

Abstract
Background
Failure of physiologic transformation of spiral arteries has been reported in preeclampsia, fetal growth restriction, fetal death, and spontaneous preterm labor with intact or ruptured membranes. Spiral arteries with failure of physiologic transformation are prone to develop atherosclerotic-like lesions of atherosis. There are striking parallels between preeclampsia and atherosclerotic disease, and between lesions of atherosis and atherosclerosis. Endothelial activation, identified by intercellular adhesion molecule-1 expression, is present in atherosclerotic-like lesions of heart transplantation and considered a manifestation of rejection. Similarly, endothelial activation/dysfunction has been implicated in the pathophysiology of atherosclerosis and preeclampsia. Intercellular adhesion molecule-1-overexpressing-activated endothelial cells are more resistant to trophoblast displacement than nonactivated endothelium and may contribute to shallow spiral artery trophoblastic invasion in obstetrical syndromes having failure of physiologic transformation.
Objective
To determine whether failure of spiral artery physiologic transformation was associated with activation of interstitial extravillous trophoblasts and/or spiral artery endothelium and presence of acute atherosis in the placental basal plate.
Study Design
A cross-sectional study of 123 placentas (19-42 weeks’ gestation) obtained from normal pregnancies (n = 22), preterm prelabor rupture of membranes (n = 26), preterm labor (n = 23), preeclampsia (n = 27), intrauterine fetal death (n = 15), and small for gestational age (n = 10) was performed. Failure of spiral artery physiologic transformation and presence of cell activation was determined using immunohistochemistry of placental basal plates containing a median of 4 (minimum: 1; maximum: 9) vessels per placenta. Endothelial/trophoblast cell activation was defined by the expression of intercellular adhesion molecule-1 (ICAM-1). Investigators examining microscopic sections were blinded to clinical diagnosis. Pairwise comparisons among placenta groups were performed with the Fisher’s exact and Wilcoxon rank sum tests using a Bonferroni-adjusted level of significance (.025).
Results
87% (94/108) of placentas having spiral arteries with failure of physiologic transformation (actin-positive and cytokeratin-negative) in the basal plate, and 0% (0/15) of placentas having only spiral arteries with complete physiologic transformation (cytokeratin-positive and actin-negative), had arterial endothelial and/or interstitial extravillous trophoblasts reactive with the ICAM-1 activation marker (P < .001). A significant correlation (R2 = 0.84) was found between expression of spiral artery endothelial and interstitial extravillous trophoblast ICAM-1 (P < .001) in activated placentas. Lesions of atherosis were found in 31.9% (30/94) of placentas with complete and/or partial failure of physiologic transformation of spiral arteries that were ICAM-1-positive, in none of the 14 placentas with failure of physiologic transformation that were ICAM-1-negative, and in none of the 15 placentas with complete spiral artery physiologic transformation without failure (P = .001). All placentas (30/30, 100%) with atherosis were identified in placentas having concomitant spiral artery endothelial and interstitial extravillous trophoblast activation.
Conclusion
Failure of spiral artery physiologic transformation in the placental basal plate is associated with interstitial extravillous trophoblast and arterial endothelial activation along with increased frequency of spiral artery atherosis. These findings may be used to improve the characterization of different disorders of the placental bed such as in refining the existing tools for the early prediction of risk for preterm, preeclamptic, and other abnormal pregnancies.
Keywords: acute atherosis, endothelial activation, failure of physiologic transformation, ICAM-1, placental basal plate, spiral arteries, trophoblast activation
Introduction
Remodeling of the spiral arteries during gestation leads to substantial dilation of these vessels, which at the point of discharge into the intervillous space, attain a luminal diameter 10 times larger than myometrial spiral arteries. This remodeling is associated with a 10-fold increase in blood supply to the fetoplacental unit in the third trimester compared to the non-pregnant uterus. Spiral artery remodeling, which increases total uteroplacental blood flow from a baseline value of 20-50 mL/min to 450-800 mL/min in singleton pregnancies (as measured utilizing the diffusion equilibrium principle [most often nitrous oxide] or electromagnetic flow probes placed directly on the uterine artery), is considered key to accommodate the increased blood flow to the uteroplacental circulation in the third trimester [1–7]. Initially, physiologic transformation of the spiral arteries includes endothelial vacuolation and smooth muscle swelling [8–11], and it has been attributed in part to immune processes within the decidua [4,12–18]. Subsequently, trophoblasts invading the spiral arteries destroy the smooth muscle in the media, which is replaced by fibrinoid material [1,3,19–21].
Physiologic transformation of the spiral arteries is a normal process with the degree of transformation being greater in the center of the placental bed than in the periphery [21–23]. Full conversion of both decidual and myometrial segments of the approximately 100-120 spiral arteries normally found in the placental bed into large, remodeled uteroplacental vessels is key for normal deep placentation [21,23]. Such deep placentation can be defective when remodeling of the junctional zone of the spiral arteries (arterial segments in the inner third of the myometrium and overlying endometrium) is absent or incomplete [21,24–26]. Insufficient physiologic transformation of the spiral arteries is associated with adverse pregnancy outcomes [3,21,27], including second-trimester spontaneous abortion [28–30], fetal death [30], abruptio placentae [31], preeclampsia [1,32–34], small for gestational age [32–35], preterm labor [36], preterm prelabor rupture of membranes [37,38], and maternal autoimmune diseases [39].
Failure of physiologic transformation of spiral arteries is seen in deep placentation disorders such as preeclampsia with or without intrauterine growth restriction, intrauterine growth restriction without preeclampsia, preterm labor and preterm prelabor rupture of membranes, abruptio placentae, and second-trimester abortion [21]. This failure is characterized by the absence of spiral artery trophoblastic invasion and remodeling [40]. Currently this condition can be identified only in placental bed biopsies because these specimens are required to demonstrate a defect of myometrial transformation [40]. However, physiologic transformation begins in the decidual segment of the spiral arteries during the first wave of trophoblast invasion. Therefore, failure of transformation of the spiral arteries in the decidual segment represents a more severe disorder than lack of transformation of the myometrial segment only. Abnormal transformation of the decidual portion of the spiral arteries can be detected by examining the placental basal plate [33–35,37,39,41–45]. This has practical implications because the basal plate of the placenta is available for examination in all cases while a placental bed biopsy requires a specialized procedure generally performed during a cesarean delivery. Immunohistochemistry [44] allows in-depth examination of the vessel wall through antigen detection of its different cellular components, which reduces the subjectivity of using only conventional light microscopy for vessel recognition in the placental basal plate.
Arteries with failure of physiologic transformation of the spiral arteries (but not those with physiologic transformation) are prone to develop atherosis. Intravascular inflammation and abnormal lipid metabolism can interfere with endothelial cell function, and predispose to atherosclerosis in non-pregnant subjects [46–53]. Women with preeclampsia, the prototypic obstetrical complication characterized by failure of physiologic transformation of the spiral arteries [19,21,32,54–58] have evidence of intravascular inflammation [57,59–62] as well as changes in lipid metabolism [63–66], such as increased concentrations of triglycerides, and low-density lipoprotein, and decreases in low-density lipoprotein particle size and high-density lipoprotein [67]. Macrophage infiltration of the intima and media is identified in both atherosis and the lesions found in transplant vasculopathy [6,43,44,58,68–70]. Therefore, there are striking parallels between preeclampsia and atherosclerosis, as well as the lesions of atherosis and those observed in patients with ischemic heart disease.
Endothelial activation, identified by the expression of intercellular adhesion molecule (ICAM)-1, is present in atherosclerotic-like lesions of heart transplantation [71–76], and is considered a manifestation of rejection. Similarly, endothelial activation/dysfunction has been implicated in the pathophysiology of atherosclerosis [77–79] and preeclampsia [80–83]. Activated endothelial cells with overexpression of cell-surface ICAM-1 [84–86] are more resistant to trophoblast displacement than non-activated endothelial cells [87], and may contribute to shallow spiral artery trophoblastic invasion in obstetrical syndromes associated with failure of physiologic transformation of the spiral arteries.
The purpose of this study was to determine whether there was an association among: (1) failure of physiologic transformation of the spiral arteries in the placental basal plate; (2) endothelial and trophoblast activation; and (3) the presence of atherosis.
Materials and Methods
Study design
A cross-sectional study was performed on 123 placentas (19-42 weeks’ gestation) collected at Hutzel Women’s Hospital, the Detroit Medical Center, under protocols approved by the Wayne State University and Eunice Kennedy Shriver National Institute of Child Health and Human Development Institutional Review Boards, and the analyses were completed by 2015 at CBL Partners for Life, Indianapolis, IN, and the California Medical Innovations Institute, San Diego, CA. Placentas were obtained from: normal pregnancies (n = 22), preterm prelabor rupture of membranes (n = 26), preterm labor (n = 23), preeclampsia (n = 27), intrauterine fetal death (n = 15), and small for gestational age (n = 10). Normal pregnancies were defined as those without medical/obstetrical complications and with birthweights adequate for gestational age (>10th percentile) at term (≥37 weeks of gestation). Seventeen babies were delivered vaginally and five by cesarean (none of the mothers or babies were infected). Preeclampsia was defined as new-onset hypertension >20 weeks of gestation (systolic blood pressure ≥140 mm Hg or diastolic blood pressure ≥90 mm Hg on at least two occasions, 4 hours to 1 week apart), edema, and proteinuria (≥300 mg in a 24-hour urine collection or one dipstick measurement of ≥2+). Infants with fetal growth restriction were considered small for gestational age if they were <10th percentile for weight for their gestational age. Pregnancies were considered preterm when the gestational age was <37 weeks. Birthweight percentiles were calculated using nationwide US natality data [88]. Placentas from mothers with obesity (body mass index ≥30), known to be associated with a pro-inflammatory state [89], were not included in the study. All women provided written informed consent prior to the collection of placenta samples. The Institutional Review Board of the sponsoring institution approved collection and use of samples for research purposes.
Definitions
The terms used in this study follow accepted nomenclature [90]. Briefly, the placental bed is part of the decidua and adjoining myometrium that underlie the placenta and provide blood supply to the intervillous space. Extravillous trophoblasts are all trophoblasts (fetal-derived cells) located outside the placental villi. Interstitial extravillous trophoblasts are placental bed trophoblasts that have entered the decidual stroma but have not yet invaded the spiral artery walls. Extravillous trophoblasts invading spiral artery walls and replacing smooth muscle and endothelium are termed “intra-arterial trophoblasts.” Physiologic transformation of the spiral arteries was defined as the complete trophoblastic invasion of the spiral arteries, partial failure of physiologic transformation of the spiral arteries as the incomplete trophoblastic invasion of the arterial muscular wall, and failure of physiologic transformation of the spiral arteries as the complete absence of trophoblastic replacement of the arterial wall.
Placental specimens and immunohistochemistry
Placental samples (10×10×5 mm) from placental septa near the center of the basal plate [3.5 (SD 0.6) per placenta] of 123 placentas obtained immediately after delivery were embedded in Tissue-Tek® optimum cutting temperature compound (Miles Inc., Elkhart, IN) and snap-frozen in liquid nitrogen. Seven serial sections (a total of seven slides) from each basal plate sample were used for the evaluation of the single-, double-, and triple-antibody techniques performed. Sample slides having basal plate spiral arteries defined immunohistochemically were included in the study, and samples without any spiral artery were excluded. A triple-antibody technique [44,73] with antibodies to α-smooth muscle actin to identify vascular smooth muscle cells, cytokeratin to identify extravillous trophoblasts and intra-arterial trophoblasts, and von Willebrand factor to identify endothelial cells was used to detect complete or partial failure of physiologic transformation and physiologic transformation of spiral arteries (median of 4 [minimum: 1; maximum: 9] vessels per placental basal plate). A triple-antibody immunohistochemistry with α-actin, cytokeratin, and ICAM-1 identified interstitial extravillous trophoblast and spiral artery endothelial cell activation [44,73,91].
Normal term placentas had cytokeratin-positive, α-actin-negative, and von Willebrand factor-positive uteroplacental arteries. Failure of physiologic transformation of the spiral arteries was characterized by cytokeratin-negative, α-actin-positive, and von Willebrand factor-positive spiral arteries. Spiral artery atherosis was cytokeratin-negative, α-actin-positive, and von Willebrand factor-positive and showed the presence of CD68+ macrophages in the intimal area of the spiral arteries. Serial sections (5 μm each) from each placental block were obtained for light microscopy (hematoxylin and eosin) and single, double, and triple immunohistochemistry. The use of immunohistochemistry allowed a clear identification of the spiral arteries in all slides from the different serial sections, demonstrating 100% reproducibility. The deposition of neutral triglycerides and other lipids were examined using oil-red O to confirm atherosis. Individuals examining microscopic sections (C.A.L., H.L.D.) were blinded to clinical diagnoses.
Statistical analysis
Continuous measures were summarized using median, minimum, and maximum, and categorical measures were summarized as frequency (percent). Demographic variables were compared using a Fisher’s exact test (for discrete measures) and the Kruskal-Wallis test (for continuous measures); these tests compared values for atherosis groups (atherosis versus no atherosis). Comparison of continuous measures for groups (atherosis versus no atherosis) were evaluated using the Wilcoxon rank sum test. We used analysis of variance to compare means from multiple groups. A Bonferroni-adjusted level of significance of ˂.025 was used to establish statistical significance for subsequent pairwise comparisons. Statistical analysis was performed by one of the authors (J.W.H).
Results
The various statuses of the spiral artery physiologic transformation in the placental basal plate were defined immunohistochemically as follows: physiologic transformation of the spiral arteries had complete muscular wall replacement by cytokeratin-positive trophoblasts that surrounded the endothelial lining of the vessels (Figure 1, A); spiral arteries with failure of physiologic transformation and atherosis maintained an actin-positive muscular wall, and cytokeratin-positive trophoblastic cells (interstitial extravillous trophoblasts) surrounded but did not invade the arterial walls (Figure 1, B). Spiral arteries with partial failure of physiologic transformation had a combination of smooth muscle cells and intra-arterial trophoblasts in the arterial walls (Figure 1, C). The immunohistochemical characteristics of the different statuses of spiral artery physiologic transformation and atherosis are summarized in Table 1.
Figure 1. Different statuses of spiral artery physiologic transformation and ICAM-1 expression.
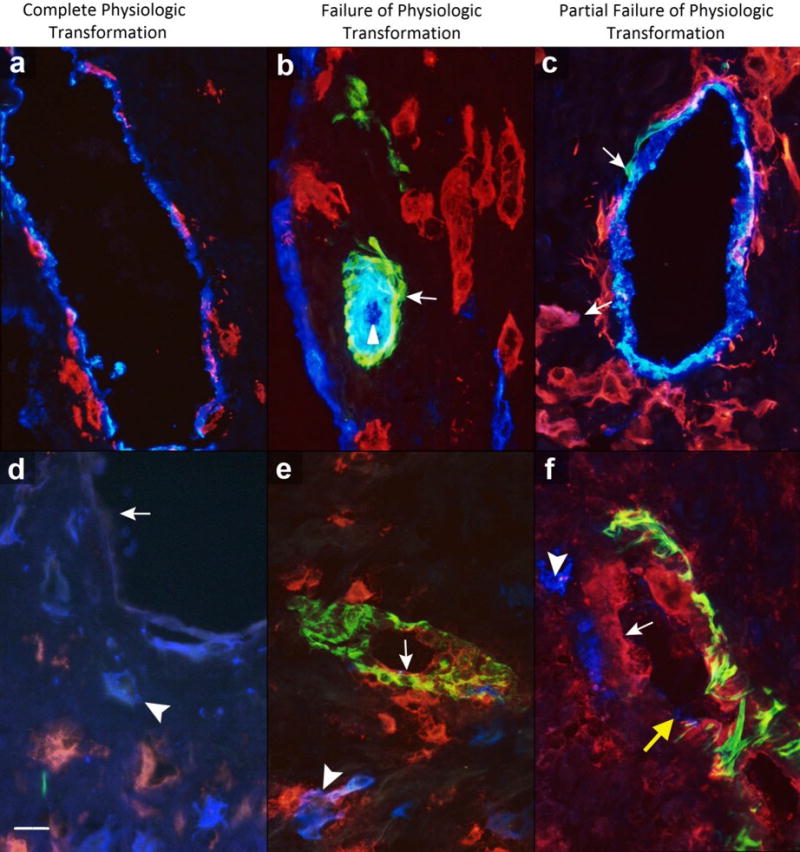
Complete physiologic transformation of spiral arteries: A, presence of endothelium (blue) and trophoblast cells (red) in vessel wall; and D, absence of endothelial intercellular adhesion molecule (ICAM)-1 expression (arrow) with ICAM-1-negative trophoblast cells (blue, arrowhead). Failure of physiologic transformation of spiral arteries: B, characterized by spiral arteries with presence of endothelium (blue, arrowhead) and smooth muscle cells (green, arrow) without arterial invasion of trophoblast cells (red) as seen in artery in center (blue linings on left and lower right of figure corresponding to neighboring veins); and E, presence of endothelial ICAM-1 (red, arrow) with trophoblast cells (blue, arrowhead). Partial physiologic transformation of spiral arteries: C, presence of endothelium (blue), trophoblast cells (red, arrow), and smooth muscle cells (green, arrow) in part of vessel; and F, presence of endothelial ICAM-1 (red, arrow) with trophoblast cells (blue, arrowhead) and smooth muscle cells (green). Note presence of intra-arterial trophoblasts (yellow arrow) in partial physiologic transformation of spiral arteries. Scale represents 50 μm. Original magnification: ×640.
Table 1.
Immunohistochemical markers in spiral artery physiologic transformation and atherosis
| Immunohistochemical characteristics | Complete physiologic transformation | Partial physiologic transformation | Failure of physiologic transformation | Atherosis |
|---|---|---|---|---|
| Cytokeratin | Positive | Positive/negative | Negative | Negative |
| Smooth muscle α-actin | Negative | Positive/negative | Positive | Positive |
| Von Willebrand factor | Positive | Positive | Positive | Positive |
| ICAM-1, endothelial | Negative | Positive/negative | Positive | Positive |
| ICAM-1, interstitial extravillous trophoblast | Negative | Positive/negative | Positive | Positive |
ICAM-1, intercellular adhesion molecule-1.
Of the 123 placentas studied, 108 had spiral arteries with failure of physiologic transformation (Table 2): 79 had failure of physiologic transformation in all spiral arteries examined and 29 had both failure of physiologic transformation and physiologic transformation (Table 2). Fifteen placentas had complete spiral artery physiologic transformation without any failure of physiologic transformation (Table 2). Spiral arteries with partial failure of physiologic transformation (n = 36) had α-actin-positive cells in part of the arterial wall and cytokeratin-positive trophoblastic cells in the rest of the wall (Table 1 and Figure 1, C). Partial failure of physiologic transformation of the spiral arteries in the basal plate was associated with failure of physiologic transformation (n = 22), physiologic transformation (n = 8), or both (n = 6).
Table 2.
Spiral artery physiologic transformation and characteristics of the study population
| Characteristics | Physiologic Transformation w/o Failure of Physiologic Transformation (n=15) | Failure of Physiologic Transformation (n=108)a
|
P-valueb | ||
|---|---|---|---|---|---|
| Failure of Physiologic Transformation w/or w/o Physiologic Transformation (n=108) | Failure of Physiologic Transformation w/Physiologic Transformation (n=29) | Failure of Physiologic Transformation w/o Physiologic Transformation (n=79) | |||
| Gestational age at delivery (weeks)c | 39.5 (26.1, 41.6) | 31.2 (18.5, 41.2) | 31.3 (18.5, 40.4) | 31.2 (19.2, 41.2) | <0.001 |
| Birthweight (percentile)c | 62.5th (7.5th, 99th) | 25.5th (1st, 99th) | 47th (1st, 99th) | 17.5th (1st, 99th) | <0.001 |
| Abnormal pregnancies (n=101)d | 1 (1%) | 100 (99%) | 22 (21.8%) | 78 (77.2%) | <0.001 |
| Preeclampsia (n=27) | 0 (0%) | 27 (100%) | 5 (18.5%) | 22 (81.5%) | |
| Small for gestational age (n=10) | 0 (0%) | 10 (100%) | 1 (10%) | 9 (90%) | |
| Fetal death (n=15) | 0 (0%) | 15 (100%) | 3 (20%) | 12 (80%) | |
| Preterm labor (n=23) | 0 (0%) | 23 (100%) | 7 (30.4%) | 16 (69.6%) | |
| Preterm prelabor rupture of membranes (n=26) | 1 (3.8%) | 25 (96.2%) | 5 (19.2%) | 20 (76.9%) | |
| Normal pregnancies (n=22) | 14 (63.6%) | 8 (36.4%) | 7 (31.8%) | 1 (4.5%) | <0.001 |
| Placentas ICAM-1-positive (n=94)e | 0 (0%) | 94 (100%) | 21 (22.3%) | 73 (77.7%) | <0.001 |
| Abnormal pregnancies (n=92) | 0 (0%) | 92 (100%) | 20 (21.7%) | 72 (78.3%) | |
| Preeclampsia (n=23) | 0 (0%) | 23 (100%) | 5 (21.7%) | 18 (78.3%) | |
| Small for gestational age (n=9) | 0 (0%) | 9 (100%) | 1 (11.1%) | 8 (88.9%) | |
| Fetal death (n=14) | 0 (0%) | 14 (100%) | 3 (21.4%) | 11 (78.6%) | |
| Preterm labor (n=23) | 0 (0%) | 23 (100%) | 7 (30.4%) | 16 (69.6%) | |
| Preterm prelabor rupture of membranes (n=23) | 0 (0%) | 23 (100%) | 4 (17.4%) | 19 (82.6%) | |
| Normal pregnancies (n=2) | 0 (0%) | 2 (100%) | 1 (50%) | 1 (50%) | |
| Placentas with atherosis (n=30) | 0 (0%) | 30 (100%) | 6 (20%) | 24 (80%) | <0.001 |
| Abnormal pregnancies (n=29) | 0 (0%) | 29 (100%) | 6 (20.7%) | 23 (79.3%) | |
| Preeclampsia (n=7) | 0 (0%) | 7 (100%) | 2 (28.6%) | 5 (71.4%) | |
| Small for gestational age (n=3) | 0 (0%) | 3 (100%) | 0 (0%) | 3 (100%) | |
| Fetal death (n=4) | 0 (0%) | 4 (100%) | 0 (0%) | 4 (100%) | |
| Preterm labor (n=6) | 0 (0%) | 6 (100%) | 2 (33.3%) | 4 (66.7%) | |
| Preterm prelabor rupture of membranes (n=9) | 0 (0%) | 9 (100%) | 2 (22.2%) | 7 (77.8%) | |
| Normal pregnancies (n=1) | 0 (0%) | 1 (100%) | 0 (0%) | 1 (100%) | |
Failure of physiologic transformation includes complete and partial failure.
P-values compare the listed characteristic for the group of physiologic transformation (n=15) and the group of failure of physiologic transformation (n=108).
Values are expressed as median (minimum, maximum) according to nationwide United States Natality data [88].
Values in both columns are expressed as number (percentage) of each clinical/immunopathological group.
ICAM-1-positive placentas include endothelial and/or extravillous trophoblast reactivity.
ICAM-1, intercellular adhesion molecule-1.
Of the 108 placentas with failure of physiologic transformation of the spiral arteries in the basal plate, 100 (100/101, 99%) were from women with complicated pregnancies, and 8 (8/22, 36.4%) from normal pregnancies (Table 2). Of the 15 placentas with complete physiologic transformation of the spiral arteries and no failure, however, only 1 (1/101, 1%) was from a woman with an abnormal pregnancy, and 14 (14/22, 63.6%) were from women with normal pregnancies (Table 2). Placentas with only physiologic transformation of the spiral arteries (Table 2) were found mostly in normal pregnancies, and placentas having only failure of physiologic transformation of the spiral arteries (Table 2) were seen predominantly in abnormal pregnancies with preeclampsia, small for gestational age, intrauterine fetal death, preterm labor, and preterm prelabor rupture of membranes (Table 2).
Of placentas from abnormal pregnancies, 99% had spiral arteries with failure of physiologic transformation in the basal plate and only 1% of them had all spiral arteries with physiologic transformation (P < .001) (Table 2). Contrarily, most placentas (63.6%) from normal pregnancies had all spiral arteries with physiologic transformation, and a smaller percentage of them (36.4%) had spiral arteries with failure of physiologic transformation (P <.001). All groups of abnormal pregnancies had a similarly high proportion of placentas with failure of physiologic transformation in all spiral arteries, which was highest in pregnancies with small for gestational age (9/10, 90%), preeclampsia (22/27, 81.5%), and intrauterine fetal death (12/15, 80%), and lowest in preterm labor (16/23, 69.6%) (Table 2). The combination of both failure of physiologic transformation and physiologic transformation of the spiral arteries was more frequently identified in preterm labor. Physiologic transformation of the spiral arteries without any failure of physiologic transformation was found predominantly in normal pregnancies (Table 2). We performed quantitative studies and evaluated the percentage of arteries with different degrees of physiologic transformation in the basal plate of the placentas of the different groups and subgroups studied (Table 3). The percentage of arteries with physiologic transformation and failure of physiologic transformation was significantly different between groups, but no differences were found in the percentage of arteries with partial failure of physiologic transformation (Table 3). The highest percentage of arteries with failure of physiologic transformation was found in placentas from fetal death and preeclampsia; the highest percentage of partial failure of physiologic transformation was found in small-for-gestational-age babies; the highest percentage of arteries with physiologic transformation was found in normal pregnancies and preterm labor; and the lowest percentage of arteries with physiologic transformation was found in small-for-gestational-age placentas (Table 3).
Table 3.
Physiologic transformation, endothelial activation, and atherosis quantification in the study groups
| Characteristics | Abnormal pregnancies (n=101)
|
Normal pregnancies (n=22) | P-valuea | |||||
|---|---|---|---|---|---|---|---|---|
| Total (n=101) | Preeclampsia (n=27) | Small for gestational age (n=10) | Fetal death (n=15) | Preterm labor (n=23) | Preterm prelabor rupture of membranes (n=26) | |||
| Percent arteries with physiologic transformation† | 8.1±18.9 | 7.7±19.4 | 1.4±4.5 | 5.1±11.9 | 11.8±20.4 | 9.6±23.3 | 81.7±29.7 | <0.0001 |
| Percent arteries with partial failure of physiologic transformation | 8.3±15.1 | 5.3±10.5 | 15.5±22.8 | 5.6±10.2 | 10.4±17.2 | 8.5±16 | 12.4±21.8 | 0.30 |
| Percent arteries with failure of physiologic transformation | 83.5±25.7 | 87.0±23 | 83.1±25.9 | 89.3±16.5 | 77.8±33.9 | 81.9±25 | 6.0±17.4 | <0.0001 |
| Percent arteries with endothelial ICAM-1 | 81.8±35.1 | 77.5±38.1 | 88.6±31.4 | 84.4±34.8 | 81.2±34.1 | 82.7±36.4 | 4.5±21.3 | <0.0001 |
| Percent arteries with atherosis | 10.4±21.7 | 10.7±19.9 | 18.3±33.7 | 16.0±30.5 | 4.3±7.7 | 9.2±20.4 | 2.3±10.7 | <0.0001 |
Values are expressed as mean ± SD for each clinical group/subgroup. All percentages were calculated by dividing the number of vessels with a particular characteristic by the total number of vessels × 100.
P-values are for comparison of total abnormal results (n=101) versus normal results (n=22).
ICAM-1, intercellular adhesion molecule-1.
Of the 94 placentas with ICAM-1 expression in arterial endothelium and/or extravillous trophoblasts, all (94/94, 100%) were associated with failure of physiologic transformation (Figure 1, E, and Table 2), and none with physiologic transformation without failure (Figure 1, D, and Table 2) (P <.001). A significant correlation (R2 = 0.84) was found between interstitial extravillous trophoblasts and spiral artery endothelial ICAM-1 expression (P < .001). Spiral arteries with partial failure of physiologic transformation also showed endothelial ICAM-1 expression (Figure 1, F). A high proportion of spiral arteries with endothelial and/or extravillous trophoblasts that were ICAM-1-positive (Table 2) was observed in complicated pregnancies. The combination of both failure of physiologic transformation of the spiral arteries and physiologic transformation was associated with a lower proportion of endothelial and/or extravillous trophoblasts that were ICAM-1-positive (Table 2). The percentage of arteries with endothelial ICAM-1 was significantly different between groups, being higher in small for gestational age and lower in normal pregnancies (Table 3). Interestingly, spiral arteries with physiologic transformation had fewer CD68+ macrophages surrounding extravillous trophoblasts (Figure 2, A) and spiral artery endothelium (Figure 2, C) than arteries with failure of physiologic transformation, which had increased number of CD68+ macrophages around smooth muscle cells of uninvaded arteries (Figure 2, B and D), suggesting that increased inflammation is associated with arterial endothelial and/or extravillous trophoblast activation.
Figure 2. Macrophages and spiral artery physiologic transformation.
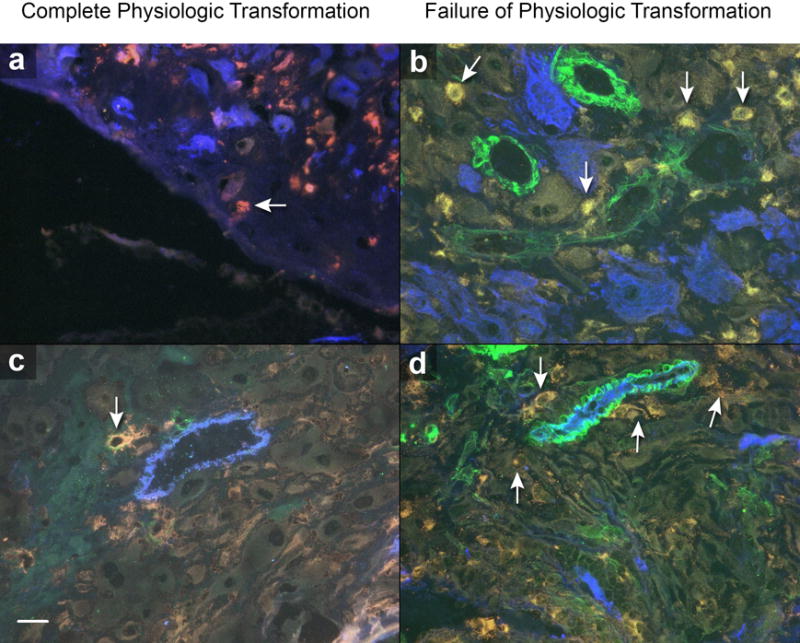
Complete physiologic transformation of spiral arteries: A and C, few CD68+ macrophages (red, arrows) around endovascular trophoblast cells (A, blue) and presence of endothelium (C, blue) in vessels with normal trophoblastic invasion. Failure of physiologic transformation of spiral arteries: B and D, numerous CD68+ macrophages (arrows) around vessels, with presence of endothelium (D, blue), absence of intra-arterial trophoblast cells (B, blue), and presence of smooth muscle cells (B and D, green) in spiral arteries. Scale represents 50 μm. Original magnification: ×640.
Atherosis, akin to placental atherosclerosis (Figure 3A), developed only in placentas with failure of physiologic transformation of the spiral arteries, and all placentas with atherosis had a high proportion of failure of physiologic transformation (Table 2) and activation of interstitial extravillous trophoblasts and spiral artery endothelium. Atherosis was identified in preterm labor (n = 6), preterm prelabor rupture of membranes (n = 9), preeclampsia (n = 7), small for gestational age (n = 3), intrauterine fetal death (n = 4), and normal (n = 1) placentas (Table 2). The percentage of arteries with atherosis lesions was significantly different between groups (P < .001); the highest percentage was found in small for gestational age and fetal death, and the lowest percentage in normal pregnancies and preterm labor (Table 3). Arteries with atherosis had numerous oil red O-positive cells, which were not observed in arteries with either physiologic transformation or failure of physiologic transformation without atherosis (Figure 3, B to D). Oil red O-positive cells were confirmed to be macrophages (Figure 3, G). Spiral arteries with atherosis were α-actin-positive (Figure 3, E and H) and cytokeratin-negative (Figure 3, F), as defined in Table 1. Lesions of atherosis were observed in 30% (30/100) of placentas with failure of physiologic transformation of the spiral arteries (Table 2) and, as expected, in none of the placentas with physiologic transformation (P = .001). Furthermore, all placentas with atherosis had concomitant spiral artery endothelial and interstitial extravillous trophoblast activation. Arteries with failure of physiologic transformation that were ICAM-1-positive had a higher frequency of atherosis than placentas without endothelial activation (P < .001). Atherosis was found in 31.9% (30/94) of placentas positive for ICAM-1 on spiral artery endothelium and/or interstitial extravillous trophoblasts; in none of the 13 placentas with failure of physiologic transformation that were ICAM-1-negative; and as expected, in none of the 15 placentas with complete physiologic transformation that were ICAM-1-negative (P = .04).
Figure 3. Histochemical and immunohistochemical characteristics of atherosis.
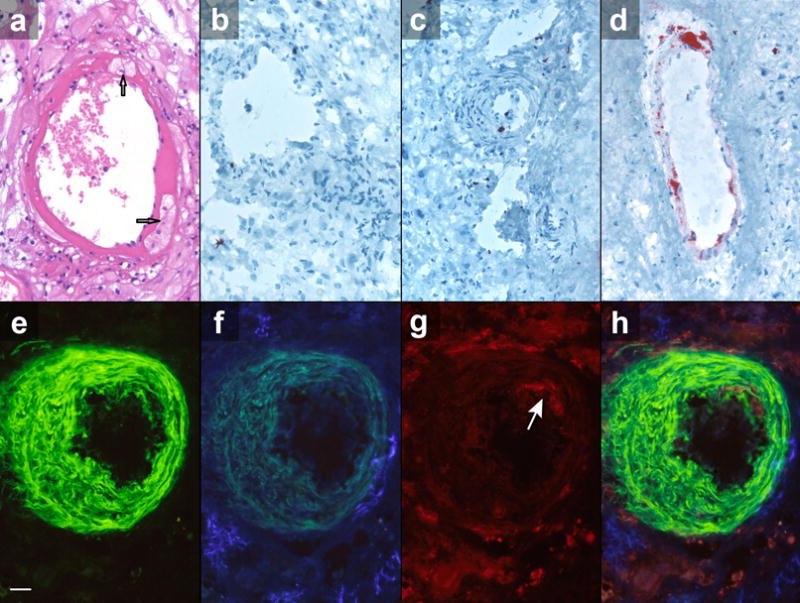
Atherosis lesion recognized with hematoxylin-eosin stain: A, note presence of foam cells (arrows) reactive with oil red O, as shown in D, confirming presence of lipids, which are B, not observed in vessels with normal trophoblastic invasion or C, absence of trophoblastic invasion also reacted with oil red O. Atherosis was observed in spiral arteries using triple-antibody technique that showed arterial smooth muscle cells (E, green), trophoblast cells surrounding but not replacing vessel walls (F, blue), and presence of macrophages in vessel’s intima (G, red, arrow). H, These cell subtypes are clearly shown in combined photograph. Triple-antibody immunohistochemistry for smooth muscle (alpha-actin, E), trophoblasts (cytokeratin, F), macrophages (CD68, G), and their combination (H). Scale represents 50 mm. Original magnification: ×640. A, Hematoxylin-eosin and B to D, oil red O stains.
We subsequently performed a comparison among the immunohistochemically defined spiral artery phenotypes in the basal plate and the different groups of pregnancies studied, the status of spiral artery transformation, and the presence of arterial endothelial/trophoblast activation and atherosis (Table 4). As expected, abnormal pregnancies had a significantly increased proportion of spiral arteries actin-positive and cytokeratin-negative; arteries actin-positive and cytokeratin-negative were characteristic of failure of physiologic transformation, and showed increased ICAM-1 expression and atherosis (Table 4). The clustered bar chart (Figure 4) shows a summary of the immunohistochemical characteristics of the spiral arteries in the basal plate of the different pregnancy conditions studied, i.e., normal and abnormal pregnancies, physiologic transformation, partial failure of physiologic transformation, failure of physiologic transformation, placental ICAM-1 expression, and atherosis.
Table 4.
Immunohistochemistry of spiral artery physiologic transformation in the different study groups
| Characteristics | Spiral artery immunohistochemistry (np= 123)a
|
P-valueb | ||
|---|---|---|---|---|
| Spiral arteries cytokeratin-positive and actin-negative (na=43) | Spiral arteries cytokeratin-positive and actin-positive (na=36) | Spiral arteries actin-positive and cytokeratin-negative (na=100) | ||
| Abnormal pregnancies (np=101)a | 22 (21.8%) | 29 (28.7%) | 97 (96.0%) | <0.001 |
| Preeclampsia (np= 27) | 5 (18.5%) | 7 (25.9%) | 27 (100%) | |
| Small for gestational age (np= 10) | 1 (10%) | 4 (40%) | 10 (100%) | |
| Fetal death (np= 15) | 3 (20%) | 4 (26.7%) | 15 (100%) | |
| Preterm labor (np= 23) | 7 (30.4%) | 7 (30.4%) | 20 (87%) | |
| Preterm prelabor rupture of membranes (np= 26) | 6 (23.1%) | 7 (26.9%) | 25 (96.2%) | |
| Normal pregnancies (np=22) | 21 (95.5%) | 7 (31.8) | 3 (13.6%) | <0.001 |
| Physiologic transformation w/o failure of physiologic transformation (np=15)c | 15 (100%) | 0 (0%) | 0 (0%) | |
| Failure of physiologic transformation w/or w/o physiologic transformation (np=108) | 28 (25.9%) | 28 (25.9%) | 98 (90.7%) | |
| Failure of physiologic transformation w/physiologic transformation (np=28) | 28 (100%) | 14 (50%) | 19 (67.9%) | |
| Failure of physiologic transformation w/o physiologic transformation (np=80) | 0 (0%) | 24 (30%) | 77 (96.3%) | |
| Placentas ICAM-1-positive (np=94)d | 21 (22.3%) | 28 (29.8%) | 88 (93.6%) | <0.001 |
| Placentas with atherosis (np=30)e | 4 (13.3%) | 10 (33.3%) | 30 (100%) | <0.001 |
Data were obtained from na=179 arteries from np=123 placentas.
P-values compare the listed characteristic for the groups with spiral arteries that are cytokeratin-positive and actin negative (na =43), cytokeratin-positive and actin-positive (na =36), and cytokeratin-negative and actin-positive (na =100).
Failure of physiologic transformation considers complete and partial failure.
Placentas ICAM-1-positive include spiral artery endothelial and/or extravillous trophoblast reactivity.
Atherosis was identified in preterm labor (np =6), preterm prelabor rupture of membranes (np =9), preeclampsia (np =7), small for gestational age (np =3), fetal death (np =4), and normal pregnancies (np =1).
ICAM-1: Intercellular adhesion molecule-1.
Figure 4. Summary of immunohistochemical characteristics of spiral artery physiologic transformation in basal plate of different pregnancy conditions studied.
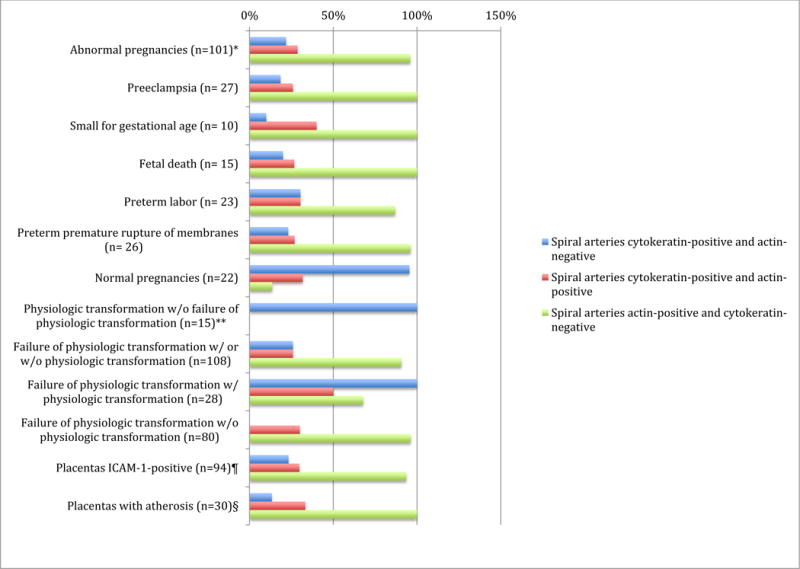
Clustered bar chart showing summary of immunohistochemical characteristics of spiral arteries in basal plate of different pregnancy conditions studied.
*Data were obtained from na = 179 arteries from np = 123 placentas;
**Failure of physiologic transformation considers complete and partial failure;
¶Placentas intercellular adhesion molecule (ICAM)-1-positive include spiral artery endothelial and/or extravillous trophoblast reactivity;
§Atherosis was identified in preterm labor (np = 6), preterm premature rupture of membranes (np = 9), preeclampsia (np = 7), small for gestational age (np = 3), fetal death (np = 4), and normal pregnancies (np = 1).
na, number of arteries; np, number of placentas.
Comment
Principal findings of the study
First, failure of physiologic transformation of the spiral arteries in the placental basal plate had ICAM-1-positive interstitial extravillous trophoblasts. Second, placentas with failure of physiologic transformation of the spiral arteries and extravillous trophoblast activation had concomitant spiral artery ICAM-1-positive endothelium. Third, failure of physiologic transformation of the spiral arteries that were ICAM-1-positive had a higher frequency of atherosis than placentas without endothelial activation.
Activated extravillous trophoblasts found around spiral arteries with failure of physiologic transformation but not arteries with physiologic transformation suggests that the presence of ICAM-1-positive interstitial extravillous trophoblasts prevents the normal invasion of spiral arteries by those cells, possibly by enabling maternal immune cells to react against activated extravillous trophoblasts. Impaired trophoblast invasion has been associated with an increased number of macrophages around spiral arteries in preeclampsia [92] that could lead to increased trophoblast apoptosis [93] and impaired arterial invasion.
Placentas with failure of physiologic transformation of the spiral arteries and endothelial activation had a significantly (P < .001) higher frequency of atherosclerotic-like lesions of atherosis than placentas without activation, suggesting that endothelial activation is paramount in the development of the lesion. Multiple pathologic processes involving excessive decidual inflammation have been implicated in the genesis of atherosis [94–97]. Local factors may contribute to lack of trophoblast invasion and lipid deposition in spiral arteries [96]. Trophoblast migration may be suppressed by an abnormal immune interaction between maternal and fetal cells [96]. It is noteworthy that atherosis and vascular lesions in rejected kidney and heart transplants are histologically similar, suggesting that atherosis may be the result of immune dysregulation at the maternal-fetal interface [43]. In the decidua, invading trophoblasts can act as ligands for maternal uterine immune cells since trophoblasts bear paternal (foreign) HLA-C antigens that can be detected by receptors on those cells [98–101]. Under normal circumstances, immune cells do not initiate a destructive response toward trophoblasts, but this response could be triggered in complicated pregnancies and explain impaired trophoblast invasion [100,102]. We have previously proposed [43] that atherosis could have an immune origin. This hypothesis has gained support, as recently reviewed by Staff et al [96]. An immunological mismatch between mother and fetus can lead to dysregulation of decidual immunity, promote local decidual inflammation, and predispose to atherosis.
Uteroplacental ischemia can lead to the production of anti-angiogenic factors [103–113], such as sFlt-1 (soluble fms-like tyrosine kinase-1) [114–117] and endoglin [69,118], which can induce endothelial dysfunction and predispose to preeclampsia or an anti-angiogenic state [57,119–129]. In addition, an excess production of trophoblast debris can result in exaggerated intravascular inflammation and endothelial cell dysfunction [130–134]. Preeclampsia is characterized by intravascular inflammation [59–62,135] and, similar to atherosclerosis [46,136,137], is marked by increased levels of circulating pro-inflammatory cytokines (such as interleukin-6 and tumor necrosis factor-α [138–143]), C-reactive protein [141,142,144–146], and markers of leukocyte activation [144,147]. Interestingly, pregnancy may constitute a metabolic/vascular “stress test,” uncovering a subclinical endothelial cell dysfunction that manifests later in life, in mothers affected with preeclampsia or an anti-angiogenic state [148–151].
Several adverse pregnancy outcomes, such as preeclampsia [1,32–34], have characteristic disordered deep placentation [28–42]. Our findings provide an association between failure of physiologic transformation of the spiral arteries in the placental basal plate and activation of spiral artery endothelial cells and extravillous trophoblasts. By secreting both tumor necrosis factor-α and a tryptophan-depleting enzyme, excess macrophages in the placental bed of women with preeclampsia have been implicated in defective extravillous trophoblast spiral artery invasion [152]. More recently, tumor necrosis factor-α has been found to mediate fetal growth restriction during abnormal maternal inflammation [58]. Macrophages inhibit extravillous trophoblast invasion both passively, by being attracted by extravillous trophoblasts, and actively, by inducing peri-arterial extravillous trophoblast apoptosis [152]. Arterial ICAM-1 up-regulation could facilitate macrophage recruitment as shown in placental villitis of unknown etiology and massive chronic intervillositis [153,154]. Aberrant ICAM-1 expression has been described in cultured syncytiotrophoblasts pre-treated with both inflammatory cytokines, which also facilitate adhesion of monocytes to the syncytiotrophoblast [155]. Monocytes adhering to placental syncytiotrophoblasts can induce tumor necrosis factor-α-dependent apoptosis accompanied by focal trophoblast disruption [156]. Focal damage of the placenta could be a route for maternal leukocyte infiltration into the villi and a possible mechanism of villitis [156]. From our findings, we postulate that activated extravillous trophoblasts could facilitate trophoblast damage, inflammation, and subsequent inhibition of spiral artery transformation. Failure of physiologic transformation of the spiral arteries could result from ICAM-1-reactive spiral artery endothelial cells impeding the normal invasion [157], or ICAM-1-enhanced reactivity could be a consequence of failure of the physiologic transformation.
The rates of failure of physiologic transformation in our study are higher than those reported previously in women with different placental disorders [20,21,36–38,158]. The differences could be explained by the number of blocks obtained from the basal plate’s placental septa [21] included in our study (a mean of 3.5 blocks per placenta) and the use of triple-antibody immunohistochemistry. The use of a triple-antibody immunohistochemical technique allows the simultaneous identification of three cell types—smooth muscle cells, trophoblasts, and endothelium—which facilitates identification not only of the type of vessel but its predominant cell type phenotypic characteristics. The immunohistochemistry technique using cytokeratin, actin, and von Willebrand factor as markers of trophoblasts, smooth muscle cells, and endothelium, respectively, allows for a more precise identification of the degree of transformation, or the lack thereof, in each vessel. Interestingly, our data in the placental basal plate are supported by recent findings of a significantly higher frequency of acute atherosis lesions in the placental basal plate than in placental bed biopsies (both decidua and myometrial segments), considering that atherosis lesions only develop in spiral arteries with failure of trophoblastic invasion [159]. As we previously discussed [36], the reasons why some women with failure of physiologic transformation of the spiral arteries develop preeclampsia, whereas others develop preterm labor, are unknown, and the possibility still exists that women with preterm labor would have developed preeclampsia if the pregnancy had continued longer. We previously proposed that pathology of the placental bed, primarily through ischemia and/or other mechanisms such as a maternal-fetal immune response, may lead to preeclampsia, small for gestational age, preterm labor with intact or ruptured membranes, abruptio placentae, and fetal death; and why a similar insult would result in different clinical phenotypes depends on genetic and environmental factors and time of onset, duration, and extent of the ischemic insult [160]. Korzeniewski et al [161] recently demonstrated that maternal plasma angiogenic index-1 concentration ratios seem to reflect the burden of uteroplacental vasculopathy indicated by histologic examination of the placenta, regardless of clinical diagnosis. The time order between changes in plasma angiogenic index-1 concentration ratios and placental features consistent with maternal vasculopathy, however, remains to be determined. The identification of the highest proportion of arteries with complete physiologic transformation in women with preterm labor with intact membranes and preterm prelabor rupture of membranes and the lowest proportion of transformed spiral arteries in small for gestational age in our study, suggests that the difference between differing pathologies is centered in the varying proportion of spiral arteries with physiologic transformation and failure of physiologic transformation. Examining more samples or perhaps the whole placental basal plate would help clarify quantitative differences of vessel transformation in different pregnancy disorders that could explain the differing outcomes. Indeed, the presence of failure of physiologic transformation in the placental basal plate in our study was associated with a significantly lower gestational age, suggesting that abnormal placentation and the vasculopathies associated with it could predispose to an early delivery. The identification of failure of spiral artery physiologic transformation in multiple pregnancy complications in general and not only in malperfusion pathologies of the placenta in particular, suggests that inflammation leading to cell activation may be the trigger for an abnormal placentation that expresses as different pathologies of pregnancy. Finally, the frequency of atherosis in our study was higher than the frequency recently described in a large population of placentas from normal and abnormal pregnancies [159,162]. We believe this is due to the high sensitivity of triple-antibody immunohistochemistry when compared with hematoxylin-eosin used to detect failure of physiologic transformation of the spiral arteries and placental atherosis [44].
In support of the central role played by trophoblast invasion, Kam et al [163] demonstrated that the physiological change in spiral arteries during pregnancy can only take place when interstitial trophoblasts are present. In further studies [36,38], frequent transformational failure was seen in the myometrial but not decidual spiral artery segments in preterm prelabor rupture of membranes, and increased transformational failure was seen in myometrial and decidual segments in preterm labor with intact membranes. The frequency of failure was higher in preeclampsia than in either preterm prelabor rupture of membranes or preterm labor with intact membranes. In this study, failure of physiologic transformation of the spiral arteries was highest in pregnancies with preeclampsia and lowest in preterm labor. The differences seen between the physiological responses of normal pregnancy and placental disorders may be only quantitative and not qualitative. Since the fetoplacental unit is a semi-allograft, normal pregnancy can be considered a challenge to the maternal immune system [164–169], and indeed, is a state of physiologic intravascular inflammation. We and other investigators have reported excessive intravascular inflammation in preeclampsia [59–62], intrauterine growth restriction [170,171], preterm labor [172], preterm prelabor rupture of membranes [173], and acute pyelonephritis [174]. An exaggerated intravascular inflammatory process can impair remodeling of the spiral arteries and lead to endothelial cell activation and atherosis.
A further point that needs to be emphasized is the role of endothelial disruption in physiologic transformation of the spiral arteries and atherosis in complicated pregnancies. Interestingly, atherosis lesions can show endothelial disruption [175], and it is tempting to postulate that such disruption is no more than the development of atherosis in arteries with partial failure of physiologic transformation. Although the study did not examine smooth muscle cells, partial failure of physiologic transformation of the spiral arteries was shown immunohistochemically by complementarity staining for endothelium and endovascular trophoblast in vessels of the placental bed [176]. Our findings demonstrated that both failure of physiologic transformation and partial failure of physiologic transformation of the spiral arteries showed endothelial activation, and interestingly, partial failure of physiologic transformation had intravascular trophoblasts (Figure 1), as found in arteries with endothelial disruption. Areas with intravascular trophoblast and absence of endothelium in partial failure of physiologic transformation of the spiral arteries would not show any expression of endothelial activation markers, as found in our study.
The nomenclature used to describe disorders of vascular remodeling in the spiral arteries can be confusing and, at times, misleading. Craven et al [10] viewed vascular changes as the beginning of physiologic change that occurs without trophoblast invasion, but others [163] adopted a more narrow view of physiological change—one that not only includes trophoblast invasion but is defined by it. Pijnenborg et al [19] suggested a new nomenclature replacing the term altogether with “trophoblast-associated remodeling.” We suggest the term “defective superficial placentation” to describe the failure of physiologic transformation of the spiral arteries in the placental basal plate during pregnancy. Although examination of the basal plate exclusively without availability of placental bed biopsies does not allow identification of defective deep placentation, the diagnosis of defective superficial placentation is also abnormal, because it suggests severe failure of physiologic transformation of the spiral arteries.
Although it is difficult to be certain regarding initial events during the placental development by examining third-trimester placentas, it is tempting to suggest what happens during early placentation based on our findings. The sequence of events is that trophoblasts first invade the superficial (or decidual) segment of the spiral arteries and, only later, the myometrial segment of the arteries. While the hallmark for disorders of deep placentation is failure of trophoblast invasion in the myometrial segment (which requires a placental bed biopsy), the fact that trophoblast has not invaded the decidual segment indicates that the process is more severe. This interpretation is indicative of failure of the first wave of spiral artery trophoblastic invasion [34]. Furthermore, the approach that we have proposed (examination of the basal plate of the placenta) has practical value, given that placental bed biopsies are rarely performed, and often require a cesarean delivery. In contrast, examination of the basal plate of the placenta can be undertaken in all placentas, whether the mode of delivery is vaginal or cesarean [34]. The concept of defective superficial placentation may be used to improve the characterization of different disorders of the placental bed. It is possible that our data will be valuable in refining the existing tools for the evaluation of risk before pregnancy or in early gestation for the prediction of subsequent pregnancy outcome.
Acknowledgments
We thank the Methodist Research Institute and the Department of Obstetrics and Gynecology at Hutzel Women’s Hospital in Detroit, Michigan.
Funding: This research was supported, in part, by the Perinatology Research Branch, Division of Intramural Research, Eunice Kennedy Shriver National Institute of Child Health and Human Development, National Institutes of Health, U.S. Department of Health and Human Services); and, in part, with Federal funds from NICHD/NIH/DHHS under Contract No. HHSN275201300006C. Dr. Romero has contributed to this work as part of his official duties as an employee of the United States Federal Government.
Footnotes
Disclosure: The authors report no conflicts of interest.
References
- 1.Brosens I, Robertson WB, Dixon HG. The physiological response of the vessels of the placental bed to normal pregnancy. J Pathol Bacteriol. 1967;93:569–579. doi: 10.1002/path.1700930218. [DOI] [PubMed] [Google Scholar]
- 2.De Wolf F, Robertson WB, Brosens I. The ultrastructure of acute atherosis in hypertensive pregnancy. Am J Obstet Gynecol. 1975;123:164–174. doi: 10.1016/0002-9378(75)90522-0. [DOI] [PubMed] [Google Scholar]
- 3.Brosens I, Khong TY. Defective spiral artery remodeling. In: Pijnenborg R, Brosens I, Romero R, editors. Placental bed disorders. 1st. New York NY: Cambridge University Press; 2010. pp. 11–21. [Google Scholar]
- 4.Whitley GS, Cartwright JE. Cellular and molecular regulation of spiral artery remodelling: lessons from the cardiovascular field. Placenta. 2010;31:465–474. doi: 10.1016/j.placenta.2010.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Osol G, Mandala M. Maternal uterine vascular remodeling during pregnancy. Physiology (Bethesda) 2009;24:58–71. doi: 10.1152/physiol.00033.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Osol G, Moore LG. Maternal uterine vascular remodeling during pregnancy. Microcirculation. 2014;21:38–47. doi: 10.1111/micc.12080. [DOI] [PubMed] [Google Scholar]
- 7.Burton GJ, Woods AW, Jauniaux E, Kingdom JCP. Rheological and physiological consequences of conversion of the maternal spiral arteries for uteroplacental blood flow during human pregnancy. Placenta. 2009;30:473–82. doi: 10.1016/j.placenta.2009.02.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Robertson WB, Brosens I, Dixon G. Uteroplacental vascular pathology. Eur J Obstet Gynecol Reprod Biol. 1975;5:47–65. doi: 10.1016/0028-2243(75)90130-6. [DOI] [PubMed] [Google Scholar]
- 9.Pijnenborg R, Bland JM, Robertson WB, et al. Uteroplacental arterial changes related to interstitial trophoblast migration in early human pregnancy. Placenta. 1983;4:397–413. doi: 10.1016/s0143-4004(83)80043-5. [DOI] [PubMed] [Google Scholar]
- 10.Craven CM, Morgan T, Ward K. Decidual spiral artery remodelling begins before cellular interaction with cytotrophoblasts. Placenta. 1998;19:241–252. doi: 10.1016/s0143-4004(98)90055-8. [DOI] [PubMed] [Google Scholar]
- 11.Staff AC, Dechend R, Pijnenborg R. Learning from the placenta: acute atherosis and vascular remodeling in preeclampsia-novel aspects for atherosclerosis and future cardiovascular health. Hypertension. 2010;56:1026–1034. doi: 10.1161/HYPERTENSIONAHA.110.157743. [DOI] [PubMed] [Google Scholar]
- 12.Smith SD, Dunk CE, Aplin JD, et al. Evidence for immune cell involvement in decidual spiral arteriole remodeling in early human pregnancy. Am J Pathol. 2009;174:1959–1971. doi: 10.2353/ajpath.2009.080995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Harris LK. Review: Trophoblast-vascular cell interactions in early pregnancy: how to remodel a vessel. Placenta. 2010;31(Suppl):S93–98. doi: 10.1016/j.placenta.2009.12.012. [DOI] [PubMed] [Google Scholar]
- 14.Hazan AD, Smith SD, Jones RL, et al. Vascular-leukocyte interactions: mechanisms of human decidual spiral artery remodeling in vitro. Am J Pathol. 2010;177:1017–1030. doi: 10.2353/ajpath.2010.091105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lash GE, Otun HA, Innes BA, et al. Regulation of extravillous trophoblast invasion by uterine natural killer cells is dependent on gestational age. Hum Reprod. 2010;25:1137–1145. doi: 10.1093/humrep/deq050. [DOI] [PubMed] [Google Scholar]
- 16.Harris LK. IFPA Gabor Than Award lecture: Transformation of the spiral arteries in human pregnancy: key events in the remodelling timeline. Placenta. 2011;32(Suppl 2):S154–158. doi: 10.1016/j.placenta.2010.11.018. [DOI] [PubMed] [Google Scholar]
- 17.Robson A, Harris LK, Innes BA, et al. Uterine natural killer cells initiate spiral artery remodeling in human pregnancy. FASEB J. 2012;26:4876–4885. doi: 10.1096/fj.12-210310. [DOI] [PubMed] [Google Scholar]
- 18.Tessier DR, Yockell-Lelievre J, Gruslin A. Uterine Spiral Artery Remodeling: The Role of Uterine Natural Killer Cells and Extravillous Trophoblasts in Normal and High-Risk Human Pregnancies. Am J Reprod Immunol. 2015;74:1–11. doi: 10.1111/aji.12345. [DOI] [PubMed] [Google Scholar]
- 19.Pijnenborg R, Vercruysse L, Hanssens M. The uterine spiral arteries in human pregnancy: facts and controversies. Placenta. 2006;27:939–958. doi: 10.1016/j.placenta.2005.12.006. [DOI] [PubMed] [Google Scholar]
- 20.Espinoza J, Romero R, Mee Kim Y, et al. Normal and abnormal transformation of the spiral arteries during pregnancy. J Perinat Med. 2006;34:447–458. doi: 10.1515/JPM.2006.089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Brosens I, Pijnenborg R, Vercruysse L, et al. The “Great Obstetrical Syndromes” are associated with disorders of deep placentation. Am J Obstet Gynecol. 2011;204:193–201. doi: 10.1016/j.ajog.2010.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Brosens I. the placental bed [thesis] London: University of London; 1965. [Google Scholar]
- 23.Brosens I, Dixon HG. The anatomy of the maternal side of the placenta. J Obstet Gynaecol Br Commonw. 1966;73:357–363. doi: 10.1111/j.1471-0528.1966.tb05175.x. [DOI] [PubMed] [Google Scholar]
- 24.Brosens I, Derwig I, Brosens J, et al. The enigmatic uterine junctional zone: the missing link between reproductive disorders and major obstetrical disorders? Hum Reprod. 2010;25:569–574. doi: 10.1093/humrep/dep474. [DOI] [PubMed] [Google Scholar]
- 25.Khong Y, Brosens I. Defective deep placentation. Best Pract Res Clin Obstet Gynaecol. 2011;25:301–311. doi: 10.1016/j.bpobgyn.2010.10.012. [DOI] [PubMed] [Google Scholar]
- 26.Pijnenborg R, Vercruysse L, Brosens I. Deep placentation. Best Pract Res Clin Obstet Gynaecol. 2011;25:273–285. doi: 10.1016/j.bpobgyn.2010.10.009. [DOI] [PubMed] [Google Scholar]
- 27.Brosens I, Benagiano G, Brosens JJ. The potential perinatal origin of placentation disorders in the young primigravida. Am J Obstet Gynecol. 2015;212:580–585. doi: 10.1016/j.ajog.2015.01.013. [DOI] [PubMed] [Google Scholar]
- 28.Khong TY, Liddell HS, Robertson WB. Defective haemochorial placentation as a cause of miscarriage: a preliminary study. Br J Obstet Gynaecol. 1987;94:649–655. doi: 10.1111/j.1471-0528.1987.tb03169.x. [DOI] [PubMed] [Google Scholar]
- 29.Michel MZ, Khong TY, Clark DA, et al. A morphological and immunological study of human placental bed biopsies in miscarriage. Br J Obstet Gynaecol. 1990;97:984–988. doi: 10.1111/j.1471-0528.1990.tb02468.x. [DOI] [PubMed] [Google Scholar]
- 30.Ball E, Bulmer JN, Ayis S, et al. Late sporadic miscarriage is associated with abnormalities in spiral artery transformation and trophoblast invasion. J Pathol. 2006;208:535–542. doi: 10.1002/path.1927. [DOI] [PubMed] [Google Scholar]
- 31.Dommisse J, Tiltman AJ. Placental bed biopsies in placental abruption. Br J Obstet Gynaecol. 1992;99:651–654. doi: 10.1111/j.1471-0528.1992.tb13848.x. [DOI] [PubMed] [Google Scholar]
- 32.Brosens IA, Robertson WB, Dixon HG. The role of the spiral arteries in the pathogenesis of preeclampsia. Obstet Gynecol Annu. 1972;1:177–191. [PubMed] [Google Scholar]
- 33.Labarrere C, Althabe O. Chronic villitis of unknown etiology and maternal arterial lesions in preeclamptic pregnancies. Eur J Obstet Gynecol Reprod Biol. 1985;20:1–11. doi: 10.1016/0028-2243(85)90077-2. [DOI] [PubMed] [Google Scholar]
- 34.Khong TY, De Wolf F, Robertson WB, et al. Inadequate maternal vascular response to placentation in pregnancies complicated by pre-eclampsia and by small-for-gestational age infants. Br J Obstet Gynaecol. 1986;93:1049–1059. doi: 10.1111/j.1471-0528.1986.tb07830.x. [DOI] [PubMed] [Google Scholar]
- 35.Althabe O, Labarrere C, Telenta M. Maternal vascular lesions in placentae of small-for-gestational-age infants. Placenta. 1985;6:265–276. doi: 10.1016/s0143-4004(85)80056-4. [DOI] [PubMed] [Google Scholar]
- 36.Kim YM, Bujold E, Chaiworapongsa T, et al. Failure of physiologic transformation of the spiral arteries in patients with preterm labor and intact membranes. Am J Obstet Gynecol. 2003;189:1063–1069. doi: 10.1067/s0002-9378(03)00838-x. [DOI] [PubMed] [Google Scholar]
- 37.Arias F, Rodriquez L, Rayne SC, et al. Maternal placental vasculopathy and infection: two distinct subgroups among patients with preterm labor and preterm ruptured membranes. Am J Obstet Gynecol. 1993;168:585–591. doi: 10.1016/0002-9378(93)90499-9. [DOI] [PubMed] [Google Scholar]
- 38.Kim YM, Chaiworapongsa T, Gomez R, et al. Failure of physiologic transformation of the spiral arteries in the placental bed in preterm premature rupture of membranes. Am J Obstet Gynecol. 2002;187:1137–1142. doi: 10.1067/mob.2002.127720. [DOI] [PubMed] [Google Scholar]
- 39.Labarrere CA, Catoggio LJ, Mullen EG, et al. Placental lesions in maternal autoimmune diseases. Am J Reprod Immunol Microbiol. 1986;12:78–86. doi: 10.1111/j.1600-0897.1986.tb00068.x. [DOI] [PubMed] [Google Scholar]
- 40.Lyall F, Robson SC, Bulmer JN. Spiral artery remodeling and trophoblast invasion in preeclampsia and fetal growth restriction. Relationship to clinical outcome Hypertension. 2013;62:1046–54. doi: 10.1161/HYPERTENSIONAHA.113.01892. [DOI] [PubMed] [Google Scholar]
- 41.Labarrere C, Althabe O. Chronic villitis of unknown etiology and decidual maternal vasculopathies in sustained chronic hypertension. Eur J Obstet Gynecol Reprod Biol. 1986;21:27–32. doi: 10.1016/0028-2243(86)90042-0. [DOI] [PubMed] [Google Scholar]
- 42.Labarrere CA, Althabe OH. Inadequate maternal vascular response to placentation in pregnancies complicated by preeclampsia and small-for-gestational-age infants. Br J Obstet Gynaecol. 1987;94:1113–8. doi: 10.1111/j.1471-0528.1987.tb02302.x. [DOI] [PubMed] [Google Scholar]
- 43.Labarrere CA. Acute atherosis. A histopathological hallmark of immune aggression? Placenta. 1988;9:95–108. doi: 10.1016/0143-4004(88)90076-8. [DOI] [PubMed] [Google Scholar]
- 44.Labarrere CA, Faulk WP. Antigenic identification of cells in spiral artery trophoblastic invasion: validation of histologic studies by triple-antibody immunocytochemistry. Am J Obstet Gynecol. 1994;171:165–71. doi: 10.1016/0002-9378(94)90464-2. [DOI] [PubMed] [Google Scholar]
- 45.Avagliano L, Bulfamante GP, Morabito A, Marconi AM. Abnormal spiral artery remodeling in the decidual segment during pregnancy: from histology to clinical correlation. J Clin Pathol. 2011;64:1064–8. doi: 10.1136/jclinpath-2011-200092. [DOI] [PubMed] [Google Scholar]
- 46.Libby P, Ridker PM, Maseri A. Inflammation and atherosclerosis. Circulation. 2002;105:1135–1143. doi: 10.1161/hc0902.104353. [DOI] [PubMed] [Google Scholar]
- 47.Libby P. Inflammation in atherosclerosis. Nature. 2002;420:868–874. doi: 10.1038/nature01323. [DOI] [PubMed] [Google Scholar]
- 48.Young JL, Libby P, Schonbeck U. Cytokines in the pathogenesis of atherosclerosis. Thromb Haemost. 2002;88:554–567. [PubMed] [Google Scholar]
- 49.Hansson GK, Libby P. The immune response in atherosclerosis: a double-edged sword. Nat Rev Immunol. 2006;6:508–519. doi: 10.1038/nri1882. [DOI] [PubMed] [Google Scholar]
- 50.Libby P. Inflammatory mechanisms: the molecular basis of inflammation and disease. Nutr Rev. 2007;65:S140–146. doi: 10.1111/j.1753-4887.2007.tb00352.x. [DOI] [PubMed] [Google Scholar]
- 51.Rocha VZ, Libby P. Obesity, inflammation, and atherosclerosis. Nat Rev Cardiol. 2009;6:399–409. doi: 10.1038/nrcardio.2009.55. [DOI] [PubMed] [Google Scholar]
- 52.Galkina E, Ley K. Immune and inflammatory mechanisms of atherosclerosis (*) Annu Rev Immunol. 2009;27:165–197. doi: 10.1146/annurev.immunol.021908.132620. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Libby P, Okamoto Y, Rocha VZ, et al. Inflammation in atherosclerosis: transition from theory to practice. Circ J. 2010;74:213–220. doi: 10.1253/circj.cj-09-0706. [DOI] [PubMed] [Google Scholar]
- 54.Brosens I, Renaer M. On the pathogenesis of placental infarcts in pre-eclampsia. J Obstet Gynaecol Br Commonw. 1972;79:794–799. doi: 10.1111/j.1471-0528.1972.tb12922.x. [DOI] [PubMed] [Google Scholar]
- 55.Brosens IA. Morphological changes in the utero-placental bed in pregnancy hypertension. Clin Obstet Gynaecol. 1977;4:573–593. [PubMed] [Google Scholar]
- 56.Kaufmann P, Black S, Huppertz B. Endovascular trophoblast invasion: implications for the pathogenesis of intrauterine growth retardation and preeclampsia. Biol Reprod. 2003;69:1–7. doi: 10.1095/biolreprod.102.014977. [DOI] [PubMed] [Google Scholar]
- 57.Chaiworapongsa T, Chaemsaithong P, Yeo L, et al. Part1: Current Understanding of its Pathophysiology. Nature Review Nephrology. 2014;10:466–480. doi: 10.1038/nrneph.2014.102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Cotechini T, Komisarenko M, Sperou A, et al. Inflammation in rat pregnancy inhibits spiral artery remodeling leading to fetal growth restriction and features of preeclampsia. J Exp Med. 2014;211:165–179. doi: 10.1084/jem.20130295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Sacks GP, Studena K, Sargent K, et al. Normal pregnancy and preeclampsia both produce inflammatory changes in peripheral blood leukocytes akin to those of sepsis. Am J Obstet Gynecol. 1998;179:80–86. doi: 10.1016/s0002-9378(98)70254-6. [DOI] [PubMed] [Google Scholar]
- 60.Redman CW, Sacks GP, Sargent IL. Preeclampsia: an excessive maternal inflammatory response to pregnancy. Am J Obstet Gynecol. 1999;180:499–506. doi: 10.1016/s0002-9378(99)70239-5. [DOI] [PubMed] [Google Scholar]
- 61.Gervasi MT, Chaiworapongsa T, Pacora P, et al. Phenotypic and metabolic characteristics of monocytes and granulocytes in preeclampsia. Am J Obstet Gynecol. 2001;185:792–797. doi: 10.1067/mob.2001.117311. [DOI] [PubMed] [Google Scholar]
- 62.Lau SY, Guild SJ, Barrett CJ, et al. Tumor necrosis factor-alpha, interleukin-6, and interleukin-10 levels are altered in preeclampsia: a systematic review and meta-analysis. Am J Reprod Immunol. 2013;70:412–427. doi: 10.1111/aji.12138. [DOI] [PubMed] [Google Scholar]
- 63.Boyd EM. The lipemia of pregnancy. J Clin Invest. 1934;13:347–363. doi: 10.1172/JCI100590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Potter JM, Nestel PJ. The hyperlipidemia of pregnancy in normal and complicated pregnancies. Am J Obstet Gynecol. 1979;133:165–170. doi: 10.1016/0002-9378(79)90469-1. [DOI] [PubMed] [Google Scholar]
- 65.Hubel CA, McLaughlin MK, Evans RW, et al. Fasting serum triglycerides, free fatty acids, and malondialdehyde are increased in preeclampsia, are positively correlated, and decrease within 48 hours post partum. Am J Obstet Gynecol. 1996;174:975–982. doi: 10.1016/s0002-9378(96)70336-8. [DOI] [PubMed] [Google Scholar]
- 66.Baker AM, Klein RL, Moss KL, et al. Maternal serum dyslipidemia occurs early in pregnancy in women with mild but not severe preeclampsia. Am J Obstet Gynecol. 2009;201:293.e1–4. doi: 10.1016/j.ajog.2009.05.037. [DOI] [PubMed] [Google Scholar]
- 67.Belo L, Santos-Silva A, Quintanilha A, et al. Similarities between pre-eclampsia and atherosclerosis: a protective effect of physical exercise? Curr Med Chem. 2008;15:2223–2229. doi: 10.2174/092986708785747553. [DOI] [PubMed] [Google Scholar]
- 68.Roberts JM, Redman CW. Pre-eclampsia: more than pregnancy-induced hypertension. Lancet. 1993;341:1447–1451. doi: 10.1016/0140-6736(93)90889-o. [DOI] [PubMed] [Google Scholar]
- 69.Lain KY, Roberts JM. Contemporary concepts of the pathogenesis and management of preeclampsia. JAMA. 2002;287:3183–3186. doi: 10.1001/jama.287.24.3183. [DOI] [PubMed] [Google Scholar]
- 70.Coelho TM, Sass N, Camano L, et al. Microvessel density in the placental bed among preeclampsia patients. Sao Paulo Med J. 2006;124:96–100. doi: 10.1590/S1516-31802006000200009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Labarrere CA, Nelson DR, Faulk WP. Endothelial activation and development of coronary artery disease in transplanted human hearts. JAMA. 1997;278:1169–1175. [PubMed] [Google Scholar]
- 72.Labarrere CA, Nelson DR, Miller SJ, et al. Value of serum-soluble intercellular adhesion molecule-1 for the noninvasive risk assessment of transplant coronary artery disease, posttransplant ischemic events, and cardiac graft failure. Circulation. 2000;102:1549–1555. doi: 10.1161/01.cir.102.13.1549. [DOI] [PubMed] [Google Scholar]
- 73.Labarrere CA, Lee JB, Nelson DR, et al. C-reactive protein, arterial endothelial activation, and development of transplant coronary artery disease: a prospective study. Lancet. 2002;360:1462–1467. doi: 10.1016/S0140-6736(02)11473-5. [DOI] [PubMed] [Google Scholar]
- 74.Labarrere CA, Jaeger BR. Biomarkers of heart transplant rejection: the good, the bad, and the ugly! Transl Res. 2012;159:238–251. doi: 10.1016/j.trsl.2012.01.018. [DOI] [PubMed] [Google Scholar]
- 75.Labarrere CA, Woods JR, Hardin JW, et al. Value of the first post-transplant biopsy for predicting long-term cardiac allograft vasculopathy (CAV) and graft failure in heart transplant patients. PLoS One. 2012;7:e36100. doi: 10.1371/journal.pone.0036100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Labarrere CA, Woods JR, Hardin JW, et al. Early inflammatory markers are independent preditors of cardiac allograft vasculopathy in heart-transplant recipients. PLoS One. 2014;9:e113260. doi: 10.1371/journal.pone.0113260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Schnoor M, Alcaide P, Voisin M-B, van Buul JD. Crossing the vascular wall: common and unique mechanisms exploited by different leukocyte subsets during extravasation. Mediators Inflamm. 2015;2015:946509. doi: 10.1155/2015/946509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Gerhardt T, Ley K. Monocyte trafficking across the vessel wall. Cardiovasc Res. 2015;107:321–30. doi: 10.1093/cvr/cvv147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Labarrere CA, Zaloga GP. C-reactive protein: from innocent bystander to pivotal mediator of atherosclerosis. Am J Med. 2004;117:499–507. doi: 10.1016/j.amjmed.2004.03.039. [DOI] [PubMed] [Google Scholar]
- 80.Roberts JM, Taylor RN, Musci TJ, et al. Preeclampsia: an endothelial cell disorder. Am J Obstet Gynecol. 1989;161:1200–1204. doi: 10.1016/0002-9378(89)90665-0. [DOI] [PubMed] [Google Scholar]
- 81.Roberts JM, Edep ME, Goldfien A, et al. Sera from preeclamptic women specifically activate human umbilical vein endothelial cells in vitro: morphological and biochemical evidence. Am J Reprod Immunol. 1992;27:101–108. doi: 10.1111/j.1600-0897.1992.tb00735.x. [DOI] [PubMed] [Google Scholar]
- 82.Wang Y, Zhang Y, Lewis DF, et al. Protease chymotrypsin mediates the endothelial expression of P- and E-selectin, but not ICAM and VCAM, induced by placental trophoblasts from pre-eclamptic pregnancies. Placenta. 2003;24:851–861. doi: 10.1016/s0143-4004(03)00132-2. [DOI] [PubMed] [Google Scholar]
- 83.Wang X, Athayde N, Trudinger B. Microvascular endothelial cell activation is present in the umbilical placental microcirculation in fetal placental vascular disease. Am J Obstet Gynecol. 2004;190:596–601. doi: 10.1016/j.ajog.2003.09.021. [DOI] [PubMed] [Google Scholar]
- 84.Chen Q, Guo F, Hensby-Bennett S, et al. Antiphospholipid antibodies prolong the activation of endothelial cells induced by necrotic trophoblastic debris: implications for the pathogenesis of preeclampsia. Placenta. 2012;33:810–815. doi: 10.1016/j.placenta.2012.07.019. [DOI] [PubMed] [Google Scholar]
- 85.Chen Q, Guo F, Liu S, et al. Calcium channel blockers prevent endothelial cell activation in response to necrotic trophoblast debris: possible relevance to pre-eclampsia. Cardiovasc Res. 2012;96:484–493. doi: 10.1093/cvr/cvs279. [DOI] [PubMed] [Google Scholar]
- 86.Shen F, Wei J, Snowise S, et al. Trophoblast debris extruded from preeclamptic placentae activates endothelial cells: a mechanism by which the placenta communicates with the maternal endothelium. Placenta. 2014;35:839–847. doi: 10.1016/j.placenta.2014.07.009. [DOI] [PubMed] [Google Scholar]
- 87.Chen Q, Stone PR, McCowan LM, et al. Activated endothelial cells resist displacement by trophoblast in vitro. Placenta. 2007;28:743–747. doi: 10.1016/j.placenta.2006.10.004. [DOI] [PubMed] [Google Scholar]
- 88.Oken E, Kleinman KP, Rich-Edwards J, Gillman MW. A nearly continuous measure of birth weight for gestational age using a United States national reference. BMC Pediatr. 2003;3:6. doi: 10.1186/1471-2431-3-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Juge-Aubry CE, Henrichot E, Meier CA. Adipose tissue: a regulator of inflammation. Best Pract Res Clin Endocrinol Metab. 2005;19:547–66. doi: 10.1016/j.beem.2005.07.009. [DOI] [PubMed] [Google Scholar]
- 90.Benirschke K, Burton GJ, Baergen RN. Pathology of the human placenta. Heidelberg, Germany: Springer-Verlag; 2012. [Google Scholar]
- 91.Labarrere CA, Faulk WP. Intercellular adhesion molecule-1 (ICAM-1) and HLA-DR antigens are expressed on endovascular cytotrophoblasts in abnormal pregnancies. Am J Reprod Immunol. 1995;33:47–53. doi: 10.1111/j.1600-0897.1995.tb01137.x. [DOI] [PubMed] [Google Scholar]
- 92.Reister F, Frank HG, Heyl W, et al. The distribution of macrophages in spiral arteries of the placental bed in pre-eclampsia differs from that in healthy patients. Placenta. 1999;20:229–233. doi: 10.1053/plac.1998.0373. [DOI] [PubMed] [Google Scholar]
- 93.DiFederico E, Genbacev O, Fisher SJ. Preeclampsia is associated with widespread apoptosis of placental cytotrophoblasts within the uterine wall. Am J Pathol. 1999;155:293–301. doi: 10.1016/S0002-9440(10)65123-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Pijnenborg R, McLaughlin PJ, Vercruysse L, et al. Immunolocalization of tumour necrosis factor-alpha (TNF-alpha) in the placental bed of normotensive and hypertensive human pregnancies. Placenta. 1998;19:231–239. doi: 10.1016/s0143-4004(98)90054-6. [DOI] [PubMed] [Google Scholar]
- 95.Harsem NK, Roald B, Braekke K, et al. Acute atherosis in decidual tissue: not associated with systemic oxidative stress in preeclampsia. Placenta. 2007;28:958–964. doi: 10.1016/j.placenta.2006.11.005. [DOI] [PubMed] [Google Scholar]
- 96.Staff AC, Dechend R, Redman CW. Review: Preeclampsia, acute atherosis of the spiral arteries and future cardiovascular disease: two new hypotheses. Placenta. 2013;34(Suppl):S73–78. doi: 10.1016/j.placenta.2012.11.022. [DOI] [PubMed] [Google Scholar]
- 97.Staff AC, Johnsen GM, Dechend R, et al. Preeclampsia and uteroplacental acute atherosis: immune and inflammatory factors. J Reprod Immunol. 2014;101–102:120–126. doi: 10.1016/j.jri.2013.09.001. [DOI] [PubMed] [Google Scholar]
- 98.Hiby SE, Walker JJ, O’shaughnessy KM, et al. Combinations of maternal KIR and fetal HLA-C genes influence the risk of preeclampsia and reproductive success. J Exp Med. 2004;200:957–965. doi: 10.1084/jem.20041214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Moffett A, Loke C. Immunology of placentation in eutherian mammals. Nat Rev Immunol. 2006;6:584–594. doi: 10.1038/nri1897. [DOI] [PubMed] [Google Scholar]
- 100.Moffett A, Hiby SE. How Does the maternal immune system contribute to the development of pre-eclampsia? Placenta. 2007;28(Suppl A):S51–56. doi: 10.1016/j.placenta.2006.11.008. [DOI] [PubMed] [Google Scholar]
- 101.Redman CW, Sargent IL. Immunology of pre-eclampsia. Am J Reprod Immunol. 2010;63:534–543. doi: 10.1111/j.1600-0897.2010.00831.x. [DOI] [PubMed] [Google Scholar]
- 102.Tilburgs T, Scherjon SA, van der Mast BJ, et al. Fetal-maternal HLA-C mismatch is associated with decidual T cell activation and induction of functional T regulatory cells. J Reprod Immunol. 2009;82:148–157. doi: 10.1016/j.jri.2009.05.003. [DOI] [PubMed] [Google Scholar]
- 103.Young J. The etiology of eclampsia and albuminuria and their relation to accidental hemorrhage: (an anatomical and experimental investigation) Proc R Soc Med. 1914;7:307–348. [PMC free article] [PubMed] [Google Scholar]
- 104.Bujold E, Romero R, Chaiworapongsa T, et al. Evidence supporting that the excess of the sVEGFR-1 concentration in maternal plasma in preeclampsia has a uterine origin. J Matern Fetal Neonatal Med. 2005;18:9–16. doi: 10.1080/14767050500202493. [DOI] [PubMed] [Google Scholar]
- 105.Gilbert JS, Babcock SA, Granger JP. Hypertension produced by reduced uterine perfusion in pregnant rats is associated with increased soluble fms-like tyrosine kinase-1 expression. Hypertension. 2007;50:1142–1147. doi: 10.1161/HYPERTENSIONAHA.107.096594. [DOI] [PubMed] [Google Scholar]
- 106.Makris A, Thornton C, Thompson J, et al. Uteroplacental ischemia results in proteinuric hypertension and elevated sFLT-1. Kidney Int. 2007;71:977–984. doi: 10.1038/sj.ki.5002175. [DOI] [PubMed] [Google Scholar]
- 107.Kusanovic JP, Romero R, Chaiworapongsa T, et al. A prospective cohort study of the value of maternal plasma concentrations of angiogenic and anti-angiogenic factors in early pregnancy and midtrimester in the identification of patients destined to develop preeclampsia. J Matern Fetal Neonatal Med. 2009;22:1021–1038. doi: 10.3109/14767050902994754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Verlohren S, Galindo A, Schlembach D, et al. An automated method for the determination of the sFlt-1/PIGF ratio in the assessment of preeclampsia. Am J Obstet Gynecol. 2010;202:161.e1–e11. doi: 10.1016/j.ajog.2009.09.016. [DOI] [PubMed] [Google Scholar]
- 109.Vaisbuch E, Whitty JE, Hassan SS, et al. Circulating angiogenic and antiangiogenic factors in women with eclampsia. Am J Obstet Gynecol. 2011;204:152.e1–e9. doi: 10.1016/j.ajog.2010.08.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Soto E, Romero R, Kusanovic JP, et al. Late-onset preeclampsia is associated with an imbalance of angiogenic and anti-angiogenic factors in patients with and without placental lesions consistent with maternal underperfusion. J Matern Fetal Neonatal Med. 2012;25:498–507. doi: 10.3109/14767058.2011.591461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.McElrath TF, Lim KH, Pare E, et al. Longitudinal evaluation of predictive value for preeclampsia of circulating angiogenic factors through pregnancy. Am J Obstet Gynecol. 2012;207:407.e1–e7. doi: 10.1016/j.ajog.2012.08.010. [DOI] [PubMed] [Google Scholar]
- 112.Wei SQ, Audibert F, Luo ZC, et al. Maternal plasma 25-hydroxyvitamin D levels, angiogenic factors, and preeclampsia. Am J Obstet Gynecol. 2013;208:390.e1–e6. doi: 10.1016/j.ajog.2013.03.025. [DOI] [PubMed] [Google Scholar]
- 113.Maynard SE, Crawford SL, Bathgate S, et al. Gestational angiogenic biomarker patterns in high risk preeclampsia groups. Am J Obstet Gynecol. 2013;209:53.e1–e9. doi: 10.1016/j.ajog.2013.03.017. [DOI] [PubMed] [Google Scholar]
- 114.Maynard SE, Min JY, Merchan J, et al. Excess placental soluble fms-like tyrosine kinase 1 (sFlt1) may contribute to endothelial dysfunction, hypertension, and proteinuria in preeclampsia. J Clin Invest. 2003;111:649–658. doi: 10.1172/JCI17189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Redman CW, Sargent IL. Latest advances in understanding preeclampsia. Science. 2005;308:1592–1594. doi: 10.1126/science.1111726. [DOI] [PubMed] [Google Scholar]
- 116.Roberts J. Pre-eclampsia a two-stage disorder: what is the linkage? Are there directed fetal/placental signals? In: Lyall F, Belfort M, editors. Pre-eclampsia: etiology and clinical practice. New York: Cambridge University Press; 2007. pp. 183–194. [Google Scholar]
- 117.Roberts JM, Hubel CA. The two stage model of preeclampsia: variations on the theme. Placenta. 2009;30(Suppl A):S32–37. doi: 10.1016/j.placenta.2008.11.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Venkatesha S, Toporsian M, Lam C, et al. Soluble endoglin contributes to the pathogenesis of preeclampsia. Nat Med. 2006;12:642–649. doi: 10.1038/nm1429. [DOI] [PubMed] [Google Scholar]
- 119.Levine RJ, Maynard SE, Qian C, et al. Circulating angiogenic factors and the risk of preeclampsia. N Engl J Med. 2004;350:672–683. doi: 10.1056/NEJMoa031884. [DOI] [PubMed] [Google Scholar]
- 120.Chaiworapongsa T, Romero R, Espinoza J, et al. Evidence supporting a role for blockade of the vascular endothelial growth factor system in the pathophysiology of preeclampsia. Young Investigator Award. Am J Obstet Gynecol. 2004;190:1541–1550. doi: 10.1016/j.ajog.2004.03.043. [DOI] [PubMed] [Google Scholar]
- 121.Chaiworapongsa T, Romero R, Kim YM, et al. Plasma soluble vascular endothelial growth factor receptor-1 concentration is elevated prior to the clinical diagnosis of pre-eclampsia. J Matern Fetal Neonatal Med. 2005;17:3–18. doi: 10.1080/14767050400028816. [DOI] [PubMed] [Google Scholar]
- 122.Widmer M, Villar J, Benigni A, et al. Mapping the theories of preeclampsia and the role of angiogenic factors: a systematic review. Obstet Gynecol. 2007;109:168–180. doi: 10.1097/01.AOG.0000249609.04831.7c. [DOI] [PubMed] [Google Scholar]
- 123.Romero R, Nien JK, Espinoza J, et al. A longitudinal study of angiogenic (placental growth factor) and anti-angiogenic (soluble endoglin and soluble vascular endothelial growth factor receptor-1) factors in normal pregnancy and patients destined to develop preeclampsia and deliver a small for gestational age neonate. J Matern Fetal Neonatal Med. 2008;21:9–23. doi: 10.1080/14767050701830480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Erez O, Romero R, Espinoza J, et al. The change in concentrations of angiogenic and anti-angiogenic factors in maternal plasma between the first and second trimesters in risk assessment for the subsequent development of preeclampsia and small-for-gestational age. J Matern Fetal Neonatal Med. 2008;21:279–287. doi: 10.1080/14767050802034545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Karumanchi SA, Stillman IE, Lindheimer MD. Angiogenesis and preeclampsia. In: Lindheimer MD, Roberts JM, Cunningham FG, editors. Chesley’s hypertensive disorders in pregnancy. Third. San Diego: Elsevier; 2009. pp. 87–104. [Google Scholar]
- 126.Espinoza J, Uckele JE, Starr RA, et al. Angiogenic imbalances: the obstetric perspective. Am J Obstet Gynecol. 2010;203:17.e1–e8. doi: 10.1016/j.ajog.2009.10.891. [DOI] [PubMed] [Google Scholar]
- 127.Chaiworapongsa T, Romero R, Savasan ZA, et al. Maternal plasma concentrations of angiogenic/anti-angiogenic factors are of prognostic value in patients presenting to the obstetrical triage area with the suspicion of preeclampsia. J Matern Fetal Neonatal Med. 2011;24:1187–1207. doi: 10.3109/14767058.2011.589932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Chaiworapongsa T, Romero R, Korzeniewski SJ, et al. Maternal plasma concentrations of angiogenic/antiangiogenic factors in the third trimester of pregnancy to identify the patient at risk for stillbirth at or near term and severe late preeclampsia. Am J Obstet Gynecol. 2013;208:287.e1–e15. doi: 10.1016/j.ajog.2013.01.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Chaiworapongsa T, Chaemsaithong P, Korzeniewski SJ, et al. Pre-eclampsia part 2: prediction, prevention and management. Nat Rev Nephrol. 2014;10:531–540. doi: 10.1038/nrneph.2014.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Ness RB, Roberts JM. Heterogeneous causes constituting the single syndrome of preeclampsia: a hypothesis and its implications. Am J Obstet Gynecol. 1996;175:1365–1370. doi: 10.1016/s0002-9378(96)70056-x. [DOI] [PubMed] [Google Scholar]
- 131.Redman CW, Sargent IL. Placental debris, oxidative stress and pre-eclampsia. Placenta. 2000;21:597–602. doi: 10.1053/plac.2000.0560. [DOI] [PubMed] [Google Scholar]
- 132.Redman CW, Sargent IL. Pre-eclampsia, the placenta and the maternal systemic inflammatory response–a review. Placenta. 2003;24(Suppl A):S21–27. doi: 10.1053/plac.2002.0930. [DOI] [PubMed] [Google Scholar]
- 133.Redman CW, Tannetta DS, Dragovic RA, et al. Review: Does size matter? Placental debris and the pathophysiology of pre-eclampsia. Placenta. 2012;33(Suppl):S48–54. doi: 10.1016/j.placenta.2011.12.006. [DOI] [PubMed] [Google Scholar]
- 134.Rajakumar A, Cerdeira AS, Rana S, et al. Transcriptionally active syncytial aggregates in the maternal circulation may contribute to circulating soluble fms-like tyrosine kinase 1 in preeclampsia. Hypertension. 2012;59:256–264. doi: 10.1161/HYPERTENSIONAHA.111.182170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Karumanchi SA, Bdolah Y. Hypoxia and sFlt-1 in preeclampsia: the “chicken-and-egg” question. Endocrinology. 2004;145:4835–4837. doi: 10.1210/en.2004-1028. [DOI] [PubMed] [Google Scholar]
- 136.Ross R. Atherosclerosis–an inflammatory disease. N Engl J Med. 1999;340:115–126. doi: 10.1056/NEJM199901143400207. [DOI] [PubMed] [Google Scholar]
- 137.Ross R. Atherosclerosis is an inflammatory disease. Am Heart J. 1999;138:S419–420. doi: 10.1016/s0002-8703(99)70266-8. [DOI] [PubMed] [Google Scholar]
- 138.Vince GS, Starkey PM, Austgulen R, et al. Interleukin-6, tumour necrosis factor and soluble tumour necrosis factor receptors in women with pre-eclampsia. Br J Obstet Gynaecol. 1995;102:20–25. doi: 10.1111/j.1471-0528.1995.tb09020.x. [DOI] [PubMed] [Google Scholar]
- 139.Conrad KP, Miles TM, Benyo DF. Circulating levels of immunoreactive cytokines in women with preeclampsia. Am J Reprod Immunol. 1998;40:102–111. doi: 10.1111/j.1600-0897.1998.tb00398.x. [DOI] [PubMed] [Google Scholar]
- 140.Yudkin JS, Kumari M, Humphries SE, et al. Inflammation, obesity, stress and coronary heart disease: is interleukin-6 the link? Atherosclerosis. 2000;148:209–214. doi: 10.1016/s0021-9150(99)00463-3. [DOI] [PubMed] [Google Scholar]
- 141.Teran E, Escudero C, Moya W, et al. Elevated C-reactive protein and pro-inflammatory cytokines in Andean women with pre-eclampsia. Int J Gynaecol Obstet. 2001;75:243–249. doi: 10.1016/s0020-7292(01)00499-4. [DOI] [PubMed] [Google Scholar]
- 142.Freeman DJ, McManus F, Brown EA, et al. Short- and long-term changes in plasma inflammatory markers associated with preeclampsia. Hypertension. 2004;44:708–714. doi: 10.1161/01.HYP.0000143849.67254.ca. [DOI] [PubMed] [Google Scholar]
- 143.Sharma A, Satyam A, Sharma JB. Leptin, IL-10 and inflammatory markers (TNF-alpha, IL-6 and IL-8) in pre-eclamptic, normotensive pregnant and healthy non-pregnant women. Am J Reprod Immunol. 2007;58:21–30. doi: 10.1111/j.1600-0897.2007.00486.x. [DOI] [PubMed] [Google Scholar]
- 144.Belo L, Santos-Silva A, Caslake M, et al. Neutrophil activation and C-reactive protein concentration in preeclampsia. Hypertens Pregnancy. 2003;22:129–141. doi: 10.1081/PRG-120021059. [DOI] [PubMed] [Google Scholar]
- 145.Braekke K, Holthe MR, Harsem NK, et al. Calprotectin, a marker of inflammation, is elevated in the maternal but not in the fetal circulation in preeclampsia. Am J Obstet Gynecol. 2005;193:227–233. doi: 10.1016/j.ajog.2004.11.055. [DOI] [PubMed] [Google Scholar]
- 146.Ustun Y, Engin-Ustun Y, Kamaci M. Association of fibrinogen and C-reactive protein with severity of preeclampsia. Eur J Obstet Gynecol Reprod Biol. 2005;121:154–158. doi: 10.1016/j.ejogrb.2004.12.009. [DOI] [PubMed] [Google Scholar]
- 147.Greer IA, Haddad NG, Dawes J, et al. Neutrophil activation in pregnancy-induced hypertension. Br J Obstet Gynaecol. 1989;96:978–982. doi: 10.1111/j.1471-0528.1989.tb03358.x. [DOI] [PubMed] [Google Scholar]
- 148.Sattar N, Greer IA. Pregnancy complications and maternal cardiovascular risk: opportunities for intervention and screening? BMJ. 2002;325:157–160. doi: 10.1136/bmj.325.7356.157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Hermes W, Franx A, van Pampus MG, et al. Cardiovascular risk factors in women who had hypertensive disorders late in pregnancy: a cohort study. Am J Obstet Gynecol. 2013;208:474.e1–e8. doi: 10.1016/j.ajog.2013.02.016. [DOI] [PubMed] [Google Scholar]
- 150.Ahmed R, Dunford J, Mehran R, et al. Pre-eclampsia and future cardiovascular risk among women: a review. J Am Coll Cardiol. 2014;63:1815–1822. doi: 10.1016/j.jacc.2014.02.529. [DOI] [PubMed] [Google Scholar]
- 151.Alsnes IV, Janszky I, Forman MR, et al. A population-based study of associations between preeclampsia and later cardiovascular risk factors. Am J Obstet Gynecol. 2014;211:657.e1–e7. doi: 10.1016/j.ajog.2014.06.026. [DOI] [PubMed] [Google Scholar]
- 152.Reister F, Frank HG, Kingdom JC, et al. Macrophage-induced apoptosis limits endovascular trophoblast invasion in the uterine wall of preeclamptic women. Lab Invest. 2001;81:1143–1152. doi: 10.1038/labinvest.3780326. [DOI] [PubMed] [Google Scholar]
- 153.Labarrere CA, Ortiz MA, Sosa MJ, et al. Syncytiotrophoblast intercellular adhesion molecule-1 expression in placental villitis of unknown cause. Am J Obstet Gynecol. 2005;193:483–488. doi: 10.1016/j.ajog.2004.12.090. [DOI] [PubMed] [Google Scholar]
- 154.Labarrere CA, Bammerlin E, Hardin JW, et al. Intercellular adhesion molecule-1 expression in massive chronic intervillositis: Implications for the invasion of maternal cells into fetal tissues. Placenta. 2014;35:311–317. doi: 10.1016/j.placenta.2014.02.006. [DOI] [PubMed] [Google Scholar]
- 155.Xiao J, Garcia-Lloret M, Winkler-Lowen B, et al. ICAM-1-mediated adhesion of peripheral blood monocytes to the maternal surface of placental syncytiotrophoblasts: implications for placental villitis. Am J Pathol. 1997;150:1845–1860. [PMC free article] [PubMed] [Google Scholar]
- 156.Garcia-Lloret MI, Winkler-Lowen B, Guilbert LJ. Monocytes adhering by LFA-1 to placental syncytiotrophoblasts induce local apoptosis via release of TNF-alpha. A model for hematogenous initiation of placental inflammations. J Leukoc Biol. 2000;68:903–908. [PubMed] [Google Scholar]
- 157.Goksu Erol AY, Nazli M, Elis Yildiz S. Significance of platelet endothelial cell adhesion molecule-1 (PECAM-1) and intercellular adhesion molecule-1 (ICAM-1) expressions in preeclamptic placentae. Endocrine. 2012;42:125–131. doi: 10.1007/s12020-012-9644-9. [DOI] [PubMed] [Google Scholar]
- 158.Romero R, Kusanovic JP, Chaiworapongsa T, Hassan SS. Placental bed disorders in preterm labor, preterm PROM, spontaneous abortion and abruptio placentae. Best Pract Res Clin Obstet Gynaecol. 2011;25:313–327. doi: 10.1016/j.bpobgyn.2011.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Kim YM, Chaemsaithong P, Romero R, et al. The frequency of acute atherosis in normal pregnancy and preterm labor, preeclampsia, small-for-gestational age, fetal death and midtrimester spontaneous abortion. J Matern Fetal Neonatal Med. 2015;28:2001–2009. doi: 10.3109/14767058.2014.976198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Romero R, Kusanovic JP, Kim CJ. Placental bed disorders in the genesis of the great obstetrical syndromes. In: Pijnenborg R, Brosens I, Romero R, editors. Placental bed disorders. New York, NY: Cambridge University Press; 2010. pp. 271–289. [Google Scholar]
- 161.Korzeniewski SJ, Romero R, Chaiworapongsa T, et al. Maternal plasma angiogenic index-1 (placental growth factor/soluble vascular endothelial growth factor receptor-1) is a biomarker for the burden of placental lesions consistent with uteroplacental underperfusion: a longitudinal case-cohort study. Am J Obstet Gynecol. 2016;214:629.e1–e17. doi: 10.1016/j.ajog.2015.11.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Kim YM, Chaemsaithong P, Romero R, et al. Placental lesions associated with acute atherosis. J Matern Fetal Neonatal Med. 2015;28:1554–1562. doi: 10.3109/14767058.2014.960835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Kam EP, Gardner L, Loke YW, et al. The role of trophoblast in the physiological change in decidual spiral arteries. Hum Reprod. 1999;14:2131–2138. doi: 10.1093/humrep/14.8.2131. [DOI] [PubMed] [Google Scholar]
- 164.Erlebacher A. Why isn’t the fetus rejected? Curr Opin Immunol. 2001;13:590–593. doi: 10.1016/s0952-7915(00)00264-8. [DOI] [PubMed] [Google Scholar]
- 165.Szekeres-Bartho J. Immunological relationship between the mother and the fetus. Int Rev Immunol. 2002;21:471–495. doi: 10.1080/08830180215017. [DOI] [PubMed] [Google Scholar]
- 166.Trowsdale J, Betz AG. Mother’s little helpers: mechanisms of maternal-fetal tolerance. Nat Immunol. 2006;7:241–246. doi: 10.1038/ni1317. [DOI] [PubMed] [Google Scholar]
- 167.Rowe JH, Ertelt JM, Xin L, et al. Pregnancy imprints regulatory memory that sustains anergy to fetal antigen. Nature. 2012;490:102–106. doi: 10.1038/nature11462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Betz AG. Immunology: Tolerating pregnancy. Nature. 2012;490:47–48. doi: 10.1038/490047a. [DOI] [PubMed] [Google Scholar]
- 169.Erlebacher A. Immunology of the maternal-fetal interface. Annu Rev Immunol. 2013;31:387–411. doi: 10.1146/annurev-immunol-032712-100003. [DOI] [PubMed] [Google Scholar]
- 170.Sabatier F, Bretelle F, D’Ercole C, et al. Neutrophil activation in preeclampsia and isolated intrauterine growth restriction. Am J Obstet Gynecol. 2000;183:1558–1563. doi: 10.1067/mob.2000.108082. [DOI] [PubMed] [Google Scholar]
- 171.Ogge G, Romero R, Chaiworapongsa T, et al. Leukocytes of pregnant women with small-for-gestational age neonates have a different phenotypic and metabolic activity from those of women with preeclampsia. J Matern Fetal Neonatal Med. 2010;23:476–487. doi: 10.3109/14767050903216033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Gervasi MT, Chaiworapongsa T, Naccasha N, et al. Phenotypic and metabolic characteristics of maternal monocytes and granulocytes in preterm labor with intact membranes. Am J Obstet Gynecol. 2001;185:1124–1129. doi: 10.1067/mob.2001.117681. [DOI] [PubMed] [Google Scholar]
- 173.Gervasi MT, Chaiworapongsa T, Naccasha N, et al. Maternal intravascular inflammation in preterm premature rupture of membranes. J Matern Fetal Neonatal Med. 2002;11:171–175. doi: 10.1080/jmf.11.3.171.175. [DOI] [PubMed] [Google Scholar]
- 174.Naccasha N, Gervasi MT, Chaiworapongsa T, et al. Phenotypic and metabolic characteristics of monocytes and granulocytes in normal pregnancy and maternal infection. Am J Obstet Gynecol. 2001;185:1118–1123. doi: 10.1067/mob.2001.117682. [DOI] [PubMed] [Google Scholar]
- 175.Khong TY, Mott C. Immunohistologic demonstration of endothelial disruption in acute atherosis in pre-eclampsia. Eur J Obstet Gynecol Reprod Biol. 1993;51:193–7. doi: 10.1016/0028-2243(93)90034-a. [DOI] [PubMed] [Google Scholar]
- 176.Khong TY, Sawyer IH, Heryet AR. An immunohistologic study of endothelialization of uteroplacental vessels in human pregnancy—evidence that endothelium is focally disrupted by trophoblast in preeclampsia. Am J Obstet Gynecol. 1992;167:751–6. doi: 10.1016/s0002-9378(11)91583-x. [DOI] [PubMed] [Google Scholar]