

Abstract
Tripartite integrative and conjugative elements (ICE3) are a novel form of ICE that exist as three separate DNA regions integrated within the genomes of Mesorhizobium spp. Prior to conjugative transfer the three ICE3 regions of M. ciceri WSM1271 ICEMcSym1271 combine and excise to form a single circular element. This assembly requires three coordinated recombination events involving three site-specific recombinases IntS, IntG and IntM. Here, we demonstrate that three excisionases–or recombination directionality factors—RdfS, RdfG and RdfM are required for ICE3 excision. Transcriptome sequencing revealed that expression of ICE3 transfer and conjugation genes was induced by quorum sensing. Quorum sensing activated expression of rdfS, and in turn RdfS stimulated transcription of both rdfG and rdfM. Therefore, RdfS acts as a “master controller” of ICE3 assembly and excision. The dependence of all three excisive reactions on RdfS ensures that ICE3 excision occurs via a stepwise sequence of recombination events that avoids splitting the chromosome into a non-viable configuration. These discoveries expose a surprisingly simple control system guiding molecular assembly of these novel and complex mobile genetic elements and highlight the diverse and critical functions of excisionase proteins in control of horizontal gene transfer.
Author summary
Bacteria evolve and adapt quickly through the horizontal transfer of DNA. A major mechanism facilitating this transfer is conjugation. Conjugative DNA elements that integrate into the chromosome are termed ‘Integrative and Conjugative Elements’ (ICE). We recently discovered a unique form of ICE that undergoes a complex series of recombination events with the host chromosome to split itself into three separate parts. This tripartite ICE must also precisely order its recombination when leaving the current host to avoid splitting the host chromosome and the ICE into non-viable parts. In this work, we show that the tripartite ICEs use chemical cell-cell communication to stimulate recombination and that recombination events are specifically ordered through cascaded transcriptional activation of small DNA-binding proteins called recombination directionality factors. Despite the inherent complexity of tripartite ICEs this work exposes a surprisingly simple system to stimulate their precise and ordered molecular assembly prior to horizontal transfer.
Introduction
Bacterial genome evolution proceeds at a rapid pace largely due to the sharing of genetic material [1]. This gene exchange is often facilitated by mobile genetic elements (MGEs) such as plasmids, bacteriophage and other chromosomally-integrating elements [2, 3]. MGEs have evolved sophisticated mechanisms to maintain themselves in their host while opportunistically infecting neighbouring organisms, maximising their dissemination through both vertical and horizontal modes of descent [4]. MGEs frequently harbour genes conferring selective benefit to hosts such as virulence, metabolism, symbiosis and antimicrobial-resistance determinants [5–10]. The rapid progress in genome sequencing this century has revealed the ubiquity of MGEs in microbial genomes and specifically, the abundance of MGE-encoded conjugation systems highlights conjugation as a major mechanism of horizontal transmission [11]. It has also become increasingly apparent that ‘non-conjugative’ plasmids and chromosomally-integrating elements may exploit conjugation systems encoded by other MGEs for their own transfer [12–14]. The bacterial mobilome can therefore be viewed as a DNA ecosystem where MGEs compete for an environmental niche defined by the hosts in which they can infect and persist.
Integrative and conjugative elements (ICEs) are the most recently defined MGE, but are probably the most abundant conjugative elements in bacteria [11]. Unlike plasmids, ICEs integrate within their host’s chromosome, negating a strict requirement for full-time extrachromosomal replication systems [15, 16]. Once stimulated to transfer, ICEs excise from the chromosome to form a circular episome capable of conjugation. Rolling-circle replication is an essential part of most conjugation systems so most ICEs likely have the capacity to replicate via this mechanism once excised [17]. Cells carrying an excised ICE can persist in this transfer-competent state and potentially donate ICEs to multiple recipients. Understanding the triggers for ICE transfer requires knowledge of regulatory cues stimulating ICE excision.
Symbiosis ICEs of Mesorhizobium spp. are a diverse family of large (~0.5-Mb) ICEs capable of converting non-symbiotic mesorhizobia into symbionts of plant legume species [8, 18–22]. The symbiosis ICE of M. loti R7A, ICEMlSymR7A, is a 502-kb ICE encoding genes enabling symbiosis with Lotus spp. [8, 18, 21, 23]. Integration of ICEMlSymR7A into mesorhizobial chromosomes is facilitated by the tyrosine recombinase (integrase) IntS [23]. The IntS attachment site attPS (the subscript denotes the integrase associated with the att site) located on the excised circular ICEMlSymR7A contains a 17-bp DNA sequence identical to the 3’-end of the sole phe-tRNA gene (attBS), which is the target for IntS-mediated recombination. Recombination between attPS and attBS produces the hybrid sites attLS and attRS, which flank the integrated ICEMlSymR7A and together form a direct 17-bp repeat demarcating the ICEMlSymR7A insertion site [18, 23].
Integrase-mediated recombination can be modulated by additional protein factors that alter the integrase-DNA complex and favoured direction of recombination [24, 25]. Recombination directionality factors (RDFs, or excisionases) are generally small winged-helix-turn-helix domain DNA-binding proteins that bend DNA within integrase att sites [25]. Excision of ICEMlSymR7A requires the RDF RdfS (S1 Fig). Overexpression of rdfS cures ICEMlSymR7A from M. loti R7A cells producing the non-symbiotic derivative R7ANS [23]. A synthetic non-replicative mini-ICE carrying only attPS and intS is able to integrate into the attBS site of R7ANS, confirming IntS is the only ICEMlSymR7A protein required for integration. Subsequent introduction of a plasmid constitutively expressing rdfS stimulates loss of the integrated mini-ICE from R7ANS [23], suggesting that like other excisionases, RdfS probably binds the IntS attachment sites to stimulate IntS-catalysed formation of attPS and attBS.
Recently we identified a new form of ICE, termed a tripartite ICE (ICE3), composed of three separated chromosomal regions of DNA α, β and γ [19, 26]. Three site-specific recombination reactions assemble these ICE3 regions into a single circular entity prior to conjugation. The ICE3 of M. ciceri WSM1271 (ICEMcSym1271) carries homologues of rdfS, intS and all genes identified as being required for horizontal transfer of ICEMlSymR7A. However, ICEMcSym1271 carries two additional tyrosine recombinases IntG and IntM, two additional predicted excisionases RdfG and RdfM and two additional sets of attachment sites attLG, attRG, attPG, attBG, and attLM, attRM. attPM, attBM (Fig 1A) [19]. Using a synthetic non-replicative mini-ICE3 element containing all three attP sites derived from ICEMcSym1271, IntS, IntG and IntM were demonstrated to mediate chromosomal integration and subsequent dispersal of this mini-ICE3 into the separate regions α, β and γ [19]. We additionally identified numerous putative tripartite ICEs in diverse symbiotic mesorhizobia, each carrying unique genetic cargo in each ICE3 region. We propose that the tripartite integration pattern serves to stabilize the ICE in the host and protect it from potential destabilisation by competing ICEs and other integrative elements [26].
Fig 1. ICEMcSym1271 assembly, excision and regulation.

(A) Schematic of the possible ICEMcSym1271 recombination states and recombination reactions leading to formation of excised ICEMcSym1271 assembled from regions α, β and γ. The recombination targets attP, attB, attL and attR (triangles) and recombination reactions (large arrows) are color-coded for each integrase: cyan for IntS; magenta for IntG; green for IntM. Primers for qPCR measurement of recombination are indicated as block-headed arrows for attB sites and triangle-headed arrows for attP sites (see Materials and Methods and S1 Table for details). Data presented here support a model where RDF-stimulated excisive reactions occur in the order IntS > IntG > IntM (highlighted in yellow) to produce excised ICEMcSym1271. (B) The regulatory model of quorum-sensing mediated stimulation of ICEMcSym1271 assembly and excision. TraR1 and TraR2 bind AHLs produced by TraI1. TraR1/2-AHL complex(es) activate transcription from the traI1 and traI2 promoters. traI2-msi172-msi171 expression leads to production of FseA and transcriptional activation of the rdfS operon. RdfS stimulates excisive IntS-mediated recombination and promotes expression of RdfG and RdfM. RdfG stimulates the excisive IntG-mediated reaction and RdfM stimulates excisive IntM-mediated recombination and excision.
The increased complexity introduced by the three separate recombination reactions required for ICEMcSym1271 integration and excision allows for the potential formation of eight distinct chromosomal recombination states [19]. The arrival at any particular state depends on the prior order and direction of the recombination reactions catalysed by IntS, IntG and IntM. Not all eight states can be reconstructed using the mini-ICE3, suggesting some states are non-viable. Specifically, our model indicates that if the first excisive reaction is catalysed by IntM, i.e. attLM + attRM > attPM + attBM, then the chromosome is split into two parts, one part lacking the likely essential phe and his-tRNA genes and the other part an origin-of-replication (Fig 1A) (ICE3 recombination reactions producing attP + attB do not necessarily result in ICE3 excision per se, but for simplicity will be referred as ‘excisive’). Quantitative PCR (qPCR) assays measuring IntM-mediated formation of attPM + attBM indicate the excisive IntM reaction occurs at the lowest frequency of the three integrase-mediated reactions [19], suggesting evolved regulatory control mechanisms might prevent IntM-mediated excisive recombination occurring before other reactions, precluding formation of the non-viable chromosome state.
In this work, we show that the three excisive reactions of ICEMcSym1271 are dependent on three distinct RDFs, RdfG, RdfM, and RdfS. ICEMcSym1271 excision and transfer is stimulated by quorum sensing (QS). RNA sequencing (RNAseq) revealed QS activation results in activation of rdfS expression (Fig 1B). Surprisingly, all three attL + attR > attP + attB reactions were dependent on rdfS and we demonstrated that this is because RdfS activates expression from the rdfG and rdfM promoters. Therefore, the ordered assembly and excision of ICEMcSym1271 is accomplished through a cascade of transcriptional activation initiated by QS and finalised by RdfS, ensuring RdfS is always the first excisionase translated and that IntS-catalysed excisive recombination occurs ahead of the IntG and IntM-catalysed reactions.
Results
RdfG and RdfM are required for excisive IntG and IntM-mediated recombination
Integration (formation of attL + attR from attP + attB) and excision (formation of attP + attB from attL + attR) of ICEMlSymR7A are catalysed by the integrase IntS, however, integration is favoured in the absence of RdfS. Overexpression of rdfS in R7A stimulates the excision reaction and results in loss of ICEMlSymR7A from the cell [23]. In contrast, the tripartite ICEMcSym1271 of WSM1271 requires the concerted action of three integrases IntG, IntM, and IntS to direct integration and excision [19]. In addition to a homologue of rdfS, two other putative excisionase genes rdfG and rdfM are present on ICEMcSym1271 [19, 26]. rdfG is oriented convergently with intG on ICEMcSym1271 region β and rdfM is encoded directly upstream of intM on ICEMcSym1271 region γ. Like RdfS, RdfG (Mesci_2550) and RdfM (Mesci_2345) are MerR superfamily proteins with a predicted winged-helix-turn-helix secondary structure (S1 Fig). To investigate potential roles of rdfG and rdfM we replaced each gene with an ΩaadA cassette producing strains 1271ΔrdfG::ΩaadA and 1271ΔrdfM::ΩaadA, respectively, and using our previously validated qPCR assay [19], measured the abundance of each the three pairs of attP and attB sites formed following each of the three excisive reactions. In wild-type WSM1271, attPG + attBG and attPS + attBS sites were detected at a frequency of 0.1–1% per chromosome and attPM + attBM sites were detected at 0.01–0.1% (Fig 2A). In contrast, attPG + attBG sites were undetectable in 1271ΔrdfG::ΩaadA and attPM + attBM sites were undetectable in 1271ΔrdfM::ΩaadA. The abundance of the two remaining pairs of attP + attB sites in each of these mutant strains was similar to that of WSM1271. Complementation of 1271ΔrdfG::ΩaadA with a cloned copy of rdfG and its native promoter partially restored attPG + attBG formation and complementation of 1271ΔrdfM::ΩaadA with a cloned copy of rdfM and its native promoter restored attPM + attBM production. These experiments therefore confirmed the roles of RdfG and RdfM in excisive IntG and IntM reactions, respectively.
Fig 2. qPCR measurement of excisive ICEMcSym1271 recombination.

Measurements represent the mean percentage of WSM1271 chromosomes in stationary-phase cultures harbouring each excisive Int-mediated recombination product (attBS, attPS, attBG, attPG, attPM, and attPM) determined by qPCR [19]. Where appropriate, plasmids carried by WSM1271 (here abbreviated as 1271) are listed in brackets after the strain name (see Table 3 for a description of plasmids). Values for each of the assay types attBS, attPS, attBG, attPG, attPM, and attPM site were individually compared between strains within the same panel (panel A, B, or C) using ANOVA and Fisher’s LSD test controlling for type I error using the Bonferroni adjustment. Groups of values from the same assay type and in the same panel that are not significantly different from each other have the same letter (a, b, c, d, e, f or g) indicated above. Expression from the IPTG inducible promoter of pSDz constructs were not induced with IPTG as they exhibit leaky expression without induction in TY medium used for assays. (A) Involvement of rdfG and rdfM in excisive recombination. (B) Quorum-sensing induction of excisive recombination. (C) Involvement of rdfS in excisive recombination.
Quorum sensing stimulates all three excisive Int-mediated recombination reactions
All three pairs of ICEMcSym1271 attP and attB products are most abundant in stationary-phase cultures and the α region of ICEMcSym1271 carries a subset of genes homologous to those involved in quorum-sensing (QS) regulation of ICEMlSymR7A excision and conjugative transfer in R7A [19, 23, 27]. These QS genes include a homologue of the ICEMlSymR7A N-acyl-homoserine lactone (AHL)-synthase gene traI1 (Mesci_5572) and a homologue of the AHL-dependent transcriptional regulator traR (Mesci_5573), here named traR1. A second traR homologue (Mesci_5676), here named traR2, is present on ICEMcSym1271-α adjacent to a homologue of the QS antiactivation gene qseM [28]. Therefore, we suspected that excision and conjugative transfer of ICEMcSym1271 was under QS control.
To confirm ICEMcSym1271 traI1 encoded a functional AHL synthase, traI1 was cloned into pPR3 downstream of the nptII promoter. The resulting plasmid pPR3-traI1 was introduced into E. coli DH5α and M. loti R7ANS and the resulting strains were screened for AHL production using the Chromobacterium violaceum CV026 AHL bioassay [29]. CV026 violacein production was induced by E. coli and M. loti strains carrying the pPR3-traI1 vector (S2 Fig), but not by strains carrying the vector-only control pPR3, indicating that traI1 produced C4-C8 AHLs in both backgrounds. In M. loti R7A, AHL-activated TraR stimulates transcription of traI1 completing a positive feedback loop of regulation [27]. To confirm TraR1 and TraR2 activated expression from the traI1 promoter of ICEMcSym1271 (PtraI1), traR1 and traR2 were cloned downstream of the lac promoter on a derivative of pSDz also carrying PtraI1 upstream of the lacZ gene. The resulting constructs were mobilized into R7ANS additionally carrying either pPR3-traI1 or pPR3. β-galactosidase assays of the resulting strains revealed that both traR1 and traR2 partially induced expression from PtraI1 in the absence of traI1 and other ICE-encoded genes, however, maximum expression from this promoter was only achieved in the presence of traI1 (S3 Fig).
traI1, traR1 and traR2 were next each individually overexpressed in WSM1271 on plasmids and ICE3 excision was measured by qPCR (Fig 2B). Constitutive expression of traI1 from the nptII promoter stimulated a 10-100-fold increase in abundance all three attP + attB sites relative to vector-only controls. Non-induced lac promoter-driven expression of traR1 or traR2 only stimulated a modest increase in att site abundance relative to WSM1271, however, unexplainedly the vector-only control exhibited ~10-fold reduced excision frequencies, so relative to this background overexpression of the traR1/2 genes each induced a 10-100-fold increase for all attP + attB sites. Overexpression of traI1 and traR1 in the same background stimulated ~1000-fold increase in abundance all three attP + attB sites relative to the vector-only control strain. To investigate effects of the QS genes on conjugative transfer, strains overexpressing traR1, traR2, and traI1 were each used as donors in mating assays where M. loti R7ANS carrying pPR3 or pFAJ1708 was the recipient (Table 1). The pattern of fold-changes in conjugation frequencies for each donor strain largely mirrored excision frequency changes observed in qPCR assays (Fig 2B) confirming that traI1, traR1 and traR2 also stimulated conjugative transfer.
Table 1. Quorum-sensing induced ICEMcSym1271 conjugative transfer.
| abDonor | Recipient | Exconjugants (per donor) | Standard deviation | cFold-change |
|---|---|---|---|---|
| WSM1271 | R7ANS(pPR3) | 8.02 x 10−8 | 1.82 x 10−8 | - |
| WSM1271(pSDz) | R7ANS(pPR3) | 2.22 x 10−8 | 9.12 x 10−9 | - |
| WSM1271(pSDz-traR1) | R7ANS(pPR3) | 4.69 x 10−7 | 1.11 x 10−7 | 21.14 |
| WSM1271(pSDz-traR2) | R7ANS(pPR3) | 5.97 x 10−7 | 1.66 x 10−7 | 26.90 |
| WSM1271(pSDz-msi172171) | R7ANS(pPR3) | 8.49 x 10−7 | 8.23 x 10−8 | 38.30 |
| WSM1271 | R7ANS(pFAJ1708) | 8.35 x 10−8 | 4.87 x 10−8 | - |
| WSM1271(pPR3) | R7ANS(pFAJ1708) | 8.74 x 10−8 | 3.89 x 10−8 | - |
| WSM1271(pPR3-traI1) | R7ANS(pFAJ1708) | 1.04 x 10−5 | 1.50 x 10−6 | 119.06 |
a Where appropriate, plasmids carried by WSM1271 are listed in brackets after the strain name (see Table 3 for a description of plasmids).
b Expression from the IPTG inducible promoter of pSDz constructs were not induced with IPTG as they exhibit leaky expression without induction in TY medium used for assays.
c Fold-change is relative to control strains carrying the appropriate pPR3 or pSDz parent vector.
Dissection of quorum sensing-induced ICE3 excision using RNA deep sequencing
QS-induced excision and conjugative transfer of ICEMlSymR7A is dependent on the transcriptional activation of rdfS [30]. In M. loti R7A, AHL-activated TraR stimulates transcription from ‘tra-box’ promoters centred 69-bp upstream of traI1 and 67-bp upstream of the traI2-msi172-msi171 operon. A programmed ribosomal frameshift site encoded in the 3’ end of msi172 facilitates translational fusion of Msi172 and Msi171, producing FseA, a regulator essential for transcription from the rdfS promoter [27, 31, 32]. Homologues of msi172-msi171 and rdfS are also located on the α region of ICEMcSym1271 [19], therefore it seemed likely these genes also stimulated ICEMcSym1271 excision. Transcriptome sequencing (RNAseq) was carried out for a QS-induced (QS+) strain carrying plasmid-borne copies of traI1 and traR1 and an uninduced strain (QS-) carrying the appropriate empty vectors. Overall, 187 significantly differentially expressed genes (adjusted P-value < 0.05) were identified (S1 Dataset) and although ICEMcSym1271 comprised only ~7.6% of the chromosome, 29 (15.5%) of the differentially expressed genes were located on ICEMcSym1271. Genes likely involved in activation of excision and conjugation including rdfS, rlxS and the type-IV conjugative pilus gene cluster msi031-trbBCDEJLFGI-msi021 were all significantly induced (Table 2).
Table 2. Quorum-sensing induced/repressed ICEMcSym1271-encoded genes.
| Gene | Locus ID | aFold-change | Standard error |
|---|---|---|---|
| Region-α | |||
| rdfS | Mesci_5530 | 19.74 | 1.20 |
| traF | Mesci_5529 | 29.21 | 1.20 |
| msi107 | Mesci_5528 | 41.10 | 1.19 |
| rlxS | Mesci_5527 | 58.14 | 1.17 |
| PtraI1 | - | 121.45 | 1.16 |
| P traI2 | - | 37.54 | 1.18 |
| traI2 | - | 141.41 | 1.16 |
| msi172 | - | 61.71 | 1.18 |
| msi171 | - | 156.99 | 1.16 |
| msi021 | Mesci_5513 | 8.28 | 1.19 |
| trbI | Mesci_5514 | 10.58 | 1.17 |
| trbG | Mesci_5515 | 18.07 | 1.19 |
| trbF | Mesci_5516 | 14.48 | 1.19 |
| trbL | Mesci_5517 | 19.35 | 1.19 |
| trbJ | Mesci_5518 | 42.31 | 1.18 |
| trbE | Mesci_5519 | 64.16 | 1.17 |
| trbD | Mesci_5520 | 14.43 | 1.20 |
| trbC | Mesci_5521 | 9.71 | 1.20 |
| trbB | Mesci_5522 | 5.39 | 1.21 |
| msi031 | Mesci_5523 | 13.88 | 1.20 |
| traG | Mesci_5524 | 2.75 | 1.16 |
| queD | Mesci_5560 | -2.35 | 0.83 |
| queC | Mesci_5561 | -2.29 | 0.82 |
| queB | Mesci_5562 | -2.34 | 0.83 |
| hypothetical | Mesci_5526 | 1.90 | 1.18 |
| Region-β | |||
| cbb3-type COx (SI) | Mesci_5510 | 1.92 | 1.16 |
| Nicotinate biosynthesis protein | Mesci_5579 | -1.85 | 0.83 |
| rdfG | Mesci_2550 | 2.46 | 1.18 |
| Hypothetical | Mesci_2555 | 2.03 | 1.19 |
| Region-γ | |||
| intS | Mesci_2349 | 2.85 | 1.15 |
a Differentially expressed genes (adjusted two-sided P-value of < 0.05) were identified using the DESeq2 package [33]. Since introduced plasmids carried copies of the traI1 and traR ORFs (not including promoter regions), reads mapping to these sequences were of an ambiguous origin and were therefore filtered and removed prior to mapping reads. Differential expression analysis of the traI1 and traI2 untranslated mRNA promoter regions, PtraI1 and PtraI2, was carried out prior to filtering–as these reads were able to be distinguished from plasmid-borne mRNAs. Reads mapping to the plasmid backbones and rRNA genes were removed prior to mapping reads for both analyses.
An alignment of the PtraI1 regions from ICEMlSymR7A and ICEMcSym1271 revealed a tra-box sequence centred 69bp upstream of the ICEMcSym1271 traI1 start codon (S4A Fig). The reads mapping to the traI1 coding sequence were filtered from our RNAseq libraries prior to differential expression analyses (Table 2) because they were also present on the introduced plasmid, however, a secondary comparison of the unfiltered RNAseq reads mapping to the PtraI1 region in our QS+ relative to the QS- WSM1271 cells revealed a sharp 121-fold increase in mapped reads beginning 44bp downstream from tra-box centre and 26bp upstream of the traI1 start codon (Table 2 & S4A Fig).
Homologues of msi172 and msi171 are present on ICEMcSym1271 (Fig 3A) [19] but our initial interrogations did not identify an ICEMlSymR7A traI2 homologue positioned upstream of these genes. traI2 of ICEMlSymR7A appears to encode an AHLsynthase paralogous with TraI1, however, mutation of traI2 has no effect on ICEMlSymR7A excision and no identifiable AHL products are produced by TraI2 [27]. Further inspection of the ICEMcSym1271 msi172-msi171 region revealed the presence of a potential tra-box sequence centred 398bp upstream of the msi172 start codon (S4A Fig). A nucleotide alignment with the corresponding ICEMlSymR7A region revealed this tra-box was also centred 66bp upstream of an internally-truncated traI2 gene remnant (S4A Fig). This traI2 pseudogene overlapped the start codon of msi172 as does traI2 on ICEMlSymR7A (Fig 3A). Interestingly, inspection of traI2-msi172 regions in M. loti USDA 3471 and M. ciceri strains WSM4083, WSM1497, and WSM1284 revealed a similar situation; the traI2 gene in each case was present as a potential protein-coding pseudogene upstream of msi172 and overlapping the msi172 start codon (S5A and S5B Fig). Therefore, although traI2 has likely become a pseudogene on ICEMcSym1271 and other symbiosis ICE/ICE3s, the transcriptional coupling of the tra-box and translational coupling of the TraI2 and Msi172 coding sequences has been maintained. In our RNAseq experiments, traI2, msi172 and msi171 reads were increased ~60-160-fold in QS+ cells (Table 2). A sharp increase in relative read depth was observed at the traI2 promoter 44bp downstream of the tra-box centre and 21bp upstream of the traI2 start codon (S4B Fig) which spanned the entire traI2-msi172-msi171 operon (Fig 3A). The likely transcription start site for traI2 observed from RNAseq reads was consistent with the previously mapped ICEMlSymR7A traI2 promoter (S4B Fig) [27]. Interestingly, comparison of the number of unfiltered transcripts mapping to the traI1 and traI2 promoter regions revealed that QS-induced expression from the traI1 promoter (2196.16 ± [SE] 434.70 TPM) is ~3-fold stronger than that of traI2 (660.88 ± 276.84 TPM) (S4A & S4B Fig). A similar ratio of traI1:traI2 expression is also observed for ICEMlSymR7A [27].
Fig 3. Quorum sensing activation of ICEMcSym1271 promoters.
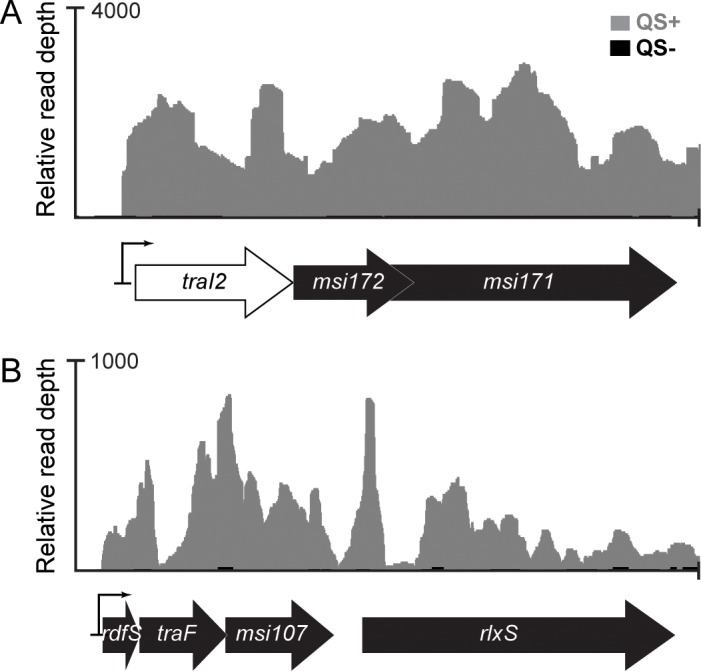
Overlayed relative read coverage (or sequencing depth) plots represent standardised values for the mean number of reads mapped to the positive strand of the regions shown in this figure from the three unfiltered QS+ (grey) and QS- (black) transcriptome libraries of WSM1271. QS+ strains were induced for QS by overexpressing both traI1 and traR1 from the plasmids pPR3-traI1 and pSDz-traR1, respectively, whereas the QS- control strains carried the parent vectors pPR3 and pSDz. The mean read depth for the (A) traI2-msi172-msi171, and (B) rdfS-traF-msi107 and rlxS regions of ICEMcSym1271 in QS- transcriptome libraries were almost non-existent relative to that of the QS+ strain (See S1 Dataset for a full list of TPM values from the filtered reads). A magnified view of reads mapping to the promoter region and the DNA sequence is shown in S4 Fig. These plots were produced using Integrated Genome Browser [73].
For ICEMlSymR7A, FseA stimulates expression from an operon containing rdfS, traF and msi107 [21, 31] (Fig 3B). The same gene cluster is present on ICEMcSym1271 and the RNAseq read depth for the corresponding ICEMcSym1271 homologues was increased 20-58-fold in QS+ cells (Table 2). A distinct read depth increase was observed 25bp upstream of the rdfS start codon corresponding closely with the mapped transcriptional start site for ICEMlSymR7A rdfS (S4C Fig) [31]. In summary, despite several genetic rearrangements, the QS regulon of ICEMcSym1271 appears functionally analogous to that of ICEMlSymR7A and importantly, QS induces the expression of msi172, msi171 and rdfS.
rdfS is required for all three excisive Int-mediated recombination reactions
To explore the involvement of RdfS in ICE3 assembly and excision, a markerless deletion in the WSM1271 rdfS gene was constructed. As expected, no attPS or attBS products were detected in this strain, but interestingly attPG + attBG and attPM + attBM products were also undetectable (Fig 2A). Introduction of rdfS expressed from its native promoter restored attP + attB production at all three sites, albeit at lower levels than wild-type WSM1271. Plasmid-based overexpression of traR1 or msi172-msi171 in the rdfS mutant did not induce excision, however, the same plasmids did induce excision and conjugative transfer in the wild-type WSM1271 (Fig 2C and Table 1). Together these data confirmed that the stimulation and coordination of all three excision reactions by QS and msi172-msi171 is dependent on rdfS.
We hypothesized that RdfS was either directly required to stimulate excisive recombination at attG and attM sites or that RdfS stimulated rdfG and rdfM expression. We overexpressed rdfG and rdfM in the rdfS mutant to see if it would restore the formation of attPG + attBG and attPM + attBM sites, respectively. rdfG was cloned downstream of the strong constitutive nptII promoter and rdfM was cloned downstream of the lac promoter. Interestingly, introduction of lac-driven rdfM resulted in growth arrest even in the absence of IPTG inducer and in the presence of glucose to repress lac expression. This was consistent with our model for excision, in which expression of rdfM alone splits the chromosome and results in loss of viability. Constitutive expression of rdfG in the rdfS mutant resulted in the restored detection of attPG + attBG products in approximately 0.01% of cells (Fig 2A) while the other two sites remained undetectable. In contrast to lac-driven expression, introduction of the cloned copy of rdfM downstream of its native promoter restored the production of attPM + attBM sites in 0.001–0.01% of cells. Therefore, it was clear that attP + attB formation was abolished in the rdfS mutant but RdfS was not directly essential for excisive IntG and IntM recombination. The observation that artificially increased levels of rdfG or rdfM compensated for the loss of rdfS implied RdfG and RdfM expression was abolished in the rdfS mutant.
Overexpression of rdfS stimulates expression of rdfG and rdfM
Inspection of RNAseq data revealed rdfG mRNA abundance was ~2.5-fold higher in QS+ cells (Table 1). rdfM was very weakly expressed in both QS+ and QS- cells and while there was ~2-fold more rdfM reads in QS+ cells, this difference was not statistically significant. To clarify the potential role for RdfS in activation of the rdfG and rdfM promoters, the non-coding regions present upstream of each gene were cloned upstream of the promoterless lacZ gene. Plasmid constructs carrying this fusion were introduced into WSM1271 carrying a constitutively expressed copy of rdfS (Fig 4A). β-galactosidase expression from the rdfG and rdfM promoters was induced ~4.5 and ~8-fold respectively in the presence of constitutively expressed rdfS. Consistent with RNAseq data, rdfM expression was much lower than rdfG expression and almost undetectable in the absence of rdfS. To discount the possibility that RdfS induced expression indirectly through other factors on ICEMcSym1271, the same set of experiments were repeated using the heterologous M. loti R7ANS background, which lacks all ICE genes (Fig 4B). These assays produced comparable results to those carried out in WSM1271, supporting the hypothesis that the transcriptional activation of rdfG and rdfM promoters by RdfS was likely direct.
Fig 4. Transcriptional regulation of rdfG and rdfM by RdfS.

β-galactosidase assays [67] were performed for (A) WSM1271 and (B) R7ANS carrying either control vector pPR3 or pPR3-rdfS (constitutively expressing rdfS) together with one of three RDF promoter-lacZ fusion constructs cloned into the pSDz vector. Assays were performed with six biological replicates and mean β-galactosidase activity values (Relative Fluorescent Units/s/OD600) were compared by Bonferroni adjusted student’s t-tests. SD is denoted by error bars.
Discussion
Excision and circularization is an essential prerequisite for conjugative transfer of ICEs. Integrase proteins of ICEs and temperate phages generally catalyse both the excision and integration reactions, but integrative recombination is generally favoured in the absence of a cognate RDF [25]. Unlike most ICEs that excise following a single Int-mediated recombination, ICEMcSym1271 requires three Int-mediated reactions to excise [19]. Here, we demonstrated that three ICEMcSym1271-encoded RDFs RdfG, RdfM, and RdfS are required for the ICEMcSym1271 excisive IntG, IntM, and IntS-mediated recombination reactions, respectively. We also demonstrated that overexpression of the QS sensors TraR1 and TraR2 or autoinducer synthase TraI1 in WSM1271 simultaneously increased the proportion of cells in a population undergoing all three ICEMcSym1271 excision reactions 10-100-fold. QS significantly induced mRNA abundance for the WSM1271 traI1, traI2-msi172-msi171, rdfS, and rdfG genes, as well as those for conjugative pilus formation [23, 27, 31]. In addition to stimulating the ICEMcSym1271 IntS-mediated excisive recombination, RdfS was shown to transcriptionally activate the rdfG and rdfM genes. Therefore, RdfS acts as the master regulator for ICEMcSym1271 excision.
Our model for the assembly and excision of ICEMcSym1271 indicates that if the first excisive reaction is catalysed by IntM, then the chromosome is split into two inviable parts (Fig 1A). However, transcription of rdfM and rdfG is dependent on RdfS, and thus excisive IntS-mediated recombination probably occurs prior to that of IntM and IntG in WSM1271 cells induced for ICEMcSym1271 assembly an excision. This hierarchical genetic regulation of the three RDFs has likely evolved to minimise the potential for formation of the non-viable split chromosome configuration following spurious rdfM expression. In wild-type WSM1271 or QS-induced WSM1271 cells, the frequency attPM + attBM site formation was also significantly less than either that of attPG + attBG and attPS + attBS, as was expression of rdfM relative to rdfG and rdfS. Moreover, introduction of a plasmid-borne copy of rdfM under the control of the relatively weak lacI promoter on pSacB [34] resulted in arrested growth of 1271ΔrdfS cells suggesting that even a low level of RdfM expression in the absence of RdfS and RdfG is deleterious. It is possible that the rdfM promoter, in addition to evolving transcriptional dependency on RdfS, has evolved to promote only subtle levels of rdfM expression to further reduce the likelihood of the formation of a non-viable chromosomal state. Considering the data, it seems probable that the in situ excisive recombination pathway of ICEMcSym1271 follows the sequence IntS > IntG > IntM (Fig 1A).
RDFs have diverse roles in the control of MGE transfer. Several bacteriophage excisionases act as both RDFs and transcriptional regulators [35–44]. Phage-P2 Cox and the coliphage-186 Apl excisionases bind and bend attP and attL DNA to promote prophage excision, but they also stimulate induction of the lytic cycle by blocking transcription of repressor genes cl and c, respectively [35–43]. The Cox protein additionally stimulates derepression of neighbouring P4 prophages by activating transcription from the late P4-phage promoter [39, 45]. Cox-bound promoter and attP regions each contain six or more repeats of a “cox-box” consensus sequence that may vary in direction or percentage identity between different binding targets, and may be bound with variable affinity [38, 39, 42]. A protein sharing structural homology with excisionases has recently been shown to be essential for relaxasome processing of the conjugative plasmid pIP501 [46]. These examples and our findings here emphasise that RDFs/excisionases have evolved differential and evolutionarily flexible roles in the control of MGE dissemination.
The RdfS proteins of R7A and WSM1271 are almost identical at the amino-acid level apart from the extreme C-terminus (S1 Fig). Therefore, it is possible that the rdfG and rdfM promoter regions could have evolved DNA-binding targets that respond to RdfS, rather than RdfS having evolved specific new functions associated with ICE3. We were unable to identify any clearly conserved DNA sequence motifs on attLS, attPS or the rdfG or rdfM promoter regions. However, excisionase binding sites are often poorly conserved at the DNA-sequence level and for most the mode of site recognition is not well understood. Most characterized RDFs have a winged-helix-turn-helix structure that contacts both major and minor DNA grooves, therefore overall DNA topology is believed to be especially critical for recognition [47]. Given that RdfS presumably binds multiple distinct sites on ICEMcSym1271, RdfS presents itself as an enticing research focus for gaining a deeper understanding of excisionase-DNA recognition characteristics and the multifaceted roles of excisionases in stimulating horizontal transfer of diverse MGE.
ICEMcSym1271-α carries two functional QS-sensor genes, traR1 and traR2. Sequence comparisons of the ICEMlSymR7A and ICEMcSym1271 QS loci suggest that the ICEMcSym1271-derived TraR2 protein is the more immediate orthologue of R7A-derived TraR. Broader comparisons of the QS loci organisation between these ICEs suggest that each ICE may have evolved from an ancestral ICE carrying two complete sets of traR-traI loci (S6 Fig). The DNA sequence upstream of traI1 on ICEMlSymR7A lacks a traR1 homologue but does contain sequence homologous to the 3’-end of traR1 from ICEMcSym1271, suggesting deletion of an ancestral copy of traR1 has occurred in R7A. The traI2 gene on ICEMcSym1271 appears to have become a pseudogene with several internal truncations, but a truncated seemingly nonsense open-reading-frame remains that has retained both its position relative to the upstream tra box and translational overlap with msi172, as is the case on other related ICEs (S5 Fig). On ICEMlSymR7A, traI2 is a complete and potentially functional gene, but ICEMlSymR7A excision or transfer is unaffected for a markerless deletion traI2 mutant, suggesting it too may be in the early stages of pseudogenisation.
For both ICEMcSym1271 and ICEMlSymR7A the functional AHL-synthase traI1 and the apparent traI2 pseudogene that is translationally coupled to msi172-msi171 are proceeded by a tra-box sequence allowing for transcriptional control by TraR. ICEMlSymR7A is exquisitely sensitive to overexpression of msi172-msi171 or rdfS, which cause growth inhibition and loss of ICEMlSymR7A respectively [23, 31, 48]. In the presence of AHLs, expression of traI2-msi172-msi171 in R7A is lower than that observed for traI1 [27]. Our RNAseq data similarly indicates that that expression from the ICEMcSym1271 traI1 promoter is stronger than from the traI2-msi172-msi171 promoter (Table 2, S4A and S4B Fig). As previously speculated [27], this separation of QS-activated genes involved in stimulation of excision (msi172-msi171) and AHL-production (traI1) has likely facilitated independent adjustment of expression levels from each QS-activated promoter. This type of genetic uncoupling of AHL synthase genes from other QS-activated genes could in some instances explain the presence of orphan–or solo—QS regulators and AHL-synthase genes frequently identified throughout gram-negative bacteria [49, 50].
ICE3s are a novel and unexpected form of MGE and the three-integrase system seemingly introduces considerable unnecessary complexity. However, in this work we show that the activity of RdfS as a master regulator of ICE3 excision greatly simplifies the pathway to excision. With RdfS in control, the excisive recombination reactions are induced in a predetermined order to excise ICEMcSym1271. As previously discussed [19, 26], despite the complex arrangement of integrase att sites, the formation of the prototype ICE3 may have occurred following only two chromosomal inversions between three single-part ICEs or non-conjugative integrating elements. We also suspect that the regulatory control of RdfS over rdfG and rdfM transcription could have pre-existed ICE3 on these ancestral single-part constituents. Several putative symbiosis ICEs carry rdfS but lack an associated IntS gene and instead carry a unique integrase and distinct attL site within one of five serine tRNA genes (Mesorhizobium spp. strains CC1192 [51]; WSM3873 (NZ_LYTM00000000.1), AA23 (NZ_LYTP00000000.1) and WSM3859 (NZ_NSGG00000000.1)). Moreover, numerous more distantly related putative ICEs in the α-proteobacteria carry a homologue of rdfS but lack an obvious intS homologue [28]. The conservation of rdfS but lack of conservation of intS on these ICEs suggests that RdfS homologues may be able stimulate excisive recombination through interactions with multiple distinct recombination systems. With this view in mind, the evolution of ICE3 and capture of unique ICE genes [26] potentially involves recombination between groups of distinct ICE3, ICEs and non-conjugative integrative elements that already share common regulatory control elements. In summary, this work provides substantial insight into the molecular control and evolution of these complex tripartite elements.
Materials and methods
Bacteria, plasmids, and growth conditions
Strains and plasmids are listed in Table 3. Strains were cultured as previously described [23, 27, 29, 52, 53]. Allelic replacement, and markerless deletion mutants were constructed using double crossover homologous recombination as previously described [23]. Plasmids for construction of mutants are described in Table 3 and primers used are listed in S1 Table. Construction of plasmids is detailed in Supplementary materials and methods (S1 File).
Table 3. Bacterial strains and plasmids.
| Strain | a Relevant Characteristics | Reference |
|---|---|---|
|
Escherichia coli DH10B |
F- endA1 deoR+ recA1 galE15 galK16 nupG rpsL Δ(lac)X74 φ80lacZΔM15 araD139 Δ(ara,leu)7697 mcrA Δ(mrr-hsdRMS-mcrBC) StrR λ– | Invitrogen |
| ST18 | S17 Δpir ΔhemA | [54] |
| Chromobacterium violaceum | ||
| CV026 | Biosensor strain for detection of C4-C8 N-acyl-homoserine lactones | [29] |
| Mesorhizobium ciceri | ||
| WSM1271 | Bisserula pelecinus symbiont, harbours ICEMcSym1271 (accession NC_014923.1) | [55] |
| 1271ΔrdfG::ΩaadA | WSM1271 rdfG ΩaadA replacement mutant | This study |
| 1271ΔrdfM::ΩaadA | WSM1271 rdfM ΩaadA replacement mutant | This study |
| 1271ΔrdfS | WSM1271 rdfS in frame deletion mutant | This study |
| M. loti | ||
| R7ANS | Symbiosis ICE cured derivative of M. loti R7A | [23] |
| Plasmids | ||
| pJQ200 SK | Suicide vector in Mesorhizobium, contains sacB, GmR | [56] |
| pEX18Tc | Suicide vector in Mesorhizobium, contains sacB, TcR | [57] |
| pHP45Ω | Insertional inactivation vector carrying an ΩaadA1 cassette, SmR, SpR | [58] |
| pJET-aadA | pJET 1.2 carrying the ΩaadA cassette from pHP45Ω amplified using primers 34 & 35, SmR, SpR, ApR | This study |
| pJQΩrdfG | pJQ200 SK carrying the ΩaadA cassette from pHP45Ω flanked by regions upstream and downstream of rdfG amplified using primers 1, 2 & 3, 4, respectively, used to create 1271ΔrdfG::ΩaadA, SmR, SpR GmR | This study |
| pJQΩrdfM | pJQ200 SK carrying the ΩaadA cassette from pHP45Ω flanked by regions upstream and downstream of rdfM amplified using primers 5, 6 & 7, 8, respectively, used to create 1271ΔrdfM::ΩaadA, SmR, SpR GmR | This study |
| pEXΔrdfS | pEX18Tc carrying regions flanking intS amplified using primers 9, 10 & 11, 12 respectively, used to create WSM1271ΔrdfS, TcR | This study |
| pJP2 | Stable (contains Par region), low copy number BHR IncP vector, TcR | [59] |
| pJP2-rdfG | pJP2 carrying rdfG from WSM1271 amplified using primers 13 & 14, TcR | This study |
| pJP2-rdfM | pJP2 carrying rdfM from WSM1271 amplified using primers 15 & 16, TcR | This study |
| pJP2-rdfS | pJP2 carrying rdfS from WSM1271 amplified using primers 17 & 18, TcR | This study |
| pPR3 | pPROBE-KT carrying the nptII promoter from pFAJ1708, NmR | [60–62] |
| pPR3-rdfG | pPR3 carrying rdfG from WSM1271 amplified using primers 19 & 20, NmR | This study |
| pPR3-traI1 | pPR3 carrying traI1 from WSM1271 amplified using primers 21 & 22, NmR | This study |
| pSacB | BHR vector carrying inducible IPTG promoter and sacB gene, NmR | [19] |
| pSacB-rdfM | pSacB carrying rdfM from WSM1271 amplified using primers 23 & 16, NmR | This study |
| pSDz | BHR plasmid, carries IPTG inducible promoter and promoterless lacZ, TcR | [31] |
| pSDz-traR1 | pSDz carrying tra1R from WSM1271 amplified using primers 24 & 25, TcR | This study |
| pSDz-traR2 | pSDz carrying traR2 from WSM1271 amplified using primers 36 & 37, TcR | This study |
| pSDz-msi172171 | pSDz carrying msi172-msi171 from WSM1271 amplified using primers 26 & 27, TcR | This study |
| pSDz-PrdfG | pSDz carrying the rdfG promoter from WSM1271 amplified using primers 28 & 29, TcR | This study |
| pSDz-PrdfM | pSDz carrying the rdfM promoter from WSM1271 amplified using primers 30 & 31, TcR | This study |
| pSDz-PrdfS | pSDz carrying the rdfS promoter from WSM1271 amplified using primers 32 & 33, TcR | This study |
| pSDzPtraI1-lacZ | pSDz carrying the traI promoter from WSM1271 amplified using primers 38 & 39, TcR | This study |
| pSDz-traR1PtraI1-lacZ | pSDz-traR1 carrying the traI promoter from WSM1271 amplified using primers 38 & 39, TcR | This study |
| pSDz-traR2PtraI1-lacZ | pSDz-traR2 carrying the traI promoter from WSM1271 amplified using primers 38 & 39, TcR | This study |
| pTHQP-1 | Standard construct for qPCR assays for ICE3 excision, GmR | [19] |
| pJET 1.2. | Commercial blunt cloning vector, ApR | Thermo Fisher Scientific |
a Abbreviation for antibiotic resistances are as follows; ApR, ampicillin; GmR, gentamycin; NmR, neomycin; SpR, spectinomycin; SmR, streptomycin; TcR, tetracycline. See S1 Table for primer details.
Molecular techniques, assays, and bioinformatics
DNA extractions, purifications, electrophoresis and PCR were carried out as previously described [18, 23]. Sanger sequencing was performed by the Australian Genome Research Facility. Nucleotide and amino acid alignments were performed using the T-Coffee multiple sequence aligner [63]. Protein secondary structures were predicted using Jpred(v4) [64]. Synteny comparisons were performed using the Artemis Comparison Tool [65] and plotted with genoplotR [66]. β-galactosidase assays were performed as previously described with three to six biological replicates per treatment [31, 67]. Mesorhizobium conjugation experiments were performed as previously described [19]. CV026 bioassays were performed on E. coli strains by streaking them adjacent to CV026 on LB agar and plates and incubating these plates for 24 h at 28°C [29]. CV026 well-diffusion bioassays were performed on M. loti strains as previously described [28, 29]. All cloning was carried out in E. coli DH10B and constructs were chemically transformed [68] into E. coli ST18 for mobilisation into Mesorhizobium spp. via biparental mating [54].
qPCR assays for ICE3 excision
Genomic DNA for qPCR was extracted from 64-h TY broth cultures as previously described [19]. Our previously validated qPCR assay [19] was used to measure the percentage of chromosomes carrying each individual attB (attBG, attBM, and attBS) and corresponding attP (attPG, attPM, and attPS) site in samples of genomic DNA extracted from WSM1271 cultures. This was achieved by comparing the standardised relative abundance of each attP and attB site to the chromosomal gene melR. Primer sites for the qPCR assay are shown in Fig 1, and described in S1 Table.
RNA-Sequencing and statistical analysis
TY broth cultures (OD600 0.8–1.0) were grown for RNAseq experiments as previously described [69] with three biological repetitions per treatment and two technical repetitions per sample. Total RNA was isolated as previously described [70]. RNA quality and concentration was analysed at various points throughout processing using Experion StdSense or HighSens analysis kit assays (Bio-Rad Technologies). DNA was removed from approximately 3 μg of total RNA using the TURBO DNA-free kit (Invitrogen) and confirmed using a Qubit fluorometer dsDNA BR assay. rRNA was depleted from total RNA using a Ribo-Zero rRNA magnetic kit (Illumina) and resulting RNA was purified using a RNA Clean & Concentrator (Zymo Research). Barcoded cDNA libraries were prepared from rRNA depleted RNA samples using Ion Total RNA-Seq kit v2 (Thermo Fisher). Each barcoded cDNA library was diluted in DEPC treated milliQ water to a final concentration of 75 pM and templates for sequencing were prepared using an Ion Chef instrument (Thermo Fisher). Sequencing was performed using the Ion Proton system (Thermo Fisher). Read sets from technical repetitions were combined. Adapter sequences were removed using nesoni clip (http://www.vicbioinformatics.com/software.nesoni.shtml). To reduce any potential rRNA/total-RNA abundance biases introduced during rRNA depletion, reads mapping to rRNA genes were removed using FastQ Screen (https://www.bioinformatics.babraham.ac.uk). Reads were mapped to the WSM1271 genome (accession NC_014923) using Bowtie 2 [71] and visualised using Artemis [72] or Integrated Genome Browser [73]. For gene expression analysis, read sets were additionally filtered to remove sequences matching plasmids pPR3-traI1 and pSDz-traR1 prior to mapping. An average (per biological replicate) of 14 million (standard deviation (SD) = 3.3 million) QS+ and 8.5 million (SD = 1.5 million) QS- post-filter reads were mapped to WSM1271 with 96.7–98.6% alignment rate. Read counts for gene features were performed using HTSeq [74] with default settings then imported into DESeq2 [33] for identification of differentially expressed genes (S1 Dataset).
To measure expression from the traI1 and traI2 promoter regions, the unfiltered reads were mapped to the WSM1271 chromosome using the procedures described above, and read counting was performed using the—nonunique all function on HTSeq so that reads mapping ambiguously to the traI1 and traI2 regions and ORFs were counted for both features.
Supporting information
Secondary structures were predicted using Jpred(v4) [64]. α-helices are highlighted in yellow, β-sheets are highlighted in blue. All three proteins carry a predicted two stranded MerR-family winged helix-turn-helix motif characteristic of RDFs [25].
(TIF)
The C. violaceum CV026 biosensor strain [29] was used to detect the production of AHLs in (A) E. coli DH10B or (B) M. loti R7ANS either constitutively expressing ICEMcSym1271-derived traI1 from the plasmid pPR3-traI1, or carry the empty vector pPR3. Production of a purple violacein halo indicated production of C4-C8 AHLs.
(TIF)
β-galactosidase assays [67] were performed on a set of R7ANS strains carrying the same traI1 promoter-lacZ fusion on either pSDz, pSDz-traR1, or pSDz-traR2. These strains were induced for expression of traR1/traR2 with 1 μM IPTG, and also carried either a constitutively expressed copy of traI1 (pPR3-traI1), or the empty vector pPR3. Assays were performed with three biological replicates and mean β-galactosidase activity values (Relative Fluorescent Units/s/OD600) were compared by Bonferroni adjusted student’s t-tests. SD is denoted by error bars.
(TIF)
The promoter regions of traI1 (A), traI2 (B), and rdfS genes (C) from WSM1271 were identified based on similarity with homologous regions in R7A. Nucleotide alignments were performed using the T-Coffee multiple sequence aligner [63]. Transcriptional start sites for R7A genes previously mapped by 5’RACE are shown in bold [27, 31]. Relative read depth (or sequencing depth) plots represent a standardised value for the mean number of reads mapped to the positive strand of the regions shown in this figure from the three unfiltered QS+ transcriptome libraries of WSM1271.These plots were produced using Integrated Genome Browser [73]. QS+ strains were induced for QS by overexpressing both traI1 and traR1 from the plasmids pPR3-traI1 and pSDz-traR1, respectively. Mean values of 2196.16 ± (SD) 434.70 TPM unfiltered reads and 660.88 ± 276.84 TPM unfiltered reads were mapped to the non-coding regions between the transcriptional start sites and start codons for traI1 and traI2, respectively. A students t-test revealed that this difference was significant (P = 0.01).
(TIF)
(A) The nucleotide sequence of traI2 promoters and (B) the TraI2 amino acid sequences from six Mesorhizobium strains were aligned using the T-coffee multiple sequence aligner [63].
(TIF)
On ICEMlSymR7A, traR is encoded upstream of an operon encoding the likely non-functional AHL-synthase gene traI2, msi172-msi171 and qseM-qseC. The functional AHL synthase TraI1 is encoded at a separate location. ICEMcSym1271 carries traR2 upstream of qseM-qseC, however, the traI2-msi172-msi171 region has been translocated to a different position and traI2 has become internally truncated. ICEMlSym1271 carries a second traR gene traR1 paired with the traI1 gene. It is likely that ICEMlSymR7A originally had a traR1 gene that has subsequently been deleted. Consistent with this notion, the 100-bp upstream of traI1 closely resembles the 3’-end of traR1. Thus, it seems likely that an ancestral ICE carried an operon comprising traR2-traI2-msi172-msi171 upstream of divergent qseC and qseM genes and a second QS locus containing traR1-traI1. Synteny comparisons were performed using the Artemis Comparison Tool [65] and plotted with genoplotR [66].
(TIF)
(PDF)
(XLSX)
(DOCX)
Acknowledgments
The authors thank Dr John Sullivan for editorial assistance.
Data Availability
Raw feature counts for filtered and unfiltered RNA-Seq libraries mapped to the WSM1271 chromosome (See Materials and methods for details), and DESeq2 ouput for differential expression analyses are available on the NCBI Gene Expression Omnibus database (https://www.ncbi.nlm.nih.gov/geo/) under the accession GSE108732. The remaining relevant data are within the paper and its Supporting Information files.
Funding Statement
TLH, JJT and GWO acknowledge support from the Grains Research and Development Corporation of Australia (https://grdc.com.au/) [GRS10939; UMU00040]. JPR is the recipient of an Australian Research Council Future Fellowship [Project ID FT170100235] funded by the Australian Government (http://www.arc.gov.au/grants). The collaboration between TLH, JJT and PSP is supported by the Sir Walter Murdoch Adjunct Professor Scheme. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Boto L. Horizontal gene transfer in evolution: facts and challenges. Proc R Soc B. 2010;277(1683):819–27. doi: 10.1098/rspb.2009.1679 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Rankin DJ, Rocha EPC, Brown SP. What traits are carried on mobile genetic elements, and why? Heredity. 2011;106(1):1–10. doi: 10.1038/hdy.2010.24 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Frost LS, Leplae R, Summers AO, Toussaint A. Mobile genetic elements: the agents of open source evolution. Nat Rev Microbiol. 2005;3(9):722–32. doi: 10.1038/nrmicro1235 [DOI] [PubMed] [Google Scholar]
- 4.Delavat F, Miyazaki R, Carraro N, Pradervand N, van der Meer JR. The hidden life of integrative and conjugative elements. FEMS Microbiol Rev. 2017;41(4):512–37. doi: 10.1093/femsre/fux008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Schmidt H, Hensel M. Pathogenicity islands in bacterial pathogenesis. Clin Microbiol Rev. 2004;17(1):14–56. doi: 10.1128/CMR.17.1.14-56.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hochhut B, Lotfi Y, Mazel D, Faruque SM, Woodgate R, Waldor MK. Molecular analysis of antibiotic resistance gene clusters in Vibrio cholerae O139 and O1 SXT constins. Antimicrob Agents Chemother. 2001;45(11):2991–3000. doi: 10.1128/AAC.45.11.2991-3000.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Dobrindt U, Hochhut B, Hentschel U, Hacker J. Genomic islands in pathogenic and environmental microorganisms. Nat Rev Microbiol. 2004;2(5):414–24. doi: 10.1038/nrmicro884 [DOI] [PubMed] [Google Scholar]
- 8.Sullivan JT, Ronson CW. Evolution of rhizobia by acquisition of a 500-kb symbiosis island that integrates into a phe-tRNA gene. PNAS. 1998;95(9):5145–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Juhas M, van der Meer JR, Gaillard M, Harding RM, Hood DW, Crook DW. Genomic islands: tools of bacterial horizontal gene transfer and evolution. FEMS Microbiol Rev. 2009;33(2):376–93. doi: 10.1111/j.1574-6976.2008.00136.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ravatn R, Studer S, Springael D, Zehnder AJ, van der Meer JR. Chromosomal integration, tandem amplification, and deamplification in Pseudomonas putida F1 of a 105-kilobase genetic element containing the chlorocatechol degradative genes from Pseudomonas sp. Strain B13. J Bacteriol. 1998;180(17):4360–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Guglielmini J, Quintais L, Garcillan-Barcia MP, de la Cruz F, Rocha EP. The repertoire of ICE in prokaryotes underscores the unity, diversity, and ubiquity of conjugation. PLoS Genet. 2011;7(8):e1002222 doi: 10.1371/journal.pgen.1002222 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Coluzzi C, Guédon G, Devignes M- D, Ambroset C, Loux V, Lacroix T, et al. A glimpse into the world of integrative and mobilizable elements in streptococci reveals an unexpected diversity and novel families of mobilization proteins. Front Microbiol. 2017;8:443 doi: 10.3389/fmicb.2017.00443 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ramsay JP, Kwong SM, Murphy RJ, Yui Eto K, Price KJ, Nguyen QT, et al. An updated view of plasmid conjugation and mobilization in Staphylococcus. Mob Genet Elements. 2016;6(4):e1208317 doi: 10.1080/2159256X.2016.1208317 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Carraro N, Rivard N, Burrus V, Ceccarelli D. Mobilizable genomic islands, different strategies for the dissemination of multidrug resistance and other adaptive traits. Mob Genet Elements. 2017;7(2):1–6. doi: 10.1080/2159256X.2017.1304193 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Johnson CM, Grossman AD. Integrative and conjugative elements (ICEs): what they do and how they work. Annu Rev Genet. 2015;49:577–601. doi: 10.1146/annurev-genet-112414-055018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wozniak RA, Waldor MK. Integrative and conjugative elements: mosaic mobile genetic elements enabling dynamic lateral gene flow. Nat Rev Microbiol. 2010;8(8):552–63. doi: 10.1038/nrmicro2382 [DOI] [PubMed] [Google Scholar]
- 17.Carraro N, Burrus V. The dualistic nature of integrative and conjugative elements. Mob Genet Elements. 2015;5(6):98–102. doi: 10.1080/2159256X.2015.1102796 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sullivan JT, Patrick HN, Lowther WL, Scott DB, Ronson CW. Nodulating strains of Rhizobium loti arise through chromosomal symbiotic gene transfer in the environment. PNAS. 1995;92(19):8985–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Haskett TL, Terpolilli JJ, Bekuma A, O'Hara GW, Sullivan JT, Wang P, et al. Assembly and transfer of tripartite integrative and conjugative genetic elements. PNAS. 2016;113(43):12268–73. doi: 10.1073/pnas.1613358113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sullivan JT, Brown SD, Yocum RR, Ronson CW. The bio operon on the acquired symbiosis island of Mesorhizobium sp. strain R7A includes a novel gene involved in pimeloyl-CoA synthesis. Microbiology (Reading, England). 2001;147(Pt 5):1315–22. [DOI] [PubMed] [Google Scholar]
- 21.Sullivan JT, Trzebiatowski JR, Cruickshank RW, Gouzy J, Brown SD, Elliot RM, et al. Comparative sequence analysis of the symbiosis island of Mesorhizobium loti strain R7A. J Bacteriol. 2002;184(11):3086–95. doi: 10.1128/JB.184.11.3086-3095.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kasai-Maita H, Hirakawa H, Nakamura Y, Kaneko T, Miki K, Maruya J, et al. Commonalities and differences among symbiosis islands of three Mesorhizobium loti strains. Microbes and environments. 2013;28(2):275–8. doi: 10.1264/jsme2.ME12201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ramsay JP, Sullivan JT, Stuart GS, Lamont IL, Ronson CW. Excision and transfer of the Mesorhizobium loti R7A symbiosis island requires an integrase IntS, a novel recombination directionality factor RdfS, and a putative relaxase RlxS. Mol Microbiol. 2006;62(3):723–34. doi: 10.1111/j.1365-2958.2006.05396.x [DOI] [PubMed] [Google Scholar]
- 24.Seah NE, Warren D, Tong W, Laxmikanthan G, Van Duyne GD, Landy A. Nucleoprotein architectures regulating the directionality of viral integration and excision. PNAS. 2014;111(34):12372–7. doi: 10.1073/pnas.1413019111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lewis JA, Hatfull GF. Control of directionality in integrase-mediated recombination: examination of recombination directionality factors (RDFs) including Xis and Cox proteins. Nucleic Acids Res. 2001;29(11):2205–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Haskett TL, Ramsay JP, Bekuma AA, Sullivan JT, O'Hara GW, Terpolilli JJ. Evolutionary persistence of tripartite integrative and conjugative elements. Plasmid. 2017;92:30–6. doi: 10.1016/j.plasmid.2017.06.001 [DOI] [PubMed] [Google Scholar]
- 27.Ramsay JP, Sullivan JT, Jambari N, Ortori CA, Heeb S, Williams P, et al. A LuxRI-family regulatory system controls excision and transfer of the Mesorhizobium loti strain R7A symbiosis island by activating expression of two conserved hypothetical genes. Mol Microbiol. 2009;73(6):1141–55. doi: 10.1111/j.1365-2958.2009.06843.x [DOI] [PubMed] [Google Scholar]
- 28.Ramsay JP, Major AS, Komarovsky VM, Sullivan JT, Dy RL, Hynes MF, et al. A widely conserved molecular switch controls quorum sensing and symbiosis island transfer in Mesorhizobium loti through expression of a novel antiactivator. Mol Microbiol. 2013;87(1):1–13. doi: 10.1111/mmi.12079 [DOI] [PubMed] [Google Scholar]
- 29.McClean KH, Winson MK, Fish L, Taylor A, Chhabra SR, Camara M, et al. Quorum sensing and Chromobacterium violaceum: exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology (Reading, England). 1997;143 (Pt 12):3703–11. [DOI] [PubMed] [Google Scholar]
- 30.Ramsay JP, Ronson CW. Genetic Regulation of Symbiosis Island Transfer in Mesorhizobium loti In: de Bruijn FJ, editor. Biological Nitrogen Fixation. 1: John Wiley & Sons, Inc; 2015. p. 217–24. [Google Scholar]
- 31.Ramsay JP, Tester LGL, Major AS, Sullivan JT, Edgar CD, Kleffmann T, et al. Ribosomal frameshifting and dual-target antiactivation restrict quorum-sensing–activated transfer of a mobile genetic element. PNAS. 2015;112(13):4104–9. doi: 10.1073/pnas.1501574112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Yang M, Sun K, Zhou L, Yang R, Zhong Z, Zhu J. Functional analysis of three AHL autoinducer synthase genes in Mesorhizobium loti reveals the important role of quorum sensing in symbiotic nodulation. Can J Microbiol. 2009;55:210+. doi: 10.1139/w08-128 [DOI] [PubMed] [Google Scholar]
- 33.Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014;15(12):550 doi: 10.1186/s13059-014-0550-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Khan SR, Gaines J, Roop RM, Farrand SK. Broad-host-range expression vectors with tightly regulated promoters and their use to examine the influence of TraR and TraM expression on Ti plasmid quorum sensing. Appl Environ Microbiol. 2008;74(16):5053–62. doi: 10.1128/AEM.01098-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lundqvist B, Bertani G. Immunity repressor of bacteriophage P2. Identification and DNA-binding activity. J Mol Biol. 1984;178(3):629–51. [DOI] [PubMed] [Google Scholar]
- 36.Dodd IB, Kalionis B, Egan JB. Control of gene expression in the temperate coliphage 186. VIII. Control of lysis and lysogeny by a transcriptional switch involving face-to-face promoters. J Mol Biol. 1990;214(1):27–37. doi: 10.1016/0022-2836(90)90144-B [DOI] [PubMed] [Google Scholar]
- 37.Reed MR, Shearwin KE, Pell LM, Egan JB. The dual role of Apl in prophage induction of coliphage 186. Mol Microbiol. 1997;23(4):669–81. [DOI] [PubMed] [Google Scholar]
- 38.Saha S, Haggard-Ljungquist E, Nordstrom K. The cox protein of bacteriophage P2 inhibits the formation of the repressor protein and autoregulates the early operon. EMBO J. 1987;6(10):3191–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Saha S, Haggård-Ljungquist E, Nordström K. Activation of prophage P4 by the P2 Cox protein and the sites of action of the Cox protein on the two phage genomes. PNAS. 1989;86(11):3973–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Yu A, Haggard-Ljungquist E. The Cox protein is a modulator of directionality in bacteriophage P2 site-specific recombination. J Bacteriol. 1993;175(24):7848–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Esposito D, Wilson JC, Scocca JJ. Reciprocal regulation of the early promoter region of bacteriophage HP1 by the Cox and Cl proteins. Virology. 1997;234(2):267–76. [DOI] [PubMed] [Google Scholar]
- 42.Ahlgren-Berg A, Cardoso-Palacios C, Eriksson JM, Mandali S, Sehlén W, Sylwan L, et al. A comparative analysis of the bifunctional Cox proteins of two heteroimmune P2-like phages with different host integration sites. Virology. 2009;385(2):303–12. doi: 10.1016/j.virol.2008.12.002 [DOI] [PubMed] [Google Scholar]
- 43.Dodd IB, Reed MR, Egan JB. The Cro-like Apl repressor of coliphage 186 is required for prophage excision and binds near the phage attachment site. Mol Microbiol. 1993;10(5):1139–50. [DOI] [PubMed] [Google Scholar]
- 44.Piazzolla D, Cali S, Spoldi E, Forti F, Sala C, Magnoni F, et al. Expression of phage P4 integrase is regulated negatively by both Int and Vis. J Gen Virol. 2006;87(Pt 8):2423–31. doi: 10.1099/vir.0.81875-0 [DOI] [PubMed] [Google Scholar]
- 45.Six EW, Lindqvist BH. Mutual derepression in the P2-P4 bacteriophage system. Virology. 1978;87(2):217–30. [DOI] [PubMed] [Google Scholar]
- 46.Goessweiner-Mohr N, Eder M, Hofer G, Fercher C, Arends K, Birner-Gruenberger R, et al. Structure of the double-stranded DNA-binding type IV secretion protein TraN from Enterococcus. Acta Crystallogr D. 2014;70(Pt 9):2376–89. doi: 10.1107/S1399004714014187 [DOI] [PubMed] [Google Scholar]
- 47.Abbani M, Iwahara M, Clubb RT. The structure of the excisionase (Xis) protein from conjugative transposon Tn916 provides insights into the regulation of heterobivalent tyrosine recombinases. J Mol Biol. 2005;347(1):11–25. doi: 10.1016/j.jmb.2005.01.019 [DOI] [PubMed] [Google Scholar]
- 48.Ramsay JP, Ronson CW. Silencing quorum sensing and ICE mobility through antiactivation and ribosomal frameshifting. Mob Genet Elements. 2015;5(6):103–8. doi: 10.1080/2159256X.2015.1107177 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Patel HK, Suárez-Moreno ZR, Degrassi G, Subramoni S, González JF, Venturi V. Bacterial LuxR solos have evolved to respond to different molecules including signals from plants. Front Plant Sci. 2013;4:447 doi: 10.3389/fpls.2013.00447 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Patankar AV, Gonzalez JE. Orphan LuxR regulators of quorum sensing. FEMS Microbiol Rev. 2009;33(4):739–56. doi: 10.1111/j.1574-6976.2009.00163.x [DOI] [PubMed] [Google Scholar]
- 51.Haskett T, Wang P, Ramsay J, O'Hara G, Reeve W, Howieson J, et al. Complete Genome Sequence of Mesorhizobium ciceri Strain CC1192, an Efficient Nitrogen-Fixing Microsymbiont of Cicer arietinum. Genome Announc. 2016;4(3) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Ronson CW, Nixon BT, Albright LM, Ausubel FM. Rhizobium meliloti ntrA (rpoN) gene is required for diverse metabolic functions. J Bacteriol. 1987;169(6):2424–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Beringer JE. R factor transfer in Rhizobium leguminosarum. J Gen Microbiol. 1974;84(1):188–98. doi: 10.1099/00221287-84-1-188 [DOI] [PubMed] [Google Scholar]
- 54.Thoma S, Schobert M. An improved Escherichia coli donor strain for diparental mating. FEMS Microbiol Lett. 2009;294(2):127–32. [DOI] [PubMed] [Google Scholar]
- 55.Nandasena K, Yates R, Tiwari R, O'Hara G, Howieson J, Ninawi M, et al. Complete genome sequence of Mesorhizobium ciceri bv. biserrulae type strain (WSM1271(T)). Stand Genomic Sci. 2014;9(3):462–72. doi: 10.4056/sigs.4458283 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Quandt J, Hynes MF. Versatile suicide vectors which allow direct selection for gene replacement in gram-negative bacteria. Gene. 1993;127(1):15–21. [DOI] [PubMed] [Google Scholar]
- 57.Hoang TT, Karkhoff-Schweizer RR, Kutchma AJ, Schweizer HP. A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: application for isolation of unmarked Pseudomonas aeruginosa mutants. Gene. 1998;212(1):77–86. [DOI] [PubMed] [Google Scholar]
- 58.Prentki P, Krisch HM. In vitro insertional mutagenesis with a selectable DNA fragment. Gene. 1984;29(3):303–13. [DOI] [PubMed] [Google Scholar]
- 59.Prell J, Boesten B, Poole P, Priefer UB. The Rhizobium leguminosarum bv. viciae VF39 gamma-aminobutyrate (GABA) aminotransferase gene (gabT) is induced by GABA and highly expressed in bacteroids. Microbiology (Reading, England). 2002;148(Pt 2):615–23. [DOI] [PubMed] [Google Scholar]
- 60.Rodpothong P, Sullivan JT, Songsrirote K, Sumpton D, Cheung KW, Thomas-Oates J, et al. Nodulation gene mutants of Mesorhizobium loti R7A-nodZ and nolL mutants have host-specific phenotypes on Lotus spp. Mol Plant Microbe Interact. 2009;22(12):1546–54. doi: 10.1094/MPMI-22-12-1546 [DOI] [PubMed] [Google Scholar]
- 61.Miller WG, Leveau JH, Lindow SE. Improved gfp and inaZ broad-host-range promoter-probe vectors. Mol Plant Microbe Interact. 2000;13(11):1243–50. doi: 10.1094/MPMI.2000.13.11.1243 [DOI] [PubMed] [Google Scholar]
- 62.Dombrecht B, Vanderleyden J, Michiels J. Stable RK2-derived cloning vectors for the analysis of gene expression and gene function in gram-negative bacteria. Mol Plant Microbe Interact. 2001;14(3):426–30. doi: 10.1094/MPMI.2001.14.3.426 [DOI] [PubMed] [Google Scholar]
- 63.Notredame C, Higgins DG, Heringa J. T-Coffee: A novel method for fast and accurate multiple sequence alignment. J Mol Biol. 2000;302(1):205–17. doi: 10.1006/jmbi.2000.4042 [DOI] [PubMed] [Google Scholar]
- 64.Drozdetskiy A, Cole C, Procter J, Barton GJ. JPred4: a protein secondary structure prediction server. Nucleic Acids Res. 2015;43(W1):W389–W94. doi: 10.1093/nar/gkv332 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Carver TJ, Rutherford KM, Berriman M, Rajandream MA, Barrell BG, Parkhill J. ACT: the Artemis Comparison Tool. Bioinformatics. 2005;21(16):3422–3. doi: 10.1093/bioinformatics/bti553 [DOI] [PubMed] [Google Scholar]
- 66.Guy L, Roat Kultima J, Andersson SGE. genoPlotR: comparative gene and genome visualization in R. Bioinformatics. 2010;26(18):2334–5. doi: 10.1093/bioinformatics/btq413 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Ramsay J. High-throughput β-galactosidase and β-glucuronidase assays using fluorogenic substrates. Bio-Protocol. 2013;3(14):e827. [Google Scholar]
- 68.Li X, Sui X, Zhang Y, Sun Y, Zhao Y, Zhai Y, et al. An improved calcium chloride method preparation and transformation of competent cells. Afr J Biotechnol. 2010;9(50):8549–54. [Google Scholar]
- 69.Fox MA, Karunakaran R, Leonard ME, Mouhsine B, Williams A, East AK, et al. Characterization of the quaternary amine transporters of Rhizobium leguminosarum bv. viciae 3841. FEMS Microbiol Lett. 2008;287(2):212–20. doi: 10.1111/j.1574-6968.2008.01307.x [DOI] [PubMed] [Google Scholar]
- 70.Karunakaran R, Ebert K, Harvey S, Leonard ME, Ramachandran V, Poole PS. Thiamine is synthesized by a salvage pathway in Rhizobium leguminosarum bv. viciae strain 3841. J Bacteriol. 2006;188(18):6661–8. doi: 10.1128/JB.00641-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Langmead B, Salzberg SL. Fast gapped-read alignment with Bowtie 2. Nat Methods. 2012;9(4):357–9. doi: 10.1038/nmeth.1923 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Rutherford K, Parkhill J, Crook J, Horsnell T, Rice P, Rajandream M-A, et al. Artemis: sequence visualization and annotation. Bioinformatics. 2000;16(10):944–5. [DOI] [PubMed] [Google Scholar]
- 73.Freese NH, Norris DC, Loraine AE. Integrated genome browser: visual analytics platform for genomics. Bioinformatics. 2016;32(14):2089–95. doi: 10.1093/bioinformatics/btw069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Anders S, Pyl PT, Huber W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics. 2015;31 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Secondary structures were predicted using Jpred(v4) [64]. α-helices are highlighted in yellow, β-sheets are highlighted in blue. All three proteins carry a predicted two stranded MerR-family winged helix-turn-helix motif characteristic of RDFs [25].
(TIF)
The C. violaceum CV026 biosensor strain [29] was used to detect the production of AHLs in (A) E. coli DH10B or (B) M. loti R7ANS either constitutively expressing ICEMcSym1271-derived traI1 from the plasmid pPR3-traI1, or carry the empty vector pPR3. Production of a purple violacein halo indicated production of C4-C8 AHLs.
(TIF)
β-galactosidase assays [67] were performed on a set of R7ANS strains carrying the same traI1 promoter-lacZ fusion on either pSDz, pSDz-traR1, or pSDz-traR2. These strains were induced for expression of traR1/traR2 with 1 μM IPTG, and also carried either a constitutively expressed copy of traI1 (pPR3-traI1), or the empty vector pPR3. Assays were performed with three biological replicates and mean β-galactosidase activity values (Relative Fluorescent Units/s/OD600) were compared by Bonferroni adjusted student’s t-tests. SD is denoted by error bars.
(TIF)
The promoter regions of traI1 (A), traI2 (B), and rdfS genes (C) from WSM1271 were identified based on similarity with homologous regions in R7A. Nucleotide alignments were performed using the T-Coffee multiple sequence aligner [63]. Transcriptional start sites for R7A genes previously mapped by 5’RACE are shown in bold [27, 31]. Relative read depth (or sequencing depth) plots represent a standardised value for the mean number of reads mapped to the positive strand of the regions shown in this figure from the three unfiltered QS+ transcriptome libraries of WSM1271.These plots were produced using Integrated Genome Browser [73]. QS+ strains were induced for QS by overexpressing both traI1 and traR1 from the plasmids pPR3-traI1 and pSDz-traR1, respectively. Mean values of 2196.16 ± (SD) 434.70 TPM unfiltered reads and 660.88 ± 276.84 TPM unfiltered reads were mapped to the non-coding regions between the transcriptional start sites and start codons for traI1 and traI2, respectively. A students t-test revealed that this difference was significant (P = 0.01).
(TIF)
(A) The nucleotide sequence of traI2 promoters and (B) the TraI2 amino acid sequences from six Mesorhizobium strains were aligned using the T-coffee multiple sequence aligner [63].
(TIF)
On ICEMlSymR7A, traR is encoded upstream of an operon encoding the likely non-functional AHL-synthase gene traI2, msi172-msi171 and qseM-qseC. The functional AHL synthase TraI1 is encoded at a separate location. ICEMcSym1271 carries traR2 upstream of qseM-qseC, however, the traI2-msi172-msi171 region has been translocated to a different position and traI2 has become internally truncated. ICEMlSym1271 carries a second traR gene traR1 paired with the traI1 gene. It is likely that ICEMlSymR7A originally had a traR1 gene that has subsequently been deleted. Consistent with this notion, the 100-bp upstream of traI1 closely resembles the 3’-end of traR1. Thus, it seems likely that an ancestral ICE carried an operon comprising traR2-traI2-msi172-msi171 upstream of divergent qseC and qseM genes and a second QS locus containing traR1-traI1. Synteny comparisons were performed using the Artemis Comparison Tool [65] and plotted with genoplotR [66].
(TIF)
(PDF)
(XLSX)
(DOCX)
Data Availability Statement
Raw feature counts for filtered and unfiltered RNA-Seq libraries mapped to the WSM1271 chromosome (See Materials and methods for details), and DESeq2 ouput for differential expression analyses are available on the NCBI Gene Expression Omnibus database (https://www.ncbi.nlm.nih.gov/geo/) under the accession GSE108732. The remaining relevant data are within the paper and its Supporting Information files.