

Abstract
Objectives
The present study aimed to investigate the overall effect of quercetin on mouse bone marrow mesenchymal stem cell (BMSC) proliferation and osteogenic differentiation in vitro.
Materials and Methods
BMSCs were treated with different concentrations of quercetin for 6 days. The effects of quercetin on cell proliferation were assessed at predetermined times using Cell Counting Kit-8 (CCK-8) assay. The cells were then treated with quercetin, estrogen, or an estrogen receptor (ER) antagonist (which was also administered in the presence of quercetin or estrogen) for 7 or 21 days. The effects of quercetin on BMSC osteogenic differentiation were analyzed by an alkaline phosphatase (ALP) assay kit, Alizarin Red S staining (ARS), quantitative real-time PCR (qPCR), and western blotting.
Results
The CCK-8 and ALP assays and ARS staining showed that quercetin significantly enhanced BMSC proliferation, ALP activity, and extracellular matrix production and mineralization, respectively. The qPCR results indicated that quercetin promoted osterix (OSX), runt-related transcription factor 2 (RUNX2), and osteopontin (OPN) transcription in the presence of osteoinduction medium, and the western blotting results indicated that quercetin enhanced bone morphogenetic protein 2 (BMP2), Smad1, Smad4, RUNX2, OSX, and OPN expression and Smad1 phosphorylation. Treatment with the ER inhibitor ICI182780 blocked the effects of quercetin.
Conclusions
Our data demonstrated that quercetin promotes BMSC proliferation and osteogenic differentiation. Quercetin enhances BMP signaling pathway activation and upregulates the expression of downstream genes, such as OSX, RUNX2, and OPN, via the ER.
1. Introduction
Mesenchymal stem cells are a class of cells that exist in multiple organs and retain the capacity for self-renewal and differentiation into multilineage tissues for life [1, 2]. Bone marrow mesenchymal stem cells (BMSCs), which are isolated from the bone marrow, are regarded as seed cells for bone tissue engineering because of their great multidirectional differentiation ability and reproductive activity [3–5]. BMSCs reportedly give rise to osteogenic, chondrogenic, and adipogenic cells, which are widely present in the bone marrow and cancellous bone and play an important role in bone metabolism. Due to these properties, BMSCs are a potential source of cells for cell and gene therapy experiments and are widely used in studies of bone regeneration [6, 7].
Estrogen is the key regulator of bone metabolism [8]. Menopause and the accompanying loss of ovarian estrogen are associated with declines in bone mineral density [9]. The protective effects of estrogen on bone in postmenopausal women were initially thought to be mediated by the suppression of bone resorption. However, recent studies have demonstrated that in addition to inhibiting osteoclast development, estrogen also promotes osteogenesis [10, 11]. Khosla et al. proposed that estrogen decreases bone resorption and maintains bone formation by exerting direct effects on osteocytes, osteoblasts, and osteoclasts [12], a process that requires the participation of the estrogen receptors (ERs) expressed on osteoblasts [13–15]. Bone morphogenetic protein (BMP), a pleiotropic cytokine belonging to the TGF-β super family, has osteogenic properties. Runt-related transcription factor 2 (RUNX2) and osterix (OSX) are the transcription factors required for osteoblastogenesis. BMP binds heterodimeric receptors to activate Smad proteins, which transactivate osteoblastogenic genes either directly or via RUNX2/OSX [16, 17]. These pathways do not work independently. Researchers have noted that strong cross-talk occurs among BMP, estrogen, and other signaling pathways. Previous studies have indicated that estrogen upregulates BMP2 expression and enhances BMP signaling pathway activation [10, 18].
Quercetin is one of the principal flavonoids and the most widely studied product of its type because of its pharmacological properties and beneficial health effects [19]. A variety of studies have begun amassing evidence regarding the anti-inflammatory [20], anticarcinogenic [21], neuroprotective, cardioprotective, and chemopreventive effects of quercetin [22]. Horcajada-Molteni et al. [23] demonstrated that rutin, a glycoside derivative of quercetin, inhibits ovariectomy-induced osteopenia in female rats. Moreover, several recent studies [6, 19, 24, 25] have indicated that quercetin promotes osteoblast differentiation. However, the mechanism by which quercetin induces osteogenic differentiation remains unclear.
Flavonoids, which are widely used in traditional Chinese medicine [17], chemically resemble estrogen. Some flavonoids have been used as estrogen substitutes, and several flavonoids, including icariin [26], genistein [27], and kaempferol [28], have been shown to promote osteogenic differentiation via their estrogen effects. As the structures of these flavonoids are similar to that of quercetin [29], we surmised that quercetin also has estrogenic effects on BMSCs and promotes osteogenic differentiation via the ER. In the present study, we investigated the estrogenic and osteogenic effects of quercetin on cultured BMSCs and elucidated the mechanisms by which the flavonoid exerts its effects. We compared ALP activity levels, OSX and OPN transcription levels, and osteogenic differentiation protein marker expression levels among quercetin-, estrogen-, and ER antagonist (ICI182780)-treated groups after osteogenic induction to analyze the effects of quercetin on cell proliferation.
2. Materials and Methods
2.1. BMSC and Quercetin Preparation
A frozen vial of mouse BMSCs isolated from C57BL/6 mice was purchased from Cyagen Biosciences Inc. (Guangzhou, China). The cells were thawed and then expanded in the complete medium (CM), that is, DMEM (Sigma–Aldrich, St. Louis, MO, USA) supplemented with 10% fetal bovine serum (FBS) (Elite Biotech, Heidelberg, GER) and 1% penicillin–streptomycin solution (Sigma–Aldrich, St. Louis, MO, USA), at 37°C in a humidified atmosphere of 95% air/5% CO2.
Quercetin was purchased from the National Institute for the Control of Pharmaceutical and Biological Products (Beijing, China) and was dissolved in DMSO (Sigma–Aldrich, St. Louis, MO, USA) to obtain a 20 mM stock solution, which was ultimately diluted in medium to obtain solutions of the desired concentrations. The control group was treated with additional DMSO to eliminate the effect of the reagent on the results. The final concentration of DMSO in the control and experimental groups was 0.1%.
2.2. Cell Proliferation Assay
Cell Counting Kit-8 (CCK-8) (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) was used to determine the effect of quercetin on cell proliferation. BMSCs were seeded in 96 well plates at a density of 2∗10∧4 cells/well and cultured in CM. The cells were cultured for 24 hours, after which they were treated with quercetin at the indicated concentrations (0-, 0.1-, 1-, 2.5-, and 5-μM) before being cultured for an additional 6 days. The 0 μM group, which was treated with DMSO, served as a control group. The cells were treated with 100 μl of fresh medium and 10 μl of CCK-8 solution at predetermined time and then incubated for 2 h. Enzyme-linked immunosorbent assay (ELISA) was used to measure the optical density (OD) values at a wavelength of 450 nm. The relative cell proliferation rate was calculated using the mean rate of the control group.
2.3. Alkaline Phosphatase (ALP) Activity Assay
ALP activity was evaluated using a 4-nitrophenyl phosphate colorimetric assay. BMSCs were seeded in 6-well plates at a density of 1∗10∧5 cells/well and cultured in CM until they reached 60% confluence. The cells were then divided into the following seven experimental groups: a negative control group (control), five quercetin-treated groups, and one estrogen-treated group. Cells cultured in CM alone served as a negative control group, and cells cultured in OIM with 10 nM 17ß-estrogen (E2) (Sigma–Aldrich, St. Louis, MO, USA) served as a positive control group. The BMSCs in the quercetin-treated groups were cultured in OIM containing 0-, 0.1-, 1-, 2.5-, and 5-μM quercetin, respectively, for 7 days. We made OIM by dissolving dexamethasone (10 nM), ascorbic acid (50 μg/ml), and ß-glycerophosphate (10 mM) (Sigma–Aldrich, St. Louis, MO, USA) in CM. ALP activity was quantified in cell lysates using an ALP assay kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China), according to the manufacturer's instructions. Each experiment was repeated twice; thus, the results are the means of three experiments.
2.4. Matrix Mineralization Assay
Extracellular matrix mineralization was observed by Alizarin Red S (ARS) staining. As in the ALP assay, BMSCs were seeded in 6-well plates at a density of 1∗10∧5 cells/well. After reaching confluence, the cells were incubated in CM or OIM containing quercetin or estrogen for 21 days. At the end of the incubation period, the cells were washed with phosphate-buffered saline (PBS) and fixed in 4% polysorbate for 4 h. The cells were then incubated with ARS (0.4%, pH 4.2, Sigma) for 30 min at room temperature.
2.5. Quantitative Real-Time PCR (qPCR)
The effects of quercetin and E2 on RUNX2, OSX, and OPN mRNA expression were investigated by real-time PCR. The cells were treated with the above reagents in the absence or presence of ICI182780 (an ER inhibitor) for 7 days, after which total RNA was extracted with TRIzol reagent (Invitrogen Corp., Carlsbad, CA, USA), and cDNA was prepared with a Transcriptor First-Strand cDNA Synthesis Kit (Thermo Fisher Scientific Inc., Waltham, MA, USA). For real-time PCR, a 20 μl reaction mixture containing 10 μl of SYBR Green Real-time PCR Master Mix (Toyobo Biotech Co., Ltd., Osaka, JP), cDNA, and primers was thermocycled in an ABI Step One Plus Real-time PCR System Thermal Cycler (Applied Biosystems Inc., Carlsbad, CA, USA). The primers, whose sequences are shown in Table 1, were purchased commercially (GeneScript Co., Ltd., Nanjing, China). The expression of RUNX2, OSX, and OPN gene was evaluated according to the threshold cycle (Ct) values and was normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene expression.
Table 1.
Sequences of the primers of the target and housekeeping genes used for RT-PCR.
| Sequence (5′-3′) | Length of product | |
|---|---|---|
| GAPDH | Sense primer: 5-GGCCTTCCGTGTTCCTACC-3 | 103 bp |
| Antisense primer: 5-TGCCTGCTTCACCACCTTC-3 | ||
| RUNX2 | Sense primer: 5-CCAAGTAGCCAGGTTCAACG-3 | 64 bp |
| Antisense primer: 5-GGTGAAACTCTTGCCTCGTC-3 | ||
| OSX | Sense primer: 5-CTTTCGTCTGCAACTGGCTT-3 | 123 bp |
| Antisense primer: 5-TAAAGCGCTTGGAACAGAGC-3 | ||
| OPN | Sense primer: 5-CAGCCATGAGTCAAGTCAGC-3 | 116 bp |
| Antisense primer: 5-TGTGGCTGTGAAACTTGTGG-3 |
2.6. Western Blot Analysis
The ER-mediated effects of quercetin on BMP signaling pathway activation and maker protein expression were investigated by western blotting analysis. BMSCs were treated with 2.5 μM quercetin or vehicle control in the absence or presence of 1 μM ICI182780 for 7 days, after which the expression of BMP2, Smad1, p-Smad1, Smad4, RUNX2, OSX, and OPN protein was assessed. For western blotting analysis, the cells were lysed in cold Nonidet P-40 (NP-40) lysis buffer (pH 7.6) containing 50 mM Tris–HCl, 150 mM NaCl, 10% glycerol, 1% NP-40, 1 mM phenylmethylsulfonyl fluoride, 1 μg/ml leupeptin, 1 μg/ml aprotinin, and 1 μg/ml pepstatin for 15 min at 4°C. The cell lysates were collected by scraping and then centrifuged at 14,000g for 10 min. Protein concentrations were assessed using BCA protein assay reagent (Jiangsu Keygen Biotech Corp., Nanjing, China). Twenty micrograms of whole-cell lysates was electrophoresed on an SDS-polyacrylamide gel before being transferred to a PVDF membrane (Jiangsu Keygen Biotech Corp., Nanjing, China), which was blocked in PBS containing 6% low-fat milk and 0.1% Tween 20 (PBST). The blots were then incubated with RUNX2-, BMP2-, Smad1-, p-Smad1-, Smad4-, OSX-, or OPN-antibodies (Jiangsu Keygen Biotech Corp., Nanjing, China), washed twice with PBST, and probed with horseradish peroxidase-conjugated goat anti-mouse secondary antibodies. The protein bands were visualized using a G:BOXChemiXR5 System (Syngene, Cambridge, UK), and the western blotting results were quantified using Gel-Pro32 (Media Cybernetics).
2.7. Statistical Analysis
Quantitative data are expressed as the mean ± standard deviation (SD) of three separate experiments and were analyzed by one-way ANOVA using SPSS 18.0. P values less than 0.05 were considered significant.
3. Results
3.1. Quercetin Enhanced Cell Proliferation
Cell proliferation was assessed using CCK-8 assay, as shown in Figure 1. The ODs in the control and quercetin groups increased during the first 5 days of the experiment and then decreased on day 6 of the experiment. The ODs in the 0.1 μM and 1 μM quercetin groups increased significantly compared with those in the control group on the first day of the experiment. We noted no significant differences in OD between the 2.5 μM and 5 μM quercetin groups and the control group. Treatment with 0.1 μM–5 μM quercetin significantly stimulated cell proliferation compared with treatment with the control on days 2, 3, and 4 of the experiment.
Figure 1.
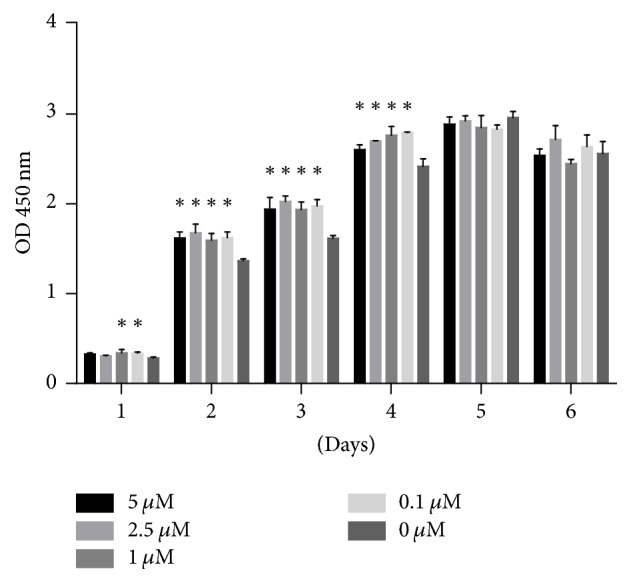
Effect of quercetin on cell proliferation in BMSCs. Cell proliferation was expressed using OD, which was measured for 6 consecutive days. Each value represents the mean ± SD from triplicate experiments. ∗ indicates a significant difference between the control and test groups at P < 0.05.
3.2. Quercetin Enhanced Osteoblastic Differentiation and Extracellular Matrix Production and Mineralization
Quantitative analysis of ALP activity was performed using an Alkaline Phosphatase Assay Kit. ALP activity was significantly enhanced in the 0 μM quercetin group compared with the control group, in which the cells were cultured in CM, on day 7 of the experiment. Treatment with additional quercetin or E2 further enhanced ALP activity. As shown in Figure 2(a), quercetin (0 μM–5 μM) induced osteoblastic differentiation in a dose-dependent manner. Notably, treatment with 5 μM quercetin had the greatest effect on ALP activity; however, we noted no significant difference in ALP activity between the 2.5 μM and 5 μM quercetin groups. Thus, 2.5 μM quercetin was used for subsequent experiments. Additionally, E2 (10 nM) was successfully utilized as a positive control in this experiment, as it also increased cell ALP activity.
Figure 2.

Effects of quercetin on ALP activity (a) and matrix mineralization (b)–(e). (a) For the ALP activity assay, cells were incubated in CM or OIM with or without quercetin or E2 for 7 days. Each value represents the mean ± SD from triplicate experiments. ∗ indicates a significant difference between the two groups at P < 0.05. (b)–(e) For the calcium nodule assay, the cells were incubated in CM (b), OIM with DMSO (c), OIM with quercetin (d), or OIM with E2 (e) for 21 days.
The effects of quercetin on extracellular matrix production and mineralization were further investigated by ARS staining for calcium deposits. As shown in Figures 2(b)–2(e), red (positive)-stained bone nodules were noted in the 0 μM quercetin group; however, no bone nodules were noted in the control group. Treatment with 10 nM E2 or 2.5 μM quercetin resulted in the formation of a large number of mineralized bone nodules. In addition to increasing the number of bone nodules, the above treatments also increased the area of each nodule. Some of the nodules joined together, forming large irregularly shaped nodules that stained a deep red.
3.3. Quercetin Upregulates Osteoblast-Specific Marker Gene Expression
As shown in Figure 3, treatment with 0.1–5 μM quercetin upregulated RUNX2, OSX, and OPN expression by 1.36–2.58-, 1.82–2.94-, and 1.50–2.17-fold, respectively, compared with treatment with the DMSO (0 μM quercetin group). These results were consistent with those of the ALP activity assay, suggesting that quercetin stimulated BMSC differentiation and function by upregulating RUNX2, OSX, and OPN expression. Treatment with quercetin at the indicated concentration (2.5 μM) elicited the maximal increases in the expression of the above proteins. Thus, this concentration was used for subsequent experiments.
Figure 3.

RUNX2, OSX, and OPN mRNA expression in BMSCs treated with quercetin. (a)–(c) The cells were incubated in osteogenic-inducing medium (OIM) with different concentrations of quercetin for 7 days. mRNA expression was then analyzed by real-time PCR. Target gene mRNA expression levels were normalized to GAPDH mRNA levels. Each value represents the mean ± SD from triplicate experiments. ∗ indicates a significant difference between the two groups at P < 0.05.
3.4. Quercetin Regulates Osteoblast-Specific Gene Expression via the ER
As shown in Figure 4, treatment with 2.5 μM quercetin and 10 nM E2 upregulated RUNX2, OSX, and OPN expression compared with treatment with DMSO (0 μM quercetin group). The addition of 1 μM ICI182780, a high-affinity ER antagonist, inhibited RUNX2, OSX, and OPN expression. RUNX2, OSX, and OPN expression levels were slightly different between the group treated with E2 and ICI182780 and the group treated with ICI182780 alone; however, there were no significant differences in RUNX2, OSX, and OPN expression between the two groups. Similarly, there were no significant differences in RUNX2, OSX, and OPN expression between the group treated with quercetin and ICI182780 and that treated with ICI182780 alone. The upregulatory effects of both quercetin and E2 were blocked by ICI182780.
Figure 4.

Effects of ICI182780 on RUNX2, OSX, and OPN mRNA expression in quercetin-treated BMSCs. The cells were cotreated with quercetin or E2 and 1 μM ICI182780 for 7 days. Each value represents the mean ± SD from triplicate experiments. ∗Significant differences between the values at P < 0.05.
3.5. Quercetin Regulates Osteoblast Marker Protein Expression
As shown in Figure 5, BMP2, Smad1, p-Smad1, Smad4, RUNX2, OST, and OPN expression levels in the 2.5 μM quercetin group were upregulated (2.19-, 2.03-, 2.76-, 2.29-, 2.22-, 2.22-, and 1.66-fold, respectively) compared with those in the control group. The relative expression levels of these proteins in the group treated with 2.5 μM quercetin and 1 μM ICI182780 were 1.040-, 1.039-, 1.312-, 1.326-, 1.123-, 0.965-, and 0.960-fold higher than those in the control group, respectively, indicating the fact that the ER is involved in mediating the effects of quercetin.
Figure 5.

Effects of ICI182780 on BMP2, Smad1, p-Smad1, Smad4, RUNX2, OSX, and OPN expression in quercetin-treated BMSCs. The cells were cotreated with quercetin and 1 μM ICI 182780 for 7 days, after which the expression levels of the above proteins were analyzed by western blotting. The data are presented as the ratio of protein expression in the treated group to protein expression in the control group. Each value represents the mean ± SD from triplicate experiments. ∗Significant differences between the values at P < 0.05.
4. Discussion
BMSCs are regarded as seed cells for bone tissue engineering because of their great multidirectional differentiation ability and reproductive activity. BMSCs reportedly give rise to several cells, including osteoblasts [3–5], and play a crucial role in the achievement and maintenance of an appropriate bone mass. Thus, it is important to promote BMSC proliferation and osteogenic differentiation.
In the current study, we first evaluated the effect of quercetin on BMSC proliferation. Our results showed that quercetin significantly increased BMSC proliferation in the quercetin-treated groups compared with the control group. ALP [30] is an early osteogenic marker and plays an important role in bone formation. A peak in ALP activity is indicative of differentiation [31, 32]. We quantified ALP activity to evaluate the effect of quercetin on BMSC osteogenic differentiation. The results indicated that quercetin significantly increased ALP activity on day 7 of the experiment and that 2.5 μM and 5 μM were the most effective quercetin concentrations utilized herein. We validated the above findings by ARS staining [33, 34]. The results showed that quercetin enhanced extracellular matrix formation.
RUNX2 is a core transcription factor that plays a pivotal role in regulating osteogenic differentiation. OSX, a zinc finger-containing transcription factor which is required for osteoblastic differentiation, acts as a downstream factor of RUNX2 [35–37]. OPN, an acidic glycoprotein that binds with hydroxyapatite in bone extracellular matrix [38], is a direct downstream target of RUNX2 [39]. In this study, we found that quercetin significantly increased RUNX2, OSX, and OPN mRNA expression levels. These results suggested that quercetin enhances BMSC osteogenic differentiation by regulating the expression of the transcription factors RUNX2 and OSX.
We then sought to determine how quercetin regulates the expression of the two transcription factors.
Estrogen is the key regulator of bone metabolism in vivo. Menopause and the accompanying loss of ovarian estrogen are associated with declines in bone mineral density [9]. Recent studies suggest that estrogen participates in the preservation of bone mass through non-nucleus-initiated mechanism via ER alpha [40]. A series of scientific studies showed that estrogen regulates bone metabolism by regulating several signaling molecules or interacting with other signaling pathways, such as the BMP/Smad signaling pathways [41–44]. In the current study, we confirmed that E2 enhances ALP activity and extracellular matrix formation and upregulates RUNX2 and OSX expression. In addition, quercetin, which is a member of a class of flavonoids that chemically resemble estrogen [28], has been shown to have biologic characteristics similar to those of estrogen in many studies regarding breast cancer [45–47]. Thus, we hypothesized that quercetin upregulates RUNX2, OSX, and OPN expression and promotes osteogenic differentiation by regulation of the estrogen signaling pathway. To confirm this hypothesis, we performed an experiment in which we investigated whether the stimulatory effects of quercetin on osteogenic differentiation in BMSCs were abolished by an ER inhibitor (ICI182780). The results showed that ICI182780 significantly reduced RUNX2, OSX, and OPN expression, indicating that ICI182780 blocked the upregulatory effects of quercetin and E2 on RUNX2, OSX, and OPN gene expression. These data clearly demonstrated that the estrogen signaling pathway plays a crucial role during quercetin-induced osteogenesis in BMSCs.
BMPs are members of the TGF-ß superfamily that were originally identified in bone-inductive extracts capable of inducing bone formation at ectopic sites [48, 49]. It is generally accepted that BMPs transmit signals to Smad proteins by binding to BMP receptors to ultimately regulate the transcription of RUNX2, OSX, and other genes [48]. As the most famous member of the BMP family, BMP2 has been shown to stimulate osteoblast differentiation and osteogenic nodule formation in vitro and bone formation in vivo [50–52]. The effect of BMP2 on osteogenic differentiation is enhanced by estrogen [10, 18], phytoestrogens [27], and flavonoids, such as genistein and icariin [26]. Other studies have shown that estrogenic compounds enhance bone formation by increasing BMP2 gene transcription [10, 18].
Quercetin is similar to genistein and icariin, as it is also a plant flavonoid and has estrogenic effects. Thus, we hypothesized that quercetin enhances BMP signaling pathway activation via the ER. First, to determine whether the BMP signaling pathway is also involved in quercetin-mediated BMSC osteogenic differentiation, we incubated cells with quercetin and OIM, after which we assessed protein marker expression using western blotting. Second, to confirm our hypothesis, we investigated whether the stimulatory effects of quercetin on osteogenic differentiation in BMSCs were abolished by the inhibitor ICI182780. Consistent with the results of previous studies, our results showed that quercetin significantly upregulated BMP2, Smad1, p-Smad1, Smad4, RUNX2, OSX, and OPN expression and that this upregulatory effect was blocked by ICI182780. Taken together, these findings showed that the E2 and BMP/Smad signaling pathways played important roles in the effects of quercetin on BMSC differentiation into osteoblasts.
Quercetin, a flavonoid, chemically resembles estrogen [28, 45–47]. Previous studies indicated that strong cross-talk occurs between the BMP and estrogen signaling pathways [18, 53]. In the present study, we found that quercetin stimulated BMP signaling by interacting with the ER, which led to increases in the expression of the downstream transcription factors RUNX2 and OSX and ultimately stimulated BMSC differentiation and mineralization.
Conflicts of Interest
The authors declare that there are no conflicts of interest regarding the publication of this paper.
References
- 1.Le Blanc K., Pittenger M. F. Mesenchymal stem cells: progress toward promise. Cytotherapy. 2005;7(1):36–45. doi: 10.1080/14653240510018118. [DOI] [PubMed] [Google Scholar]
- 2.Pittenger M. F., Mackay A. M., Beck S. C., et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284(5411):143–147. doi: 10.1126/science.284.5411.143. [DOI] [PubMed] [Google Scholar]
- 3.Teng Z. W., Zhu Y., Na Q., et al. Regulatory effect of miRNA on multi-directional differentiation ability of mesenchymal stem cell in treatment of osteoporosis. Journal of Biological Regulators and Homeostatic Agents. 2016;30(2):345–352. [PubMed] [Google Scholar]
- 4.Qu B., Chu Y., Zhu F., et al. Granulocyte colony-stimulating factor enhances the therapeutic efficacy of bone marrow mesenchymal stem cell transplantation in rats with experimental acute pancreatitis. Oncotarget . 2017;8(13):21305–21314. doi: 10.18632/oncotarget.15515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Liu X., Bao C., Hu J., Yin G., Luo E. Effects of clodronate combined with hydroxyapatite on multi-directional differentiation of mesenchymal stromal cells. Archives of Medical Science. 2010;6(5):670–677. doi: 10.5114/aoms.2010.17079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kim Y. J., Bae Y. C., Suh K. T., Jung J. S. Quercetin, a flavonoid, inhibits proliferation and increases osteogenic differentiation in human adipose stromal cells. Biochemical Pharmacology. 2006;72(10):1268–1278. doi: 10.1016/j.bcp.2006.08.021. [DOI] [PubMed] [Google Scholar]
- 7.Wang C., Meng H., Wang X., Zhao C., Peng J., Wang Y. Differentiation of bone marrow mesenchymal stem cells in osteoblasts and adipocytes and its role in treatment of osteoporosis. Medical Science Monitor. 2016;22:226–233. doi: 10.12659/MSM.897044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Klein-Nulend J., van Oers R. F. M., Bakker A. D., Bacabac R. G. Bone cell mechanosensitivity, estrogen deficiency, and osteoporosis. Journal of Biomechanics. 2015;48(5):855–865. doi: 10.1016/j.jbiomech.2014.12.007. [DOI] [PubMed] [Google Scholar]
- 9.Cauley J. A. Estrogen and bone health in men and women. Steroids. 2015;99:11–15. doi: 10.1016/j.steroids.2014.12.010. [DOI] [PubMed] [Google Scholar]
- 10.Matsumoto Y., Otsuka F., Takano M., et al. Estrogen and glucocorticoid regulate osteoblast differentiation through the interaction of bone morphogenetic protein-2 and tumor necrosis factor-α in C2C12 cells. Molecular and Cellular Endocrinology. 2010;325(1-2):118–127. doi: 10.1016/j.mce.2010.05.004. [DOI] [PubMed] [Google Scholar]
- 11.Martin A., Xiong J., Koromila T., et al. Estrogens antagonize RUNX2-mediated osteoblast-driven osteoclastogenesis through regulating RANKL membrane association. Bone. 2015;75:96–104. doi: 10.1016/j.bone.2015.02.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Khosla S., Oursler M. J., Monroe D. G. Estrogen and the skeleton. Trends in Endocrinology & Metabolism. 2012;23(11):576–581. doi: 10.1016/j.tem.2012.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Yuan F.-L., Xu R.-S., Jiang D.-L., et al. Leonurine hydrochloride inhibits osteoclastogenesis and prevents osteoporosis associated with estrogen deficiency by inhibiting the NF-κB and PI3K/Akt signaling pathways. Bone. 2015;75:128–137. doi: 10.1016/j.bone.2015.02.017. [DOI] [PubMed] [Google Scholar]
- 14.Gray T. K., Flynn T. C., Gray K. M., Nabell L. M. 17 beta-estradiol acts directly on the clonal osteoblastic cell line UMR106. Proceedings of the National Acadamy of Sciences of the United States of America. 1987;84(17):6267–6271. doi: 10.1073/pnas.84.17.6267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Komm B. S., Terpening C. M., Benz D. J., et al. Estrogen binding, receptor mRNA, and biologic response in osteoblast-like osteosarcoma cells. Science. 1988;241(4861):81–84. doi: 10.1126/science.3164526. [DOI] [PubMed] [Google Scholar]
- 16.Lin G. L., Hankenson K. D. Integration of BMP, Wnt, and notch signaling pathways in osteoblast differentiation. Journal of Cellular Biochemistry. 2011;112(12):3491–3501. doi: 10.1002/jcb.23287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zhang N.-D., Han T., Huang B.-K. Traditional Chinese Medicine formulas for the treatment of osteoporosis: implication for antiosteoporotic drug discovery. Journal of Ethnopharmacology. 2016;189:61–80. doi: 10.1016/j.jep.2016.05.025. [DOI] [PubMed] [Google Scholar]
- 18.Zhou S., Turgeman G., Harris S. E., et al. Estrogens activate bone morphogenetic protein-2 gene transcription in mouse mesenchymal stem cells. Molecular Endocrinology. 2003;17(1):56–66. doi: 10.1210/me.2002-0210. [DOI] [PubMed] [Google Scholar]
- 19.Zhou C., Lin Y. Osteogenic differentiation of adipose-derived stem cells promoted by quercetin. Cell Proliferation. 2014;47(2):124–132. doi: 10.1111/cpr.12097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Skibola C. F., Smith M. T. Potential health impacts of excessive flavonoid intake. Free Radical Biology & Medicine. 2000;29(3-4):375–383. doi: 10.1016/S0891-5849(00)00304-X. [DOI] [PubMed] [Google Scholar]
- 21.Hollman P. C. H., Katan M. B. Dietary flavonoids: Intake, health effects and bioavailability. Food and Chemical Toxicology. 1999;37(9-10):937–942. doi: 10.1016/s0278-6915(99)00079-4. [DOI] [PubMed] [Google Scholar]
- 22.Kelly G. S. Quercetin. Monograph. Alternative medicine review. A Journal of Clinical Therapeutic. 2011;16(2):172–194. [PubMed] [Google Scholar]
- 23.Horcajada-Molteni M.-N., Crespy V., Coxam V., Davicco M.-J., Remesy C., Barlet J.-P. Rutin inhibits ovariectomy-induced osteopenia in rats. Journal of Bone and Mineral Research. 2000;15(11):2251–2258. doi: 10.1359/jbmr.2000.15.11.2251. [DOI] [PubMed] [Google Scholar]
- 24.Zhou Y., Wu Y., Jiang X., et al. The effect of quercetin on the osteogenesic differentiation and angiogenic factor expression of bone marrow-derived mesenchymal stem cells. PLoS ONE. 2015;10(6) doi: 10.1371/journal.pone.0129605.e0129605 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Casado-Díaz A., Anter J., Dorado G., Quesada-Gómez J. M. Effects of quercetin, a natural phenolic compound, in the differentiation of human mesenchymal stem cells (MSC) into adipocytes and osteoblasts. The Journal of Nutritional Biochemistry. 2016;32:151–162. doi: 10.1016/j.jnutbio.2016.03.005. [DOI] [PubMed] [Google Scholar]
- 26.Zhang D., Fong C., Jia Z., Cui L., Yao X., Yang M. Icariin stimulates differentiation and suppresses adipocytic transdifferentiation of primary osteoblasts through estrogen receptor-mediated pathway. Calcified Tissue International. 2016;99(2):187–198. doi: 10.1007/s00223-016-0138-2. [DOI] [PubMed] [Google Scholar]
- 27.Dai J., Li Y., Zhou H., Chen J., Chen M., Xiao Z. Genistein promotion of osteogenic differentiation through BMP2/SMAD5/RUNX2 signaling. International Journal of Biological Sciences. 2013;9(10):1089–1098. doi: 10.7150/ijbs.7367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Guo A. J., Choi R. C., Zheng K. Y., et al. Kaempferol as a flavonoid induces osteoblastic differentiation via estrogen receptor signaling. Chinese Medicine. 2012;7, article no. 10 doi: 10.1186/1749-8546-7-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Swioklo S., Watson K. A., Williamson E. M., Farrimond J. A., Putnam S. E., Bicknell K. A. Defining Key Structural Determinants for the Pro-osteogenic Activity of Flavonoids. Journal of Natural Products. 2015;78(11):2598–2608. doi: 10.1021/acs.jnatprod.5b00075. [DOI] [PubMed] [Google Scholar]
- 30.Najafabadi B. M., Abnosi M. H. Boron induces early matrix mineralization via calcium deposition and elevation of alkaline phosphatase activity in differentiated rat bone marrow mesenchymal stem cells. Cell Journal. 2016;18(1):62–73. doi: 10.22074/cellj.2016.3988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Cao F.-Y., Fan J.-X., Long Y., Zeng X., Zhang X.-Z. A smart fluorescence nanoprobe for the detection of cellular alkaline phosphatase activity and early osteogenic differentiation. Nanomedicine: Nanotechnology, Biology and Medicine. 2016;12(5):1313–1322. doi: 10.1016/j.nano.2016.01.010. [DOI] [PubMed] [Google Scholar]
- 32.Ji Z., Cheng Y., Yuan P., Dang X., Guo X., Wang W. Panax notoginseng stimulates alkaline phosphatase activity, collagen synthesis, and mineralization in osteoblastic MC3T3-E1 cells. In Vitro Cellular & Developmental Biology - Animal. 2015;51(9):950–957. doi: 10.1007/s11626-015-9915-x. [DOI] [PubMed] [Google Scholar]
- 33.Moriguchi T., Yano K., Nakagawa S., Kaji F. Elucidation of adsorption mechanism of bone-staining agent alizarin red S on hydroxyapatite by FT-IR microspectroscopy. Journal of Colloid and Interface Science. 2003;260(1):19–25. doi: 10.1016/S0021-9797(02)00157-1. [DOI] [PubMed] [Google Scholar]
- 34.Orimo H., Shimada T. The role of tissue-nonspecific alkaline phosphatase in the phosphate-induced activation of alkaline phosphatase and mineralization in SaOS-2 human osteoblast-like cells. Molecular and Cellular Biochemistry. 2008;315(1-2):51–60. doi: 10.1007/s11010-008-9788-3. [DOI] [PubMed] [Google Scholar]
- 35.Komori T., Yagi H., Nomura S., et al. Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell. 1997;89(5):755–764. doi: 10.1016/S0092-8674(00)80258-5. [DOI] [PubMed] [Google Scholar]
- 36.Fakhry M., Hamade E., Badran B., Buchet R., Magne D. Molecular mechanisms of mesenchymal stem cell differentiation towards osteoblasts. World Journal of Stem Cells. 2013;5(4):136–148. doi: 10.4252/wjsc.v5.i4.136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Nishio Y., Dong Y., Paris M., O'Keefe R. J., Schwarz E. M., Drissi H. Runx2-mediated regulation of the zinc finger Osterix/Sp7 gene. Gene. 2006;372(1-2):62–70. doi: 10.1016/j.gene.2005.12.022. [DOI] [PubMed] [Google Scholar]
- 38.Franzen A., Heinegard D. Isolation and characterization of two sialoproteins present only in bone calcified matrix. Biochemical Journal. 1985;232(3):715–724. doi: 10.1042/bj2320715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Liu W., Toyosawa S., Furuichi T., et al. Overexpression of Cbfa1 in osteoblasts inhibits osteoblast maturation and causes osteopenia with multiple fractures. The Journal of Cell Biology. 2001;155(1):157–166. doi: 10.1083/jcb.200105052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Manolagas S. C., O'Brien C. A., Almeida M. The role of estrogen and androgen receptors in bone health and disease. Nature Reviews Endocrinology. 2013;9(12):699–712. doi: 10.1038/nrendo.2013.179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Abdallah B. M., Ditzel N., Mahmood A., et al. DLK1 is a novel regulator of bone mass that mediates estrogen deficiency-induced bone loss in mice. Journal of Bone and Mineral Research. 2011;26(7):1457–1471. doi: 10.1002/jbmr.346. [DOI] [PubMed] [Google Scholar]
- 42.Kamiya Y., Chen J., Xu M., et al. Increased mandibular condylar growth in mice with estrogen receptor beta deficiency. Journal of Bone and Mineral Research. 2013;28(5):1127–1134. doi: 10.1002/jbmr.1835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Song L., Zhao J., Zhang X., Li H., Zhou Y. Icariin induces osteoblast proliferation, differentiation and mineralization through estrogen receptor-mediated ERK and JNK signal activation. European Journal of Pharmacology. 2013;714(1-3):15–22. doi: 10.1016/j.ejphar.2013.05.039. [DOI] [PubMed] [Google Scholar]
- 44.Pierroz D. D., Rufo A., Biancht E. N., et al. β-Arrestin2 regulates RANKL and Ephrins gene expression in response to bone remodeling in mice. Journal of Bone and Mineral Research. 2009;24(5):775–784. doi: 10.1359/jbmr.081237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Wilkinson A. S., Taing M.-W., Pierson J. T., et al. Estrogen modulation properties of mangiferin and quercetin and the mangiferin metabolite norathyriol. Food & Function. 2015;6(6):1847–1854. doi: 10.1039/c5fo00133a. [DOI] [PubMed] [Google Scholar]
- 46.Wang H., Tao L., Qi K., et al. Quercetin reverses tamoxifen resistance in breast cancer cells. Journal of B.U.ON. 2015;20(3):707–713. [PubMed] [Google Scholar]
- 47.Ruotolo R., Calani L., Brighenti F., Crozier A., Ottonello S., Del Rio D. Glucuronidation does not suppress the estrogenic activity of quercetin in yeast and human breast cancer cell model systems. Archives of Biochemistry and Biophysics. 2014;559:62–67. doi: 10.1016/j.abb.2014.03.003. [DOI] [PubMed] [Google Scholar]
- 48.Peng Y., Kang Q., Cheng H., et al. Transcriptional Characterization of Bone Morphogenetic Proteins (BMPs)-Mediated Osteogenic Signaling. Journal of Cellular Biochemistry. 2003;90(6):1149–1165. doi: 10.1002/jcb.10744. [DOI] [PubMed] [Google Scholar]
- 49.Yu Y., Yang J. L., Chapman-Sheath P. J., Walsh W. R. TGF-beta, BMPS, and their signal transducing mediators, Smads, in rat fracture healing. Journal of Biomedical Materials Research Part B: Applied Biomaterials. 2002;60(3):392–397. doi: 10.1002/jbm.1289. [DOI] [PubMed] [Google Scholar]
- 50.Yamaguchi A., Ishizuya T., Kintou N., et al. Effects of BMP-2, BMP-4, and BMP-6 on osteoblastic differentiation of bone marrow-derived stromal cell lines, ST2 and MC3T3-G2/PA6. Biochemical and Biophysical Research Communications. 1996;220(2):366–371. doi: 10.1006/bbrc.1996.0411. [DOI] [PubMed] [Google Scholar]
- 51.Xiao G., Gopalakrishnan R., Jiang D., Reith E., Benson M. D., Franceschi R. T. Bone morphogenetic proteins, extracellular matrix, and mitogen-activated protein kinase signaling pathways are required for osteoblast-specific gene expression and differentiation in MC3T3-E1 cells. Journal of Bone and Mineral Research. 2002;17(1):101–110. doi: 10.1359/jbmr.2002.17.1.101. [DOI] [PubMed] [Google Scholar]
- 52.Harris S. E., Bonewald L. F., Harris M. A., et al. Effects of transforming growth factor β on bone nodule formation and expression of bone morphogenetic protein 2, osteocalcin, osteopontin, alkaline phosphatase, and type I collagen mRNA in long-term cultures of fetal rat calvarial osteoblasts. Journal of Bone and Mineral Research. 1994;9(6):855–863. doi: 10.1002/jbmr.5650090611. [DOI] [PubMed] [Google Scholar]
- 53.Foroughinia G., Fazileh A., Eghbalsaied S. Expression of genes involved in BMP and estrogen signaling and AMPK production can be important factors affecting total number of antral follicles in ewes. Theriogenology. 2017;91:36–43. doi: 10.1016/j.theriogenology.2016.12.023. [DOI] [PubMed] [Google Scholar]