

Abstract
Many human populations are undergoing an extinction of experience, with a progressive decline in interactions with nature. This is a consequence both of a loss of opportunity for, and orientation towards, such experiences. The trend is of concern in part because interactions with nature can be good for human health and wellbeing. One potential means of redressing these losses is through the intentional provision of resources to increase wildlife populations in close proximity to people, thereby increasing the potential for positive human–nature experiences, and thence the array of benefits that can result. In this paper, we review the evidence that these resource subsidies have such a cascade of effects. In some Westernized countries, the scale of provision is extraordinarily high, and doubtless leads to both positive and negative impacts for wildlife. In turn, these impacts often lead to more frequent, reliable and closer human–nature interactions, with a greater variety of species. The consequences for human wellbeing remain poorly understood, although benefits documented in the context of human–nature interactions more broadly seem likely to apply. There are also some important feedback loops that need to be better characterized if resource provisioning is to contribute effectively towards averting the extinction of experience.
This article is part of the theme issue ‘Anthropogenic resource subsidies and host–parasite dynamics in wildlife’.
Keywords: bird feeding, extinction of experience, resource provisioning, urbanization, urban wildlife, wildlife gardening
1. Introduction
It is widely accepted that many human populations are undergoing a progressive extinction of experience of nature [1–3]. Particularly in more developed countries and regions, over recent decades regular daily contact of people with nature has been in persistent decline. This is especially marked in children, with often increasingly stark differences between the nature experiences (e.g. visiting natural areas, watching wild animals, climbing trees) of the present generation when compared with those of their parents, grandparents and great-grandparents [4].
The extinction of experience is arguably an inevitable consequence of the growing urbanization of the human population, which in 2007 passed the point at which across the world one in every two people lived in a town or city [5]. Urban lifestyles tend to reduce the likelihood of inevitable daily interactions with nature, and increase the necessity of intentionality (e.g. in visiting greenspaces) to experience such interactions. The extinction of experience has also been fuelled by global and regional losses of natural habitat and biodiversity (e.g. [6,7]), and the growth of sedentary pastimes such as watching television, and engaging with the Internet and social media (e.g. [8,9]). Indeed, for increasing numbers of people ‘nature’ has become something primarily accessed through filters of parents, peers and/or the media; there is virtually no personal interaction involved.
The consequences of the extinction of experience are increasingly thought to be profound. First, there is compelling evidence that the loss of nature interactions has negative impacts on multiple dimensions of human health and wellbeing [10,11]. These include effects on physical health [12,13], mental health [14,15] and social wellbeing [16]. Second, there is evidence that reduced nature experiences can lead to people having less affinity to and interest in nature (e.g. [17,18]), placing less value on nature (e.g. [3,18]) and being less likely to participate in pro-environmental behaviours (e.g. [19,20]).
One conscious or subconscious response to the extinction of experience might be the intentional provision of resources to wildlife, as people seek to enable or to increase daily nature experiences. Indeed, such provisioning of resources is conducted on a massive scale in some regions. In this paper, we examine the links between such resource provisioning, impacts on wildlife, and the impacts on human–nature experiences and their consequences (figure 1). We focus on the purposeful provision of resources by the general public for wildlife. We do not address intentional provisioning for scientific research, conservation, management or tourism (e.g. see [21]) or the unintentional provision, such as from human food waste, that can occur, often at scale, in and around towns and cities. We also focus almost exclusively on urban areas, because these are environments in which people not only have reduced exposure to nature and so the extinction of experience is particularly prevalent, but also are where levels of wildlife provisioning tend to be greatest (e.g. [22]). These are also areas in which high densities of people and some animals live together, and so where the benefits and costs of resource provisioning are most starkly revealed. In the main, our examples are drawn from Western societies, which at present are the foci of documented resource provision activities. The extent to which this is a cultural and/or economic constraint remains to be determined.
Figure 1.
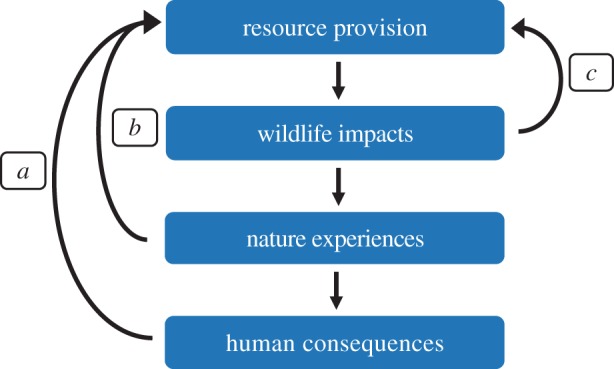
The potential links between urban resource provisioning, wildlife, and human–nature interactions and their consequences. The human consequences can lead to feedback loops in which the consequences accelerate or decelerate further resource provisioning: feedback a, health and wellbeing benefits from experiencing wildlife increase resource provision; feedback b, anticipation of wildlife experiences as yet unrealized increases resource provision; feedback c, a concern for wildlife welfare without experiencing wildlife increases resource provision. Note that the schematic diagram does not represent all potential factors and processes. (Online version in colour.)
2. Resource provision
The intentional provision of resources for wildlife, often under the banners of ‘urban greening’ and ‘wildlife gardening’, can include food, water, general habitat and breeding, roosting and wintering sites [23–25]. The most common location in which people provide such resources is within their domestic gardens [24]. These land parcels comprise a substantial proportion of the mosaic of land use in urban areas and are often in aggregate the largest component of greenspace. For example, domestic gardens constitute 16% of the area of Stockholm, Sweden [26], 36% of Dunedin, New Zealand [27], and 19–27% of cities in the UK [28]. In the UK, an estimated 87% of households have access to a private garden, varying from a few square metres to several hectares [24]. Although small in themselves, scaled-up to the national level the resources they provide are significant, such that in the UK it has been estimated that domestic gardens contain 2.5–3.5 million freshwater ponds and 28.7 million trees (just under a quarter of all trees occurring outside of woodlands; [24]). Resource provisioning includes that of both native and non-native plants, provided not only for the aesthetic appeal, but also to attract butterflies, bees (and other pollinators) and birds (e.g. [29,30]). The extent of these activities within individual gardens can vary enormously, from leaving a nettle Urtica dioica patch untouched or the installation of an artificial home for invertebrates, to full-scale management for biodiversity [31,32].
The provision of supplementary resources for birds is undoubtedly the most popular form of resource provisioning, driving a multi-billion dollar global industry [33,34]. The level and range of supplementary feeding can be astounding [35]. In the UK, there are approximately 12.6 million (48%) households providing food for birds [24], that is an average feeder density across the UK of 100 per km2 and about 200 per km2 in one city of half a million people [36]. Or, put another way, equating to one bird feeder for every nine potentially feeder-using birds in the UK [24], and providing enough resource to feed almost three times the breeding populations of 10 feeder-using songbird species [35]. Likewise, annually in the USA an estimated 54.3 million (73%) households provide 500 000 tonnes of food (figure 2a), sufficient to feed 300 million chickadees Poecile spp. if they fed on nothing else [39]. Similarly, there are a minimum of 4.7 million nest-boxes within gardens in the UK, at least one nest-box for every six breeding pairs of cavity nesting birds in the country [24]. Further, people who feed birds or put up nest-boxes are also more likely to provide other resources for birds, such as through planting trees or providing water (figure 2b). Songbirds are not the only beneficiaries, with other examples including the provision of meat for red kites (Milvus milvus) in the UK [40], and for butcherbirds and magpies in Australia [41], hand-feeding of bread to American white ibis (Eudocimus albus [42]) or fruit for cassowaries (Casuarius casuarius johnsonii) in northeast Australia [43].
Figure 2.

Empirical evidence demonstrating levels of resource provisioning around the home. (a) Feeding wildlife is the most common nature interaction in the USA [37]; (b) in New Zealand people who feed birds are more likely to engage in other resource provisioning activities [38]; (c) trends of wildlife feeding around the home in the USA [37]; and (d) birds are the most observed wildlife around the home in the USA [37]. (Online version in colour.)
Usually more opportunistically than with garden bird feeding, people also provide food resources for mammals in urban areas. In the USA, the scale of backyard feeding of racoons, squirrels, skunks, bears, coyote and deer either intentionally, or indirectly via bird feeders, is difficult to quantify, because unlike supplementary food for birds there is no equivalent measurable product for mammals. However, by surveying local residents it is possible to estimate the energetic quantity supplied [44]. There is evidence that in some urban areas provisioning may be significant, with the opportunistic feeding of mule deer, Odocoileus hemionus, and white-tailed deer, O. virginianus [21], and bears [45] by locals being cited as a factor contributing to the subsequent culling of animals [46]. In Western Europe, nocturnal urban mammals, such as hedgehogs, Erinaceus europaeus [47], and foxes, Vulpes vulpes [44], are the focus of subsidy provisioning, with 92% of 253 dead hedgehogs in Finland being found with human-fed foods in their stomachs (especially fish and milk; [48]).
3. Wildlife impacts
As resource provisioning continues to grow in popularity, it is increasingly important to understand the ecological impacts that these huge quantities of additional material have on both target and non-target wildlife populations. The provision of wildlife-friendly habitats and/or food-bearing plants has been associated with attracting a wider community of invertebrate species [30], birds (e.g. [49]) and an increase in the proportion of gardens used frequently by hedgehogs and mice (e.g. [50]). Indeed, small gardens managed for wildlife can be as rich in terms of biodiversity as large gardens, overriding the effects of size and location within the urban matrix [23,28,51].
Many of the songbirds that are the target of resource subsidies are already relatively common [52], with provisioning being associated with their increased local abundance and distribution [53,54]. Given that these species are in some cases also in major regional decline (e.g. [7]), this begs the question as to the extent to which their populations are effectively being ‘propped up’ by such provisioning. The impacts of resource subsidies can also influence the local abundances, particularly in urban areas, of species as diverse as red kites, Milvus milvus [55], hummingbirds (e.g. [56]), blackcaps, Sylvia atricapilla (e.g. [57]), and macaques [58]. It is currently unclear whether, and under what circumstances, the higher abundances of these species are associated with real increases in population abundances, or whether they are a result of displacement effects whereby resource provisioning drives local immigration [59].
The provision of easily accessible resource subsidies has been associated with positive outcomes for some target bird species, including but not limited to, increased adult overwinter survival [56,60,61], earlier lay dates and increased egg and clutch sizes [39]. Its effects on bird productivity are variable, and overwinter feeding has been found both to increase [39] and reduce [62,63] breeding success in subsequent seasons. Similarly, experiments during the breeding period have found mixed results, with evidence for both increases (e.g. [64]) and reductions [65] in productivity. Birds are not the only recipients potentially to benefit from resource provisioning, with around a third as many people in the US feeding other wildlife (figure 2c). The intentional feeding of Eastern chipmunks, Tamias striatus, allows them to maintain their activity levels from spring until autumn, without the summer lull that is seen in their rural counterparts [66]. High densities of food allow some urban mammal species to reduce their home range size compared to their rural counterparts (e.g. racoon, Procyon lotor [67]; Florida Key deer, Odocoileus virginianus clavium [68]) and has been attributed to buffering urban populations of species against severe weather events (e.g. Hanuman langurs, Semnopitheaus entellus, in India [69]).
As well as conveying benefits, supplementary feeding can also have negative consequences for target species of wildlife. Most prominent, is the increased risk of pathogen transmission through increased contact rates between hosts, and pathogens accumulating at feeders and in the surrounding environment (reviewed by [70,71]). Indeed, pathogen transmission in house finches (Haemorhous mexicanus) has been found to be significantly higher in birds in areas with high densities of bird feeders [72]. Feeder-related disease transmission is thought to have contributed towards the rapid population declines of once common species (e.g. Trichomonosis gallinae in greenfinches, Carduelis chloris; [73]). There is also evidence of an increased risk from supplementary feeding of local songbird nest predation [74], delays in the start of dawn singing [75] and changing predator–prey dynamics [76], while the provision of inappropriate foods can result in poor welfare (e.g. magpies and butcherbirds [77], cassowary [43]). It would be surprising if the provision of resource subsidies did not also impact on some other species. For example, the provision of mostly bread through garden bird feeding in New Zealand benefits introduced species (which are predominantly granivores or omnivores) at the expense of native ones (which are predominantly insectivores and nectarivores; [59]). Further, resource subsidies can result in decreases in local abundances of some non-target species, such as ground beetles that fall prey to ground-foraging birds attracted to the feeders [78], and increases in others, such as introduced grey squirrels, Sciurus carolinensis [79], and ring-necked parakeets, Psittacula karameri, in the UK [80]. Grey squirrels, for example, have been shown negatively to impact on resource acquisition by songbirds by over 90% [79], likely because of strong interference competition between songbirds and this dominant aggressive species monopolising resources [39].
Opportunistic backyard feeding of large, potentially dangerous animals has been associated with few benefits, but numerous costs for wildlife. Feeding inevitably leads to changes in behaviour and ecology of these species, including increased aggression and frequency of conflict behaviours, which can often lead to the need for the removal of problem individuals (e.g. southern cassowary in Australia [43], deer and bears in USA [21]). The hand feeding of primates appears to be common across cultures and countries, leading to behavioural changes, particularly increases in aggression and enhanced risk of road traffic fatalities (e.g. long-tailed macaques, Macaca fascicularis, in Singapore [81] and Hanuman langurs, Semnopitheaus entellis, in India [82]).
4. Nature experiences
Inevitably, such a wide range of ecological impacts resulting from resource provisioning is bound to influence people's experiences of the wildlife around them. Fundamentally, the increased density of resources in a garden, or group of gardens, will increase the flow of target (and some non-target) species into these areas (figure 3a). The resulting increased abundances are then likely to increase the frequency and duration of human–wildlife interactions. Further, the increased density of subsidies is also likely to increase the number of individuals and species that are seen at any one time, especially when a variety of resources are provided (figure 3b). Resources are usually placed in visible locations, thereby increasing the reliability of sightings, particularly of rarer species or ones with more cryptic behaviour (e.g. nocturnal ones such as hedgehogs [47]). Resource subsidies can also be associated with behavioural shifts, including increased boldness and neophilia [85], reduced flight initiation distances (figure 3c), facilitating an ability to discover new food sources more quickly [86] and allowing overwintering of some otherwise migratory species (e.g. [57]). In sum, these changes can allow people to view animals at a closer proximity, more reliably, for longer and throughout the year.
Figure 3.

Changing wildlife experiences. (a) Increased bird abundance with feeder density in Sheffield, UK [54]; (b) increased bird species richness with the number of wildlife-friendly features in gardens in Leeds, UK [25]; (c) flight initiation distances of different bird species decrease in the presence of bird feeders in urban areas [83]; and (d) the proportion of people feeding birds decreases in compact urban designs [84]. (Online version in colour.)
People tend to feed wildlife that they experience daily, with birds being the most commonly encountered (at least in terms of that wildlife which is recognized) around the home (figure 2d). In many urban areas introduced species outcompete native ones [38], therefore changing people's experiences of local bird communities. Indeed, increasing numbers of children [87] and adults [88] can now more easily recognize introduced as opposed to native species. A potential consequence of this desire to interact with everyday nature is the provision of subsidies appropriate for introduced but not native species [38], thereby exacerbating the problem. It is currently unclear what effect, if any, this shifting species baseline has on the extinction of experience or the conservation of native species (discussed in [89]).
One consequence of the extinction of experience is not only a reduced exposure to nature, but also that people may fail to note the nature that they do encounter, through a lack of familiarity and knowledge [3]. Resource provisioning means that people expect to see wildlife in, on and around these subsidies, and so are more likely to experience the wildlife that is there. However, increasing local abundances is not necessarily sufficient to maintain or increase people's daily connection to nature. As the global human population becomes increasingly urbanized, a critical question is how can we design urban areas not only to foster positive daily experiences of nature, but also people's desire to seek out these experiences [18]. Those communities living in compact urban landscape designs have reduced associated wildlife populations, with households being less likely actively or passively to engage with nearby nature [90], or to provide food for birds (figure 3d). Therefore, communities that are already deprived across multiple dimensions of health further lose access to these experiences. More sprawling urban landscape designs, with increased numbers of neighbourhood greenspaces promote population sizes of wildlife species for multiple taxa [91], facilitating more frequent daily wildlife experiences [91,92].
5. Human consequences
As resource subsidies change people's everyday experiences of wildlife, so too are they likely to influence the health and wellbeing outcomes that people receive from exposure to nature. Evidence suggests that an increase in the intensity of exposure is associated with improved health outcomes, with health gains increasing with both the quantity and quality of the natural elements that are encountered [93]. Resource subsidies can be seen positively to influence exposure quantity through an increased abundance of target and non-target species, and exposure quality through attracting an increased number of species.
Increasing exposure quantity through an increase in the abundance of resources provided has been positively associated with increases in psychological benefits, such as feelings of pleasure [33,94], and of being relaxed and connected to nature [95]. A greater number of birds means more birdsong, which contributes towards perceived attention restoration and stress reduction [96]. Cox et al. [15] found positive associations between people's mental health and the numbers of birds in their neighbourhood in the afternoon (when people are more active), but not the numbers of birds in the morning (when birds are more active; figure 4a). They concluded that mental health benefits from neighbourhood nature were likely associated with the birds that people encounter, as opposed to their intrinsic abundance. A logical next step is that a feeder, which attracts birds to where they can more easily be seen by people, has the potential to provide a focal point that might contribute to the prevention and treatment of poor mental health.
Figure 4.

Variation in health and wellbeing benefits associated with nature and nature experiences. (a) A lower population prevalence of depression was associated with higher afternoon abundances of neighbourhood birds in the UK [15]; (b) in the UK people who fed birds regularly felt more connected to nature when they watched birds in their garden [95]; and (c) people who provided resources in Australia had a higher connectedness to nature, than those with ecologically poor gardens [97,98]. (Online version in colour.)
Increasing exposure quality through an increase in the variety of resource subsidies has been positively associated with people's preferences for urban gardens [99], birds at feeders [100] and bird song [101]. However, the paradox is that most people are usually unable to appreciate richness by themselves, and so instead may gain benefits from perceived richness (discussed in [102]). A bird feeder can be seen to close the gap between actual and perceived richness, even for people with limited knowledge about the species, because it allows the viewing of multiple species within a short timeframe.
Resource subsidies provide easier access to daily interactions with wildlife around the home, which has been associated with a greater empathy for, and understanding of, the natural world [34,92,103]. Indeed, an increased connection to nature is associated with greener more diverse gardens [90], and with people being more likely to provide resources for wildlife (figure 4b). In Australia, those people with an increased orientation towards nature were more likely than those with a low nature orientation to have native plants in their garden and live in a bush setting, and be less likely to have ecologically poor gardens (figure 4c). People who fed birds regularly tended to know the names of more garden species, to consider these species to be more likeable [38,100], and to feel a greater connection to nature when they watched birds in the garden [95].
Although the opportunistic feeding of large dangerous animals doubtless provides those doing it with immediate feelings of connection to nature and wellbeing, the inevitable habituation of fed individuals can, and often does, lead to longer term negative consequences for human health and wellbeing. Feeding has been associated with an increase in the number of human–wildlife conflicts (see [21]). At the worst, attacks can lead to serious injury and death (e.g. by cassowaries [43] or dingoes [104]), while other issues include noise, mess, the destruction of property and attacks on domestic animals (e.g. [105]). The costs of conflict resulting from resource provisioning are difficult to quantify because most conflict is relatively minor, and it is difficult to assess costs such as diminished psychological wellbeing and disruption of livelihoods [106].
6. Feedbacks
To this point, we have presented a rather linear sequence of events in which resource provisioning affects wildlife, which in turn affects human nature experiences, which has consequences for the recipients of those experiences. However, the situation is doubtless more complex. There are, of course, likely to be a number of feedback loops (figure 1). Most obviously, if people gain health and wellbeing benefits from resource provisioning [107,108], and from seeing wildlife in their garden (e.g. [34,95]), then they may be more likely to persist with providing resources where these encourage more wildlife [25,44,109]; figure 1, feedback a).
Further, resource provisioning may be an expression of an orientation towards nature. Nature orientated people may be responding to a reduced opportunity to regularly experience nature, by attempting to manipulate local wildlife populations, thereby allowing closer, more meaningful interactions. Cox and Gaston [95] found that people who regularly fed birds were willing to do so even if there were none currently in the garden, although this willingness decreased in people who fed birds irregularly or did not feed birds. This suggests a feedback loop whereby those people who are orientated towards nature may provide resources because they anticipate positive human consequences (figure 1, feedback b). Conversely, a failure to use resources by wildlife, for example due to a decline in the local wildlife population [44], may decrease the desire to continue with provision by people with a low nature orientation.
Undoubtedly, many people provide resources due to motivations grounded in species conservation and welfare (e.g. [95,110]), such as ‘helping’ songbirds during periods of harsh winter weather [34]. This is despite during these periods daylight hours being shorter, with people spending less time in their gardens so arguably there being less likelihood of experiencing birds directly. Thus, perceived positive impacts for wildlife can feedback into providing resources, without the need necessarily to experience wildlife (figure 1, feedback c).
Given the multiple potential negative impacts for wildlife of resource provisioning and that a concern for wildlife is clearly a significant motivating factor for people (e.g. [110]), it is interesting that so many people provide resources across such a broad range of species. This may represent a missing feedback loop, where people do not experience the negative impacts for wildlife, and so do not associate their actions with potential welfare issues (e.g. [38]). This may be a worrying symptom of a disconnect with the natural world. Negative social feedback and peer to peer dissemination of information to change behaviours is essential to raise awareness where provisioning is inappropriate [111]. Better management and education campaigns incorporating animal welfare into a framework to evaluate feeding activities may help people to recognize the harm that feeding often causes. Encouragingly, in the USA at least, although the numbers of animals observed in urban areas is reasonably stable (figure 2d) there appears to be a steadily decreasing trend in the number of people feeding wildlife other than birds around the home (figure 2c), suggesting that there may be cultural feedback towards recognizing potential problems associated with provisioning large animals.
7. In conclusion
Resource subsidies attract an increased abundance and often richness of species in close proximity to people, thereby enabling an increased frequency, duration and intensity of daily nature experiences. In the urban landscape, increased nature exposure across these three dimensions of dose has been positively associated with the health and wellbeing of people [12,13,15,96]. Ultimately, it is unclear to what degree variation in resource provisioning is driven by a desire to connect to nature, or that people who are connected to nature are more orientated towards providing resources. However, a strong sense of connection with nature is not a prerequisite for engaging in resource provisioning, so encouraging such activities, possibly through wildlife media, has the potential to reach those who are currently unengaged [103]. As urbanization continues, understanding how these areas can be best designed to foster people's desire to connect with everyday nature, while minimizing the potential negative impacts for wildlife, is of growing importance. For example, neighbourhoods with greater greenspace connectivity allow wildlife to move between gardens (and public greenspaces) more easily, thereby promoting interactions with a greater number of people [98]. Resource provisioning has the potential to contribute towards averting the extinction of experience, for the benefit of both people and wildlife.
Acknowledgements
We are grateful to S. Gaston and two anonymous reviewers for comments.
Data accessibility
This article has no additional data.
Authors' contributions
D.T.C.C. and K.J.G. jointly conceived, designed and wrote the paper.
Competing interests
We have no competing interests to declare.
Funding
This work was funded by the Fragments, Functions, Flows and Urban Ecosystem Services project, NERC grant NE/J015237/1, with support from the Biodiversity and Ecosystem Service Sustainability (BESS) programme. BESS is a six-year programme (2011–2017) funded by the UK Natural Environment Research Council (NERC) and the Biotechnology and Biological Sciences Research Council (BBSRC) as part of the UK's Living with Environmental Change (LWEC) programme. This work presents the outcomes of independent research funded by NERC through the BESS programme. The views expressed are those of the authors and not necessarily those of the BESS Directorate or NERC.
References
- 1.Pyle RM. 1993. The thunder tree: lessons from an urban wildland. Boston, MA: Houghton Mifflin. [Google Scholar]
- 2.Miller JR. 2005. Biodiversity conservation and the extinction of experience. Trends Ecol. Evol. 20, 430–434. ( 10.1016/j.tree.2005.05.013) [DOI] [PubMed] [Google Scholar]
- 3.Soga M, Gaston KJ. 2016. Extinction of experience: evidence, consequences and challenges of loss of human-nature interactions. Front. Ecol. Environ. 14, 94–101. ( 10.1002/fee.1225) [DOI] [Google Scholar]
- 4.Louv R. 2005. Last child in the woods: saving our children from nature-deficit disorder. Chapel Hill, NC: Algonquin Books. [Google Scholar]
- 5.United Nations, Department of Economic and Social Affairs, Population Division. 2015. World Urbanization Prospects: The 2014 Revision (ST/ESA/SER.A/366). New York, NY: United Nations.
- 6.Dirzo R, Young HS, Galetti M, Ceballos G, Isaac NJB, Collen B. 2014. Defaunation in the Anthropocene. Science 345, 401–406. ( 10.1126/science.1251817) [DOI] [PubMed] [Google Scholar]
- 7.Inger R, Gregory R, Duffy JP, Stott I, Voříšek P, Gaston KJ. 2015. Common European birds are declining rapidly while less abundant species' numbers are rising. Ecol. Lett. 18, 28–36. ( 10.1111/ele.12387) [DOI] [PubMed] [Google Scholar]
- 8.Pergams ORW, Zaradic PA. 2006. Is love of nature in the US becoming love of electronic media? 16-year downtrend in national park visits explained by watching movies, playing video games, internet use, and oil prices. J. Environ. Manage. 80, 387–393. ( 10.1016/j.jenvman.2006.02.001) [DOI] [PubMed] [Google Scholar]
- 9.Ballouard JM, Brischoux F, Bonnet X. 2011. Children prioritize virtual exotic biodiversity over local biodiversity. PLoS ONE 6, e23152 ( 10.1371/journal.pone.0023152) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Keniger LE, Gaston KJ, Irvine KN, Fuller RA. 2013. What are the benefits of interacting with nature? Int. J. Environ. Res. Public Health 10, 913–935. ( 10.3390/ijerph10030913) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hartig T, Mitchell R, de Vries S, Frumkin H. 2014. Nature and health. Annu. Rev. Public Health 35, 207–228. ( 10.1146/annurev-publhealth-032013-182443) [DOI] [PubMed] [Google Scholar]
- 12.Barton J, Pretty J. 2010. What is the best dose of nature and green exercise for improving mental health? A multi-study analysis. Environ. Sci. Tech. 44, 3947–3955. ( 10.1021/es903183r) [DOI] [PubMed] [Google Scholar]
- 13.Shanahan DF, Bush R, Gaston KJ, Lin BB, Dean J, Barber E, Fuller RA. 2016. Health benefits from nature experiences depend on dose. Sci. Rep. 6, 28551 ( 10.1038/srep28551) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bratman GN, Hamilton JP, Hahn KS, Daily GC, Gross JJ. 2015. Nature experience reduces rumination and subgenual prefrontal cortex activation. Proc. Natl Acad. Sci. USA 112, 8567–8572. ( 10.1073/pnas.1510459112) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cox DTC, Shanahan DF, Hudson HL, Plummer KE, Siriwardena GM, Fuller RA, Anderson K, Hancock S. 2017. Doses of neighbourhood nature: benefits for mental health of living with nature. BioScience 67, 147–152. ( 10.1093/biosci/biw173) [DOI] [Google Scholar]
- 16.Weinstein N, Balmford A, Dehaan CR, Gladwell V, Bradbury RB, Amano T. 2015. Seeing community for the trees: the links among contact with natural environments, community cohesion, and crime. BioScience 65, 1141–1153. ( 10.1093/biosci/biv151) [DOI] [Google Scholar]
- 17.Zhang W, Goodale E, Chen J. 2014. How contact with nature affects children's biophilia, biophobia and conservation attitude in China. Biol. Conserv. 177, 109–116. ( 10.1016/j.biocon.2014.06.011) [DOI] [Google Scholar]
- 18.Soga M, Gaston KJ, Koyanagi TF, Kurisu K, Hanaki K. 2016. Urban residents' perceptions of neighbourhood nature: does the extinction of experience matter? Biol. Conserv. 203, 143–150. ( 10.1016/j.biocon.2016.09.020) [DOI] [Google Scholar]
- 19.Nord M, Luloff AE, Bridger JC. 1998. The association of forest recreation with environmentalism. Environ. Behav. 30, 235–246. ( 10.1177/0013916598302006) [DOI] [Google Scholar]
- 20.Collado S, Corraliza JA, Staats H, Ruiz M. 2015. Effect of frequency and mode of contact with nature on children's self-reported ecological behaviors. J. Environ. Psychol. 41, 65–73. ( 10.1016/j.jenvp.2014.11.001) [DOI] [Google Scholar]
- 21.Dubois S, Fraser D. 2013. A framework to evaluate wildlife feeding in research, wildlife management, tourism and recreation. Animals 3, 978–994. ( 10.3390/ani3040978) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Tryjanowski P, et al. 2015. Urban and rural habitats differ in number and type of bird feeders and in bird species consuming supplementary food. Environ. Sci. Pollut. Res. 22, 15 097–15 103. ( 10.1007/s11356-015-4723-0) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gaston KJ, Warren PH, Thompson K, Smith RM. 2005. Urban domestic gardens (IV): the extent of the resource and its associated features. Biodivers. Conserv. 14, 3327–3349. ( 10.1007/s10531-004-9513-9) [DOI] [Google Scholar]
- 24.Davies ZG, Fuller RA, Loram A, Irvine KN, Sims V, Gaston KJ. 2009. A national scale inventory of resource provision for biodiversity within domestic gardens. Biol. Conserv. 142, 761–771. ( 10.1016/j.biocon.2008.12.016) [DOI] [Google Scholar]
- 25.Goddard MA, Dougill AJ, Benton TG. 2013. Why garden for wildlife? Social and ecological drivers, motivations and barriers for biodiversity management in residential landscapes. Ecol. Econ. 86, 258–273. ( 10.1016/j.ecolecon.2012.07.016) [DOI] [Google Scholar]
- 26.Colding J, Lundberg J, Folke C. 2012. Incorporating green-area user groups in urban ecosystem management. Ambio 35, 237–244. ( 10.1579/05-A-098R.1) [DOI] [PubMed] [Google Scholar]
- 27.Mathieu R, Freeman C, Aryal J. 2007. Mapping private gardens in urban areas using object-oriented techniques and very high-resolution satellite imagery. Landsc. Urban Plan. 81, 179–192. ( 10.1016/j.landurbplan.2006.11.009) [DOI] [Google Scholar]
- 28.Loram A, Tratalos J, Warren PH, Gaston KJ. 2007. Urban domestic gardens (X): the extent and structure of the resource in five major cities. Landsc. Ecol. 22, 601–615. ( 10.1007/s10980-006-9051-9) [DOI] [Google Scholar]
- 29.Goddard MA, Dougill AJ, Benton TG. 2010. Scaling up from gardens: biodiversity conservation in urban environments. Trends Ecol. Evol. 25, 90–98. ( 10.1016/j.tree.2009.07.016) [DOI] [PubMed] [Google Scholar]
- 30.Shackleton K, Ratnieks FLW. 2016. Garden varieties: how attractive are recommended garden plants to butterflies? J. Insect Conserv. 20, 141–148. ( 10.1007/s10841-015-9827-9) [DOI] [Google Scholar]
- 31.Gaston KJ, Fuller RA, Loram A, MacDonald C, Power S, Dempsey N. 2007. Urban domestic gardens (XI): variation in urban wildlife gardening in the United Kingdom. Biodivers. Conserv. 16, 3227–3238. ( 10.1007/s10531-007-9174-6) [DOI] [Google Scholar]
- 32.Loram A, Warren PH, Thompson K, Gaston KJ. 2011. Urban domestic gardens: the effects of human interventions on garden composition. Environ. Manage. 48, 808–824. ( 10.1007/s00267-011-9723-3) [DOI] [PubMed] [Google Scholar]
- 33.Jones D. 2011. An appetite for connection: why we need to understand the effect and value of feeding wild birds. Emu 111, i–vii. ( 10.1071/MUv111n2_ED) [DOI] [Google Scholar]
- 34.Jones DN, Reynolds SJ. 2008. Feeding birds in our towns and cities: a global research opportunity. J. Avian Biol. 39, 265–271. ( 10.1111/j.2008.0908-8857.04271.x) [DOI] [Google Scholar]
- 35.Orros ME, Fellowes MDE. 2015. Wild bird feeding in an urban area: intensity, economics and numbers of individuals supported. Acta Ornithol. 50, 43–58. ( 10.3161/00016454AO2015.50.1.006) [DOI] [Google Scholar]
- 36.Fuller RA, Irvine KN, Davies ZG, Armsworth PR, Gaston KJ. 2012. Interactions between people and birds in urban landscapes. In Studies in avian biology (eds Lepczyk CA, Warren PS), pp. 249–266. Berkeley, CA: University of California Press. [Google Scholar]
- 37.U.S. Department of the Interior, U.S. Fish and Wildlife Service, and U.S. Department of Commerce. U.S. Census Bureau. 1991–2011 National survey of fishing, hunting, and wildlife-associated recreation. Available: https://www.census.gov/prod/www/fishing.html.
- 38.Galbraith JA, Beggs JR, Jones DN, McNaughton EJ, Krull CR, Stanley MC. 2014. Risks and drivers of wild bird feeding in urban areas of New Zealand. Biol. Conserv. 180, 64–74. ( 10.1016/j.biocon.2014.09.038) [DOI] [Google Scholar]
- 39.Robb GN, McDonald RA, Chamberlain DE, Bearhop S. 2008. Food for thought: supplementary feeding as a driver of ecological change in avian populations. Front. Ecol. Environ. 6, 476–484. ( 10.1890/060152) [DOI] [Google Scholar]
- 40.Orros ME, Fellowes MDE. 2014. Supplementary feeding of the reintroduced Red Kite Milvus milvus in UK gardens. Bird Study 61, 260–263. ( 10.1080/00063657.2014.885491) [DOI] [Google Scholar]
- 41.Rollinson DJ, O'Leary R, Jones DN. 2003. The practice of wildlife feeding in suburban Brisbane. Corella 27, 52–58. [Google Scholar]
- 42.Murray MH, Kidd AD, Curry SE, Hepinstall-Cymerman J, Yabsley MJ, Adams HC, Ellison T, Welch CN, Hernandez SM. 2018. From wetland specialist to hand-fed generalist: shifts in diet and condition with provisioning for a recently urbanized wading bird. Phil. Trans. R. Soc. B 373, 20170100 ( 10.1098/rstb.2017.0100) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kofron CP. 1999. Attacks to humans and domestic animals by the southern cassowary (Casuarius casuarius johnsonii) in Queensland, Australia. J. Zool. 249, 375–381. ( 10.1111/j.1469-7998.1999.tb01206.x) [DOI] [Google Scholar]
- 44.Baker P, Funk S, Harris S, Newman T, Saunders G, White P. 2004 The impact of human attitudes on the social and spatial organisation of urban foxes (Vulpes vulpes) before and after an outbreak of sarcoptic mange. In Proc. 4th Int. Symp. Urban Wildlife Conservation (eds WW Shaw, LK Harris, L VanDruff), pp. 153–163. Tucson, AZ: University of Arizona.
- 45.Dubois S, Fraser D. 2013. Local attitudes towards bear management after illegal feeding and problem bear activity. Animals 3, 935–950. ( 10.3390/ani3030935) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Hesse G.2010. British Columbia urban ungulate conflict analysis. See http://www2.gov.bc.ca/assets/gov/environment/plants-animals-and-ecosystems/wildlife-wildlife-habitat/staying-safe-around-wildlife/urbanungulatesconflictanalysisfinaljuly5-2010.pdf. (accessed on 12 June 2017).
- 47.Morris PA. 1985. The effects of supplementary feeding on movements of hedgehogs (Erinaceus europaeus). Mammal Rev. 15, 23–33. ( 10.1111/j.1365-2907.1985.tb00383.x) [DOI] [Google Scholar]
- 48.Rautio A, Isomursu M, Valtonen A, Hirvela-Koski V, Kunnasranta M. 2016. Mortality, diseases and diet of European hedgehogs (Erinaceus europaeus) in an urban environment in Finland. Mammal Res. 61, 161–169. ( 10.1007/s13364-015-0256-7) [DOI] [Google Scholar]
- 49.Belaire JA, Whelan CJ, Minor ES. 2014. Having our yards and sharing them too: the collective effects of yards on native bird species in an urban landscape. Ecol. Appl. 24, 2132–2143. ( 10.1890/13-2259.1) [DOI] [PubMed] [Google Scholar]
- 50.Baker PJ, Harris S. 2007. Urban mammals: what does the future hold? An analysis of the factors affecting patterns of use of residential gardens in Great Britain. Mammal Rev. 37, 297–315. ( 10.1111/j.1365-2907.2007.00102.x) [DOI] [Google Scholar]
- 51.Loram A, Thompson K, Warren PH, Gaston KJ. 2008. Urban domestic gardens (XII): the richness and composition of the flora in five UK cities. J. Veg. Sci. 19, 321–330. ( 10.3170/2007-8-18373) [DOI] [Google Scholar]
- 52.Cannon AR, Chamberlain DE, Toms MP, Hatchwell BJ, Gaston KJ. 2005. Trends in the use of private gardens by wild birds in Great Britain 1995–2002. J. Appl. Ecol. 42, 659–671. ( 10.1111/j.1365-2664.2005.01050.x) [DOI] [Google Scholar]
- 53.Chamberlain DE, Vickery JA, Glue DE, Robinson RA, Conway GJ, Woodburn RJW, Cannon AR. 2005. Annual and seasonal trends in the use of garden feeders by birds in winter. Ibis 147, 563–575. ( 10.1111/j.1474-919x.2005.00430.x) [DOI] [Google Scholar]
- 54.Fuller RA, Warren PH, Armsworth PR, Barbosa O, Gaston KJ. 2008. Garden bird feeding predicts the structure of urban avian assemblages. Divers. Distrib. 14, 131–137. ( 10.1111/j.1472-4642.2007.00439.x) [DOI] [Google Scholar]
- 55.Orros ME, Fellowes MDE. 2015. Widespread supplementary feeding in domestic gardens explains the return of reintroduced Red Kites Milvus milvus to an urban area. Ibis 157, 230–238. ( 10.1111/ibi.12237) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Greig EI, Wood EM, Bonter DN. 2017. Winter range expansion of a hummingbird is associated with urbanization and supplementary feeding. Proc. R. Soc. B 284, 20170256 ( 10.1098/rspb.2017.0256) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Plummer KE, Siriwardena GM, Conway GJ, Risely K, Toms MP. 2015. Is supplementary feeding in gardens a driver of evolutionary change in a migratory bird species? Glob. Change Biol. 21, 4353–4363. ( 10.1111/gcb.13070) [DOI] [PubMed] [Google Scholar]
- 58.Southwick CH, Siddiqi MF, Farooqui MY, Pal BC. 1976. Effects of artificial feeding on aggressive-behaviour of Rhesus monkeys in India. Anim. Behav. 24, 11–15. ( 10.1016/s0003-3472(76)80093-0) [DOI] [PubMed] [Google Scholar]
- 59.Galbraith JA, Beggs JR, Jones DN, Stanley MC. 2015. Supplementary feeding restructures urban bird communities. Proc. Natl Acad. Sci. USA 112, E2648–E2657. ( 10.1073/pnas.1501489112) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Jansson C, Ekman J, von Brömssen A. 1981. Winter mortality and food supply in tits Parus spp. Oikos 37, 313–322. ( 10.2307/3544122) [DOI] [Google Scholar]
- 61.Brittingham MC, Temple SA. 1988. Impacts of supplemental feeding on survival rates of Black-capped chickadees. Ecology 69, 581–589. ( 10.2307/1941007) [DOI] [Google Scholar]
- 62.Plummer KE, Bearhop S, Leech DI, Chamberlain DE, Blount JD. 2013. Fat provisioning in winter impairs egg production during the following spring: a landscape-scale study of blue tits. J. Anim. Ecol. 82, 673–682. ( 10.1111/1365-2656.12025) [DOI] [PubMed] [Google Scholar]
- 63.Plummer KE, Bearhop S, Leech DI, Chamberlain DE, Blount JD. 2013. Winter food provisioning reduces future breeding performance in a wild bird. Sci. Rep. 3, 2002 ( 10.1038/srep02002) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Peach WJ, Mallord JW, Orsman CJ, Ockendon N, Haines WG. 2013. Testing assumptions of a supplementary feeding experiment aimed at suburban House Sparrows Passer domesticus. Bird Study 60, 308–320. ( 10.1080/00063657.2013.809048) [DOI] [Google Scholar]
- 65.Harrison TJE, Smith JA, Martin GR, Chamberlain DE, Bearhop S, Robb GN, Reynolds SJ. 2010. Does food supplementation really enhance productivity of bird nestlings? Oecologia 164, 311–320. ( 10.1007/s00442-010-1645-x) [DOI] [PubMed] [Google Scholar]
- 66.Ryan DA, Larson JS. 1976. Chipmunks in residential environments. Urban Ecol. 2, 173–178. ( 10.1016/0304-4009(76)90022-X) [DOI] [Google Scholar]
- 67.Prange S, Gehrt SD, Wiggers EP. 2004. Influences of anthropogenic resources on raccoon (Procyon lotor) movements and spatial distribution. J. Mammal. 85, 483–490. ( 10.1644/bos-121) [DOI] [Google Scholar]
- 68.Harveson PM, Lopez RR, Collier BA, Silvy NJ. 2007. Impacts of urbanization on Florida Key deer behavior and population dynamics. Biol. Conserv. 134, 321–331. ( 10.1016/j.biocon.2006.07.022) [DOI] [Google Scholar]
- 69.Waite TA, Chhangani AK, Campbell LG, Rajpurohit LS, Mohnot SM. 2007. Sanctuary in the city: urban monkeys buffered against catastrophic die-off during ENSO-related drought. Ecohealth 4, 278–286. ( 10.1007/s10393-007-0112-6) [DOI] [Google Scholar]
- 70.Becker DJ, Streicker DG, Altizer S. 2015. Linking anthropogenic resources to wildlife pathogen dynamics: a review and meta-analysis. Ecol. Lett. 18, 483–495. ( 10.1111/ele.12428) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Murray MH, Becker DJ, Hall RJ, Hernandez SM. 2016. Wildlife health and supplemental feeding: a review and management recommendations. Biol. Conserv. 204, 163–174. ( 10.1016/j.biocon.2016.10.034) [DOI] [Google Scholar]
- 72.Moyers SC, Adelman JS, Farine DR, Thomason CA, Hawley DM. 2018. Feeder density enhances house finch disease transmission in experimental epidemics. Phil. Trans. R. Soc. B 373, 20170090 ( 10.1098/rstb.2017.0090) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Robinson RA, et al. 2010. Emerging infectious disease leads to rapid population declines of common British birds. PLoS ONE 5, e12215 ( 10.1371/journal.pone.0012215) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Hanmer HJ, Thomas RL, Fellowes MDE. 2017. Provision of supplementary food for wild birds may increase the risk of local nest predation. Ibis 159, 158–167. ( 10.1111/ibi.12432) [DOI] [Google Scholar]
- 75.Saggese K, Korner-Nievergelt F, Slagsvold T, Amrhein V. 2011. Wild bird feeding delays start of dawn singing in the great tit. Anim. Behav. 81, 361–365. ( 10.1016/j.anbehav.2010.11.008) [DOI] [Google Scholar]
- 76.Malpass JS, Rodewald AD, Matthews SN. 2017. Species-dependent effects of bird feeders on nest predators and nest survival of urban American Robins and Northern Cardinals. Condor 119, 1–16. ( 10.1650/condor-16-72.1) [DOI] [Google Scholar]
- 77.Stanley J, Siepen G. 1996. Please don't feed the animals. Ranger 35, 22–24. [Google Scholar]
- 78.Orros M, Thomas R, Holloway G, Fellowes ME. 2014. Supplementary feeding of wild birds indirectly affects ground beetle populations in suburban gardens. Urban Ecosyst. 18, 1–11. ( 10.1007/s11252-014-0404-x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Bonnington C, Gaston KJ, Evans KL. 2014. Assessing the potential for Grey Squirrels Sciurus carolinensis to compete with birds at supplementary feeding stations. Ibis 156, 220–226. ( 10.1111/ibi.12107) [DOI] [Google Scholar]
- 80.Peck HL, Pringle HE, Marshall HH, Owens IPF, Lord AM. 2014. Experimental evidence of impacts of an invasive parakeet on foraging behavior of native birds. Behav. Ecol. 25, 582–590. ( 10.1093/beheco/aru025) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Yeo JH, Neo H. 2010. Monkey business: human-animal conflicts in urban Singapore. Soc. Cult. Geogr. 11, 681–699. ( 10.1080/14649365.2010.508565) [DOI] [Google Scholar]
- 82.Chhangani AK. 2004. Killing of Hanuman langur (Semnopithecus entellus) in road accidents in Kumbhalgarh wildlife sanctuary, Rajasthan, India. Primate Rep. 69, 49–57. [Google Scholar]
- 83.Møller AP, Tryjanowski P, Diaz M, Kwiecinski Z, Indykiewicz P, Mitrus C, Golawski A, Polakowski M. 2016. Urban habitats and feeders both contribute to flight initiation distance reduction in birds. Behav. Ecol. 26, 861–865. ( 10.1093/beheco/arv024) [DOI] [Google Scholar]
- 84.Davies ZG, Fuller RA, Dallimer M, Loram A, Gaston KJ. 2012. Household factors influencing participation in bird feeding activity: a national scale analysis. PLoS ONE 7, e39692 ( 10.1371/journal.pone.0039692) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Tryjanowski P, et al. 2016. Urbanization affects neophilia and risk-taking at bird-feeders. Sci. Rep. 6, 7 ( 10.1038/srep28575) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Tryjanowski P, Morelli F, Skórka P, Goławski A, Indykiewicz P, Møller AP, Mitrus C, Wysocki D, Zduniak P. 2015. Who started first? Bird species visiting novel birdfeeders. Sci. Rep. 5, 11858 ( 10.1038/srep11858) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Genovart M, Tavecchia G, Ensenat JJ, Laiolo P. 2013. Holding up a mirror to the society: children recognize exotic species much more than local ones. Biol. Conserv. 159, 484–489. ( 10.1016/j.biocon.2012.10.028) [DOI] [Google Scholar]
- 88.Parker J. 2009. An analysis of urban ecological knowledge and behaviour in Wellington, New Zealand. Ph.D thesis, University of Wellington, Victoria. [Google Scholar]
- 89.Dunn PO, Gavin MC, Sanchez MC, Solomon JN. 2006. The pigeon paradox: dependence of global conservation on urban nature. Conserv. Biol. 20, 1814–1816. ( 10.1111/j.1523-1739.2006.00553.x) [DOI] [PubMed] [Google Scholar]
- 90.Lin BB, Gaston KJ, Fuller RA, Wu D, Bush R, Shanahan DF. 2017. How green is your garden?: urban form and socio-demographic factors influence yard vegetation, visitation, and ecosystem service benefits. Landsc. Urban Plan. 157, 239–246. ( 10.1016/j.landurbplan.2016.07.007) [DOI] [Google Scholar]
- 91.Sushinsky JR, Rhodes JR, Possingham HP, Gill TK, Fuller RA. 2013. How should we grow cities to minimize their biodiversity impacts? Glob. Change Biol. 19, 401–410. ( 10.1111/gcb.12055) [DOI] [PubMed] [Google Scholar]
- 92.Soga M, Yamaura Y, Aikoh T, Shoji Y, Kubo T, Gaston KJ. 2015. Reducing the extinction of experience: association between urban form and recreational use of public greenspace. Landsc. Urban Plan. 143, 69–75. ( 10.1016/j.landurbplan.2015.06.003) [DOI] [Google Scholar]
- 93.Shanahan DF, Fuller RA, Bush R, Lin BB, Gaston KJ. 2015. The health benefits of urban nature: how much do we need? BioScience 65, 476–485. ( 10.1093/biosci/biv032) [DOI] [Google Scholar]
- 94.Belaire JA, Westphal LM, Whelan J, Minor ES. 2015. Urban residents’ perceptions of birds in the neighborhood: biodiversity, cultural ecosystem services, and disservices. Condor 117, 192–202. ( 10.1650/CONDOR-14-128.1) [DOI] [Google Scholar]
- 95.Cox DTC, Gaston KJ. 2016. Urban bird feeding: connecting people with nature. PLoS ONE 11, e0158717 ( 10.1371/journal.pone.0158717) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Ratcliffe E, Gatersleben B, Sowden PT. 2013. Bird sounds and their contributions to perceived attention restoration and stress recovery. J. Environ. Psychol. 36, 221–228. ( 10.1016/j.jenvp.2013.08.004) [DOI] [Google Scholar]
- 97.Shaw A, Miller KK, Wescott G. 2017. Australian native gardens: is there scope for a community shift? Landsc. Urban Plan. 157, 322–330. ( 10.1016/j.landurbplan.2016.07.009) [DOI] [Google Scholar]
- 98.Cox DTC, Inger R, Hancock S, Anderson K, Gaston KJ. 2016. Movement of feeder-using songbirds: the influence of urban features. Sci. Rep. 6, 37660 ( 10.1038/srep37669) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Lindemann-Matthies P, Marty T. 2013. Does ecological gardening increase species richness and aesthetic quality of a garden? Biol. Conserv. 159, 37–44. ( 10.1016/j.biocon.2012.12.011) [DOI] [Google Scholar]
- 100.Cox DTC, Gaston KJ. 2015. Likeability of garden birds: importance of species knowledge and richness in connecting people to nature. PLoS ONE 10, e0141505 ( 10.1371/journal.pone.0141505) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Hedblom M, Heyman E, Antonsson H, Gunnarsson B. 2014. Bird song diversity influences young people's appreciation of urban landscapes. Urban For. Urban Green. 13, 469–474. ( 10.1016/j.ufug.2014.04.002) [DOI] [Google Scholar]
- 102.Pett TJ, Shwartz A, Irvine KN, Dallimer M, Davies ZG. 2016. Unpacking the people-biodiversity paradox: a conceptual framework. BioScience 66, 576–583. ( 10.1093/biosci/biw036) [DOI] [Google Scholar]
- 103.Shaw A, Miller K, Wescott G. 2013. Wildlife gardening and connectedness to nature: engaging the unengaged. Environ. Value. 22, 483–502. ( 10.3197/096327113X13690717320748) [DOI] [Google Scholar]
- 104.Burns GL, Howard P. 2003. When wildlife tourism goes wrong: a case study of stakeholder and management issues regarding Dingoes on Fraser Island, Australia. Tourism Manage. 24, 699–712. ( 10.1016/s0261-5177(03)00146-8) [DOI] [Google Scholar]
- 105.Soulsbury CD, White PCL. 2015. Human-wildlife interactions in urban areas: a review of conflicts, benefits and opportunities. Wildlife Res. 42, 541–553. ( 10.1071/wr14229) [DOI] [Google Scholar]
- 106.Barua M, Bhagwat SA, Jadhav S. 2013. The hidden dimensions of human-wildlife conflict: health impacts, opportunity and transaction costs. Biol. Conserv. 157, 309–316. ( 10.1016/j.biocon.2012.07.014) [DOI] [Google Scholar]
- 107.Van Den Berg AE, Custers MHG. 2011. Gardening promotes neuroendocrine and affective restoration from stress. J. Health Psychol. 16, 3–11. ( 10.1177/1359105310365577) [DOI] [PubMed] [Google Scholar]
- 108.Curtin S, Fox D. 2014. Human dimensions of wildlife gardening: its development, controversies and psychological benefits. In Horticulture: plants for people and places (eds Dixon GR, Aldous DE), pp. 1025–1046. Amsterdam, Netherlands: Springer. [Google Scholar]
- 109.Head L, Muir P. 2006. Suburban life and the boundaries of nature: resilience and rupture in Australian backyard gardens. Trans. Inst. Br. Geogr. 31, 505–524. ( 10.1111/j.1475-5661.2006.00228.x) [DOI] [Google Scholar]
- 110.Brock M, Perino G, Sugden R. 2017. The warden attitude: an investigation of the value of interaction with everyday wildlife. Environ. Resour. Econ. 67, 127–155. ( 10.1007/s10640-015-9979-9) [DOI] [Google Scholar]
- 111.McCleery RA. 2009. Improving attitudinal frameworks to predict behaviours in human wildlife conflicts. Soc. Nat. Resour. 22, 353–368. ( 10.1080/08941920802064414) [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
This article has no additional data.