

Abstract
Purpose of review
The intestine is highly efficient at absorbing and packaging dietary lipids onto the structural protein apoB48 for distribution throughout the body. Here, we summarize recent advances into understanding the physiological and pharmacological actions of the proglucagon-derived peptides: glucagon like peptide 1 (GLP-1) and glucagon like peptide 2 (GLP-2) on intestinal lipoprotein secretion.
Recent findings
Several recent studies have elucidated mechanisms underlying the paradoxical effects of GLP-1 and GLP-2 on intestinal production of triglyceride-rich lipoproteins (TRLs). Both gut-derived peptides are secreted on an equimolar basis in response to the same nutrient stimulus. Despite neither receptor demonstrating clear localization to enterocytes, a single injection of a GLP-1R agonist rapidly decreases delivery of intestinally packaged fatty acids into the plasma, while conversely GLP-2 receptor (GLP-2R) activation acutely increases TRL concentrations in plasma.
Summary
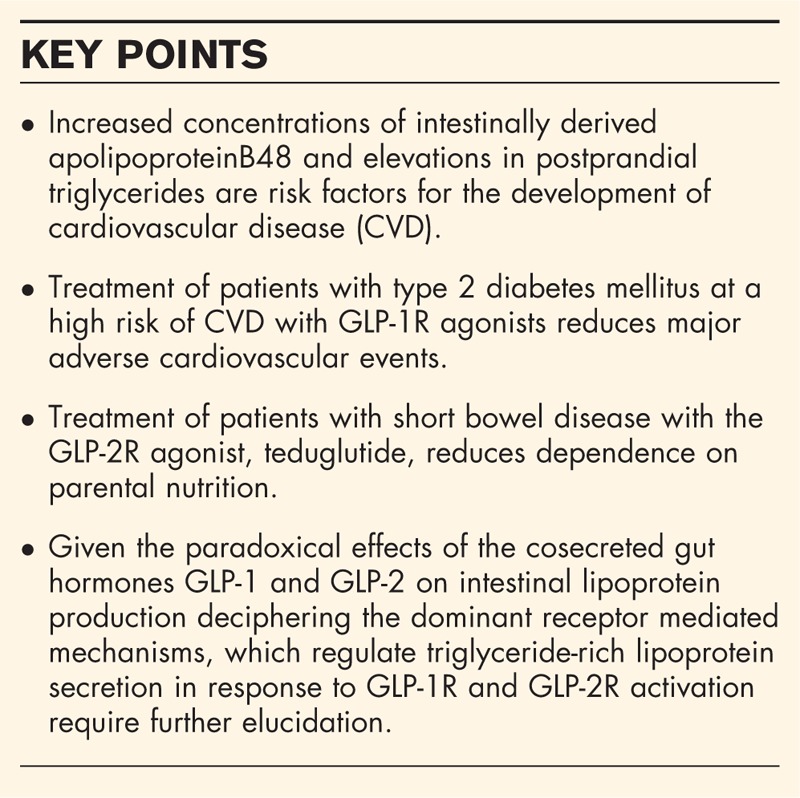
The regulation of TRL secretion is dependent on the coordination of many processes: fatty acid availability uptake, assembly onto the apoB48 polypeptide backbone, secretion and reuptake, which the hormonal, neural, inflammatory and metabolic milieu can all strongly influence. Understanding of how GLP-1 and GLP-2 receptor agonists control TRL production has clinical importance given that GLP1R agonists were recently demonstrated not only to provide glycemic control but also to prevent major adverse cardiovascular events in patients with T2DM and the success of GLP-2R agonists in treating short bowel disease.
Keywords: glucagon like peptide 1, glucagon like peptide 2, intestine, lipoprotein, postprandial
INTRODUCTION
Patients with type 2 diabetes mellitus (T2DM) are at an increased risk of death from cardiovascular disease (CVD) [1,2]. The particulars of the disturbed metabolic milieu responsible for the increase in CVD risk are complex. Intensive glucose lowering has been definitively linked to improved microvascular health; however, a clear benefit on macrovascular vessel health has been elusive [3–5]. The atherogenic, dyslipidemia in T2DM is characterized by an overproduction and/or delayed catabolism of triglyceride-rich particles (TRLs), including apolipoprotein (apoB)48-containing chylomicrons and apoB100-containing VLDLs, cholesterol-rich remnant particles, small dense LDLs and a reduction in circulating HDLs [6]. LDL-cholesterol lowering with statins is a mainstay in the management of dyslipidemia in patients with T2DM [7]. However, even when cholesterol targets are met, the risk of CVD is still significant. Prospective clinical trials have demonstrated that hypertriglyceridemia is an additional independent risk factor for the development of CVD [8,9] and the extent of coronary atherosclerosis in patients with T2DM is positively correlated with plasma concentrations of TRL [10]. Nonfasting triglyceride concentrations are correlated with cholesterol-rich remnants and when elevated are associated with an increased risk of myocardial infarction (MI) and death [9]. Results from the Improved Reduction of Outcomes: Vytorin Efficacy International Trial (IMPROVE IT) have demonstrated that the addition of ezetimibe to statin treatment offered further benefit in the reduction of major adverse cardiovascular events (MACEs) [11]. Also, recent studies have demonstrated that apoB48 levels better predict increased adiposity than traditional lipid risk factors [12▪], suggesting it may also be a harbinger of a disturbed metabolic milieu. Therefore, therapies that target both the atherogenic dyslipidemia and hyperglycemia observed in patients with T2DM are of significant interest for potential CVD risk reduction.
Glucagon like peptide -1 (GLP-1) is a gut-derived peptide hormone, which is an exquisite regulator of glucose-dependent insulin secretion. GLP-1, together with the incretin hormone glucose-dependent insulinotropic polypeptide (GIP), is responsible for up to 70% of nutrient-stimulated insulin secretion and therefore indirectly facilitates postprandial glucose clearance [13]. Activation of the β-cell GLP-1 receptor (GLP-1R) has well described effects, including potentiation of glucose stimulated insulin secretion, increased synthesis of insulin, prevention of β-cell apoptosis and stimulation of somatostatin contributing to a reduction in glucagon secretion [14–16]. However, levels of GLP-1 in circulation are very low, owing to degradation by the serine protease dipeptidyl peptidase 4 (DPP4) [17]. Sustained activation of the GLP-1 receptor (GLP-1R) can be achieved through long-acting agonists resistant to cleavage by DPP4 or through compounds which inhibit DPP4 activity [18]. In addition to the sodium glucose cotransporter 2 (SGLT2) inhibitor empagliflozin, two GLP-1R agonists, liraglutide and semaglutide, have demonstrated a decrease in MACE in patients with T2DM [19,20▪▪,21▪▪]. Therefore, in addition to its significant effects on the endocrine pancreas, the less prominent but potentially equally important postprandial lipid-lowering aspects of GLP-1 physiology may improve the postprandial dyslipidemia associated with T2DM and reduce the risk for CVD [22]. However, understanding the physiological role of GLP-1 on intestinal lipoprotein secretion and how it may be disturbed in T2DM is complicated due to the cosecretion on an equimolar basis with glucagon like peptide 2 (GLP-2). In contrast to the reduction in postprandial TRL observed with induction of GLP-1 signalling, GLP-2 receptor (GLP-2R) activation has been demonstrated to promote lipid absorption [23]. In this review, we focus on new developments in both the physiological and pharmacological regulation of TRL metabolism in response to treatment with GLP-1 and GLP-2.
Box 1.
no caption available
REGULATION OF INTESTINAL-DERIVED TRIGLYCERIDE-RICH LIPOPROTEIN SECRETION
Unlike the liver, which can control its intake of lipid by regulating receptor-mediated endocytosis, the intestine must act in a dynamic fashion at the extremes of nutrient exposure [24,25]. Luminal fat content is a key signal for TRL production but represents only one of the many hormone and nutrition signals, which are integrated to control the process [26]. Within the intestinal lumen, fats are hydrolyzed and associate with bile acids to be taken up by both passive and active transport [27]. Upon entry, the fatty acids (FAs) can be re-esterified and incorporated from the endoplasmic reticulum (ER) into cytosolic lipids droplets or loaded onto the apoB48 polypeptide, the obligate scaffold for the assembly of dietary lipids through the actions of microsomal triglyceride transfer protein (MTP). These triglyceride -rich particles undergo maturation and association with other and secreted from the enterocyte into lymphatic lacteal. Intestinal lymphatics within the villi are the main route of entry for dietary lipid into the bloodstream. Chylomicrons are too large to cross endothelial barriers and have been described to enter the lacteal through both paracellular transport in endothelial vesicles and transcytosis through open junctions [28]. The transport process is initiated minutes after a meal and continues for hours. The lymphatics drain from all sections of the small bowel to mesenteric and celiac nodes and onto intestinal lymph trunks and the venous circulation through the thoracic duct [29]. This routing bypasses the liver, making absorbed nutrients immediately available to all tissues.
SECRETION OF PREPROGLUCAGON PEPTIDES
Preproglucagon (Gcg)+ enteroendocrine cells compose approximately 1% of the intestinal epithelium [30] and are dispersed throughout the duodenum but more enriched in the ileum in rats and pigs and distal colon in mice [31]. Single cell sequencing has determined that Gcg+ cells can be classified into three overlapping but distinct hormone-secreting populations [32▪▪]. Activation of a number of nutrient receptors residing on the luminal side of the L-cell stimulate release of stored peptide hormones in secretory vesicles at the basolateral membrane in close proximity to capillaries and nerve endings [33,34]. In healthy participants, fasting plasma concentrations of GLP-1 are within the 5–10 pmol/l range, which elevates to approximately 50 pmol/l after ingestion of a mixed meal in blood [35,36]. Interestingly, recent work has suggested that FA sensing by L-cells may also occur basolaterally in a process involving postlipoprotein lipase (LPL)-mediated hydrolysis of formed chylomicrons [37▪▪]. These data support previous studies whereby disruption of chylomicron synthesis by treatment with a surfactant reduced GLP-1 secretion [38].
PRECLINICAL STUDIES EVALUATING PHYSIOLOGICAL AND PHARMACOLOGICAL CONCENTRATIONS OF GLUCAGON LIKE PEPTIDE-1 ON INTESTINAL LIPOPROTEIN METABOLISM
Despite being pharmacologically exploited for its potent effects on the islet, GLP-1 is best described as an intestinal signal peptide that works as an elegant messenger during the postprandial period to fine-tune endocrine responses to nutrients. GLP-1 has been characterized to inhibit gastric acid secretion [39], reduce gastric emptying [40,41], slow gastric motility [42], reduce lymph flow [43], relax the gastric fundus [44], inhibit antro-pyloroduodenal motility [45] and act as an ‘ileal break’ [46]. Both elevations in endogenous GLP-1 and pharmacological concentrations reached with GLP-1R agonists have demonstrated significant reductions in postprandial lipoprotein secretion [47,48]. Alternatively, Glp1r−/− mice demonstrate an increase in triglyceride-rich, apoB48-containing lipoproteins postfat load when compared with littermate controls. Similarly, administration of the GLP-1R antagonist exendin 9–39 (50 nmol/kg) to hamsters increased apoB48 mass in the plasma TRL fraction thus supporting a role for endogenous or physiological GLP-1 signaling in regulating triglyceride secretion [47].
GLP-1R expression has been reported in mouse and human bowel [49–51] and treatment of intestinal segments with exendin-4 reduce apoB48 secretion to the media [47]. Flow cytometry analysis of the absorptive layer of the intestine has demonstrated that the GLP-1R is not expressed in Villin+ enterocytes but instead is localized to intraepithelial lymphocytes [52]. Consistent with its effects throughout the alimentary canal, the GLP-1R has also been localized in mice using in-situ hybridization to gastric parietal cells in the glandular stomach, Brunners glands and the nerve plexus throughout the small and large bowel [53▪▪].
In the absence of a direct signalling mechanism in enterocytes to regulate postprandial lipid handling, a number of secondary pathways have been investigated. Acute treatment of mice with exendin-4 (24 nmol/kg) decreases the rate of appearance of triglyceride and apoB48 into plasma [47], suggesting while the chronic GLP-1R-mediated anorectic effects or weight loss may contribute clinically to improved lipid profiles, they are not the exclusive driver of reduced lipoprotein secretion (Fig. 1a). Also, the lowering of triglyceride and apoB48 production in response to exendin-4 persisted even when oil was given 1 h prior to treatment to allow FA to exit the stomach and enter the small bowel, therefore eliminating inhibition of antral contractility and reduced gastric emptying as the main mechanism for decreasing TRL-production (Fig. 1a). As GLP-1R agonists increase meal-stimulated insulin secretion, a significant regulator of postprandial lipoprotein secretion [54], analysis of plasma insulin levels and the relative effects of these concentrations on intestinal lipoprotein secretion determined that the reduction in the number of TRL cannot solely be explained by increased insulin secretion [47] (Fig. 1a).
FIGURE 1.
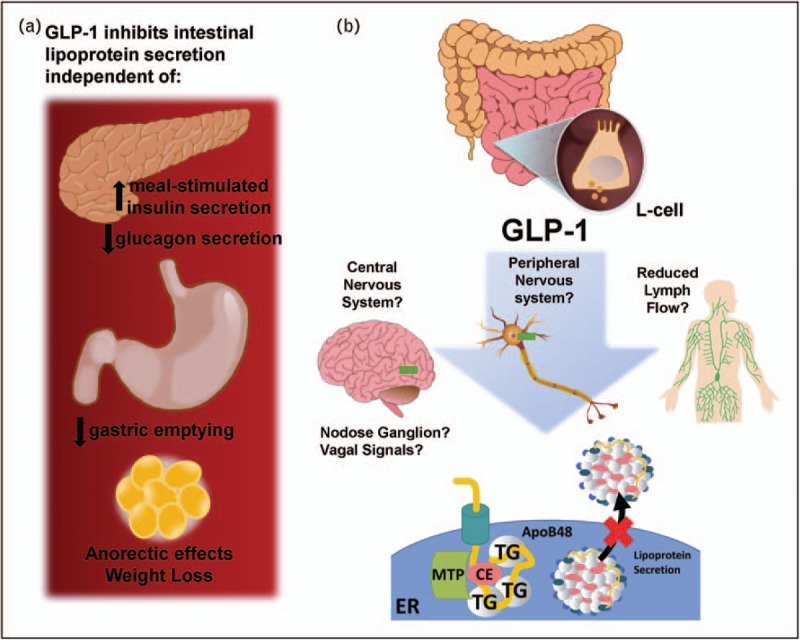
(a) GLP-1R mediated decrease in the production rate of intestinally derived, triglyceride-rich lipoproteins is independent of increased insulin secretion and reduced glucagon secretion, reduced gastric emptying and weight loss or anorectic effects. (b) Potential mechanisms in need of validation for GLP-1R mediated reductions in TRL-TG and TRL- apoB48 synthesis and secretion including signalling through the peripheral gut:brain axis, lymph flow and the central nervous system.
The intestine has been proposed to play a role in control of lymph flow [28]. Initial lymphatics lack the ability to generate tone due to an absence of smooth muscle cells and therefore are dependent on the peristaltic motion of the intestinal wall to propel flow. During the transition from a fasted to fed state, lymph flow in the intestine is normally increased, particularly in response to ingested fat [55]. The concentrations of GLP-1 in lymph have been reported to significantly exceed those in plasma [56] and injection of recombinant GLP-1 (20 pmol/kg/min) into lipid-infused rats demonstrated a 50% decrease in the rate of lymph flow [43]. Studies in cannulated rats that received an intraduodenal infusion with 3H-triolein demonstrated a significant decrease in the recovery of 3H-triolein in the lymph suggesting a decrease in entry of lipid into the lymphatics with GLP-1 treatment (20 pmol/kg/min) [43]. An increase in label was found within the mucosa layer and not the lumen suggesting uptake and esterification of FA into triglyceride within enterocytes was not disturbed [43]. In accordance with this, evaluation of feces from Glp1r−/− mice or mice treated with the GLP-1R agonist taspoglutide demonstrated no significant increase in lipid content [57,58]. Exendin-4 treatment has been reported to result in a modest (∼20%) reduction in MTP activity but did not affect MTP protein levels. These data were associated with reduced Oil red O staining in Villi of the jejunum 2 h after oil gavage [59]. Therefore, it is clear GLP-1 has significant effects on lipid handling within the intestine but interpreting the pathways involved requires further study. GLP-1R activation also has a described role in both the small and large intestine to mediate mucosal expansion [49]; however, what role secondary mediators such as fibroblast growth factor 7, (Fgf7) have in lipid handling and lipoprotein secretion remain unclear (Fig. 1b).
Studies in rats have demonstrated that intestinal transit is sensitive to treatment with native GLP-1; however, concentrations of GLP-1 greatly exceeding physiological concentrations do not affect the contractility of gastrointestinal muscle strips suggesting an indirect and potential neuronal effect to reduce intestinal transit time [60]. GLP-1R are distributed throughout the brain and peripherally in the nodose ganglion [61,62]. Injection of exendin-4 into the ventricles of the brain reduces intestinal lipoprotein secretion in hamsters. However, intracerebroventricular (i.c.v.) injections of exendin 9–39 did not prevent the inhibition of TRL production by exendin-4 suggesting a potential combination of peripheral and neuronal pathways [59]. The plasticity observed in lipoprotein regulation is consistent with recent studies evaluating the GLP-1R+ circuits within the brain that control glucose and body weight. Deletion of the Glp1r in the hypothalamus, proopiomelanocortin neurons,or paraventricular nucleus did not individually disrupt GLP-1R feeding and glucose responses, although clearly each population can contribute [63▪▪].
CLINICAL STUDIES EVALUATING GLUCAGON LIKE PEPTIDE-1 RECEPTOR AGONISTS AND INTESTINAL LIPOPROTEIN METABOLISM
Collectively, placebo-controlled clinical trials with GLP-1R agonists as a monotherapy have illustrated a consistent 0.2–0.3 mmol reduction in triglyceride concentrations [22]. Importantly, many of the mechanistic insights provided from preclinical studies have been reproduced in clinical settings. Kinetic studies in healthy men in which a primed constant infusion of deuterated leucine was administered under pancreatic clamp conditions in the fed state have demonstrated that insulin acutely suppresses apoB48 production rates in the postprandial state (47–62%) [54]. In-vivo support for GLP-1R activation to reduce TRL production independent of increased insulin secretion and reduced gastric emptying came from similar kinetic study protocols undertaken in which healthy participants were administrated a fixed high fat, mixed macronutrient liquid meal through a nasoduodenal tube. Under these experimental conditions, acute treatment with the GLP-1R agonist, exenatide, was still able to significantly reduce postprandial apoB48 production [64].
Patients with impaired glucose tolerance or recent onset T2DM injected with exenatide [10 μg, subcutaneous (sc)] demonstrated a marked decrease in serum concentrations of triglyceride, remnant-triglyceride and apoC3 [65]. A null mutation (R19X) discovered in APOC3, in which carriers express half the amount of protein, is associated with lower fasting and postprandial triglyceride (1–6 h) postfat load [66] and apoC3-enriched TRLS have a slower rate of triglyceride hydrolysis particularly when LPL was bound to GPIHBP1 [67▪▪]. Therefore, chronic changes in apoC3 do have the potential to influence lipoprotein secretion. However, kinetic studies have not to date demonstrated a reduction in fractional catabolic rate of TRL-particles and analysis of patients treated for 1 year with exenatide (10 μg twice daily) versus insulin glargine have demonstrated significant reductions in triglyceride, free fatty acids (FFA), HDL-cholesterol, VLDL-cholesterol, and apoB48; however, no differences were noted in apoB100, apoA1, apoA2 or importantly apoC3 [67▪▪].
PRECLINICAL STUDIES EVALUATING THE PHYSIOLOGICAL AND PHARMACOLOGICAL EFFECTS OF GLUCAGON LIKE PEPTIDE-2 ON INTESTINAL LIPOPROTEIN SECRETION
Initial studies characterizing the effects of treatment of female CD-1 mice with synthetic, rat GLP-2 (2.5 μg sc, twice daily for 10 days) demonstrated an increase in intestinal lipid absorption as GLP-2 enhanced plasma triolein levels [68]. Consistent with this acute, intravenous treatment of chow-fed Syrian golden hamsters (GLP-2, 20 pmol/kg body weight/min) after an oral fat load increased plasma levels of radiolabelled triolein and apoB48 [69]. Studies in Glp2r−/− mice and experiments utilizing GLP-2 (3–33), an antagonist of the GLP-2R [70], which can also exert weak agonist activity [71], have demonstrated that endogenous GLP-2R activation regulates mucosal expansion in response to refeeding; however, a direct effect on lipoprotein action has not been elucidated. Interestingly, high fat diet-fed CD-1 mice treated with GLP-2(3–33) had no change in lipid content in faeces, suggesting that inhibition of endogenous GLP-2 action does not disturb lipid absorption [72▪].
Like GLP-1, GLP-2 signals through a G-protein coupled receptor, which has been localized within the gastrointestinal tract and found to be enriched within the jejunum [73,74▪]. GLP-2Rs have also been associated in intestinal compartments rich in enteric neurons [75]. Consistent with these studies, recent in-situ hybridization analysis in mice has determined that GLP-2R expression is negligible in absorptive epithelial cells and abundant in the nerve plexus throughout the gut [53▪▪]. In addition, GLP-2Rs were localized to the muscle layers of the nonglandular stomach, lamina propria of the mucosal layer, in the circular and longitudinal muscles of the duodenum [53▪▪].
Fast performance liquid chromatography analysis of plasma lipoproteins isolated after a fat load demonstrated an enrichment in the chylomicron remnant/VLDL range in hamsters administered GLP-2 (1–33) (0.25 mg/kg) and also in wildtype mice treated with the GLP-2 analogue (hGly2-GLP-2, 0.25 mg/kg) compared with controls [76]. Mechanistically, the increase in TRL was demonstrated to be due to a three-fold increase in intestinal lipoprotein production. In support of increased production, radiolabelling experiments in which primary hamster jejunal segments were treated with GLP-2 demonstrated an increase in newly synthesized apoB48 into the media and these data were not due to an overall increase in protein synthesis or increased enterocyte survival [76]. Treatment of hamsters with GLP-2 accelerated the rate of incorporation of gavaged radiolabelled triolein into plasma TG. Consistent with increased FA uptake, acute GLP-2 treatment increased levels of glycosylated CD36 in hamster-derived enterocytes and increased localization to the subapical domain of enterocyte villi. Further experiments in mice lacking CD36 (Cd36−/−), demonstrated that treatment with hGly2-GLP-2 (0.25 mg/kg) failed to increase luminal triolein uptake and increase plasma TRL-TG and apoB48 [76]. It is clear that CD36 facilitates the increase in FA uptake in response to GLP-2R signalling; however, what signalling pathway GLP-2 exploits to increase glycosylation and activity of the CD36 transmembrane receptor remains undetermined (Fig. 2).
FIGURE 2.
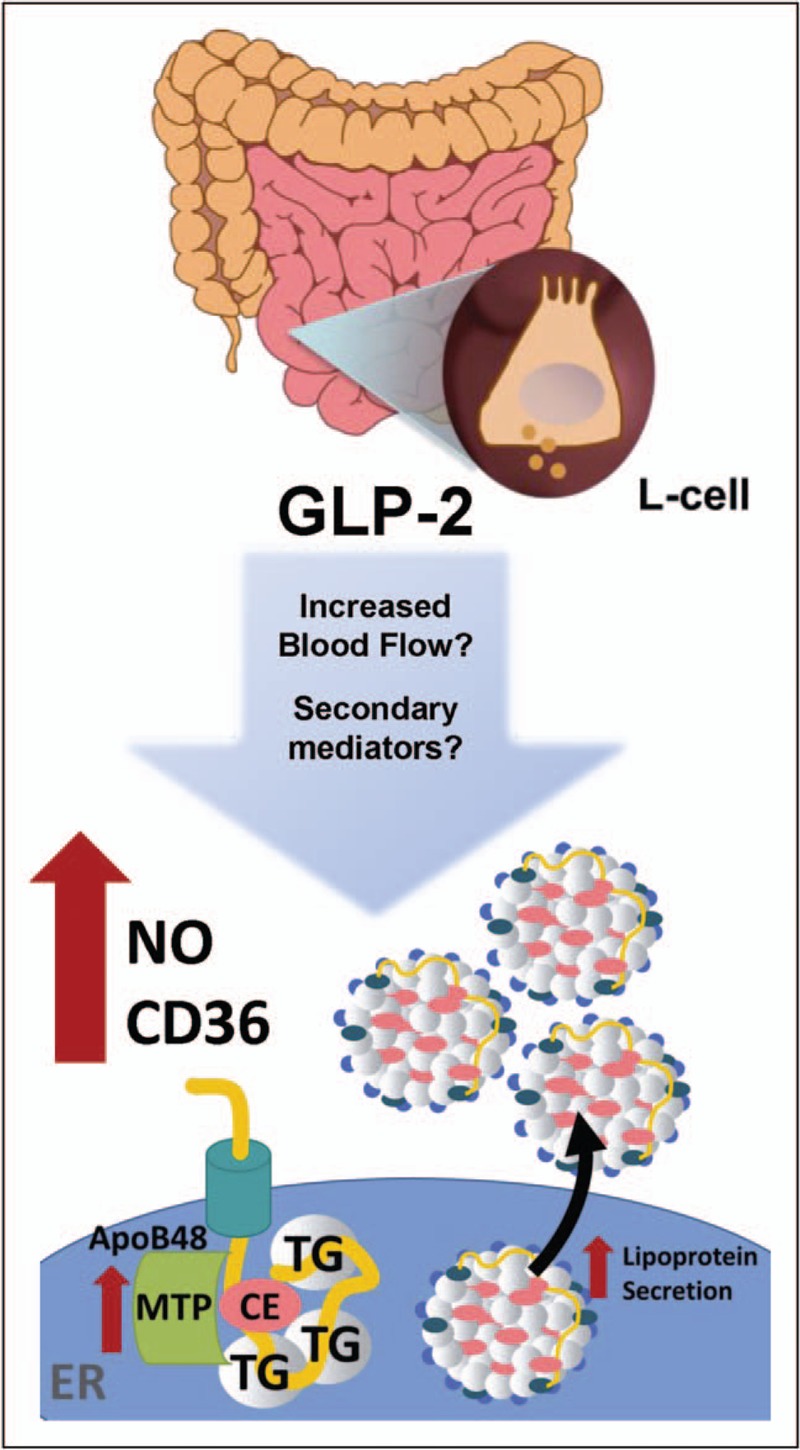
Emerging evidence suggests that GLP-2R activation increases the secretion of preformed chylomicron particles. Potential mechanisms in need of validation for GLP-2R mediated increases in TRL-TG and TRL-apoB48 synthesis and secretion including increased blood flow and secondary mediators.
In 12-day-old piglets fed by parental nutrition, human GLP-2 (500 pmol/kg/h) increased portal blood flow, which was inhibited by NG-Nitro-L-arginine methyl ester (L-NAME, 50 μmol/kg/h) suggesting a nitric oxide dependent mechanism [77]. Nitric oxide is the product of nitric oxide synthase (NOS), which is freely permeable and can easily diffuse through biological membranes [78]. Therefore, as nitric oxide is a known mediator of GLP-2 action on increasing mesenteric blood flow, its role in lipoprotein secretion was further investigated. Consistent with a role in the regulation of lipoprotein secretion by GLP-2, genetic elimination of eNOS resulted in fewer TRLs and triglyceride accumulation in the jejunum, which was unable to be rescued by intraperitoneal (i.p.) injection of Gly2-GLP-2 [79]. In addition, a significant increase in MTP activity was described in hamsters 90 min after GLP-2 (0.25 mg/kg) treatment and this effect was completely abrogated by treatment with L-NAME [79] (Fig. 2).
Not surprisingly similar to GLP-1, significant concentrations of GLP-2 appear within intestinal lymph in response to oral administration of a range of FAs [80]; however, the significance of elevating endogenous concentrations remains to be determined. Also, many secondary mediators have been identified as targets of GLP-2 action, including the growth factors: keratinocyte growth factor, insulin-like growth factors, epidermal growth factor and related ligands in the ErbB family [23]; however, it remains to be determined what role if any these secondary mediators have in GLP-2 mediated effects on lipoprotein secretion.
CLINICAL STUDIES EVALUATING GLUCAGON LIKE PEPTIDE-2 ACTION ON INTESTINAL LIPOPROTEIN SECRETION
GLP-2 (2 pmol/kg/min) treatment of healthy male participants fed a low-calorie content meal significantly increased postprandial plasma triglyceride and FFA concentrations [81]. Administration of a single, pharmacological dose of native GLP-2 (1500 μg subcutaneously) during a high-fat, macronutrient-fixed formula administered in a nasoduodenal tube resulted in a rapid increase in TRL-apoB48 and TRL-TG with peak concentrations of plasma triglyceride and apoB48 being reached within 1 h after administration. Further studies using retinyl palmitate to label lipid incorporation in triglyceride-rich lipoproteins 7 h prior to treatment with GLP-2 clearly demonstrated an acute increase in retinyl palmitate labelled TRL triglyceride and apoB-48 concentrations suggesting GLP-2 mediates the release of preformed chylomicron particles [82]. Studies in healthy, obese males regarding the endogenous contribution of GLP-1 and GLP-2 demonstrate that significant variation occurs in the postprandial TRL fraction and variation in GLP-1 and GLP-2 concentrations did not account for this disparity [83]. However, similar to the kinetic studies discussed above, GLP-2 levels significantly correlated with the rapid elevation in area under the curve for both TRL-TG concentrations and apoB48 [83].
CONCLUSION
GLP-1 and GLP-2 are secreted in equimolar concentrations in response to nutrients; clearly, they both have significant effects on lipoprotein secretion, however, deciphering the precise mechanisms has been difficult. Both have their biological activity regulated through cleavage by the serine protease DPP4, GLP-1 has a half-life of 1–2 min [84], while GLP-2 cleavage is much slower at 7 min [85]. Many factors challenge our interpretation of how GLP-1 and GLP-2 regulate TRL secretion, including controlling for fluctuations in pancreatic hormones, gastric emptying, intestinal motility, lymphatic and splanchnic blood flow, exchangeable apolipoproteins and the differences in metabolic milieu and inflammation, which arise from studying healthy participants versus diabetic patients. In hamsters, a physiological infusion of GLP-1 and GLP-2 results in an initial increase in lipid absorption and increased plasma concentrations of TRL-apoB48, which after prolonged coinfusion leads to decreased levels of TRL-apoB48 in plasma. Interestingly, under conditions of insulin resistance, hamsters displayed an increased postprandial lipid response consistent with the dominance of a GLP-2 response [69]. As GLP-1R agonists become more widely prescribed to prevent cardiovascular death in patients with T2DM and GLP-2R agonists are tested in a variety of intestinal disorders understanding both the physiological actions and pharmacological properties takes on a new importance.
Acknowledgements
I would like to thank Natasha Trzaskalski for her help with the figures.
Financial support and sponsorship
Work in the author's laboratory is funded through an NSERC Discovery Grant RGPIN-2017-05996.
Conflicts of interest
Erin Mulvihill has received honoraria from Merck in the past 12 months.
This study has not been published in its current form or a substantially similar form and is not under consideration by another publication.
REFERENCES AND RECOMMENDED READING
Papers of particular interest, published within the annual period of review, have been highlighted as:
▪ of special interest
▪▪ of outstanding interest
REFERENCES
- 1.Buse JB, Ginsberg HN, Bakris GL, et al. Primary prevention of cardiovascular diseases in people with diabetes mellitus: a scientific statement from the American Heart Association and the American Diabetes Association. Diabetes Care 2007; 30:162–172. [DOI] [PubMed] [Google Scholar]
- 2.Inzucchi SE, Bergenstal RM, Buse JB, et al. Management of hyperglycemia in type 2 diabetes: a patient-centered approach: position statement of the American Diabetes Association (ADA) and the European Association for the Study of Diabetes (EASD). Diabetes Care 2012; 35:1364–1379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Group UPDSU. Intensive blood-glucose control with sulphonylureas or insulin compared with conventional treatment and risk of complications in patients with type 2 diabetes (UKPDS 33). UK Prospective Diabetes Study (UKPDS) Group. Lancet 1998; 352:837–853. [PubMed] [Google Scholar]
- 4.Group UPDSU. Effect of intensive blood-glucose control with metformin on complications in overweight patients with type 2 diabetes (UKPDS 34). UK Prospective Diabetes Study (UKPDS) Group. Lancet 1998; 352:854–865. [PubMed] [Google Scholar]
- 5.Fonarow GC. Diabetes medications and heart failure: recognizing the risk. Circulation 2014; 130:1565–1567. [DOI] [PubMed] [Google Scholar]
- 6.Chahil TJ, Ginsberg HN. Diabetic dyslipidemia. Endocrinol Metab Clin North Am 2006; 35:491–510. vii-viii. [DOI] [PubMed] [Google Scholar]
- 7.Colhoun HM, Betteridge DJ, Durrington PN, et al. Primary prevention of cardiovascular disease with atorvastatin in type 2 diabetes in the Collaborative Atorvastatin Diabetes Study (CARDS): multicentre randomised placebo-controlled trial. Lancet 2004; 364:685–696. [DOI] [PubMed] [Google Scholar]
- 8.Bansal S, Buring JE, Rifai N, et al. Fasting compared with nonfasting triglycerides and risk of cardiovascular events in women. JAMA 2007; 298:309–316. [DOI] [PubMed] [Google Scholar]
- 9.Nordestgaard BG, Benn M, Schnohr P, Tybjaerg-Hansen A. Nonfasting triglycerides and risk of myocardial infarction, ischemic heart disease, and death in men and women. JAMA 2007; 298:299–308. [DOI] [PubMed] [Google Scholar]
- 10.Tkac I, Kimball BP, Lewis G, et al. The severity of coronary atherosclerosis in type 2 diabetes mellitus is related to the number of circulating triglyceride-rich lipoprotein particles. Arterioscler Thromb Vasc Biol 1997; 17:3633–3638. [DOI] [PubMed] [Google Scholar]
- 11.Cannon CP, Blazing MA, Giugliano RP, et al. Ezetimibe added to statin therapy after acute coronary syndromes. N Engl J Med 2015; 372:2387–2397. [DOI] [PubMed] [Google Scholar]
- 12▪.Wilke MS, Maximova K, Henderson M, et al. Adiposity in children and CVD risk: ApoB48 has a stronger association with central fat than classic lipid markers. J Clin Endocrinol Metab 2016; 101:2915–2922. [DOI] [PubMed] [Google Scholar]; In this clinical study, apoB48 is identified to associate more strongly than traditional lipid risk factors with increased adiposity in overweight and obese children.
- 13.Nauck MA, Homberger E, Siegel EG, et al. Incretin effects of increasing glucose loads in man calculated from venous insulin and C-peptide responses. J Clin Endocrinol Metab 1986; 63:492–498. [DOI] [PubMed] [Google Scholar]
- 14.Lamont BJ, Li Y, Kwan E, et al. Pancreatic GLP-1 receptor activation is sufficient for incretin control of glucose metabolism in mice. J Clin Invest 2012; 122:388–402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Fehmann HC, Habener JF. Functional receptors for the insulinotropic hormone glucagon-like peptide-I(7-37) on a somatostatin secreting cell line. FEBS Lett 1991; 279:335–340. [DOI] [PubMed] [Google Scholar]
- 16.Fehmann H-C, Goke B. Characterization of GIP(1-30) and GIP(1-42) as stimulators of proinsulin gene transcription. Peptides 1995; 16:1149–1152. [DOI] [PubMed] [Google Scholar]
- 17.Deacon CF, Johnsen AH, Holst JJ. Degradation of glucagon-like peptide-1 by human plasma in vitro yields an N-terminally truncated peptide that is a major endogenous metabolite in vivo. J Clin Endocrinol Metab 1995; 80:952–957. [DOI] [PubMed] [Google Scholar]
- 18.Baggio LL, Drucker DJ. Biology of incretins: GLP-1 and GIP. Gastroenterology 2007; 132:2131–2157. [DOI] [PubMed] [Google Scholar]
- 19.Zinman B, Wanner C, Lachin JM, et al. Empagliflozin, cardiovascular outcomes, and mortality in type 2 diabetes. N Engl J Med 2015; 373:2117–2128. [DOI] [PubMed] [Google Scholar]
- 20▪▪.Marso SP, Daniels GH, Brown-Frandsen K, et al. Liraglutide and cardiovascular outcomes in type 2 diabetes. N Engl J Med 2016; 375:311–322. [DOI] [PMC free article] [PubMed] [Google Scholar]; In this CVD safety trial treatment with the GLP1R agonist, Liraglutide is demonstrated to reduce MACE in patients with type 2 diabetes at a high risk of CVD.
- 21▪▪.Marso SP, Bain SC, Consoli A, et al. Semaglutide and cardiovascular outcomes in patients with type 2 diabetes. N Engl J Med 2016; 375:1834–1844. [DOI] [PubMed] [Google Scholar]; In this study powered to demonstrate noninferiority, a relatively short follow-up resulted in a significantly lower risk of the MACE among T2DM patients at a high risk for cardiovascular treated with semaglutide.
- 22.Nauck MA, Meier JJ, Cavender MA, et al. Cardiovascular actions and clinical outcomes with glucagon-like peptide-1 receptor agonists and dipeptidyl peptidase-4 inhibitors. Circulation 2017; 136:849–870. [DOI] [PubMed] [Google Scholar]
- 23.Drucker DJ, Yusta B. Physiology and pharmacology of the enteroendocrine hormone glucagon-like peptide-2. Annu Rev Physiol 2014; 76:561–583. [DOI] [PubMed] [Google Scholar]
- 24.Karmen A, Whyte M, Goodman DS. Fatty acid esterification and chylomicron formation during fat absorption. 1. Triglycerides and cholesterol esters. J Lipid Res 1963; 4:312–321. [PubMed] [Google Scholar]
- 25.Whyte M, Karmen A, Goodman DS. Fatty acid esterification and chylomicron formation during fat absorption. 2. Phospholipids. J Lipid Res 1963; 4:322–329. [PubMed] [Google Scholar]
- 26.Xiao C, Dash S, Morgantini C, Lewis GF. New and emerging regulators of intestinal lipoprotein secretion. Atherosclerosis 2014; 233:608–615. [DOI] [PubMed] [Google Scholar]
- 27.Pan X, Hussain MM. Gut triglyceride production. Biochim Biophys Acta 2012; 1821:727–735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Dixon JB. Mechanisms of chylomicron uptake into lacteals. Ann N Y Acad Sci 2010; 1207 (Suppl 1):E52–E57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Randolph GJ, Miller NE. Lymphatic transport of high-density lipoproteins and chylomicrons. J Clin Invest 2014; 124:929–935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Grun D, Lyubimova A, Kester L, et al. Single-cell messenger RNA sequencing reveals rare intestinal cell types. Nature 2015; 525:251–255. [DOI] [PubMed] [Google Scholar]
- 31.Dube PE, Brubaker PL. Nutrient, neural and endocrine control of glucagon-like peptide secretion. Horm Metab Res 2004; 36:755–760. [DOI] [PubMed] [Google Scholar]
- 32▪▪.Glass LL, Calero-Nieto FJ, Jawaid W, et al. Single-cell RNA-sequencing reveals a distinct population of proglucagon-expressing cells specific to the mouse upper small intestine. Mol Metab 2017; 6:1296–1303. [DOI] [PMC free article] [PubMed] [Google Scholar]; In this preclinical study, it was determined that enteroendocrine cell populations produce a greater range of peptides than was initially defined. Importantly, they also identify different receptor expression within each cell type.
- 33.Hansen L, Deacon CF, Orskov C, Holst JJ. Glucagon-like peptide-1-(7-36)amide is transformed to glucagon-like peptide-1-(9-36)amide by dipeptidyl peptidase IV in the capillaries supplying the L cells of the porcine intestine. Endocrinology 1999; 140:5356–5363. [DOI] [PubMed] [Google Scholar]
- 34.Gutierrez-Aguilar R, Woods SC. Nutrition and L and K-enteroendocrine cells. Curr Opin Endocrinol Diabetes Obes 2011; 18:35–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Orskov C, Rabenhoj L, Wettergren A, et al. Tissue and plasma concentrations of amidated and glycine-extended glucagon-like peptide I in humans. Diabetes 1994; 43:535–539. [DOI] [PubMed] [Google Scholar]
- 36.Orskov C, Wettergren A, Holst JJ. Biological effects and metabolic rates of glucagonlike peptide-1 7-36 amide and glucagonlike peptide-1 7-37 in healthy subjects are indistinguishable. Diabetes 1993; 42:658–661. [DOI] [PubMed] [Google Scholar]
- 37▪▪.Psichas A, Larraufie PF, Goldspink DA, et al. Chylomicrons stimulate incretin secretion in mouse and human cells. Diabetologia 2017; 60:2475–2485. [DOI] [PMC free article] [PubMed] [Google Scholar]; In this cell culture study, the authours provide evidence that GLP-1 secretion may occur in response to the lipolysis of chylomicrons by lipoprotein lipase.
- 38.Lu WJ, Yang Q, Yang L, et al. Chylomicron formation and secretion is required for lipid-stimulated release of incretins GLP-1 and GIP. Lipids 2012; 47:571–580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.O’Halloran DJ, Nikou GC, Kreymann B, et al. Glucagon-like peptide-1 (7-36)-NH2: a physiological inhibitor of gastric acid secretion in man. J Endocrinol 1990; 126:169–173. [DOI] [PubMed] [Google Scholar]
- 40.Schirra J, Katschinski M, Weidmann C, et al. Gastric emptying and release of incretin hormones after glucose ingestion in humans. J Clin Invest 1996; 97:92–103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Schirra J, Kuwert P, Wank U, et al. Differential effects of subcutaneous GLP-1 on gastric emptying, antroduodenal motility, and pancreatic function in men. Proc Assoc Am Physicians 1997; 109:84–97. [PubMed] [Google Scholar]
- 42.Anvari M, Paterson CA, Daniel EE, McDonald TJ. Effects of GLP-1 on gastric emptying, antropyloric motility, and transpyloric flow in response to a nonnutrient liquid. Dig Dis Sci 1998; 43:1133–1140. [DOI] [PubMed] [Google Scholar]
- 43.Qin X, Shen H, Liu M, et al. GLP-1 reduces intestinal lymph flow, triglyceride absorption, and apolipoprotein production in rats. Am J Physiol Gastrointest Liver Physiol 2005; 288:G943–G949. [DOI] [PubMed] [Google Scholar]
- 44.Schirra J, Wank U, Arnold R, et al. Effects of glucagon-like peptide-1(7-36)amide on motility and sensation of the proximal stomach in humans. Gut 2002; 50:341–348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Schirra J, Nicolaus M, Roggel R, et al. Endogenous GLP-1 controls endocrine pancreatic secretion and antro-pyloro-duodenal motility in humans. Gut 2006; 55:243–251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Giralt M, Vergara P. Glucagonlike peptide-1 (GLP-1) participation in ileal brake induced by intraluminal peptones in rat. Dig Dis Sci 1999; 44:322–329. [DOI] [PubMed] [Google Scholar]
- 47.Hsieh J, Longuet C, Baker CL, et al. The glucagon-like peptide 1 receptor is essential for postprandial lipoprotein synthesis and secretion in hamsters and mice. Diabetologia 2010; 53:552–561. [DOI] [PubMed] [Google Scholar]
- 48.Farr S, Taher J, Adeli K. Glucagon-like peptide-1 as a key regulator of lipid and lipoprotein metabolism in fasting and postprandial states. Cardiovasc Hematol Disord Drug Targets 2014; 14:126–136. [DOI] [PubMed] [Google Scholar]
- 49.Koehler JA, Baggio LL, Yusta B, et al. GLP-1R agonists promote normal and neoplastic intestinal growth through mechanisms requiring Fgf7. Cell Metab 2015; 21:379–391. [DOI] [PubMed] [Google Scholar]
- 50.Koehler JA, Kain T, Drucker DJ. Glucagon-like peptide-1 receptor activation inhibits growth and augments apoptosis in murine CT26 colon cancer cells. Endocrinology 2011; 152:3362–3372. [DOI] [PubMed] [Google Scholar]
- 51.Kedees MH, Guz Y, Grigoryan M, Teitelman G. Functional activity of murine intestinal mucosal cells is regulated by the glucagon-like peptide-1 receptor. Peptides 2013; 48:36–44. [DOI] [PubMed] [Google Scholar]
- 52.Yusta B, Baggio LL, Koehler J, et al. GLP-1R agonists modulate enteric immune responses through the intestinal intraepithelial lymphocyte GLP-1R. Diabetes 2015; 64:2537–2549. [DOI] [PubMed] [Google Scholar]
- 53▪▪.Wismann P, Barkholt P, Secher T, et al. The endogenous preproglucagon system is not essential for gut growth homeostasis in mice. Mol Metab 2017; 6:681–692. [DOI] [PMC free article] [PubMed] [Google Scholar]; This preclinical study conducted in C57BL6 mice provides a comprehensive examination of the location of GLP-1R and GLP-2R expression within the gut utilizing in-situ hybridization.
- 54.Pavlic M, Xiao C, Szeto L, et al. Insulin acutely inhibits intestinal lipoprotein secretion in humans in part by suppressing plasma free fatty acids. Diabetes 2010; 59:580–587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Alexander JS, Ganta VC, Jordan PA, Witte MH. Gastrointestinal lymphatics in health and disease. Pathophysiology 2010; 17:315–335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ohlsson L, Kohan AB, Tso P, Ahren B. GLP-1 released to the mesenteric lymph duct in mice: effects of glucose and fat. Regul Pept 2014; 189:40–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Flock G, Baggio LL, Longuet C, Drucker DJ. Incretin receptors for glucagon-like peptide 1 and glucose-dependent insulinotropic polypeptide are essential for the sustained metabolic actions of vildagliptin in mice. Diabetes 2007; 56:3006–3013. [DOI] [PubMed] [Google Scholar]
- 58.Panjwani N, Mulvihill EE, Longuet C, et al. GLP-1 receptor activation indirectly reduces hepatic lipid accumulation but does not attenuate development of atherosclerosis in diabetic male ApoE−/− mice. Endocrinology 2013; 154:127–139. [DOI] [PubMed] [Google Scholar]
- 59.Farr S, Baker C, Naples M, et al. Central nervous system regulation of intestinal lipoprotein metabolism by glucagon-like peptide-1 via a brain-gut axis. Arterioscler Thromb Vasc Biol 2015; 35:1092–1100. [DOI] [PubMed] [Google Scholar]
- 60.Tolessa T, Gutniak M, Holst JJ, et al. Glucagon-like peptide-1 retards gastric emptying and small bowel transit in the rat: effect mediated through central or enteric nervous mechanisms. Dig Dis Sci 1998; 43:2284–2290. [DOI] [PubMed] [Google Scholar]
- 61.Baggio LL, Drucker DJ. Glucagon-like peptide-1 receptors in the brain: controlling food intake and body weight. J Clin Invest 2014; 124:4223–4226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Sisley S, Gutierrez-Aguilar R, Scott M, et al. Neuronal GLP1R mediates liraglutide's anorectic but not glucose-lowering effect. J Clin Invest 2014; 124:2456–2463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63▪▪.Burmeister MA, Ayala JE, Smouse H, et al. The hypothalamic glucagon-like peptide 1 receptor is sufficient but not necessary for the regulation of energy balance and glucose homeostasis in mice. Diabetes 2017; 66:372–384. [DOI] [PMC free article] [PubMed] [Google Scholar]; In this preclinical study, the authours utilize mouse genetics to systematically eliminate Glp1r signalling in a number of neuronal populations and provide insight into the contribution of specific neuronal populations that are important to mediate GLP1R agonist effects on food intake and regulation of glucose metabolism.
- 64.Xiao C, Bandsma RH, Dash S, et al. Exenatide, a glucagon-like peptide receptor agonist, acutely inhibits intestinal lipoprotein production in healthy humans. Arterioscler Thromb Vasc Biol 2012; 32:1513–1519. [DOI] [PubMed] [Google Scholar]
- 65.Schwartz EA, Koska J, Mullin MP, et al. Exenatide suppresses postprandial elevations in lipids and lipoproteins in individuals with impaired glucose tolerance and recent onset type 2 diabetes mellitus. Atherosclerosis 2010; 212:217–222. [DOI] [PubMed] [Google Scholar]
- 66.Pollin TI, Damcott CM, Shen H, et al. A null mutation in human APOC3 confers a favorable plasma lipid profile and apparent cardioprotection. Science 2008; 322:1702–1705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67▪▪.Larsson M, Allan CM, Jung RS, et al. Apolipoprotein C-III inhibits triglyceride hydrolysis by GPIHBP1-bound LPL. J Lipid Res 2017; 58:1893–1902. [DOI] [PMC free article] [PubMed] [Google Scholar]; This preclinical study provides mechanistic insight into how apoC3-mediated inhibition of lipoprotein lipase mediated hydrolysis of triglyceride occurs.
- 68.Brubaker PL, Izzo A, Hill M, Drucker DJ. Intestinal function in mice with small bowel growth induced by glucagon-like peptide-2. Am J Physiol 1997; 272:E1050–E1058. [DOI] [PubMed] [Google Scholar]
- 69.Hein GJ, Baker C, Hsieh J, et al. GLP-1 and GLP-2 as yin and yang of intestinal lipoprotein production: evidence for predominance of GLP-2-stimulated postprandial lipemia in normal and insulin-resistant states. Diabetes 2013; 62:373–381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Shin ED, Estall JL, Izzo A, et al. Mucosal adaptation to enteral nutrients is dependent on the physiologic actions of glucagon-like peptide-2 in mice. Gastroenterology 2005; 128:1340–1353. [DOI] [PubMed] [Google Scholar]
- 71.Thulesen J, Knudsen LB, Hartmann B, et al. The truncated metabolite GLP-2 (3-33) interacts with the GLP-2 receptor as a partial agonist. Regul Pept 2002; 103:9–15. [DOI] [PubMed] [Google Scholar]
- 72▪.Baldassano S, Amato A, Rappa F, et al. Influence of endogenous glucagon-like peptide-2 on lipid disorders in mice fed a high-fat diet. Endocr Res 2016; 41:317–324. [DOI] [PubMed] [Google Scholar]; This preclinical study conducted in high fat fed mice demonstrated that 4-week treatment with the GLP-2R antagonist GLP-2 (3-33) increased hepatic lipids but did not impact intestinal lipid absorption.
- 73.Orskov C, Hartmann B, Poulsen SS, et al. GLP-2 stimulates colonic growth via KGF, released by subepithelial myofibroblasts with GLP-2 receptors. Regul Pept 2005; 124:105–112. [DOI] [PubMed] [Google Scholar]
- 74▪.Yusta B, Matthews D, Flock GB, et al. Glucagon-like peptide-2 promotes gallbladder refilling via a TGR5-independent, GLP-2R-dependent pathway. Mol Metab 2017; 6:503–511. [DOI] [PMC free article] [PubMed] [Google Scholar]; In this preclinical study, the authors provide evidence for a novel bile acid -Tgr5-GLP-2 signalling cascade to regulate gall bladder contraction and subsequent emptying in response to nutrients.
- 75.Pedersen J, Pedersen NB, Brix SW, et al. The glucagon-like peptide 2 receptor is expressed in enteric neurons and not in the epithelium of the intestine. Peptides 2015; 67:20–28. [DOI] [PubMed] [Google Scholar]
- 76.Hsieh J, Longuet C, Maida A, et al. Glucagon-like peptide-2 increases intestinal lipid absorption and chylomicron production via CD36. Gastroenterology 2009; 137:997–1005. [DOI] [PubMed] [Google Scholar]
- 77.Guan X, Stoll B, Lu X, et al. GLP-2-mediated up-regulation of intestinal blood flow and glucose uptake is nitric oxide-dependent in TPN-fed piglets. Gastroenterology 2003; 125:136–147. [DOI] [PubMed] [Google Scholar]
- 78.Pacher P, Beckman JS, Liaudet L. Nitric oxide and peroxynitrite in health and disease. Physiol Rev 2007; 87:315–424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Hsieh J, Trajcevski KE, Farr SL, et al. Glucagon-like peptide 2 (GLP-2) stimulates postprandial chylomicron production and postabsorptive release of intestinal triglyceride storage pools via induction of nitric oxide signaling in male hamsters and mice. Endocrinology 2015; 156:3538–3547. [DOI] [PubMed] [Google Scholar]
- 80.Sato SHR, Hozumi H, Ueda T, et al. Effect of different kind of dietary lipid on glucagon-like peptide2 (GLP-2) concentration in intestinal lymph of rats. Gastroenterology 2010; 138:S-406. [Google Scholar]
- 81.Meier JJ, Nauck MA, Pott A, et al. Glucagon-like peptide 2 stimulates glucagon secretion, enhances lipid absorption, and inhibits gastric acid secretion in humans. Gastroenterology 2006; 130:44–54. [DOI] [PubMed] [Google Scholar]
- 82.Dash S, Xiao C, Morgantini C, et al. Glucagon-like peptide-2 regulates release of chylomicrons from the intestine. Gastroenterology 2014; 147:1275–1284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Matikainen N, Bjornson E, Soderlund S, et al. Minor contribution of endogenous GLP-1 and GLP-2 to postprandial lipemia in obese men. PLoS One 2016; 11:e0145890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Deacon CF, Pridal L, Klarskov L, et al. Glucagon-like peptide 1 undergoes differential tissue-specific metabolism in the anesthetized pig. Am J Physiol 1996; 271:E458–E464. [DOI] [PubMed] [Google Scholar]
- 85.Drucker DJ, Shi Q, Crivici A, et al. Regulation of the biological activity of glucagon-like peptide 2 in vivo by dipeptidyl peptidase IV. Nat Biotechnol 1997; 15:673–677. [DOI] [PubMed] [Google Scholar]