

Abstract
The skin is a bilayered organ that serves as a key barrier between an organism and its environment. In addition to protecting against microbial invasion, physical trauma and environmental damage, skin participates in maintaining homeostasis. Skin is also capable of spontaneous self‐repair following injury. These functions are mediated by numerous pleiotrophic growth factors, including members of the vascular endothelial growth factor (VEGF), fibroblast growth factor (FGF), and transforming growth factor β (TGFβ) families. Although growth factor expression has been well documented in mammals, particularly during wound healing, for groups such as reptiles less is known. Here, we investigate the spatio‐temporal pattern of expression of multiple growth factors in normal skin and following a full‐thickness cutaneous injury in the representative lizard Eublepharis macularius, the leopard gecko. Unlike mammals, leopard geckos can heal cutaneous wounds without scarring. We demonstrate that before, during and after injury, keratinocytes of the epidermis express a diverse panel of growth factor ligands and receptors, including: VEGF, VEGFR1, VEGFR2, and phosphorylated VEGFR2; FGF‐2 and FGFR1; and phosphorylated SMAD2, TGFβ1, and activin βA. Unexpectedly, only the tyrosine kinase receptors VEGFR1 and FGFR1 were dynamically expressed, and only during the earliest phases of re‐epithelization; otherwise all the proteins of interest were constitutively present. We propose that the ubiquitous pattern of growth factor expression by keratinocytes is associated with various roles during tissue homeostasis, including protection against ultraviolet photodamage and coordinated body‐wide skin shedding.
Keywords: cytokine, keratinocytes, lizard, scar‐free wound healing, skin
The skin is the primary interface between an organism and its environment and serves a variety of barrier functions related to protection against mechanical abrasion, microorganisms, water loss, and ultraviolet radiation (Martin, 1997; Singer & Clark, 1999; Seifert & Maden, 2014). In addition, skin also participates in thermoregulation, synthesis of vitamin D, neurosensory perception, camouflage and crypsis, and various immune functions (Slominski et al. 1993; Singer & Clark, 1999). Underpinning the physiological and structural resiliency of the skin are impressive capacities for both homeostatic self‐renewal and injury‐mediated self‐repair. Both processes involve a variety of cytokines and growth factors that regulate the proliferation and differentiation programs.
Across vertebrates, the basic structure of the skin consists of two compartments: a superficial epidermis, dominated by keratinocytes; and a deeper dermis, rich in connective tissues (Fig. 1). The epidermis is a stratified squamous epithelium that in squamates (lizards, snakes and amphisbaenians) consists of five main layers: the Oberhaütchen, β‐keratin layer, α‐keratin layer, intermediate zone, and stratum germinativum (Jensen‐Jarolim, 2013; Allam et al. 2016; Fig. 1B). The Oberhaütchen, β‐keratin layer, and α‐keratin layer are often combined together as the stratum corneum (Jensen‐Jarolim, 2013; Pastar et al. 2014). Deep to the epidermis is the dermis (Fig. 1C,D), a dense connective tissue network that includes adipocytes, blood vessels, nerves, lymphatics, resident inflammatory cells, and two types of pigment cells: xanthophores (yellow pigments) and melanophores (black pigments) (Szydłowski et al. 2015).
Figure 1.
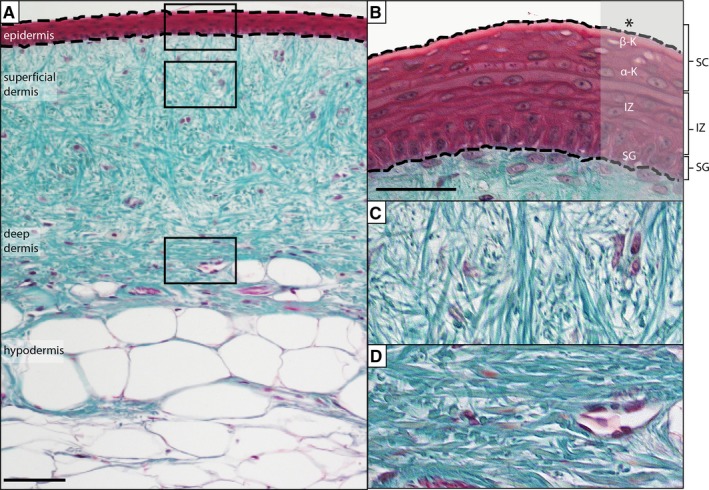
Anatomy of gecko skin. (A) Skin is organized into two compartments: epidermis and dermis. (B) The epidermis is a stratified, squamous epithelium. The most superficial layers are united as the stratum corneum (SC), and include the Oberhaütchen (*), β‐keratin, and α‐keratin layers. The Oberhaütchen and portions of the β‐keratin layers are frequently lost during histological preparation. Deep to the SC is the intermediate zone (IZ) and then the stratum germinativum (SG). The subadjacent dermis can be divided into (C) a superficial dermis (loose connective tissue dominated by thin bundles of collagen) and (D) a deep dermis (a more compact and densely arranged connective tissue with larger bundles of collagen). Masson's Trichrome. Scale bar = 10 μm
The epidermis undergoes constant physiological renewal, with cells of the stratum germinativum (= stratum basale) serving as the progenitor source (Pastar et al. 2014). Following proliferation, neo‐keratinocytes begin migrating through the intermediate zone towards the stratum corneum. During their migration, presumptive keratinocytes synthesize keratin and break down their organelles (Pastar et al. 2014). Once they reach the stratum corneum, keratinocytes are terminally differentiated and characteristically squamous‐shaped and anucleate. Over time, they are desquamated and replaced by the next generation.
In addition to homeostatic renewal, the skin is also capable of spontaneous self‐repair. In adult mammals, the most common mode of repair involves the formation of scar tissue. Although scars restore tissue homeostasis and prevent pathogen entry, they are non‐specific, fibrous replacements of the original organ. In contrast, some species of salamanders (Lévesque et al. 2010; Seifert et al. 2012a; Godwin & Rosenthal, 2014) and fetal mammals (up until mid‐gestation; Lorenz & Adzick, 1993; Ferguson & O'Kane, 2004; Larson et al. 2010; Satish & Kathju, 2010), as well as zebrafish (e.g. Azevedo et al. 2011; Stewart & Stankunas, 2012; Richardson et al. 2013) are capable of healing without scarring. This scar‐free mode of wound healing is a tissue‐specific process, substituting lost or damaged tissues with a near‐perfect restoration of tissue architecture, pigmentation, and the re‐establishment of integumentary organs (i.e. glands, hair, scales). Recently, scar‐free wound healing has also been demonstrated in African spiny mice (Acomys spp.; Seifert et al. 2012b; Santos et al. 2016) and various species of lizard (Wu et al. 2014; Peacock et al. 2015).
In mammals, both homeostatic renewal and wound‐healing programs are tightly regulated processes, orchestrated by multiple growth factors and cytokines including members of the vascular endothelial growth factor (VEGF), fibroblast growth factor (FGF), and transforming growth factor β (TGFβ) families. VEGF is a heparin‐binding protein widely recognized as a potent pro‐angiogenic, involved in endothelial cell proliferation, migration and (ultimately) blood vessel formation, and hence is crucial for restoring vasculature to regenerating tissues (Brown et al. 1992; Nissen et al. 1998; Gurtner et al. 2008). In mammals, VEGF also acts in an autocrine manner to promote keratinocyte proliferation and survival prior to and throughout wound healing (Wilgus et al. 2005; Man et al. 2006; Yang et al. 2010; Zhu et al. 2013). VEGF function is primarily mediated through two main receptor tyrosine kinases, VEGFR2 (Flk‐1) and VEGFR1 (flt‐1), both of which are expressed by endothelial cells and keratinocytes (Brown et al. 1992; Ferrara, 2004). In the skin, the major sources of VEGF include keratinocytes, as well as platelets and macrophages (Brown et al. 1992).
Like VEGF, FGFs are also heparin‐binding proteins, with potent pro‐angiogenic functions (Battegay, 1995; Tonnesen et al. 2000). In addition, FGFs are involved in various cell functions, including differentiation, migration, proliferation and cytoprotective/cell survival roles following stress (Abraham & Klagsbrun, 1988; Basilico & Moscatelli, 1992; Werner, 1998; Ornitz & Itoh, 2001; Werner & Grose, 2003; Yang et al. 2010). Although all FGF ligands and receptors have been detected in uninjured and regenerating skin, FGF‐2 (also known as basic FGF) appears to be particularly important during wound healing, inducing granulation tissue, re‐epithelialization and tissue remodelling (Werner & Grose, 2003; Barrientos et al. 2008). For example, FGF‐2 null mice demonstrate a significant delay in re‐epithelialization, decreased collagen deposition and increased scab thickness following injury (Ortega et al. 1998). Although less is known about the role of FGFs in reptiles, they have been reported during lizard tail regeneration. Previous work on two distantly related species of lizards (Lamprophilis guichenoti and Podarcis sicula) has revealed that FGF‐2 and FGF‐1 are largely absent from the epidermis prior to injury, but robustly expressed by keratinocytes during tail regeneration (Alibardi & Lovicu, 2010; Alibardi, 2012).
The TGFβ superfamily includes the prototypic members, TGFβ1‐3, along with the activins, inhibins, bone morphogenic proteins, growth differentiation factors, nodals and myostatin (Derynck & Miyazono, 2008). TGFβ1‐3/activins signal through a common canonical [Small Body and Mothers Against Decepentaplegic Homolog (SMAD)]‐mediated] pathway (Moustakas & Heldin, 2009; Wu & Hill, 2009; Ogunjimi et al. 2012). Once activated, TGFβ and activin phosphorylate SMAD2 and SMAD3 (Ross & Hill, 2008) and are then translocated to the nucleus to regulate gene expression (Ross & Hill, 2008). TGFβ and activins have pleiotropic effects across the body, including both tissue homeostasis and wound healing (Werner & Grose, 2003; Gilbert et al. 2016). However, the diversity of effects are context and cell‐type dependent (Roberts, 1998). For example, although all three TGFβ isoforms are known mitogens, they also suppress proliferation in various cell types, including keratinocytes (Roberts, 1998). In mammals, TGFβ1 (and TGFβ2) is typically considered to be pro‐fibrotic (and thus associated with scar formation) during wound healing, whereas TGFβ3 has been shown to be anti‐fibrotic/pro‐regenerative (O'Kane & Ferguson, 1997; Järvinen & Ruoslahti, 2010). Interestingly, TGFβ3 is not expressed during tail regeneration in lizards (Delorme et al. 2012), whereas TGFβ1 is almost ubiquitously expressed by keratinocytes before and following tail loss, with one conspicuous exception – it is transiently absent during the early phase of new tail outgrowth (Gilbert et al. 2013).
Activins are dimer proteins made up of the subunits activin βA and activin βB. In combination, these subunits create the three activin ligands: activin A (βA, βA), activin B (βB, βB) and activin AB (βA, βB) (Werner & Grose, 2003; Werner & Alzheimer, 2006). Within the integument, activins are involved in skin morphogenesis, keratinocyte proliferation, and cutaneous wound healing (Sulyok et al. 2004; Bamberger et al. 2005). Although little is known about their function among reptiles, activin βA is upregulated following tail loss in geckos (Gilbert et al. 2013).
Here, we performed a spatio‐temporal characterization of VEGF, FGF‐2, TGFβ1 and activin βA expression in normal (uninjured) and healing skin of a representative lizard Eublepharis macularius (Blyth, 1854), the leopard gecko (hereafter ‘gecko’). We hypothesized that growth factor expression would be dynamically regulated and would contribute to the scar‐free wound healing seen in geckos. Similar to some salamanders and zebrafish, geckos are capable of spontaneous scar‐free wound healing (Delorme et al. 2012; Peacock et al. 2015). We determined that gecko keratinocytes, like those of mammals, express a diversity of growth factor ligands and receptors under homeostatic and injury‐mediated conditions. Combined, the observed expression patterns underscore roles for each of these cytokines beyond classic pro‐angiogenic functions in a species capable of healing without forming a scar.
Methods
Animal care
Captive bred E. macularius were acquired from a commercial supplier (Global Exotic Pets, Kitchener, ON, Canada). At the beginning of the experiment, all animals were sexually immature and less than 1 year old, with a body mass range of 8.3–29.7 g. Animal Usage Protocols were approved by the University of Guelph Animal Care Committee (Protocol Number 2493) and followed the procedures of the Canadian Council on Animal Care. Geckos were housed and maintained following the work of McLean & Vickaryous (2011). Briefly, the gecko colony was kept in an isolated, temperature‐controlled environmental chamber with a 12 : 12 h photoperiod and an average room temperature of 27.5 °C. Individual geckos were housed in 5‐gal polycarbonate containers, with a subsurface heating cable (Hagen Inc., Baie d'Urfe, Quebec, Canada) set to 32 °C placed under one end to create a temperature gradient. Geckos were fed three to five larval Tenebrio spp. (mealworm) dusted with powdered calcium and vitamin D3 (cholecalciferol) (Zoo Med Laboratories Inc., San Luis Obispo, CA, USA) daily and had free access to clean drinking water. A total of 20 geckos (n = 4 for each of four time points, plus four sentinel/environmental controls) were used to characterize the spatio‐temporal expression of endogenous growth factors throughout wound healing and regeneration. Geckos were randomly assigned into one of five groups: sentinel; original uninjured tissue; biopsy and tissue collection at 2, 8 and 45 days following injury. This work was completed in accordance with the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines.
Biopsies
Prior to biopsy, geckos were anaesthetized using a 30 mg kg−1 intramuscular injection of Alfaxan (diluted to 2 mg mL−1 in sterile injectable 0.9% sodium chloride, using a 0.5‐cm3 insulin syringe; Abbott Laboratories, Saint‐Laurent, Quebec, Canada). Injections were done bilaterally into the cervical epaxial musculature. Geckos were considered to have reached the surgical plane of anaesthesia once the righting reflex was lost (Schumacher & Yelen, 2006). A biopsy punch tool (Integra Miltex, Burlington, ON, Canada) was then used to create a 3‐mm full‐thickness (epidermis and dermis) wound into the dorsal skin of the tail. Each gecko received two parasagittal biopsy wounds: one proximal (along the proximal third of the tail) and one distal (along the middle third of the tail). Proximal and distal wounds were on alternate sides of the midline. Each biopsy wound was separated by 5 cm. Excised tissue was removed with the use of both #11 scalpel blade and forceps. Both biopsies on each tail healed in a similar manner and as such they were both treated the same and not as independent groups for the purposes of this study.
Tissue collection
Experimental gecko tissues were collected at four time points either prior to (original tissue/biopsy controls) or following biopsy (2, 8 and 45 days); tissues were not collected from sentinel geckos. Geckos were euthanized with an intra‐abdominal injection of 250–500 mg ethyl 3‐aminobenzoate methane sulphonic acid (tricaine methansulphonate, MS222), then placed in 10% NBF (neutral buffered formalin; Fisher Scientific, Waltham, MA, USA) for approximately 24 h (using either transcardial perfusion followed by immersion or the tail tissues were dissected into regions of interest and these were directly immersed). Following fixation, the tissue was then rinsed with distilled water and transferred to 70% ethanol. Regions of interest were dissected/trimmed as necessary, before being de‐calcified with Cal‐Ex® (Fisher Scientific, Waltham, MA, USA) for 30 min. Tissues were then put in 100% isopropanol, cleared in xylene and infiltrated with paraffin wax using an automated processor (Shandon Excelsior ES Tissue Processor, Thermo Fisher Scientific). Tissue samples were then embedded in paraffin blocks and sectioned at 5 μm using a rotary microtome (Shandon Finesse ME+ Thermo Fisher Scientific), before being mounted on charged slides (Surgipath® X‐tra®, Leica Microsystems, Concord, ON, Canada), and baked at 60 °C overnight.
Haematoxylin and eosin
Representative sections from each tissue sample were stained with haematoxylin and eosin to examine tissue structure. Briefly, slide‐mounted tissue sections were first rehydrated to water: three rinses of xylene (2 min each); three rinses of 100% isopropanol (2 min each); one rinse in 70% isopropanol (2 min); and one rinse in deionized water (2 min). Once rehydrated, sections were stained with modified Harris haematoxylin (Fisher Scientific) for 10 min and then dipped 6–10 times in a solution of 1% hydrochloric acid in 70% isopropanol, before being rinsed in deionized water. Next, slides were blued in ammonia water for about 15 s rinsed in deionized water, and then dipped in 70% isopropanol six times, followed by staining with eosin (1 min). Stained sections were dehydrated with four rinses of absolute isopropanol (2 min each) and then cleared with three rinses of xylene (2 min each). Finally, slides were cover‐slipped using Cytoseal (Fisher Scientific).
Masson's trichrome
To differentiate fibrous connective tissue, representative sections were stained with a modified Masson's trichrome (McLean & Vickaryous, 2011). After rehydration (see above), slide‐mounted sections were stained with Mayer's haematoxylin (10 min), blued in ammonia water for approximately 15 s and rinsed with deionized water. Sections were stained in 0.5% ponceau xylidine/0.5% acid fuschin in 1% acetic acid solution (2 min); rinsed in deionized water; stained in 1% phosphomolybdic acid (10 min); rinsed in deionized water; stained in 2% light green (90 s); and rinsed in deionized water. Slides were then dehydrated [one rinse in 95% isopropanol (2 min); three rinses in absolute isopropanol (2 min)] and cleared with three rinses in xylene (2 min each). Finally, slides were cover‐slipped using Cytoseal (Fisher Scientific).
Immunohistochemistry
Immunohistochemistry was performed to identify localized protein expression of pVEGFR2, FGF‐2, FGFR1, pSMAD2, TGFβ1 and activin βA (Table 1). Once rehydrated, slide‐mounted sections were quenched in 3% hydrogen peroxide (20 min), then rinsed three times in phosphate‐buffered saline (PBS; 2 min each). For three proteins (pVEGFR2, FGF‐2 and activin βA), heat‐induced antigen retrieval was employed to unmask the epitope of interest (citrate buffer at 90 °C for 12 min, after which the buffer was allowed to cool for 20 min). Sections were then rinsed three times in PBS (2 min each) and blocked using 3% normal goat serum (Vector Laboratories, Burlingame, CA, USA) diluted in sterile PBS for 1 h at room temperature. Sections were then incubated with the primary antibody diluted in sterile PBS overnight at 4 °C; omission (negative) controls were incubated without the primary antibody. The next day, sections were rinsed three times in PBS (2 min each), then incubated with the secondary antibody diluted in sterile PBS for 1 h at room temperature. Sections were then rinsed three times in PBS (2 min each), before being incubated with horseradish peroxidase conjugated streptavidin (Jackson ImmunoResearch Laboratories, Inc. West Grove, PA, USA, code: 016‐030‐084) diluted in sterile PBS for 1 h at room temperature. Sections were then rinsed three times in PBS (2 min each) and then 3,3′‐diaminobenzidine peroxidase substrate (DAB; Vector Laboratories) was applied for a time optimized to each antibody (Table 1). The chromogenic reaction was stopped by immersing the sections in deionized water, after which the sections were counterstained with Mayer's haematoxylin (1 min), rinsed in deionized water, blued in ammonia water, and rinsed again in deionized water. Slides were then dehydrated (see above) and cover‐slipped using Cytoseal (Fisher Scientific).
Table 1.
A summary of all optimized immunohistochemistry and immunofluorescence protocols for proteins of interest
| Antigen | Type | Retrieval | Block | Primary | Secondary | HRP | DAB |
|---|---|---|---|---|---|---|---|
| vWF | IF | None | 3% NGS 1 h RT | Rabbit anti‐von Willebrand factor 1 : 500 (Dako Canada, A0082) | Cy3 goat anti‐rabbit 1 : 400 (Jackson ImmunoResearch Laboratories, Inc. 111‐165‐144) | ||
| α‐SMA | IF | None | 3% NGS 1 h RT | Mouse anti‐α‐actin 1 : 400 (Santa Cruz BioTechnology, Inc. sc‐32252) | Goat anti‐mouse AlexaFluor‐488 1 : 400 (Life Technologies, A‐11001) | ||
| VEGF | IHC | 12 min citrate buffer | 3% NGS 1 h RT | Rabbit anti‐VEGF 1 : 100 (Santa Cruz BioTechnology, Inc. sc‐152) | Biotinylated goat anti‐rabbit 1 : 500 (Jackson ImmunoResearch Laboratories, Inc. 111‐066‐003) | 1 : 200 | 25 s |
| IF | 12 min citrate buffer | 3% NGS 1 h RT | Rabbit anti‐VEGF 1 : 50 (Santa Cruz BioTechnology, Inc. sc‐152) | Cy3 goat anti‐rabbit 1 : 50) (Jackson ImmunoResearch Laboratories, Inc. 111‐154‐144) | |||
| VEGFR2 (Flk‐1) | IHC | 12 min citrate buffer | 3% NGS 1 h RT | Mouse anti‐Flk‐1 1 : 600 (Santa Cruz BioTechnology, Inc. sc‐6251) | Biotinylated goat anti‐mouse 1 : 200 (Vector Laboratories BA‐9200) | 1 : 200 | 40 s |
| IF | 12 min citrate buffer | 3% NGS 1 h RT | Mouse anti‐Flk‐1 1 : 100 (Santa Cruz BioTechnology, Inc. sc‐6251) | Goat anti‐mouse AlexaFluor‐488 1 : 100 (Life Technologies, A‐11001) | |||
| VEGFR1 (flt‐1) | IHC | 12 min citrate buffer | 3% NGS 1 h RT | Rabbit anti‐Flt‐1 1 : 200 (Santa Cruz BioTechnology, Inc. sc‐316) | Biotinylated goat anti‐rabbit 1 : 200 (Jackson ImmunoResearch Laboratories, Inc. 111‐165‐144) | 1 : 200 | 50 s |
| IF | 12 min citrate buffer | 3% NGS 1 h RT | Rabbit anti‐Flt‐1 1 : 50 (Santa Cruz BioTechnology, Inc. sc‐316) | Cy3 goat anti‐rabbit 1 : 200) (Jackson ImmunoResearch Laboratories, Inc. 111‐154‐144) | |||
| pVEGFR2 Y1054/1059 | IHC | 12 min citrate buffer | 3% NGS 1 h RT | Rabbit anti‐VEGF receptor 2 Y1054/1059 1 : 200 (Abcam, ab5473) | Biotinylated goat anti‐rabbit 1 : 200 (Jackson ImmunoResearch Laboratories, Inc. 111‐066‐003) | 1 : 200 | 1 min |
| FGF‐2 | IHC | 12 min citrate buffer | 5% NGS 1 h RT | Rabbit anti‐FGF‐2 1 : 100 (Santa Cruz BioTechnology, Inc. sc‐79) | Biotinylated goat anti‐rabbit 1 : 200 (Jackson ImmunoResearch Laboratories, Inc. 111‐165‐144) | 1 : 200 | 40 s |
| FGFR1 | IHC | None | 3% NGS 1 h RT | Rabbit anti‐FGFR1 1 : 50 (Cell Signaling Technology, #3472) | Biotinylated goat anti‐rabbit 1 : 200 (Jackson ImmunoResearch Laboratories, Inc. 111‐066‐003) | 1 : 200 | 70 s |
| PCNA | IF | None | 3% NGS 1 h RT | Rabbit anti‐PCNA 1 : 100 (Santa Cruz BioTechnology, Inc. sc‐7907) | Cy3 goat anti‐rabbit 1 : 200) (Jackson ImmunoResearch Laboratories, Inc. 111‐154‐144) | ||
| pSMAD2 | IHC | None | 3% NGS 1 h RT | Rabbit anti‐pSMAD2 1 : 800 (Cell Signaling Technology, ser465/467) | Biotinylated goat anti‐rabbit 1 : 200 (Jackson ImmunoResearch Laboratories, Inc.111‐066‐003) | 1 : 200 | 40 s |
| TGFβ1 | IHC | None | 3% NGS 1 h RT | Rabbit anti‐TGFβ1 1 : 500 (Santa Cruz BioTechnology Inc., sc‐146) | Biotinylated goat anti‐rabbit 1 : 500 (Jackson ImmunoResearch Laboratories, Inc. 111‐066‐003) | 1 : 200 | 30 s |
| Activin βA | IHC | 12 min citrate buffer | Blocking buffer 1 h RT | Kind donation from Dr Paul Sawchenko; 1 : 500 Salk Institute for Biological Studies, La Jolla, CA PBL #207‐234 | Biotinylated goat anti‐rabbit 1 : 200 (Jackson ImmunoResearch Laboratories, Inc.111‐066‐003) | 1 : 50 | 45 s |
α‐SMA, α‐smooth muscle actin; DAB, 3,3′‐diaminobenzidine; FGF‐2, fibroblast growth factor 2; FGFR1, fibroblast growth factor receptor 1; HRP, horseradish peroxidise; IF, immunofluorescence; IHC, immunohistochemistry; NGS, normal goat serum; PBS, phosphate‐buffered saline; pSMAD2, phosphorylated SMAD2; pVEGFR2, phosphorylated vascular endothelial growth factor receptor 2 (tyrosine residues Y1054/1059); RT, room temperature; TGFβ1, transforming growth factor β1; VEGF, vascular endothelial growth factor; VEGFR1, vascular endothelial growth factor receptor 1 (flt‐1); VEGFR2, vascular endothelial growth factor receptor 2 (Flk‐1); vWF, von Willebrand factor.
For activin βA, a modified immunohistochemistry protocol was utilized to detect the epitope. There are three primary differences between the above‐mentioned immunohistochemistry protocol and the modified protocol: (i) Tris‐buffered saline with Tween20® (TBST; Sigma‐Aldrich, Oakville, ON, Canada) rinses are used in place of PBS rinses; (ii) a blocking buffer comprising 3% bovine serum albumin (BSA; Santa Cruz BioTechnology, Santa Cruz, CA, USA), 10% normal goat serum in TBST is used instead of the 3% NGS block; (iii) the secondary antibody is only incubated for 30 min at room temperature as opposed to 1 h.
Immunofluorescence
Immunofluorescence was performed to visualize PCNA, as well as to co‐localize three pairs of proteins: VEGF and VEGFR2; VEGFR1 and VEGFR2; and vWF and α‐SMA (Table 1). Once rehydrated, heat‐induced antigen retrieval was employed to unmask the epitope of interest for all antibodies used except for vWF and α‐SMA (citrate buffer at 90 °C for 12 min, after which the buffer was allowed to cool for 20 min). Sections were then rinsed three times in PBS (2 min each), and blocked using 3% normal goat serum (Vector Laboratories) diluted in sterile PBS for 1 h at room temperature. Sections were then incubated with the primary antibodies diluted in sterile PBS overnight at 4 °C; omission (negative) controls were incubated without either primary antibody. The next day, sections were rinsed three times in PBS (2 min each), then incubated with the secondary antibodies diluted in sterile PBS for 1 h at room temperature. Sections were then rinsed three times in PBS (2 min each) before being incubated with 4′‐6′‐diamidino‐2‐phenylindole (DAPI; Life Technologies, ThermoFisher Scientific, D1306) diluted 1 : 10 000 in PBS (2 min). Sections were then rinsed three times in PBS (2 min each) and cover‐slipped with Dako Fluorescent Mounting Medium (Dako Canada S3022, Burlington, ON, Canada).
Results
Prior to injury, the skin across the dorsal body surface demonstrated two main scale morphologies: large conical‐shaped tubercles and smaller low profile scales (Fig. S1A). Although pigmentation was often variable, it typically involved countershading with a dorsal pattern of brown‐black spots on a background of white, orange and yellow, and a near solid white ventral surface.
To initiate cutaneous wound healing, we created full‐thickness (epidermal and dermal) wounds to the dorsal surface of the tail (Supporting Information Fig. S1). The pattern of wound healing and regeneration observed was consistent with previous reports (Peacock et al. 2015). Blood loss from the biopsy wounds was minimal, and haemostasis was naturally achieved within several minutes, no exogenous methods to control haemorrhaging were required. All wounds healed through secondary intention (like humans, but unlike mice; Galiano et al. 2004; Christenson et al. 2005; Dunn et al. 2013). No changes in gecko behaviour or growth were observed following biopsy, nor were there any signs of infection or inflammation at the site of injury. The pattern and mode of wound healing was identical across biopsy wounds created on proximal and distal locations along the tail, and therefore these data are presented together.
As has been previously demonstrated (Peacock et al. 2015), full‐thickness biopsies to gecko skin, healed without scarring. Briefly, an exudate clot was present within 12 h post‐wounding and remained visible 2 days ppost‐wounding (DPW; Fig. S1B). By 8 DPW, this clot had self‐detached, revealing the presence of a shiny, smooth wound epithelium (WE; Fig. S1C). The WE remained unpigmented until ~ 14 DPW (Peacock et al. 2015). By 45 DPW scalation and pigmentation had essentially been restored (Fig. S1D). As observed during previous experiments investigating complete tail regeneration (e.g. McLean & Vickaryous, 2011; Delorme et al. 2012; see also Bellairs & Bryant, 1985), and cutaneous wound healing (Peacock et al. 2015), tubercle scales were not regenerated. Instead, the regenerated skin was clad exclusively in small (~ 2 mm) low‐profile scales, comparable to those of the original (uninjured) tissue. In addition, although the new skin was pigmented, the pattern observed did not replicate that of the original. Although the absence of tubercles cannot be explained at present, it is worth noting that the mechanism involved in scale regeneration (invagination of the WE) differs from that involved in scale development (viz. evagination) (Wu et al. 2014).
Histology of the reptilian epidermis
In section, the original (uninjured) epidermis ranged in preserved thickness from three to four cell layers, primarily representing the stratum germinativum, the intermediate zone, and the basal‐most layers of the stratum corneum (Supporting Information Fig. S2A); the more superficial layers of the stratum corneum were generally lost during tissue processing. As evidenced by immunostaining, cell proliferation (visualized with PCNA) within the epidermis was restricted to the stratum germinativum (Fig. S2B). The dermis was invested with structurally mature blood vessels co‐expressing the endothelial marker von Willebrand Factor (vWF) and the mural cell marker α‐smooth muscle actin (α‐SMA) (Fig. S2C).
Following excisional injury, a program of scar‐free wound healing was initiated. Restoration began with the formation of an exudate clot, followed by re‐epithelization (Fig. S2D). At 2 DPW the WE was present along the wound margins, spanning centripetally. Although PCNA+ cells were present within the tongue‐like epithelial outgrowth (Fig. S2E), there was no evidence of blood vessels within the sub‐adjacent wound bed (Fig. S2F).
By 8 DPW, re‐epithelialization was complete and the WE had achieved its maximal preserved thickness, 6–10 cell layers (Fig. S2G). This increased thickening of the WE is consistent with previous investigations of cutaneous wound healing in lizards (Peacock et al. 2015; see also Wu et al. 2014). Furthermore, it closely resembled the transient thickening of the WE observed following tail loss (e.g. Whimster, 1978; McLean & Vickaryous, 2011; Delorme et al. 2012). In addition to the stratum germinativum, there were large numbers of PCNA+ cells within the intermediate zone of the WE (Fig. S2H). Blood vessels were also found to be present within the wound bed and were characteristically mural cell‐supported (Fig. S2I).
By 45 DPW, the epidermis was restored to its original organization (preserved as three to four cell layers thick; Fig. S2J), and the pattern of small scales had re‐developed. Both the distribution of PCNA+ cells (restricted to the stratum germinativum; Fig. S2K) and the structural maturity of blood vessels within the dermis (Fig. S2L) matched that of the original skin.
Keratinocytes of the original epidermis and WE express VEGF
Although VEGF is best known as a potent pro‐angiogenic factor and mitogen of endothelial cells, in mammals it is also constitutively expressed by keratinocytes (Weninger et al. 1996) and is upregulated during periods of wound healing (Brown et al. 1992; see also Frank et al. 1995). Using immunostaining, we determined that prior to injury, virtually all gecko keratinocytes (Fig. 2A–C; see also Supporting Information Fig. S3) and many fibroblasts of the dermis (data not shown), robustly co‐localized with both VEGF and VEGFR2. This widespread pattern of VEGF/VEGFR2 expression was largely maintained throughout wound healing (Fig. 2D–F). Prior to the completion of re‐epithelialization, neo‐keratinocytes did not uniformly express VEGF/VEGFR2, but both the ligand and receptor were still detected in the wound margin (WM) as well as the leading edge (Fig. 2D). By 45 DPW, the original near‐uniform pattern of overlapping expression had returned (Fig. 2F).
Figure 2.
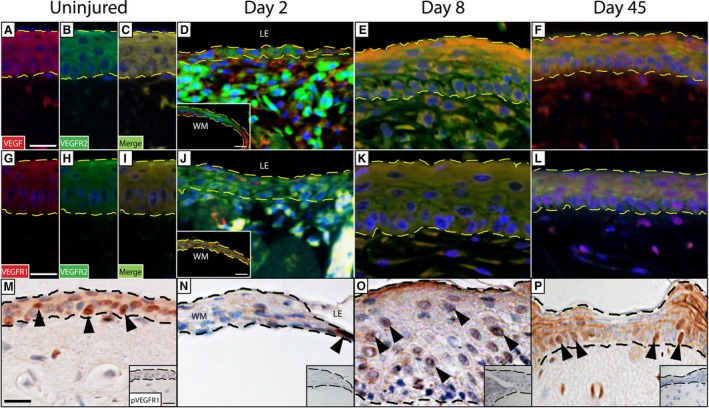
VEGF ligand and receptor expression by keratinocytes. Prior to injury, expression of VEGF (A) and VEGFR2 (B) by keratinocytes is widespread. As evidenced by the merged image (C), these markers are frequently co‐localized. At the start of wound healing (2 DPW), VEGF and VEGFR2 are detected in both the wound margin (WM; inset, bottom left‐hand corner) as well as the leading edge (LE), although this expression is not uniform (D). By 8 DPW, VEGF and VEGFR2 are strongly co‐localized and are expressed by nearly all keratinocytes (E). After wound healing is complete, the expression at 45 DPW resembles that of the original epidermis (F). Keratinocytes also express a second VEGF receptor, VEGFR1, in the original epidermis (G). VEGFR1 and VEGFR2 (H) are also frequently co‐localized (I). Prior to re‐epithelialization (2 DPW), VEGFR1 and VEGFR2 are co‐localized in the wound margin (inset, bottom left‐hand corner), but VEGFR1 is notably absent in the leading edge (J). However, the two receptors show co‐expression following re‐epithelialization at 8 (K) and 45 days (L) following injury. Phosphorylated VEGFR2 (pVEGFR2) was assessed to determine pathway activation. We noted constitutive expression of pVEGFR2 in the uninjured epidermis (indicated by the black arrowheads; M), as well as throughout wound healing (N‐P). Dashed line indicates the junction between the overlying epidermis and the deeper dermis. Omission controls = insets, bottom right hand corner. See Fig. S3 for negative controls. Scale bar: 10 μm.
Next, we sought to determine whether VEGFR1, another high‐affinity VEGF receptor, was expressed by gecko keratinocytes. Prior to injury, cells of the original epidermis demonstrated widespread VEGFR1 immunoreactivity, often revealed to co‐localize with VEGFR2 (Fig. 2G–I). Although VEGFR1 and VEGFR2 were strongly co‐localized in the WM at 2 DPW, VEGFR1 (but not VEGFR2) was conspicuously absent from keratinocytes at the leading edge (Fig. 2J). However, by 8 DPW (once re‐epithelialization of the wound was complete) co‐expression of both receptors throughout the WE had resumed (Fig. 2K). Once wound healing was complete (45 DPW), the expression pattern of VEGFR1 and VEGFR2 by keratinocytes closely resembled that of the original epidermis (Fig. 2L).
Most VEGF functions take place when VEGF binds to its main receptor, VEGFR2. This results in phosphorylation of tyrosine receptor residues and the activation of various downstream cellular signals (Clegg & Gabhann, 2015). Using an antibody raised against tyrosine residues Y1054 and Y1059, we investigated VEGFR2 phosphorylation before, during and after creating the cutaneous biopsies. Most keratinocytes in the uninjured gecko epidermis expressed phosphorylated VEGFR2 (pVEGFR2) (Fig. 2M). At 2 DPW, there were comparatively few pVEGFR2‐positive cells (Fig. 2N), but by 8 DPW there was a marked increase in the number of labelled cells (Fig. 2O). By 45 DPW, the pattern of pVEGFR2 expression was virtually identical to that of the original epidermis (Fig. 2P).
Keratinocytes of the original and regenerating epidermis express FGF‐2
To expand our characterization, we then investigated the expression pattern of FGF‐2 and TGFβ. Like VEGF, FGF‐2 is involved in regulating angiogenesis and cell proliferation (Kottakis et al. 2011). In mammals, FGF‐2 is also reported to play roles in self‐renewal of stem/progenitor populations and DNA repair of keratinocytes following radiation‐induced damage (Harfouche et al. 2010). Furthermore, exogenous treatment with FGF‐2 stimulates keratinocyte migration in vitro (Sogabe et al. 2006). Prior to injury, keratinocytes of the gecko epidermis demonstrated strong immunoreactivity for FGF‐2 (Fig. 3A). Prior to wound closure (2 DPW), FGF‐2 expression was only observed among individual neo‐keratinocytes (Fig. 3B), but once wound closure was complete (8 DPW), FGF‐2 expression resumed throughout the WE (Fig. 3C). By 45 DPW, the pattern of FGF‐2 (Fig. 3D) expression had returned to baseline and closely resembled that of the original epidermis. We then investigated FGFR1, a high‐affinity receptor for FGF‐2. In the uninjured epidermis, FGFR1 was ubiquitously expressed (Fig. 3E), but at 2 DPW, FGFR1 was conspicuously absent (Fig. 3F). At 8 DPW, FGFR1 expression had returned, although immunopositive cells were primarily restricted to the outermost layers of the WE (Fig. 3G). By 45 DPW, FGFR1 immunoreactivity had returned to its pre‐injury appearance (Fig. 3H).
Figure 3.
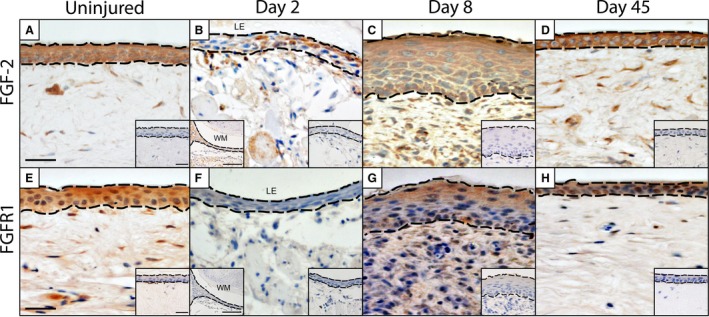
FGF‐2 ligand and receptor expression by keratinocytes. Prior to injury, expression of FGF‐2 (A) by keratinocytes is widespread. This robust pattern of expression is maintained by keratinocytes at the wound margin (WM; inset, bottom left‐hand corner) and throughout the WE before (B) and following (C, D) wound closure. Although FGFR1 is robustly expressed by keratinocytes prior to injury (E), it is notably absent from the 2 DPW wound epithelium (F), including the wound margin (inset, bottom left‐hand corner). FGFR1 expression resumes by 8 DPW (G) and resembles its pre‐injury pattern of expression by day 45 (H). Dashed line indicates the junction between the overlying epidermis and the deeper dermis. Omission controls = insets, bottom right hand corner. Scale bar: 10 μm.
TGFβ signalling detected in keratinocytes of the original and regenerating epidermis
Next, we investigated TGFβ, focusing on the intracellular mediator phosphorylated SMAD2 (pSMAD2), and the ligands TGβ1 and activin βA. Phosphorylation of SMAD2 is considered to represent a readout of canonical TGFβ activation (Penn et al. 2012). Prior to injury, pSMAD2 expression was observed throughout the epidermis (Fig. 4A). Two days following the biopsy (2 DPW), pSMAD2 expression was restricted to only a few scattered neo‐keratinocytes across the wound site (Fig. 4B). However, following wound closure (8 DPW), pSMAD2 was strongly expressed by most keratinocytes of the WE (Fig. 4C). Once wound healing was complete (45 DPW), the widespread pattern of pSMAD2 expression closely resembled that of the original (uninjured) epidermis (Fig. 4D).
Figure 4.
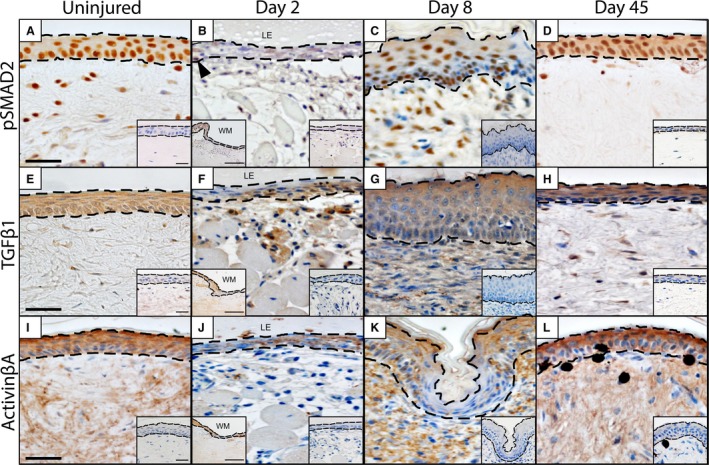
TGFβ1 and activin βA expression by keratinocytes. Phosphorylated SMAD2 (pSMAD2), a readout of TGFβ signalling, is widely expressed by keratinocytes prior to injury (A). Following injury, pSMAD2 expression is present within the wound margin (inset, bottom left‐hand corner) but otherwise rare among cells of the WE (2 DPW; indicated by black arrow heads; B). However, as the WE thickens (8 DPW) there is a marked increase in pSMAD2 expression (C). At 45 DPW, pSMAD2 expression resembles that of the original epidermis (D). We also observed a similar pattern of expression for both TGFβ1 (E–H) and activin βA (I–L). Dashed line indicates the junction between the overlying epidermis and the deeper dermis. Omission controls = insets, bottom right hand corner. Scale bar: 10 μm.
Given our evidence for canonical TGFβ signalling, we shifted our investigation to two important ligands: TGFβ1 and activin βA. As with pSMAD2, TGFβ1 was strongly expressed by the keratinocytes of the uninjured epidermis (Fig. 4E). Prior to re‐epithelialization (2 DPW), protein expression of TGFβ1 was limited and no longer observed as widespread among keratinocytes (Fig. 4F). However, following re‐epithelialization (8 DPW), TGFβ1 immunostaining was again demonstrated by most keratinocytes (Fig. 4G) and by 45 DPW it closely resembled that of the uninjured epidermis (Fig. 4H).
Matching the pattern of distribution for pSMAD2 (and TGFβ1), activin βA demonstrated a near ubiquitous pattern of expression within the uninjured epidermis (Fig. 4I). Prior to re‐epithelialization (2 DPW), keratinocytes expressing activin βA were limited (Fig. 4J). By 8 DPW, a non‐uniform pattern of protein expression was observed, with some regions of the WE expressing a near ubiquitous pattern of activin βA expression, whereas adjacent regions were completely devoid of immunoreactivity (Fig. 4K). At 45 DPW, activin βA has returned to its pre‐injury expression profile of widespread expression (Fig. 4L).
Discussion
Our data reveal that normal (uninjured) and regenerating gecko epidermis robustly expresses a diverse panel of growth factor ligands and receptors: VEGF and its receptors VEGFR1, VEGFR2 and pVEGFR2; FGF‐2 and its receptor FGFR1; as well as pSMAD2, TGFβ1 and activin βA. Curiously, only VEGFR1 and FGFR1 are dynamically expressed, and only so during the earliest phases of re‐epithelialization (Fig. 5). Otherwise, our proteins of interest were constitutively present. Among mammals, a comparable pattern of protein expression has been documented for keratinocytes in rodents and humans (Vrabec et al. 1994; Weninger et al. 1996; Mildner et al. 1999; Steiling & Werner, 2003; Man et al. 2006; Zhu et al. 2012). Combined, our data point towards both homeostatic and injury‐mediated roles for each of these cytokines.
Figure 5.
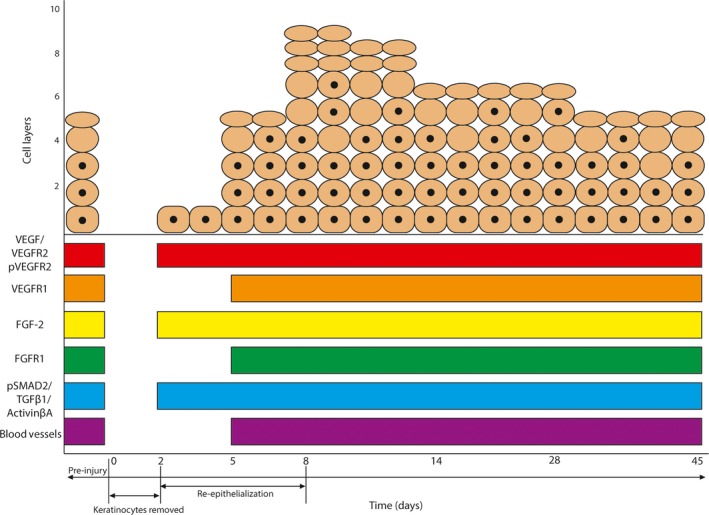
Summary of growth factor expression in the homeostatic and regenerating epidermis. With the exception of FGFR1 and VEGFR1, which were notably absent during re‐epithelialization, all other growth factors were constitutively expressed throughout scar‐free wound healing.
VEGF signalling
Prior to and throughout the wound healing program, we documented robust co‐localization of VEGF/VEGFR2, along with an overlapping pattern of expression of pVEGFR2, by virtually all keratinocytes of the normal epidermis and WE. In addition, most keratinocytes prior to and following injury also co‐express VEGFR1/VEGFR2. A comparable pattern of VEGF receptor and ligand expression has also been reported for humans (Weninger et al. 1996; Mildner et al. 1999; Zhu et al. 2012) and mice (Man et al. 2006). In mammals, VEGF signaling by keratinocytes is most commonly interpreted as functioning in a paracrine manner to regulate vascular homeostasis in the dermis, and to induce neo‐vascularization during wound healing and tumour formation (Brown et al. 1992; Lauer et al. 2000; Zhang et al. 2004). Although a similar function cannot be excluded for geckos, we did not observe the induction of exuberant neovascularization or the formation of granulation tissue. Instead, as has been previously reported (Peacock et al. 2015), the density of newly formed blood vessels during wound healing was comparable to the original (uninjured) dermis, despite the widespread evidence for VEGF signalling. Moreover, most newly formed blood vessels observed are structurally mature (i.e. mural cell‐supported), and likely no longer VEGF‐dependent (Alon et al. 1995; Banks et al. 1998). Combined, these data indicate that VEGF may be functioning in a role beyond angiogenesis.
VEGFR1 is noticeably absent from the leading edge of the WE at 2 DPW. In contrast, when mice receive cutaneous wounds, VEGFR1 expression is necessary for keratinocyte proliferation and rapid wound closure (Wilgus et al. 2005). Whether these differences in VEGFR1 expression represent a fundamental distinction between scar‐free and scar‐forming repair programs remains to be determined. It is also worth noting that many stimulatory effects of VEGF are mediated through VEGFR2, whereas VEGFR1 is hypothesized to participate in a more regulatory capacity (Gerber et al. 1999; Rahimi et al. 2000; Zeng et al. 2001a,b; Roberts et al. 2004).
Our demonstration of VEGF/VEGFR2 co‐expression, and overlapping pVEGFR2 expression, by keratinocytes points towards autocrine signalling. Among mammals, autocrine VEGF signalling promotes keratinocyte survival and proliferation (Johnson & Wilgus, 2012), and attenuates ultraviolet (UV) radiation damage (Zhu et al. 2012). UV irradiation is a well‐known mutagen and damaging stressor of the skin, leading to erythema, inflammation and carcinogenesis (Brauchle et al. 1996; Johnson & Wilgus, 2012). Various in vivo and in vitro investigations have demonstrated that VEGF and VEGFRs are widely expressed by keratinocytes in mammals (Brauchle et al. 1996; Wilgus et al. 2005; Man et al. 2006; Zhu et al. 2012), and that exposure to UV radiation activates VEGFR‐mediated pro‐survival (anti‐apoptotic) mechanisms in these cells (Zhu et al. 2013). Paradoxically, UV irradiation (at low doses) of the epidermis is beneficial, promoting vitamin D3 (cholecalciferol) photobiosynthesis (Brauchle et al. 1996). Indeed, for many reptiles – including nocturnal and/or crepuscular species – cutaneous vitamin D3 synthesis and metabolism is an essential physiological process (Holick, 1995; Carman et al. 2000; Acierno et al. 2008). In the absence of UV (especially UVB; spectrum range 290–315 nm) exposure or dietary supplementation, reptiles may develop metabolic bone disease, a complex spectrum of disorders that includes rickets, lethargy and anorexia (Klaphake, 2010). Perhaps not surprisingly, species with less exposure to UV, such as those active at dawn and dusk (i.e. crepuscular taxa), appear to have more sensitive mechanisms for photobiosynthesis of vitamin D3 compared with diurnal species (Carman et al. 2000). We propose that constitutive VEGFR expression by gecko keratinocytes may participate in attenuating photodamage while maximizing optimal photobiosynthesis.
FGF signalling
Like VEGF, FGF‐2 is robustly expressed by gecko keratinocytes prior to injury and throughout the process of wound healing. In mammals, FGF‐2 has a well‐documented role in stimulating keratinocyte proliferation (Vrabec et al. 1994; Steiling & Werner, 2003) and may also play a protective role by regulating DNA repair and maintaining genomic integrity in keratinocyte progenitor populations (Harfouche et al. 2010). Treatment of keratinocyte progenitor cells with exogenous FGF‐2 prior to irradiation not only increased the rate of double‐strand break repair but also demonstrated a pro‐survival effect (Harfouche et al. 2010). Although we were unable to co‐localize FGF‐2 with its receptor FGFR1, data from adjacent sections clearly demonstrate that both proteins are detected in most keratinocytes (except prior to re‐epithelialization). Hence, FGF‐2/FGFR1 are almost certainly co‐expressed, indicating that FGF signalling may be activated. The absence of FGFR1 expression during re‐epithelization (at 2 DPW) suggests that FGF‐2 could be signalling through another one of its other receptors, possibly FGFR2 (for which it has the highest binding affinity after FGFR1; Mansukhani et al. 1992; Ornitz et al. 1996).
In contrast to previous studies investigating FGF expression in lizard skin (L. guichenoti and P. sicula; Alibardi & Lovicu, 2010; Alibardi, 2012), we determined that FGF‐2 is robustly expressed in the normal epidermis, as well as during cutaneous wound healing. Although this may represent a taxonomic difference, it may also relate to the ability of geckos to heal scar‐free. Across lizard species there is a spectrum of cutaneous wound healing responses, from scarring to scarless (Wu et al. 2014). Whereas L. guichenoti and P. sicula are capable of regenerating their tails, it remains unclear if their scar‐free capabilities include wounds restricted to the skin.
TGFβ signalling
TGFβ is a multi‐functional cytokine superfamily with numerous roles related to tissue homeostasis and wound healing (Penn et al. 2012). We determined that the canonical TGFβ/activin signalling pathway is activated in the gecko epidermis, based on the near ubiquitous expression of pSMAD2 in the uninjured skin as well as during scar‐free wound healing. We investigated two key factors known to play a role in this signalling pathway: TGFβ1 and activin βA. In mammals, TGFβ1 is known to limit keratinocyte proliferation in normal skin (Ramirez et al. 2014). Curiously, although TGFβ1 is localized to the uppermost layers of keratinocytes in human epidermis (Gold et al. 2000), we found it expressed throughout all epidermal strata in the original and fully regenerated epidermis. One possible explanation is that geckos, as squamate reptiles, undergo coordinated body‐wide skin‐shedding or ecdysis events (Maderson, 1964, 1965). This periodic process requires the synchronization of stratum germinativum proliferation and keratinocyte differentiation to duplicate the epidermis, resulting in an inner and outer epidermal generation (Fig. 6; see also Maderson, 1964, 1965). During ecdysis, the outer generation is sloughed off while the inner generation is retained. We propose that constitutive expression of TGFβ1 could play a role in coordinating ecdysis by restricting keratinocyte proliferation. TGFβ1 also participates in wound closure by promoting keratinocyte migration. More specifically, TGFβ1 drives epithelial‐to‐mesenchymal transitions (Räsänen & Vaheri, 2010; Weber et al. 2012; Lamouille et al. 2014), and increases the expression of, while acting on, the integrins (Gailit et al. 1994; Li et al. 2006; Margadant & Sonnenberg, 2010).
Figure 6.
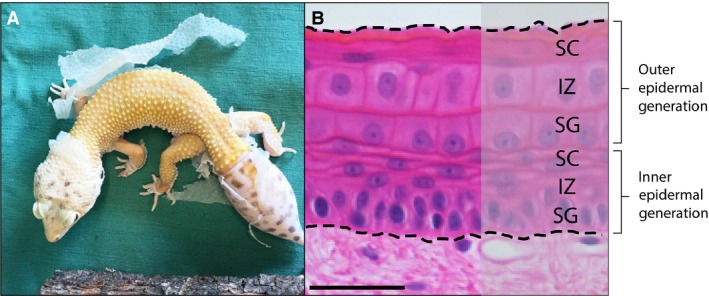
Synchronized body‐wide epidermal shedding, and a possible role for TGFβ/activin signalling. Geckos, along with other squamates and tuatara, undergo discontinuous episodes of ecdysis, resulting in the coordinated sloughing of large patches of the epidermis (A). Coordinated ecdysis requires the synchronization of stratum germinativum proliferation to duplicate the epidermis, creating an inner and outer epidermal generation. The outer generation (B) is eventually sloughed off. We hypothesize a role for TGFβ1 in coordinating body‐wide ecdysis. TGFβ1 is known to inhibit keratinocyte proliferation and prevent hyperproliferation during wound healing. As evidenced by pSMAD2 expression, the canonical TGFβ/activin signalling pathway is constitutively activated, suggesting that it may participate in the suppression of continuous shedding. Haematoxoylin & eosin. Scale bar: 10 μm.
Compared with TGFβ1, less is known about the function(s) of activin in the epidermis although a role in resolving the epidermal defect is indicated (Wankell et al. 2001). Activin, and the closely related inhibin, are dimer proteins best known for their roles in reproductive organs. Activin βA contributes to one homodimer, activin A (βA, βA), and one heterodimer, activin AB (βA, βB) (Werner & Alzheimer, 2006); therefore, one or both could be expressed in geckos. Human (unpublished, but cited in Werner & Alzheimer, 2006) and gecko (Gilbert et al. 2013) keratinocytes upregulate activin βA in response to injury, whereas mouse keratinocytes upregulate activin βA and activin βB and express activin A (Werner & Alzheimer, 2006). In mammals, inhibiting activin (using a transgenic mouse line that overexpresses the antagonist follistatin) delays wound healing but also reduces the amount of granulation tissue formed, resulting in a smaller scar (Wankell et al. 2001). Outside of mammals, the importance of activin βA expression during wound healing has also been demonstrated in zebrafish (Jaźwińska et al. 2007).
Conclusions
Our findings provide important new evidence about normal and injury‐mediated growth factor expression by keratinocytes, and expand the comparative framework of keratinocyte biology to include a reptilian species capable of scar‐free wound healing. Although the functional implications of these expression patterns remain poorly understood, roles beyond angiogenesis are strongly indicated. We propose that constitutive expression of growth factor ligands and receptors by keratinocytes may participate in photo‐protective and skin‐shedding functions, in addition to re‐epithelialization. Moreover, our data provide evidence of autocrine signalling by reptilian epidermal cells.
Author contributions
N. Subramaniam contributed to the experimental design, data analysis and interpretation, and drafted the manuscript. J. Petrik contributed to the experimental design, data analysis and interpretation, and made critical revisions to the drafted manuscript. M. Vickaryous contributed to the experimental design, data analysis and interpretation, and drafted the manuscript. All authors read and approved the submitted manuscript. Contributors who do not qualify as authors include E. A. B. Gilbert, K. Jacyniak and R. P. McDonald, who assisted with tissue collection and animal care, and J. Hung, R. Tari and S. Donato, who performed technical replicates of immunostaining.
Supporting information
Fig. S1. Macroscopic sequence of scar‐free wound healing. (A‐D) Biopsy wounds (3 mm diameter; indicated by black hatched lines) were created on the dorsal surface of the tail. Four time points were collected: uninjured (original) skin; day 2 (prior to re‐epithelialization); day 8 (following re‐epithelialization); day 45 (after wound healing is complete). Note: this panel represents four different individuals.
Fig. S2. Histology of scar‐free wound healing. At each sampled time point, representative sections were stained with haematoxylin and eosin (H&E) to visualize structure and morphology. (E‐H). Prior to injury, proliferating cell nuclear antigen (PCNA) immunostaining is primarily restricted to the stratum germinativum. There is very little proliferation at day 2 (indicated by white arrowhead), but by 8 DPW there is evidence of cell proliferation within multiple layers of the epidermis. At 45 DPW, proliferation returns to pre‐injury levels and is largely confined to the stratum germinativum. (I‐L). Scale bar: 10 μm.
Fig. S3. Negative controls for immunofluorescent figures. Top row: vascular endothelial growth factor A (VEGF) expression is absent in the omission control (OC), and in the trial applying both the blocking peptide and the primary antibody (P+Ab). Middle row: VEGF receptor 2 (VEGFR2) expression is absent in the omission control (OC). Bottom row: VEGFR1 expression is absent in the omission control (OC). Scale bar: 10 μm.
Acknowledgements
The authors sincerely thank H. Coates, E. A. B. Gilbert, J. Hung, K. Jacyniak, R. P. McDonald, K. Osz, H. M. Peacock, S. Donato and R. Tari for their technical assistance and discussion, and two anonymous reviewers for their valuable input. The activin βA antibody was a kind donation from Dr Paul Sawchenko from the Salk Institute for Biological Studies, in La Jolla, California. This work was supported by the Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant (400358) to M. K. Vickaryous, NSERC Discovery Grant (401013) to J. Petrik, and Ontario Veterinary College Scholarship to N. Subramaniam.
References
- Abraham JA, Klagsbrun M (1988) Modulation of wound repair by members of the fibroblast growth factor family In: The Molecular and Cellular Biology of Wound Repair (ed. Clark RAF.), pp. 195–248. Boston: Springer USA. [Google Scholar]
- Acierno MJ, Mitchell MA, Zachariah TT, et al. (2008) Effects of ultraviolet radiation on plasma 25‐hydroxyvitamin D3 concentrations in corn snakes (Elaphe guttata). Am J Vet Res 69, 294–297. [DOI] [PubMed] [Google Scholar]
- Alibardi L (2012) Observations on FGF immunoreactivity in the regenerating tail blastema, and in the limb and tail scars of lizard suggest that FGFs are required for regeneration. Belg J Zoo 142, 23–38. [Google Scholar]
- Alibardi L, Lovicu FJ (2010) Immunolocalization of FGF1 and FGF2 in the regenerating tail of the lizard Lampropholis guichenoti: implications for FGFs as trophic factors in lizard tail regeneration. Acta Histochem 112, 459–473. [DOI] [PubMed] [Google Scholar]
- Allam AA, Daza JD, Abo‐Eleneen RE (2016) Histology of the skin of three limbless squamates dwelling in mesic and arid environments. Anat Rec 299, 979–989. [DOI] [PubMed] [Google Scholar]
- Alon T, Hemo I, Itin A, et al. (1995) Vascular endothelial growth factor acts as a survival factor for newly formed retinal vessels and has implications for retinopathy of prematurity. Nat Med 1, 1024–1028. [DOI] [PubMed] [Google Scholar]
- Azevedo AS, Grotek B, Jacinto A, et al. (2011) The regenerative capacity of the zebrafish caudal fin is not affected by repeated amputations. PLoS ONE 6, e22820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bamberger C, Schärer A, Antsiferova M, et al. (2005) Activin controls skin morphogenesis and wound repair predominantly via stromal cells and in a concentration‐dependent manner via keratinocytes. Am J Pathol 167, 733–747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banks RE, Forbes MA, Kinsey SE, et al. (1998) Release of the angiogenic cytokine vascular endothelial growth factor (VEGF) from platelets: significance for VEGF measurements and cancer biology. Br J Cancer 77, 956–964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barrientos S, Stojadinovic O, Golinko MS, et al. (2008) Growth factors and cytokines in wound healing. Wound Repair Regen 16, 585–601. [DOI] [PubMed] [Google Scholar]
- Basilico C, Moscatelli D (1992) The FGF family of growth factors and oncogenes. Adv Cancer Res 59, 115–165. [DOI] [PubMed] [Google Scholar]
- Battegay EJ (1995) Angiogenesis: mechanistic insights, neovascular diseases, and therapeutic prospects. J Med 73, 333–346. [DOI] [PubMed] [Google Scholar]
- Bellairs AA, Bryant SV (1985) Autotomy and regeneration in reptiles. Biol Reptilia 15, 310–401. [Google Scholar]
- Blyth E (1854) Proceedings of the Society. Report of the Curator, Zoology Department. J Asiat Soc Bengal 23, 737–740. [Google Scholar]
- Brauchle M, Funk JO, Kind P, et al. (1996) Ultraviolet B and H2O2 are potent inducers of vascular endothelial growth factor expression in cultured keratinocytes. J Biol Chem 271, 21793–21797. [DOI] [PubMed] [Google Scholar]
- Brown LF, Yeo K, Berse B, et al. (1992) Expression of vascular permeability factor (vascular endothelial growth factor) by epidermal keratinocytes during wound healing. J Exp Med 176, 1375–1379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carman EN, Ferguson GW, Gehrmann WH, et al. (2000) Photobiosynthetic opportunity and ability for UV‐B generated vitamin D synthesis in free‐living house geckos (Hemidactylus turcicus) and Texas spiny lizards (Sceloporus olivaceous). Copeia 2000, 245–250. [Google Scholar]
- Christenson LJ, Phillips PK, Weaver AL, et al. (2005) Primary closure vs second‐intention treatment of skin punch biopsy sites: a randomized trial. Arch Dermatol 141, 1093–1099. [DOI] [PubMed] [Google Scholar]
- Clegg LW, Gabhann F (2015) Site‐specific phosphorylation of VEGFR2 is mediated by receptor trafficking: insights from a computational model. PLoS Comp Biol 11, e1004158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delorme SL, Lungu IM, Vickaryous MK (2012) Scar‐free wound healing and regeneration following tail loss in the leopard gecko, Eublepharis macularius . Anat Rec 295, 1575–1595. [DOI] [PubMed] [Google Scholar]
- Derynck R, Miyazono K. (eds.) 2008) The TGF‐[beta] Family, vol. 50 Cold Spring Harbor, New York: CSHL Press. [Google Scholar]
- Dunn L, Prosser HC, Tan JT, et al. (2013) Murine model of wound healing. J Vis Exp 75, e50265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferguson MW, O'Kane S (2004) Scar‐free healing: from embryonic mechanisms to adult therapeutic intervention. Philos Trans R Soc Lond B Biol Sci 359, 839–850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrara N (2004) Vascular endothelial growth factor: basic science and clinical progress. Endocr Rev 25, 581–611. [DOI] [PubMed] [Google Scholar]
- Frank S, Hübner G, Breier G, et al. (1995) Regulation of vascular endothelial growth factor expression in cultured keratinocytes. Implications for normal and impaired wound healing. J Biol Chem 270, 12607–12613. [DOI] [PubMed] [Google Scholar]
- Gailit J, Clark RA, Welch MP (1994) TGF‐β1 stimulates expression of keratinocyte integrins during re‐epithelialization of cutaneous wounds. J Invest Dermatol 103, 221–227. [DOI] [PubMed] [Google Scholar]
- Galiano RD, Michaels V, Dobryansky M, et al. (2004) Quantitative and reproducible murine model of excisional wound healing. Wound Repair Regen 12, 485–492. [DOI] [PubMed] [Google Scholar]
- Gerber HP, Hillan KJ, Ryan AM, et al. (1999) VEGF is required for growth and survival in neonatal mice. Development 12, 1149–1159. [DOI] [PubMed] [Google Scholar]
- Gilbert RW, Vickaryous MK, Viloria‐Petit AM (2013) Characterization of TGFβ signaling during tail regeneration in the leopard gecko (Eublepharis macularius). Dev Dynam 242, 886–896. [DOI] [PubMed] [Google Scholar]
- Gilbert RW, Vickaryous MK, Viloria‐Petit AM (2016) Signalling by transforming growth factor β isoforms in wound healing and tissue regeneration. J Dev Biol 4, 21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Godwin JW, Rosenthal N (2014) Scar‐free wound healing and regeneration in amphibians: immunological influences on regenerative success. Differentiation 87, 66–75. [DOI] [PubMed] [Google Scholar]
- Gold LI, Jussila T, Fusenig NE, et al. (2000) TGF‐β isoforms are differentially expressed in increasing malignant grades of HaCaT keratinocytes, suggesting separate roles in skin carcinogenesis. J Pathol 190, 579–588. [DOI] [PubMed] [Google Scholar]
- Gurtner GC, Werner S, Barrandon Y, et al. (2008) Wound repair and regeneration. Nature 453, 314–321. [DOI] [PubMed] [Google Scholar]
- Harfouche G, Vaigot P, Rachidi W, et al. (2010) Fibroblast growth factor type 2 signaling is critical for DNA repair in human keratinocyte stem cells. Stem Cells 28, 1639–1648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holick MF (1995) Environmental factors that influence the cutaneous production of vitamin D. Am J Clin Nutr 61, 638S–645S. [DOI] [PubMed] [Google Scholar]
- Järvinen TA, Ruoslahti E (2010) Target‐seeking antifibrotic compound enhances wound healing and suppresses scar formation in mice. Proc Natl Acad Sci U S A 107, 21671–21676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaźwińska A, Badakov R, Keating MT (2007) Activin‐βA signaling is required for zebrafish fin regeneration. Curr Biol 17, 1390–1395. [DOI] [PubMed] [Google Scholar]
- Jensen‐Jarolim E. (ed.) (2013) Comparative Medicine: Anatomy and Physiology. New York: Springer Science & Business Media. [Google Scholar]
- Johnson KE, Wilgus TA (2012) Multiple roles for VEGF in non‐melanoma skin cancer: angiogenesis and beyond. J Skin Cancer 2012, 483439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klaphake E (2010) A fresh look at metabolic bone diseases in reptiles and amphibians. Vet Clin North Am Exot Anim Pract 13, 375–392. [DOI] [PubMed] [Google Scholar]
- Kottakis F, Polytarchou C, Foltopoulou P, et al. (2011) FGF‐2 regulates cell proliferation, migration, and angiogenesis through an NDY1/KDM2B‐miR‐101‐EZH2 pathway. Mol Cell 43, 285–298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamouille S, Xu J, Derynck R (2014) Molecular mechanisms of epithelial–mesenchymal transition. Nat Rev Mol Cell Biol 15, 178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larson BJ, Longaker MT, Lorenz HP (2010) Scarless fetal wound healing: a basic science review. Plast Reconstr Surg 126, 1172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lauer G, Sollberg S, Cole M, et al. (2000) Expression and proteolysis of vascular endothelial growth factor is increased in chronic wounds. J Invest Dermatol 115, 12–18. [DOI] [PubMed] [Google Scholar]
- Lévesque M, Villiard É, Roy S (2010) Skin wound healing in axolotls: a scarless process. J Exp Zool B Mol Dev Evol 314, 684–697. [DOI] [PubMed] [Google Scholar]
- Li Y, Fan J, Chen M, et al. (2006) Transforming growth factor‐alpha: a major human serum factor that promotes human keratinocyte migration. J Invest Dermatol 126, 2096–2105. [DOI] [PubMed] [Google Scholar]
- Lorenz HP, Adzick NS (1993) Scarless skin wound repair in the fetus. Western J Med 159, 350. [PMC free article] [PubMed] [Google Scholar]
- Maderson PFA (1964) The skin of lizards and snakes. J Herpet 3, 151–154. [Google Scholar]
- Maderson PFA (1965) Histological changes in the epidermis of snakes during the sloughing cycle. Proc Zool Soc Long 146, 98–113. [Google Scholar]
- Man XY, Yang XH, Cai SQ, et al. (2006) Immunolocalization and expression of vascular endothelial growth factor receptors (VEGFRs) and neuropilins (NRPs) on keratinocytes in human epidermis. Mol Med 12, 127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mansukhani A, Dell'Era P, Moscatelli D, et al. (1992) Characterization of the murine BEK fibroblast growth factor (FGF) receptor: activation by three members of the FGF family and requirement for heparin. Proc Natl Acad Sci U S A 89, 3305–3309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Margadant C, Sonnenberg A (2010) Integrin–TGF‐β crosstalk in fibrosis, cancer and wound healing. EMBO Rep 11, 97–105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin P (1997) Wound healing – aiming for perfect skin regeneration. Science 276, 75–81. [DOI] [PubMed] [Google Scholar]
- McLean KE, Vickaryous MK (2011) A novel amniote model of epimorphic regeneration: the leopard gecko, Eublepharis macularius . BMC Dev Biol 11, 1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mildner M, Weninger W, Trautinger F, et al. (1999) UVA and UVB radiation differentially regulate vascular endothelial growth factor expression in keratinocyte‐derived cell lines and in human keratinocytes. Photochem Photobiol 70, 674–679. [PubMed] [Google Scholar]
- Moustakas A, Heldin CM (2009) The regulation of TGFbeta signal transduction. Development. J Cell Sci 136, 3699–3714. [DOI] [PubMed] [Google Scholar]
- Nissen NN, Polverini P, Koch AE, et al. (1998) Vascular endothelial growth factor mediates angiogenic activity during the proliferative phase of wound healing. Am J Pathol 152, 1445–1452. [PMC free article] [PubMed] [Google Scholar]
- Ogunjimi AA, Zeqiraj E, Ceccarelli DF, et al. (2012) Structural basis for specificity of TGFβ family receptor small molecule inhibitors. Cell Signal 24, 476–483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Kane S, Ferguson MW (1997) Transforming growth factor βs and wound healing. Int J Biochem Cell Biol 29, 63–78. [DOI] [PubMed] [Google Scholar]
- Ornitz DM, Itoh N (2001) Fibroblast growth factors. Genome Biol 2, 1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ornitz DM, Xu J, Colvin JS, et al. (1996) Receptor specificity of the fibroblast growth factor family. J Biol Chem 271, 15292–15297. [DOI] [PubMed] [Google Scholar]
- Ortega S, Ittmann M, Tsang SH, et al. (1998) Neuronal defects and delayed wound healing in mice lacking fibroblast growth factor 2. Proc Natl Acad Sci U S A 95, 5672–5677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pastar I, Stojadinovic O, Yin NC, et al. (2014) Epithelialization in wound healing: a comprehensive review. Adv Wound Care 3, 445–464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peacock HM, Gilbert EA, Vickaryous MK (2015) Scar‐free cutaneous wound healing in the leopard gecko, Eublepharis macularius . J Anat 227, 596–610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Penn JW, Grobbelaar AO, Rolfe KJ (2012) The role of the TGF‐β family in wound healing, burns and scarring: a review. Int J Burns Trauma 2, 18–28. [PMC free article] [PubMed] [Google Scholar]
- Rahimi N, Dayanir V, Lashkari K (2000) Receptor chimeras indicate vascular endothelial growth factor receptor‐1 (VEGFR‐1) modulate mitogenic activity of VEGFR‐2 in endothelial cells. J Biol Chem 275, 16986–16992. [DOI] [PubMed] [Google Scholar]
- Ramirez H, Patel SB, Pastar I (2014) The role of TGFβ signaling in wound epithelialization. Adv Wound Care 3, 482–491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Räsänen K, Vaheri A (2010) TGF‐beta1 causes epithelial‐mesenchymal transition in HaCaT derivatives, but induces expression of COX‐2 and migration only in benign, not in malignant keratinocytes. J Dermatol Sci 58, 97–104. [DOI] [PubMed] [Google Scholar]
- Richardson R, Slanchev K, Kraus C, et al. (2013) Adult zebrafish as a model system for cutaneous wound‐healing research. J Invest Dermatol 133, 1655–1665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts AB (1998) Molecular and cell biology of TGF‐β. Miner Electrolyte Metab 24, 111–119. [DOI] [PubMed] [Google Scholar]
- Roberts DM, Kearney JB, Johnson JH (2004) The vascular endothelial growth factor (VEGF) receptor Flt‐1 (VEGFR‐1) modulates Flk‐1 (VEGFR‐2) signaling during blood vessel formation. Am J Pathol 164, 1531–1535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ross S, Hill CS (2008) How the Smads regulate transcription. Int J Biochem Cell Biol 40, 383–408. [DOI] [PubMed] [Google Scholar]
- Santos D, Rita AM, Casanellas I, et al. (2016) Ear wound regeneration in the African spiny mouse Acomys cahirinus . Regeneration 3, 52–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Satish L, Kathju S (2010) Cellular and molecular characteristics of scarless versus fibrotic wound healing. Dermatol Res 2010, 790234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schumacher J, Yelen T (2006) Anesthesia and analgesia. Reptile Med Surg 2, 442–452. [Google Scholar]
- Seifert AW, Maden M (2014) New insights into vertebrate skin regeneration. Int Rev Cell Mol Biol 310, 129–169. [DOI] [PubMed] [Google Scholar]
- Seifert AW, Monaghan JR, Voss SR, et al. (2012a) Skin regeneration in adult axolotls: a blueprint for scar‐free healing in vertebrates. PLoS ONE 7, e32875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seifert AW, Kiama SG, Seifert MG, et al. (2012b) Skin shedding and tissue regeneration in African spiny mice (Acomys). Nature 489, 561–565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singer AJ, Clark RA (1999) Cutaneous wound healing. N Engl J Med 341, 738–746. [DOI] [PubMed] [Google Scholar]
- Slominski A, Paus R, Wortsman J (1993) On the potential role of proopiomelanocortin in skin physiology and pathology. Mol Cell Endocrinol 93, C1–C6. [DOI] [PubMed] [Google Scholar]
- Sogabe Y, Abe M, Yokoyama Y, et al. (2006) Basic fibroblast growth factor stimulates human keratinocyte motility by Rac activation. Wound Repair Regen 14, 457–462. [DOI] [PubMed] [Google Scholar]
- Steiling H, Werner S (2003) Fibroblast growth factors: key players in epithelial morphogenesis, repair and cytoprotection. Curr Opin Biotechnol 14, 533–537. [DOI] [PubMed] [Google Scholar]
- Stewart S, Stankunas K (2012) Limited dedifferentiation provides replacement tissue during zebrafish fin regeneration. Dev Biol 365, 339–349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sulyok S, Wankell M, Alzheimer C, et al. (2004) Activin: an important regulator of wound repair, fibrosis, and neuroprotection. Mol Cell Endocrinol 225, 127–132. [DOI] [PubMed] [Google Scholar]
- Szydłowski P, Madej JP, Mazurkiewicz‐Kania M (2015) Ultrastructure and distribution of chromatophores in the skin of the leopard gecko (Eublepharis macularius). Acta Zool 97, 370–375. [Google Scholar]
- Tonnesen MG, Feng X, Clark RA (2000) Angiogenesis in wound healing. J Investig Dermatol Symp Proc 5, 40–46. [DOI] [PubMed] [Google Scholar]
- Vrabec JT, Schwaber MK, Davidson JM, et al. (1994) Evaluation of basic fibroblast growth factor in tympanic membrane repair. Laryngoscope 104, 1059–1064. [DOI] [PubMed] [Google Scholar]
- Wankell M, Munz B, Hübner G, et al. (2001) Impaired wound healing in transgenic mice overexpressing the activin antagonist follistatin in the epidermis. EMBO J 20, 5361–5372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weber CE, Li NY, Wai PY, et al. (2012) Epithelial‐mesenchymal transition, TGF‐β, and osteopontin in wound healing and tissue remodeling after injury. J Burn Care Res 33, 311–318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weninger W, Uthman A, Pammer J, et al. (1996) Vascular endothelial growth factor production in normal epidermis and in benign and malignant epithelial skin tumors. Lab Invest 75, 647–657. [PubMed] [Google Scholar]
- Werner S (1998) Keratinocyte growth factor: a unique player in epithelial repair processes. Cytokine Growth Factor Rev 9, 153–165. [DOI] [PubMed] [Google Scholar]
- Werner S, Alzheimer C (2006) Roles of activin in tissue repair, fibrosis, and inflammatory disease. Cytokine Growth Factor Rev 17, 157–171. [DOI] [PubMed] [Google Scholar]
- Werner S, Grose R (2003) Regulation of wound healing by growth factors and cytokines. Physiol Rev 83, 835–870. [DOI] [PubMed] [Google Scholar]
- Whimster IW (1978) Nerve supply as a stimulator of the growth of tissues including skin. Clin Exp Dermatol 3, 389–410. [DOI] [PubMed] [Google Scholar]
- Wilgus TA, Matthies AM, Radek KA, et al. (2005) Novel function for vascular endothelial growth factor receptor‐1 on epidermal keratinocytes. Am J Pathol 167, 1257–1266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu MY, Hill CS (2009) TGF‐β superfamily signaling in embryonic development and homeostasis. Dev Cell 16, 329–343. [DOI] [PubMed] [Google Scholar]
- Wu P, Alibardi L, Chuong CM (2014) Regeneration of reptilian scales after wounding: neogenesis, regional difference, and molecular modules. Regeneration 1, 15–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Y, Andresen BT, Yang K, et al. (2010) Association of vascular entothelial growth factor‐634C/G polymorphism and diabetic retinopathy in type 2 diabetic Han Chinese. Exp Biol Med 235, 1204–1211. [DOI] [PubMed] [Google Scholar]
- Zeng H, Dvorak HF, Mukhopadhyay D (2001a) Vascular permeability factor (VPF)/vascular endothelial growth factor (VEGF) receptor‐1 down‐modulates VPF/VEGF receptor‐2‐mediated endothelial cell proliferation, but not migration, through phosphatidylinositol 3‐kinase‐dependent pathways. J Biol Chem 276, 26969–26979. [DOI] [PubMed] [Google Scholar]
- Zeng H, Sanyal S, Mukhopadhyay D (2001b) Tyrosine residues 951 and 1059 of vascular endothelial growth factor receptor‐2 (KDR) are essential for vascular permeability factor/vascular endothelial growth factor‐induced endothelium migration and proliferation, respectively. J Biol Chem 276, 32714–32719. [DOI] [PubMed] [Google Scholar]
- Zhang N, Fang Z, Contag PR, et al. (2004) Tracking angiogenesis induced by skin wounding and contact hypersensitivity using a Vegfr2‐luciferase transgenic mouse. Blood 103, 617–626. [DOI] [PubMed] [Google Scholar]
- Zhu JW, Wu XJ, Luo D, et al. (2012) Activation of VEGFR‐2 signaling in response to moderate dose of ultraviolet B promotes survival of normal human keratinocytes. Int J Biochem Cell Biol 44, 246–256. [DOI] [PubMed] [Google Scholar]
- Zhu JW, Wu XJ, Lu ZF, et al. (2013) Role of VEGF receptors in normal and psoriatic human keratinocytes: evidence from irradiation with different UV sources. PLoS ONE 8, e55463. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fig. S1. Macroscopic sequence of scar‐free wound healing. (A‐D) Biopsy wounds (3 mm diameter; indicated by black hatched lines) were created on the dorsal surface of the tail. Four time points were collected: uninjured (original) skin; day 2 (prior to re‐epithelialization); day 8 (following re‐epithelialization); day 45 (after wound healing is complete). Note: this panel represents four different individuals.
Fig. S2. Histology of scar‐free wound healing. At each sampled time point, representative sections were stained with haematoxylin and eosin (H&E) to visualize structure and morphology. (E‐H). Prior to injury, proliferating cell nuclear antigen (PCNA) immunostaining is primarily restricted to the stratum germinativum. There is very little proliferation at day 2 (indicated by white arrowhead), but by 8 DPW there is evidence of cell proliferation within multiple layers of the epidermis. At 45 DPW, proliferation returns to pre‐injury levels and is largely confined to the stratum germinativum. (I‐L). Scale bar: 10 μm.
Fig. S3. Negative controls for immunofluorescent figures. Top row: vascular endothelial growth factor A (VEGF) expression is absent in the omission control (OC), and in the trial applying both the blocking peptide and the primary antibody (P+Ab). Middle row: VEGF receptor 2 (VEGFR2) expression is absent in the omission control (OC). Bottom row: VEGFR1 expression is absent in the omission control (OC). Scale bar: 10 μm.