Nafion–double‐coated electrodes. MAO‐B inhibitors are typically used in DA measurements to prevent the metabolism of the released DA (Burrell, Atcherley, Heien, & Lipski, 2015; Crespi, 1990), although MAO‐A and MAO‐B metabolize DA equally (Finberg & Rabey, 2016). In our preliminary experiment, we failed to suppress the responses to 5‐HT and NE using MAO‐A–coated electrodes (unpublished data). However, we clearly showed that the cross‐linked MAO‐B–coated electrode discriminated DA from 5‐HT and NE, as shown in Figure 3. It is notable that MAO‐B cross‐linking is a key step in ensuring high selectivity for DA. It is probable that MAO‐B cross‐linking changed both the catalytic properties of MAO‐B and its long‐term stability. A base‐hydrolyzed cellulose membrane was used as a container for the MAO‐B; it also served as a fouling‐resistant coating membrane for the electrodes because of its ability to exclude large biomolecules (Wang & Hutchins, 1985). Nafion is a perfluorosulfonate proton‐exchange membrane polymer that allows only DA, which is cationic near neutrality, to reach the electrode, while blocking ascorbic acid, which is anionic near neutrality (Burmeister et al., 2008; Hu & Wilson, 1997). Ascorbic acid disturbs the detection of DA and cannot be neglected because high concentrations of ascorbate are present in the rodent striatum (Ewing, Wightman, & Dayton, 1982).
Any surface modification (except nanoparticles) will decrease the electrode response: The significant difference of the slope in dose–curve response between bare electrode and the coated electrode might indicate a diffusion barrier due to the two membranes. It should also be noted that the response of MAO‐B–impregnated electrodes could be inhibited by numerous substances such asβ‐carboline or neurologic drugs in vivo, and these effects were not investigated in our study.
The implantation of electrodes for chronic measurement in waking animals is an important use of FSCV that takes advantage of its fine temporal resolution. Here, the coated electrodes were implanted into the rats’ striatum to provide in vivo measurements. The MAO‐B–coated electrode worked well for at least 1 week after implantation, showing that it is useful for chronic DA measurements. However, all recordings in this study were conducted in anesthetized rather than awake animals. A high signal‐to‐noise ratio is required for measurements in awake animals because of the additional noise caused by body motion. In addition, more complex neural activities are likely to occur in the awake brain than the anesthetized brain. The present electrode achieved a substantial improvement in selectivity for DA versus other monoamines and in signal‐to‐noise ratio. Thus, the electrode might be usable for chronic measurements in awake animals. However, by 2 weeks post implantation, unstable baseline and atypical voltammograms emerged, possibly due to fouling of the electrode by radical species, damage to the membrane, and/or formation of glial scarring around the electrode. Although most of these problems seem to be unavoidable in electrode implantation experiments, further experiments should be conducted on the development of a novel electrode with higher durability and longer service life in situ.
It should be noted that the immersion of the biosensor for 7 days in culture medium at 37 °C had few effects on sensitivity. The probable explanation of this in vitro different response to the in vivo response is probably due to the inhibitory effect by glial responses in the host brain.
An important advantage of the novel coated electrode was that it increased the signal‐to‐noise ratio for detecting DA in the presence of other monoamines, rather than merely enhancing the sensitivity to DA. Although the sensitivities of the bare and coated electrodes were comparable at the calibration stage, it is possible that the general sensitivity of the coated electrode decayed faster. Although we confirmed the stability of the cross‐linked, MAO‐B–coated electrodes in vitro, the long‐term activity of the cross‐linked MAO‐B remains unclear. This point might be important, especially in chronic experiments. Further examination of long‐term changes in the enzymatic activity of MAO‐B is needed. A potential limitation of the present method‐validation study is that it was not possible to compare the results for male and female rats because only male rats were used.
We conclude that it is possible to enhance the FSCV measurement of DA using a newly developed double‐layered, membrane‐coated electrode that includes MAO‐B enzymes to increase selectivity for DA. The system successfully detected continuous and phasic increases in DA while decreasing the undesirable detection of 5‐HT and NE, both in vitro and in vivo. The present study validates a new strategy and insight—namely, the enzyme‐based discrimination of DA in FSCV.
CONFLICT OF INTEREST
The authors declare that they have no conflicts of interest.
AUTHOR CONTRIBUTIONS
All authors had full access to all the data in the study and take responsibility for the integrity of the data and the accuracy of the data analysis. Conceptualization and Methodology: A.I. (Imamura) and H.H. Software and Resources: A.I. (Imamura). Investigation and Formal Analysis: A.I., A.I., Y.U. R.M. T.S. and H.H. Writing ‐ Original Draft: AI (Ishida). Writing ‐ Review & Editing: AI (Imamura) and HH. Funding Acquisition: A.I. (Ishida) and H.H. Supervision: HH.
ACKNOWLEDGMENTS
The authors thank K. Yoshimi for providing technical support with the experiments. We also thank H. Aizawa for contributing useful discussions. We thank Rachel Baron, PhD, from Edanz Group (www.edanzediting.com/ac) for editing a draft of this manuscript.
Ishida A, Imamura A, Ueda Y, et al. A novel biosensor with high signal‐to‐noise ratio for real‐time measurement of dopamine levels in vivo. J Neuro Res. 2018;96:817–827. https://doi.org/10.1002/jnr.24193
Significance Real‐time measurement of dopamine (DA) in vivo is an important technique in neuroscience research, especially research on movement disorders, addiction, and some forms of learning. However, it is hard to discriminate DA from other monoamines using cyclic voltammetry because of their electrochemical similarities. To overcome this problem, we developed an implantable double‐layered electrode. The biosensor can discriminate DA from other monoamines using monoamine oxidase B diluted in the cellulose membrane, thus allowing the highly selective and chronic measurement of DA in vitro and in vivo. The novel biosensor is a promising tool to facilitate further research into the physiology of DA.
REFERENCES
- Alexander, G. E. , DeLong, M. R. , & Strick, P. L. (1986). Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annual Review of Neuroscience, 9, 357–381. [DOI] [PubMed] [Google Scholar]
- Anastassiou, C. A. , Patel, B. A. , Arundell, M. , Yeoman, M. S. , Parker, K. H. , & O'Hare, D. (2006). Subsecond voltammetric separation between dopamine and serotonin in the presence of ascorbate. Analytical Chemistry, 78, 6990–6998. [DOI] [PubMed] [Google Scholar]
- Bourdy, R. , & Barrot, M. (2012). A new control center for dopaminergic systems: Pulling the VTA by the tail. Trends in Neurosciences, 35, 681–690. [DOI] [PubMed] [Google Scholar]
- Bromberg‐Martin, E. S. , Matsumoto, M. , & Hikosaka, O. (2010). Dopamine in motivational control: Rewarding, aversive, and alerting. Neuron, 68, 815–834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burmeister, J. J. , Pomerleau, F. , Huettl, P. , Gash, C. R. , Werner, C. E. , Bruno, J. P. , & Gerhardt, G. A. (2008). Ceramic‐based multisite microelectrode arrays for simultaneous measures of choline and acetylcholine in CNS. Biosensors & Bioelectronics, 23, 1382–1389. [DOI] [PubMed] [Google Scholar]
- Burrell, M. H. , Atcherley, C. W. , Heien, M. L. , & Lipski, J. (2015). A novel electrochemical approach for prolonged measurement of absolute levels of extracellular dopamine in brain slices. ACS Chemical Neuroscience, 6, 1802–1812. [DOI] [PubMed] [Google Scholar]
- Cadet, J. L. , Brannock, C. , Krasnova, I. N. , Ladenheim, B. , McCoy, M. T. , Chou, J. , … Wang, Y. (2010). Methamphetamine‐induced dopamine‐independent alterations in striatal gene expression in the 6‐hydroxydopamine hemiparkinsonian rats. PLoS One, 5, e15643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crespi, F. (1990). In vivo voltammetry with micro‐biosensors for analysis of neurotransmitter release and metabolism. Journal of Neuroscience Methods, 34, 53–65. [DOI] [PubMed] [Google Scholar]
- Crespi, F. , England, T. G. , & Trist, D. G. (1995). Simultaneous, selective detection of catecholaminergic and indolaminergic signals using cyclic voltammetry with treated micro‐sensor. Journal of Neuroscience Methods, 61, 201–212. [DOI] [PubMed] [Google Scholar]
- Ewing, A. G. , Wightman, R. M. , & Dayton, M. A. (1982). In vivo voltammetry with electrodes that discriminate between dopamine and ascorbate. Brain Research, 249, 361–370. [DOI] [PubMed] [Google Scholar]
- Fallon, J. H. (1981). Collateralization of monoamine neurons: Mesotelencephalic dopamine projections to caudate, septum, and frontal cortex. Journal of Neuroscience, 1, 1361–1368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finberg, J. P. , & Rabey, J. M. (2016). Inhibitors of MAO‐A and MAO‐B in psychiatry and neurology. Frontiers in Pharmacology, 7, 340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Floresco, S. B. , & Magyar, O. (2006). Mesocortical dopamine modulation of executive functions: Beyond working memory. Psychopharmacology, 188, 567–585. [DOI] [PubMed] [Google Scholar]
- Fortin, S. M. , Cone, J. J. , Ng‐Evans, S. , McCutcheon, J. E. , & Roitman, M. F. (2015). Sampling phasic dopamine signaling with fast‐scan cyclic voltammetry in awake, behaving rats. Current Protocols in Neuroscience, 70, 7.25.1‐20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garris, P. A. , Walker, Q. D. , & Wightman, R. M. (1997). Dopamine release and uptake rates both decrease in the partially denervated striatum in proportion to the loss of dopamine terminals. Brain Research, 753, 225–234. [DOI] [PubMed] [Google Scholar]
- Grace, A. A. , & Bunney, B. S. (1983). Intracellular and extracellular electrophysiology of nigral dopaminergic neurons—1. Identification and characterization. Neuroscience, 10, 301–315. [DOI] [PubMed] [Google Scholar]
- Grace, A. A. , Floresco, S. B. , Goto, Y. , & Lodge, D. J. (2007). Regulation of firing of dopaminergic neurons and control of goal‐directed behaviors. Trends in Neuroscience, 30, 220–227. [DOI] [PubMed] [Google Scholar]
- Hafizi, S. , Kruk, Z. L. , & Stamford, J. A. (1990). Fast cyclic voltammetry: Improved sensitivity to dopamine with extended oxidation scan limits. Journal of Neuroscience Methods, 33, 41–49. [DOI] [PubMed] [Google Scholar]
- Heien, M. L. , Phillips, P. E. , Stuber, G. D. , Seipel, A. T. , & Wightman, R. M. (2003). Overoxidation of carbon‐fiber microelectrodes enhances dopamine adsorption and increases sensitivity. Analyst, 128, 1413–1419. [DOI] [PubMed] [Google Scholar]
- Hida, H. , Masuda, T. , Sato, T. , Kim, T. S. , Misumi, S. , & Nishino, H. (2007). Pleiotrophin promotes functional recovery after neural transplantation in rats. Neuroreport, 18, 179–183. [DOI] [PubMed] [Google Scholar]
- Hosp, J. A. , Pekanovic, A. , Rioult‐Pedotti, M. S. , & Luft, A. R. (2011). Dopaminergic projections from midbrain to primary motor cortex mediate motor skill learning. Journal of Neuroscience, 31, 2481–2487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu, Y. , & Wilson, G. S. (1997). Rapid changes in local extracellular rat brain glucose observed with an in vivo glucose sensor. Journal of Neurochemistry, 68, 1745–1752. [DOI] [PubMed] [Google Scholar]
- Jung, M. C. , Shi, G. , Borland, L. , Michael, A. C. , & Weber, S. G. (2006). Simultaneous determination of biogenic monoamines in rat brain dialysates using capillary high‐performance liquid chromatography with photoluminescence following electron transfer. Analytical Chemistry, 78, 1755–1760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kovach, P. M. , Ewing, A. G. , Wilson, R. L. , & Wightman, R. M. (1984). In vitro comparison of the selectivity of electrodes for in vivo electrochemistry. Journal of Neuroscience Methods, 10, 215–227. [DOI] [PubMed] [Google Scholar]
- Lindvall, O. , Bjorklund, A. , Moore, R. Y. , & Stenevi, U. (1974). Mesencephalic dopamine neurons projecting to neocortex. Brain Research, 81, 325–331. [DOI] [PubMed] [Google Scholar]
- McNamara, C. G. , Tejero‐Cantero, A. , Trouche, S. , Campo‐Urriza, N. , & Dupret, D. (2014). Dopaminergic neurons promote hippocampal reactivation and spatial memory persistence. Nature Neuroscience, 17, 1658–1660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morisset, S. , Pilon, C. , Tardivel‐Lacombe, J. , Weinstein, D. , Rostene, W. , Betancur, C. , … Arrang, J. M. (2002). Acute and chronic effects of methamphetamine on tele‐methylhistamine levels in mouse brain: Selective involvement of the D(2) and not D(3) receptor. Journal of Pharmacology and Experimental Therapeutics, 300, 621–628. [DOI] [PubMed] [Google Scholar]
- Phillips, P. E. , Robinson, D. L. , Stuber, G. D. , Carelli, R. M. , & Wightman, R. M. (2003). Real‐time measurements of phasic changes in extracellular dopamine concentration in freely moving rats by fast‐scan cyclic voltammetry. Methods in Molecular Medicine, 79, 443–464. [DOI] [PubMed] [Google Scholar]
- Ramsson, E. S. , Cholger, D. , Dionise, A. , Poirier, N. , Andrus, A. , & Curtiss, R. (2015). Characterization of fast‐scan cyclic voltammetric electrodes using paraffin as an effective sealant with in vitro and in vivo applications. PLoS One, 10, e0141340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson, D. L. , Venton, B. J. , Heien, M. L. , & Wightman, R. M. (2003). Detecting subsecond dopamine release with fast‐scan cyclic voltammetry in vivo. Clinical Chemistry, 49, 1763–1773. [DOI] [PubMed] [Google Scholar]
- Wang, J. , & Hutchins, L. D. (1985). Thin‐layer electrochemical detector with a glassy carbon electrode coated with a base‐hydrolyzed cellulosic film. Analytical Chemistry, 57, 1536–1541. [DOI] [PubMed] [Google Scholar]
- Wu, Q. , Reith, M. E. , Wightman, R. M. , Kawagoe, K. T. , & Garris, P. A. (2001). Determination of release and uptake parameters from electrically evoked dopamine dynamics measured by real‐time voltammetry. Journal of Neuroscience Methods, 112, 119–133. [DOI] [PubMed] [Google Scholar]
- Yoshimi, K. , Kumada, S. , Weitemier, A. , Jo, T. , & Inoue, M. (2015). Reward‐induced phasic dopamine release in the monkey ventral striatum and putamen. PLoS One, 10, e0130443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zachek, M. K. , Hermans, A. , Wightman, R. M. , & McCarty, G. S. (2008). Electrochemical dopamine detection: Comparing gold and carbon fiber microelectrodes using background subtracted fast scan cyclic voltammetry. Journal of Electroanalytical Chemistry, 614, 113–120. [DOI] [PMC free article] [PubMed] [Google Scholar]